Abstract
Gangliosides (GGs) are a glycolipid class present on Mesenchymal Stem Cells (MSCs) surfaces with a critical appearance role in stem cell differentiation, even though their mechanistic role in signaling and differentiation remains largely unknown. This review aims to carry out a critical analysis of the predictive role of gangliosides as specific markers of the cellular state of undifferentiated and differentiated MSCs, towards the osteogenic, chondrogenic, neurogenic, and adipogenic lineage. For this reason, we analyzed the role of GGs during multilineage differentiation processes of several types of MSCs such as Umbilical Cord-derived MSCs (UC-MSCs), Bone Marrow-derived MSCs (BM-MSCs), Dental Pulp derived MSCs (DPSCs), and Adipose derived MSCs (ADSCs). Moreover, we examined the possible role of GGs as specific cell surface markers to identify or isolate specific stem cell isotypes and their potential use as additional markers for quality control of cell-based therapies.
1. Introduction
Stem cells (SCs) can be classified based on several criteria like the differentiative capacity (totipotent, pluripotent, multipotent, etc.) or the source of derivation (embryos or adult tissues). Embryonic stem cells (ESCs) are pluripotent cells derived from the inner cell mass of the blastocyst (ICM). Once extracted, these cells can be cultured and proliferate as undifferentiated lines or they can differentiate into desired cell lines [1,2].
MSCs are part of adult stem cells (ASCs), non-specialized cells that reproduce to provide some specific cytotypes. The primary function of ASCs is to act as a reservoir of new cellular elements of a given tissue, providing for its structural maintenance and repair of any losses due to trauma or pathologies. MSCs were discovered and characterized for the first time in the bone marrow stroma and called BM-MSCs [3]. Several studies have shown the presence of these cells not only in bone marrow (BM) but also in adipose tissue (AT), umbilical cord (UC), dental pulp (DP), and other tissues [4,5]. MSCs have a degree of plasticity higher than other types of ASCs and can differentiate in vitro in different cell populations; for this reason, they are considered promising in regenerative-reparative medicine, cell therapy, and tissue engineering [6,7].
GGs are the main membrane components expressed on the external cell surface in direct contact with the extracellular environment [8]. GGs are glycosphingolipids (GSLs) complexes consisting of ceramide and a bulky sugar chain containing one or more sialic acids [9,10,11] (Figure 1A). They are also involved in the formation of lipid rafts [10,12,13]. Lipid rafts are highly dynamic structures envisaged as lateral assemblies of specific lipids and proteins in cellular membranes involved in several cellular processes [11,14] such as membrane transport, viruses’ entry, signal transduction, cell adhesion, migration, or apoptosis [15,16,17].
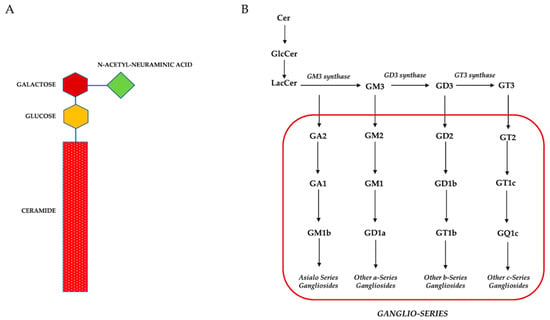
Figure 1.
(A): Schematic representation of gangliosides structure (i.e., GM3). (B): Schematic representation of gangliosides metabolism.
In recent years, several scientists showed the switch of GGs pattern during MSCs’ differentiation, suggesting a possible role of GGs in the mechanism of stem cells’ differentiation. Therefore, GGs are gaining increasing attention in the stem biology field (Table 1). Several specific ganglioside markers have been identified in SCs [18,19]. For example, SSEA-4 (a globo-series ganglioside having a NeuAcα2–3Galβ1–3GalNAcβ1-R structure) [20] is specifically expressed in human pluripotent embryonic stem cells (hESCs) [1] and in induced pluripotent stem cells (hiPSCs) [21,22].

Table 1.
Gangliosides and their role in stem cells.
Furthermore, a comparison of the glycome, including glycosphingolipids of hESCs and hiPSCs has also been reported [23]. Overall, these studies have demonstrated that carbohydrate markers can be used as important tools to confirm the pluripotency, quality, identity, and safety of pluripotent cell lines.
GD3 is expressed in mouse and human neural stem cells [24,25], and GD2 and SSEA-4 are expressed in human MSCs [25,26,27]. Moreover, it was shown that GGs play a fundamental role both in mouse embryonic stem cells (mESCs) and in DPSCs’ neuronal differentiation process [28,29].
Martinez et al., intriguingly, demonstrated the expression of GD2 only by MSCs and no other cells within the bone marrow (hematopoietic cells), suggesting that this antigen may represent a single definitive marker of marrow-derived MSCs [26].
GGs were first isolated from ganglion cells by Ernst Klenk in 1942 [30,31]. They are GSLs, composed of an amphiphilic molecule consisting of a hydrophobic ceramide tail which is usually anchored to the outer leaflet of the plasma membrane, and a glycan headgroup represented by a bulky chain sugar containing one or more sialic acids (Figure 1A) [9,10,11]. Currently, the gangliosides nomenclature proposed by Svennerholm in the 1960s remains widely used [32]. According to this nomenclature, GGs are named and classified following the number of sialic acid residues and fitting to their chromatographic mobility. GGs are abbreviated with two letters followed by a number, e.g., in GM3 the G refers to the ganglio-series of glycosphingolipids; the M stands for ‘mono’ which means 1 sialic acid, and the number 3 is based on the migration order in thin layer chromatography (TLC) (Figure 1B) [33].
GGs have important biological characteristics and functions [34], including regulation of cell proliferation, adhesion, migration, apoptosis, cell-cell interactions, cell differentiation [35,36,37], and interaction with proteins [38,39].
Moreover, GGs have been implicated in numerous pathophysiological processes, such as human malignancies [40]. One hallmark of cancer is the ability of cancer cells to undergo epithelial-to-mesenchymal transition (EMT). Through this cellular process, cancer cells lose their epithelial features to acquire mesenchymal features, becoming more aggressive and resistant to drugs [41]. Liang et al. demonstrated an association between GSLs alteration and cancer EMT [42]. During the EMT transition, they found an increase in ganglioside expression accompanied by a drastic reduction of globosides, suggesting that the last one is involved in the maintenance of epithelial characteristics [43]. Furthermore, the dynamic balance between gangliosides and globosides has been reported in mesenchymal-type pancreatic cancer cells compared to their epithelial counterparts [44], suggesting once again the involvement of GSLs during EMT.
In this review, we focused our attention on MSCs because these cells have fewer restrictions than ESCs despite showing great potential in regenerative medicine. Some authors have reported that GGs are important for modulating cell proliferation and neuronal and osteoblast differentiation of MSCs [18,45,46]. They also observed a change in ganglioside expression patterns in cells during differentiation and in response to cytokine and growth factor exposure [47,48]. Numerous studies have confirmed that GGs and their expression levels are controlled during development [49] and are cell type-specific [50], supporting the idea that these molecules are key players in cell commitment [51]. GGs could also play an important role in DPSCs’ differentiation [45,52,53], and our previous studies have shown how specifically lipid rafts are involved in DPSCs’ neuronal differentiation process [4,5,53].
For this reason, the aim of this review is to summarize the role of GGs during the multilineage differentiation processes of MSCs. In addition, since gangliosides localize on the cell surface, we would examine the possible function of GGs as specific cell surface markers to identify or isolate specific stem cell clones and their potential use as additional markers for evaluation of state and quality control of cell-based therapy products [54].
2. Main MSCs’ Properties
MSCs are undifferentiated cells relatively easy to isolate from different sources such as BM, umbilical cord blood (UCB), AT, placenta, and dental tissues, and they grow well in culture [67]. MSCs are multilineage cells able to self-renew and differentiate in multiple cell types, which play prominent roles in tissue healing and regenerative medicine. In response to some stimuli such as inflammation, trauma, and necrosis, MSCs can proliferate in vitro and can initiate the differentiative process for repairing damaged tissues in various degenerative diseases, both in animal models and in human clinical trials [68,69,70]. MSCs’ homing ability confers them the capacity to differentiate into local components of injured sites. They are also able to secrete chemokines, cytokines, and growth factors that help tissue regeneration [71,72]. The use of MSCs may reduce some of the controversial ethical issues and technical problems associated with ESCs’ use, as it is harvested from human embryos before preimplantation [73]. MSCs have a distinct morphology and express a characteristic set of surface markers, such as clusters of differentiation (CD) [74,75,76,77,78]. The biological characteristics of MSCs depend on the sources currently used for their isolation. Flow cytometry can be used to define and validate either homogeneity or heterogeneity (i.e., potential contamination) of MSCs in a tissue-specific way and to study their differentiation potential depending on the tissue origin.
3. Role of Gangliosides during Multilineage Differentiation of MSCs Isolated from Different Sources
3.1. Umbilical Cord Derived Mesenchymal Stem Cells (UC-MSCs)
The UC is a deciduous and therefore temporary, anatomical formation containing the blood vessels that connect the fetus to the placenta. For many years the UC has been treated as a waste material. In the last decade, however, it has been reconsidered and used for biomedical products and in regenerative medicine, thanks to the presence of MSCs obtained directly from the umbilical cord in toto or from its compartments (amniotic epithelium, sub-amniotic stroma, intravascular stroma named classically as Wharton’s jelly, perivascular stroma, and vessels) [79,80].
UC-MSCs are adherent to plastic and displayed fibroblastic morphology. UC-MSCs have immunomodulatory properties such as the ability to inhibit the proliferation of T, B, and NK lymphocytes and reduce inflammation by secreting interleukin-10 (IL-10) and interleukin-4 (IL-4). Also, they have anti-inflammatory effects, such as suppressing the secretion of inflammatory factor interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α), and interleukin-8 (IL-8). Next to the immunomodulatory properties, UC-MSCs have a secretory and paracrine activity important in regenerative medicine as they secrete soluble molecules such as keratinocyte growth factor (KGF), hepatocyte growth factor (HGF), epidermal growth factor (EGF), and other cytokines [81,82]. UC-MSCs have a high proliferative potential and capability to differentiate into the three germ layers: mesoderm (adipocytes, osteocytes, and chondrocytes); ectoderm (neurons, astrocytes, and glial cells), and endoderm (islet cells and liver) [83]. Cell differentiation is a highly regulated process that depends on various extracellular and intracellular factors of its modulation, including GGs. Several authors have demonstrated that GGs, such as GM3, GM1, and GD2, are already present in undifferentiated UC-MSCs [84].
Nan et al. demonstrated that GM1 induces the differentiation of UC-MSCs into neuron-like cells in vitro, characterized by the expression of the neuron-specific proteins, microtubule-associated protein 2 (MAP-2), and neurofilament protein (NFH), but not glial fibrillary acidic protein (GFAP), a marker for astrocyte development. As UC-MSCs are sub-totipotent stem cells, GM1 may provide a microenvironment to activate the specific expression programs of nerve cells and thereby induce them to differentiate into neural cells [85,86].
Jin et al. were the first to show the presence of GD2 ganglioside in the membrane and to suggest that GD2 expression is closely associated with neuronal differentiation of human UC-MSCs. GD2 can be used as a marker of neuronal differentiation as its expression is also related to the upregulation of early neurogenic transcriptional factors [84].
Xu et al. have found that UC-MSCs express GD2 in early passages, and they are the only ones in the chord that express this marker. Their data show that GD2-expressing cells are a subpopulation of primitive precursor cells and suggest that these GGs may be used as markers for isolation in the early culture steps. Furthermore, their study demonstrated that undifferentiated UC-MSCs expressed GD2, as well as differentiated cells, and GD2+ MSCs subpopulation which also showed a large upregulation of specific adipogenic, osteogenic, and neurogenic genes, suggesting the involvement of GD2 in differentiation (Table 2) [80].
Table 2.
Change in gangliosides pattern during multilineage differentiation of MSCs.
3.2. Bone Marrow-Derived Mesenchymal Stem Cells (BM-MSCs)
BM-MSCs are adult, multipotent, non-hematopoietic stem cells located in the bone marrow stroma, which can be readily harvested and isolated in humans from the sternum, vertebral body, iliac crest, and femoral shaft [93,94,95]. Today, the most common source of MSCs is BM, but bone marrow from healthy donors is a harmful and painful source. BM-MSCs are fibroblast-like cells with colony-forming and multilineage differentiation capabilities [96,97]. These are a population of cells with different commitments [98] and the multipotency capacity to differentiate into osteoblasts, chondrocytes, and adipocytes, as well as into cardiomyocytes, skeletal muscle, and neural precursors [99,100,101,102]. Among ASCs, BM-MSCs are the most studied, and due to their characteristics, such as multipotent differentiation potential, myelo-supportive capacity, anti-inflammatory and immunomodulatory properties, are used in cell therapy and tissue repair [103,104,105,106]. In response to injury signals, BM-MSCs can potentially move from their niche into the peripheral circulation and reach target tissues through vessel walls [104,107]. Gangliosides, GM3, GM2, GM1, GD3, GD2, and GD1a, are usually expressed on the cell surface of undifferentiated cells but also during the differentiation processes in which they seem to be involved [26,52]. During BM-MSCs’ osteogenic differentiation, a drastic decrease in GM3 and GD3 concentration has been demonstrated. In parallel, GM2, GM1, and GD1a increase, and GD1a becomes the ganglioside with the highest relative increase in expression during the differentiation process [29], thus demonstrating that the increase of GD1a ganglioside is crucial for BM-MSCs differentiation [52]. On the other hand, after the chondrogenic differentiation of BM-MSCs, the expression of GM3 temporarily increases. Ryu et al. reported that GM3 and GD3 are expressed after the chondrogenic differentiation and GM3 enhanced transforming growth factor-beta (TGF-β) signaling via SMAD 2/3 [37]. However, GM3 levels significantly decreased, whereas GM1, GD3, and GD1a levels increased during further differentiation into chondrocytes [87]. Furthermore, GM3 is related to the differentiation of megakaryocytes, CD4+ T cells, CD8+ T cells, osteoblasts, and neural cells (Table 2) [28,35,47,88,89].
3.3. Dental Pulp Mesenchymal Stem Cells (DPSCs)
DPSCs are multipotent stem cells [108,109] from highly vascularized connective tissue located in the center of the cavity of permanent third molars or in other dental tissues, such as the periodontal ligament, the gingival mucosa, the apical papilla, the dental follicle, or the dental pulp of childhood deciduous teeth [110] that show the properties of MSCs [111,112,113]. DPSCs were initially isolated and characterized by Gronthos et al. [114] and, in recent decades, they became the most studied type of dental stem cells, thanks to their easy extraction, absence of ethical issues, and relative abundance as biological waste from dental clinics [115]. Several researchers reviewed the isolation procedures, characterization, differentiation, and banking activity of DPSCs, providing non-invasive techniques and it has no notable ethical constraints [116,117]. DPSCs exhibit promising characteristics such as BM-MSCs, including fibroblast-like structure, clonogenicity, plasticity, rapid proliferation, regeneration, high proliferative, self-renewal, and multilineage differentiation ability [109,118]. They have the potential to differentiate into endodermal (respiratory and gastrointestinal tracts, liver, pancreas, thyroid, prostate, and bladder lineages), mesodermal (adipogenic, osteogenic, and chondrogenic lineages), and ectodermal (skin and neural lineages) [119,120] when placed in specific culture conditions. Several recent reviews have documented the current knowledge and understanding of DPSCs’ differentiation into vital lines, including their angiogenic, neurogenic, odontogenic, and chondrogenic potential and regeneration of periodontal and dental tissues [121,122,123]. Several studies have shown that GM3, GM2, and GD1a are expressed on the cell surface of undifferentiated DPSCs [36,45]. Therefore, the roles of GGs in differentiation depend on the origin of the MSCs. Lee et al. compared ganglioside expression for human adipose mesenchymal stem cells (ADSCs) and DPSCs differentiation into osteoblasts cells. They found that during DP-MSCs’ osteogenic differentiation, GM3, GM2, and GD1a were mostly increased [36,45] with a significant increase in GD1a expression compared to ADSCs [39]. Previous studies suggest that ganglioside GD1a is essential in regulating the differentiation of MSCs into osteoblasts [28,35]. Moussavou et al. suggest that GGs might play a role in the differentiation of ADSCs and DPSCs into osteoblasts and that this role is more important in regulating the osteoblast differentiation of DP-MSCs compared to ADSCs [36]. Immunostaining and high-performance thin-layer chromatography (HPTLC) analysis showed that an increase in ganglioside biosynthesis was associated with the neural differentiation of DPSCs. In fact, during DPSCs’ neuronal differentiation, GM3, GD3, and GD1a are expressed, with a significant increase in GD3 and GD1a expression [29] and furthermore, Martellucci et al., have shown that GM2 was the most representative ganglioside in DPSCs whilst GD3 was present exclusively during DPSCs’ neuronal differentiation (Table 2) [4,5].
3.4. Adipose Mesenchymal Derived Stem Cells (ADSCs)
AT is derived from the mesoderm during embryonic development, and it is present in every mammalian species, located throughout the body. AT serves as an endocrine organ, functioning to maintain energy metabolism through the storage of lipids [124]. The long-held belief about AT was that it was a relatively inert tissue in terms of biological activity, and it was believed that its main role was energy storage. However, this old theory was changed thanks to the discovery of the large abundance of adult stem/stromal cells in this tissue. ADSCs are multipotent stem cells and the actual progenitors of fat cells/adipocytes [125]. Today, AT is the predominant cell source for MSCs [126] which can be obtained more easily and efficiently than from BM [127]. The most common source of ADSCs is the white adipose tissue present in subcutaneous tissues, in the intraperitoneal compartment (visceral fat surrounding organs), and spread throughout the body as a padding for vital structures. SCs are principally isolated from adipose tissue of the abdomen, thighs, and hips/buttocks by physical methods of enzymatic digestion, typically with collagenase [128]. The resulting cellular pellet is indicated as the stromal-vascular fraction (SVF) and it is composed of all cell types present in AT, such as endothelial cells, resident, or infiltrated macrophages, pericytes, lymphocytes, and adipocyte precursors. ADSCs are selected from the other SVF cells by their adhesion on culture plates; they are a relatively homogeneous population of spindle-shaped fibroblast-like cells that expand after 7–14 days of culture [129,130]. ADSCs constitute a communication network that regulates the activity and function of adipose tissue deposits. ADSCs can interact with other cells and modulate important processes, including inflammation, apoptosis, and angiogenesis [131]. They can regulate angiogenesis through paracrine mechanisms by releasing growth factors such as vascular endothelial growth factor (VEGF), HGF, and basic fibroblast growth factor (bFGF). ADSCs have immune-modulatory activity through the secretion of IL-6 and TGF-β1 and release other important factors such as IGF-1, IL-8, and BDNF by promoting their application in tissue engineering and regenerative medicine [132]. When placed in contact with specific growth factors, ADSCs can differentiate into many different cell types of the mesodermal (bone, fat, cartilage, cardiac, and muscle cells) and non-mesodermal lineages (neuron-like cells, endothelial cells, hepatocytes, pancreatic cells) [133]. Many studies have shown that GGs play a crucial role in the differentiation of MSCs [85] and were differentially expressed during neural and osteogenic differentiation of MSCs [36,51,52,55]. GM3, GM2, and GD1a gangliosides are also expressed in undifferentiated ADSCs, and GD1a expression increased during differentiation towards an osteoblastic phenotype [36,52]. The ganglioside GD3 is known to be involved in neurogenesis and it is also considered important in the maintenance and proliferation of Neural Stem Cells (NSCs). Cho et al. demonstrated the critical role of ganglioside GD3 in the neuronal differentiation of pig adipose stem cells. Specifically, enzyme ganglioside synthase inhibition causes the blocking of ganglioside GD3 expression followed by reduced neuronal differentiation of ADSCs [90]. Regarding the adipogenic differentiation of MSCs, GM3 is the highest in adipocytes. Rampler et al. showed a significant increase in GM3, which is known to be expressed in adipocytes differentiated from adipose tissue [91,92]. In addition to these studies, Hohenwallner et al., recently found that GGs are upregulated in adipocytes compared to their human MSC progenitors, and confirmed the presence of these classes (GM3, GM2, GM1, GD3, GD2, and GD1a) in undifferentiated MSCs. They found that GM1 is abundant during adipogenic differentiation, it increases in osteogenic and decreases in chondrogenic differentiation. GM3 is expressed more in adipogenic than in osteogenic differentiated ADSCs. GD2 is not expressed in chondrogenic or osteogenic cells, but in ADSCs and differentiated adipogenic cells (Table 2) [54].
4. Gangliosides as a Potential New Class of Stem Cell Markers
The expressions of GGs are frequently and drastically changed during development and differentiation; therefore, GGs can be useful as lineage-specific differentiation markers for identifying or isolating stem cells [18,19]. Recently, attention has been placed on GGs that appear to act as molecular markers for the recognition of multipotent stem cell subclasses (Figure 2). For example, GM1, GM3, and GD2 were expressed in UC-MSCs and BM-MSCs [26,84]. This ability depends on the fact that they can be addressed by specific monoclonal antibodies, thus providing a tool for cell recognition and separation [18,134]. GGs expressed in several stem cells (pluripotent, multipotent, and cancer) have been identified by biochemical and immunological analysis [55,84,135]. Several studies have hypothesized that GGs could be used as specific markers for cell lineage or cell status [36,136]. It is currently known that GGs were differentially expressed during neural [55] and osteogenic [36,52] differentiation of MSCs. Little is known about the other lineage differentiation processes. Ganglioside GD2 may serve as a marker for the identification and purification of murine bone marrow mesenchymal stem cells (mBM-MSCs) [137]. Moreover, Xu et al., demonstrate that the cells selected by GD2 are a subpopulation of MSCs with primitive precursor cells features and provide evidence that GD2 can be considered a cell surface marker suitable for the isolation and purification of UC-MSCs in early-passage culture [80]. Meanwhile, Martinez et al., confirmed that cells selected by GD2 are a subpopulation of MSCs with primitive precursor cells features and that GD2 is highly expressed in BM-MSCs, therefore, this marker is being used for the prospective isolation of these cells [26]. Bergante et al. assessed that ganglioside GD1a could play a role in BM-MSCs differentiation, immunomagnetically sorted BM-MSCs with an anti-GD1a antibody, obtaining a cell fraction highly positive for GD1a (GD1a+) and a cell fraction poorly positive for GD1a (GD1a−). GD1a+ cell fraction revealed a significantly higher expression of osteogenic markers than GD1a− BM-MSCs, confirming their higher commitment toward the osteogenic phenotype. These results support the hypothesis that GGs could potentially be used not only as stem cell markers but also to direct stem cell differentiation [52]. Nakatani et al. examined the expression of GD3 in NSCs and evaluated the usefulness of GD3 as a cell-surface biomarker for identifying and isolating NSCs, especially from postnatal and adult brain tissue in which GD3 is rare but GM1 and GD1a are abundantly expressed. They found that in embryonic, postnatal, and adult NSCs, the percentage of GD3+ cells were more than 80% and possessed all the characteristics of NSCs, such as marker expression (nestin, Sox2, Musashi-1, and SSEA-1), a neurosphere-forming ability, and a multipotency to differentiate into neurons, astrocytes, and oligodendrocytes [24]. Wang et al. suggest that GM1 and neural growth factor (NGF) induce NSCs proliferation and differentiation, respectively, in a dose-dependent manner [138]. Rossdam et al., for the first time, describe GM3 and GD3 as being highly abundant GSLs on the cell surface of stem cell-derived cardiomyocytes [139]. Rampler et al. propose several GM3 ganglioside species as potential markers for the characterization of differentiated adipogenic cells and found that there are other classes of ganglioside including GM4 and GT3 significantly upregulated in the chondrogenic lineage compared to native MSCs [92]. Battula et al. report that ganglioside GD2 identifies a small fraction of cells in human breast cancer cell lines. Their findings suggest that GD2 is a possible novel biomarker of breast cancer stem cells (CSCs) and that GD3S, the enzyme involved in GD2 biosynthesis, is essential for breast CSCs function [140,141]. Also, in the brain, a subpopulation of brain cancer cells has been reported. These cells exhibit stem cell-like characteristics, such as the ability for self-renewal and multipotency, in addition to the capability to sustain brain tumor formation. These cells also express c-series gangliosides, also known as A2B5 antigens, characteristic of embryonic cells [142,143]. These GGs can be utilized not only as biomarkers for CSCs but also as targets for the treatment of tumors [144,145]. Considering this, we hypothesize that studies of stem cell gangliosides and their role in stem cell differentiation and other processes should prove to be a fertile area of research in the future.
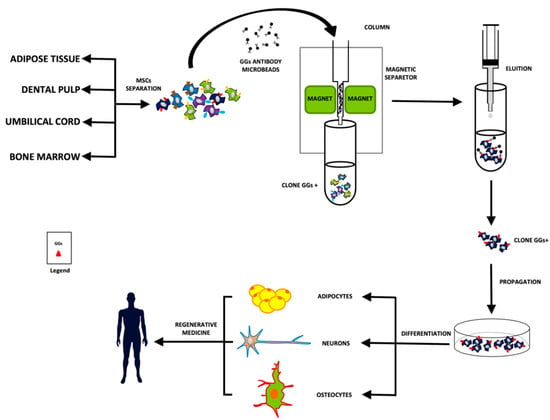
Figure 2.
Schematic procedure for the separation of the specific clones of MSCs.
5. Conclusions and Future Perspectives
In this review, we analyzed the role of GGs during MSCs’ multilineage differentiation process and the possibility of using GGs as specific markers to isolate MSC clones and maintain their cellular identity and functionalities during ex vivo culture in therapeutic applications. Several authors have shown a switch in the gangliosides pattern during the multilineage differentiation process of MSCs. Indeed, GGs’ expression frequently and drastically changed during development and differentiation, so GGs have been discussed as possible stem cell and lineage-specific differentiation markers. As for hiPSCs, their identification may contribute to the knowledge about their role in several biological processes including maintenance, proliferation, and differentiation. Hence the identification of expression of specific GGs at a different stage of MSC commitment may enable the use of these molecules as a potential target to isolate specific MSC clones (magnetic immunodetection) or as novel glycolipid-based CD markers for stem cell phenotype characterization and targeting of human stem cell clones. In conclusion, a better knowledge of gangliosides expression in stem cells will definitely allow a better separation of specific clones for their utilization in regenerative medicine. Moreover, as suggested for human embryonic and induced pluripotent stem cells [146], MSCs represent also a new tool of regenerative medicine to study MSCs-based cell therapies for disease treatments. It is noted that one of the main limitations of this work was the poor and little detailed scientific literature on this subject which made it difficult having a conclusive overview of the gangliosides pattern variation during MSCs differentiation processes.
Author Contributions
Conceptualization, V.M. and S.D.M.; writing—original draft preparation, F.S., J.F., F.P., C.S., S.D.M. and V.M.; writing—review and editing, V.M., S.D.M., F.P. and F.S.; supervision, V.M., S.D.M. and M.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomson, J.A.; Itskovitz-Eldor, J.; Shapiro, S.S.; Waknitz, M.A.; Swiergiel, J.J.; Marshall, V.S.; Jones, J.M. Embryonic stem cell lines derived from human blastocysts. Science 1998, 282, 1145–1147, Erratum in Science 1998, 282, 1827. [Google Scholar] [CrossRef] [PubMed]
- Ying, Q.L.; Stavridis, M.; Griffiths, D.; Li, M.; Smith, A. Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat. Biotechnol. 2003, 21, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar] [PubMed]
- Mattei, V.; Santacroce, C.; Tasciotti, V.; Martellucci, S.; Santilli, F.; Manganelli, V.; Piccoli, L.; Misasi, R.; Sorice, M.; Garofalo, T. Role of lipid rafts in neuronal differentiation of dental pulp-derived stem cells. Exp. Cell Res. 2015, 339, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Martellucci, S.; Manganelli, V.; Santacroce, C.; Santilli, F.; Piccoli, L.; Sorice, M.; Mattei, V. Role of prion protein-EGFR multimolecular complex during neuronal differentiation of human dental pulp-derived stem cells. Prion 2018, 12, 117–126. [Google Scholar] [CrossRef]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef]
- Rohban, R.; Pieber, T.R. Mesenchymal Stem and Progenitor Cells in Regeneration: Tissue Specificity and Regenerative Potential. Stem Cells Int. 2017, 2017, 5173732. [Google Scholar] [CrossRef]
- He, X.; Guan, F.; Lei, L. Structure, and function of glycosphingolipids on small extracellular vesicles. Glycoconj. J. 2022, 39, 197–205. [Google Scholar] [CrossRef]
- Moran, A.B.; Gardner, R.A.; Wuhrer, M.; Lageveen-Kammeijer, G.S.M.; Spencer, D.I.R. Sialic Acid Derivatization of Fluorescently Labeled N-Glycans Allows Linkage Differentiation by Reversed-Phase Liquid Chromatography-Fluorescence Detection-Mass Spectrometry. Anal. Chem. 2022, 94, 6639–6648. [Google Scholar] [CrossRef]
- Martellucci, S.; Santacroce, C.; Santilli, F.; Manganelli, V.; Sorice, M.; Mattei, V. Prion Protein in Stem Cells: A Lipid Raft Component Involved in the Cellular Differentiation Process. Int. J. Mol. Sci. 2020, 21, 4168. [Google Scholar] [CrossRef]
- Codini, M.; Garcia-Gil, M.; Albi, E. Cholesterol and Sphingolipid Enriched Lipid Rafts as Therapeutic Targets in Cancer. Int. J. Mol. Sci. 2021, 22, 726. [Google Scholar] [CrossRef] [PubMed]
- Hannun, Y.A.; Obeid, L.M. Sphingolipids and their metabolism in physiology and disease. Nat. Rev. Mol. Cell Biol. 2018, 19, 175–191, Erratum in Nat. Rev. Mol. Cell Biol. 2018, 19, 673. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Qin, Y.; Yu, X.; Xu, X.; Yu, W. Lipid raft involvement in signal transduction in cancer cell survival, cell death and metastasis. Cell Prolif. 2022, 55, e13167. [Google Scholar] [CrossRef] [PubMed]
- Mattei, V.; Manganelli, V.; Martellucci, S.; Capozzi, A.; Mantuano, E.; Longo, A.; Ferri, A.; Garofalo, T.; Sorice, M.; Misasi, R. A multimolecular signaling complex including PrPC and LRP1 is strictly dependent on lipid rafts and is essential for the function of tissue plasminogen activator. J. Neurochem. 2020, 152, 468–481. [Google Scholar] [CrossRef]
- Sorice, M.; Mattei, V.; Tasciotti, V.; Manganelli, V.; Garofalo, T.; Misasi, R. Trafficking of PrPC to mitochondrial raft-like microdomains during cell apoptosis. Prion 2012, 6, 354–358. [Google Scholar] [CrossRef]
- Sorice, M.; Mattei, V.; Matarrese, P.; Garofalo, T.; Tinari, A.; Gambardella, L.; Ciarlo, L.; Manganelli, V.; Tasciotti, V.; Misasi, R.; et al. Dynamics of mitochondrial raft-like microdomains in cell life and death. Commun. Integr. Biol. 2012, 5, 217–219. [Google Scholar] [CrossRef]
- Manganelli, V.; Capozzi, A.; Recalchi, S.; Signore, M.; Mattei, V.; Garofalo, T.; Misasi, R.; Degli Esposti, M.; Sorice, M. Altered Traffic of Cardiolipin during Apoptosis: Exposure on the Cell Surface as a Trigger for Antiphospholipid Antibodies. J. Immunol. Res. 2015, 2015, 847985. [Google Scholar] [CrossRef]
- Yanagisawa, M. Stem cell glycolipids. Neurochem. Res. 2011, 36, 1623–1635. [Google Scholar] [CrossRef]
- Yu, R.K.; Suzuki, Y.; Yanagisawa, M. Membrane glycolipids in stem cells. FEBS Lett. 2010, 584, 1694–1699. [Google Scholar] [CrossRef]
- Kannagi, R.; Cochran, N.A.; Ishigami, F.; Hakomori, S.; Andrews, P.W.; Knowles, B.B.; Solter, D. Stage-specific embryonic antigens (SSEA-3 and -4) are epitopes of a unique globo-series ganglioside isolated from human teratocarcinoma cells. EMBO J. 1983, 2, 2355–2361. [Google Scholar] [CrossRef]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef]
- Yu, J.; Vodyanik, M.A.; Smuga-Otto, K.; Antosiewicz-Bourget, J.; Frane, J.L.; Tian, S.; Nie, J.; Jonsdottir, G.A.; Ruotti, V.; Stewart, R.; et al. Induced pluripotent stem cell lines derived from human somatic cells. Science 2007, 318, 1917–1920. [Google Scholar] [CrossRef]
- Fujitani, N.; Furukawa, J.; Araki, K.; Fujioka, T.; Takegawa, Y.; Piao, J.; Nishioka, T.; Tamura, T.; Nikaido, T.; Ito, M.; et al. Total cellular glycomics allows characterizing cells and streamlining the discovery process for cellular biomarkers. Proc. Natl. Acad. Sci. USA 2013, 110, 2105–2110. [Google Scholar] [CrossRef]
- Nakatani, Y.; Yanagisawa, M.; Suzuki, Y.; Yu, R.K. Characterization of GD3 ganglioside as a novel biomarker of mouse neural stem cells. Glycobiology 2010, 20, 78–86. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Yoshimura, S.; Yu, R.K. Expression of GD2 and GD3 gangliosides in human embryonic neural stem cells. ASN Neuro 2011, 3, e00054. [Google Scholar] [CrossRef]
- Martinez, C.; Hofmann, T.J.; Marino, R.; Dominici, M.; Horwitz, E.M. Human bone marrow mesenchymal stromal cells express the neural ganglioside GD2: A novel surface marker for the identification of MSCs. Blood 2007, 109, 4245–4248, Erratum in Blood 2007, 110, 826. [Google Scholar] [CrossRef]
- Gang, E.J.; Bosnakovski, D.; Figueiredo, C.A.; Visser, J.W.; Perlingeiro, R.C. SSEA-4 identifies mesenchymal stem cells from bone marrow. Blood 2007, 109, 1743–1751. [Google Scholar] [CrossRef]
- Lee, D.H.; Koo, D.B.; Ko, K.; Ko, K.; Kim, S.M.; Jung, J.U.; Ryu, J.S.; Jin, J.W.; Yang, H.J.; Do, S.I.; et al. Effects of daunrubcin on ganglioside expression and neuronal differentiation of mouse embryonic stem cells. Biochem. Biophys. Res. Commun. 2007, 362, 313–318. [Google Scholar] [CrossRef]
- Ryu, J.S.; Ko, K.; Lee, J.W.; Park, S.B.; Byun, S.J.; Jeong, E.J.; Ko, K.; Choo, Y.K. Gangliosides are involved in neural differentiation of human dental pulp-derived stem cells. Biochem. Biophys. Res. Commun. 2009, 387, 266–271. [Google Scholar] [CrossRef]
- Klenk, E. Über die Ganglioside, eine neue Gruppe von zuckerhaltigen Gehirnlipoiden. Biol. Chem. 1942, 273, 76–86. [Google Scholar] [CrossRef]
- Xu, Y.; Sun, J.; Yang, L.; Zhao, S.; Liu, X.; Su, Y.; Zhang, J.; Zhao, M. Gangliosides play important roles in the nervous system by regulating ion concentrations. Neurochem. Res. 2022, 47, 1791–1798. [Google Scholar] [CrossRef] [PubMed]
- Svennerholm, L. The gangliosides. J. Lipid. Res. 1964, 5, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Svennerholm, L. Chromatographic separation of human brain gangliosides. J. Neurochem. 1963, 10, 613–623. [Google Scholar] [CrossRef] [PubMed]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.J.; Jung, K.Y.; Kwak, D.H.; Lee, S.H.; Ryu, J.S.; Kim, J.S.; Chang, K.T.; Lee, J.W.; Choo, Y.K. Inhibition of ganglioside GD1a synthesis suppresses the differentiation of human mesenchymal stem cells into osteoblasts. Dev. Growth Differ. 2011, 53, 323–332. [Google Scholar] [CrossRef]
- Moussavou, G.; Kwak, D.H.; Lim, M.U.; Kim, J.S.; Kim, S.U.; Chang, K.T.; Choo, Y.K. Role of gangliosides in the differentiation of human mesenchymal-derived stem cells into osteoblasts and neuronal cells. BMB Rep. 2013, 46, 527–532. [Google Scholar] [CrossRef]
- Ryu, J.S.; Seo, S.Y.; Jeong, E.J.; Kim, J.Y.; Koh, Y.G.; Kim, Y.I.; Choo, Y.K. Ganglioside GM3 Up-Regulate Chondrogenic Differentiation by Transform Growth Factor Receptors. Int. J. Mol. Sci. 2020, 21, 1967. [Google Scholar] [CrossRef]
- Bieberich, E. Sphingolipids, and lipid rafts: Novel concepts and methods of analysis. Chem. Phys. Lipids 2018, 216, 114–131. [Google Scholar] [CrossRef]
- Kim, D.H.; Triet, H.M.; Ryu, S.H. Regulation of EGFR activation and signaling by lipids on the plasma membrane. Prog. Lipid. Res. 2021, 83, 101115. [Google Scholar] [CrossRef]
- Cumin, C.; Huang, Y.L.; Everest-Dass, A.; Jacob, F. Deciphering the Importance of Glycosphingolipids on Cellular and Molecular Mechanisms Associated with Epithelial-to-Mesenchymal Transition in Cancer. Biomolecules 2021, 11, 62. [Google Scholar] [CrossRef]
- Pastushenko, I.; Blanpain, C. EMT Transition States during Tumor Progression and Metastasis. Trends Cell Biol. 2019, 29, 212–226. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.J.; Din, Y.; Levery, S.B.; Lobaton, M.; Handa, K.; Hakomori, S.I. Differential expression profiles of glycosphingolipids in human breast cancer stem cells vs. cancer non-stem cells. Proc. Natl. Acad. Sci. USA 2013, 110, 4968–4973. [Google Scholar] [CrossRef] [PubMed]
- Jacob, F.; Alam, S.; Konantz, M.; Liang, C.Y.; Kohler, R.S.; Everest-Dass, A.V.; Huang, Y.L.; Rimmer, N.; Fedier, A.; Schötzau, A.; et al. Transition of Mesenchymal and Epithelial Cancer Cells Depends on α1-4 Galactosyltransferase-Mediated Glycosphingolipids. Cancer Res. 2018, 78, 2952–2965. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; van Die, I.; Tefsen, B.; van Vliet, S.J.; Laan, L.C.; Zhang, J.; Ten Dijke, P.; Wuhrer, M.; Belo, A.I. Differential O- and Glycosphingolipid Glycosylation in Human Pancreatic Adenocarcinoma Cells with Opposite Morphology and Metastatic Behavior. Front. Oncol. 2020, 10, 732. [Google Scholar] [CrossRef]
- Lee, S.H.; Ryu, J.S.; Lee, J.W.; Kwak, D.H.; Ko, K.; Choo, Y.K. Comparison of ganglioside expression between human adi-pose and dental pulp-derived stem cell differentiation into osteoblasts. Arch. Pharm. Res. 2010, 33, 585–591. [Google Scholar] [CrossRef]
- Porter, M.J.; Zhang, G.L.; Schnaar, R.L. Ganglioside Extraction, Purification and Profiling. J. Vis. Exp. 2021, 169, e62385. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, J.U.; Ryu, J.S.; Jin, J.W.; Yang, H.J.; Ko, K.; You, H.K.; Jung, K.Y.; Choo, Y.K. Effects of gangliosides on the differentiation of human mesenchymal stem cells into osteoblasts by modulating epidermal growth factor receptors. Biochem. Biophys. Res. Commun. 2008, 371, 866–871. [Google Scholar] [CrossRef]
- Liu, Y.; Li, R.; Ladisch, S. Exogenous ganglioside GD1a enhances epidermal growth factor receptor binding and dimerization. J. Biol. Chem. 2004, 279, 36481–36489. [Google Scholar] [CrossRef]
- Yu, R.K. Development regulation of ganglioside metabolism. Prog. Brain Res. 1994, 101, 31–44. [Google Scholar] [CrossRef]
- Inokuchi, J.; Nagafuku, M.; Ohno, I.; Suzuki, A. Heterogeneity of gangliosides among T cell subsets. Cell Mol. Life Sci. 2013, 70, 3067–3075. [Google Scholar] [CrossRef]
- Bergante, S.; Creo, P.; Piccoli, M.; Ghiroldi, A.; Menon, A.; Cirillo, F.; Rota, P.; Monasky, M.M.; Ciconte, G.; Pappone, C.; et al. GM1 Ganglioside Promotes Osteogenic Differentiation of Human Tendon Stem Cells. Stem Cells Int. 2018, 2018, 4706943. [Google Scholar] [CrossRef]
- Bergante, S.; Torretta, E.; Creo, P.; Sessarego, N.; Papini, N.; Piccoli, M.; Fania, C.; Cirillo, F.; Conforti, E.; Ghiroldi, A.; et al. Gangliosides as a potential new class of stem cell markers: The case of GD1a in human bone marrow mesenchymal stem cells. J. Lipid Res. 2014, 55, 549–560. [Google Scholar] [CrossRef]
- Martellucci, S.; Santacroce, C.; Santilli, F.; Piccoli, L.; Delle Monache, S.; Angelucci, A.; Misasi, R.; Sorice, M.; Mattei, V. Cellular and Molecular Mechanisms Mediated by recPrPC Involved in the Neuronal Differentiation Process of Mesenchymal Stem Cells. Int. J. Mol. Sci. 2019, 20, 345. [Google Scholar] [CrossRef]
- Hohenwallner, K.; Troppmair, N.; Panzenboeck, L.; Kasper, C.; El Abiead, Y.; Koellensperger, G.; Lamp, L.M.; Hartler, J.; Egger, D.; Rampler, E. Decoding distinct ganglioside patterns of native and differentiated mesenchymal stem cells by a novel glycolipidomics profiling strategy. JACS Au 2022, 2, 2466–2480. [Google Scholar] [CrossRef]
- Kwak, D.H.; Yu, K.; Kim, S.M.; Lee, D.H.; Kim, S.M.; Jung, J.U.; Seo, J.W.; Kim, N.; Lee, S.; Jung, K.Y.; et al. Dynamic changes of gangliosides expression during the differentiation of embryonic and mesenchymal stem cells into neural cells. Exp. Mol. Med. 2006, 38, 668–676. [Google Scholar] [CrossRef]
- Fazzari, M.; Lunghi, G.; Chiricozzi, E.; Mauri, L.; Sonnino, S. Gangliosides, and the Treatment of Neurodegenerative Diseases: A Long Italian Tradition. Biomedicines 2022, 10, 363. [Google Scholar] [CrossRef]
- Cavallini, L.; Venerando, R.; Miotto, G.; Alexandre, A. Ganglioside GM1 protection from apoptosis of rat heart fibroblasts. Arch. Biochem. Biophys. 1999, 370, 156–162. [Google Scholar] [CrossRef]
- Tang, F.L.; Wang, J.; Itokazu, Y.; Yu, R.K. Ganglioside GD3 regulates dendritic growth in newborn neurons in adult mouse hippocampus via modulation of mitochondrial dynamics. J. Neurochem. 2021, 156, 819–833. [Google Scholar] [CrossRef]
- Wang, J.; Cheng, A.; Wakade, C.; Yu, R.K. Ganglioside GD3 is required for neurogenesis and long-term maintenance of neural stem cells in the postnatal mouse brain. J. Neurosci. 2014, 34, 13790–13800. [Google Scholar] [CrossRef]
- Todeschini, A.R.; Dos Santos, J.N.; Handa, K.; Hakomori, S.I. Ganglioside GM2/GM3 complex affixed on silica nanospheres strongly inhibits cell motility through CD82/cMet-mediated pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 1925–1930. [Google Scholar] [CrossRef]
- Schnaar, R.L.; Lopez, P.H. Myelin-associated glycoprotein and its axonal receptors. J. Neurosci. Res. 2009, 87, 3267–3276. [Google Scholar] [CrossRef] [PubMed]
- Jaiswal, R.K.; Jaiswal, N.; Bruder, S.P.; Mbalaviele, G.; Marshak, D.R.; Pittenger, M.F. Adult human mesenchymal stem cell differentiation to the osteogenic or adipogenic lineage is regulated by mitogen-activated protein kinase. J. Biol. Chem. 2000, 275, 9645–9652. [Google Scholar] [CrossRef] [PubMed]
- Itokazu, Y.; Wang, J.; Yu, R.K. Gangliosides in Nerve Cell Specification. Prog. Mol. Biol. Transl. Sci. 2018, 156, 241–263. [Google Scholar] [CrossRef] [PubMed]
- Maric, D.; Maric, I.; Chang, Y.H.; Barker, J.L. Prospective cell sorting of embryonic rat neural stem cells and neuronal and glial progenitors reveals selective effects of basic fibroblast growth factor and epidermal growth factor on self-renewal and differentiation. J. Neurosci. 2003, 23, 240–251. [Google Scholar] [CrossRef]
- Kwak, D.H.; Seo, B.B.; Chang, K.T.; Choo, Y.K. Roles of gangliosides in mouse embryogenesis and embryonic stem cell differentiation. Exp. Mol. Med. 2011, 43, 379–388. [Google Scholar] [CrossRef]
- Tsuji, S.; Arita, M.; Nagai, Y. GQ1b, a bioactive ganglioside that exhibits novel nerve growth factor (NGF)-like activities in the two neuroblastoma cell lines. J. Biochem. 1983, 94, 303–306. [Google Scholar] [CrossRef]
- Wolfe, M.; Pochampally, R.; Swaney, W.; Reger, R.L. Isolation and culture of bone marrow-derived human multipotent stromal cells (hMSCs). Methods Mol. Biol. 2008, 449, 3–25. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef]
- Oh, E.J.; Lee, H.W.; Kalimuthu, S.; Kim, T.J.; Kim, H.M.; Baek, S.H.; Zhu, L.; Oh, J.M.; Son, S.H.; Chung, H.Y.; et al. In vivo migration of mesenchymal stem cells to burn injury sites and their therapeutic effects in a living mouse model. J. Control. Release 2018, 279, 79–88. [Google Scholar] [CrossRef]
- Kim, H.K.; Lee, S.G.; Lee, S.W.; Oh, B.J.; Kim, J.H.; Kim, J.A.; Lee, G.; Jang, J.D.; Joe, Y.A. A Subset of Paracrine Factors as Efficient Biomarkers for Predicting Vascular Regenerative Efficacy of Mesenchymal Stromal/Stem Cells. Stem Cells 2019, 37, 77–88. [Google Scholar] [CrossRef]
- Gnecchi, M.; Danieli, P.; Malpasso, G.; Ciuffreda, M.C. Paracrine Mechanisms of Mesenchymal Stem Cells in Tissue Repair. Methods Mol. Biol. 2016, 1416, 123–146. [Google Scholar] [CrossRef] [PubMed]
- Lozano Navarro, L.V.; Chen, X.; Giratá Viviescas, L.T.; Ardila-Roa, A.K.; Luna-Gonzalez, M.L.; Sossa, C.L.; Arango-Rodríguez, M.L. Mesenchymal stem cells for critical limb ischemia: Their function, mechanism, and therapeutic potential. Stem Cell Res. Ther. 2022, 13, 345. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Wang, P.; Chen, W.; Weir, M.D.; Bao, C.; Xu, H.H. Human embryonic stem cells and macroporous calcium phosphate construct for bone regeneration in cranial defects in rats. Acta Biomater. 2014, 10, 4484–4493. [Google Scholar] [CrossRef] [PubMed]
- Martellucci, S.; Santacroce, C.; Manganelli, V.; Santilli, F.; Piccoli, L.; Cassetta, M.; Misasi, R.; Sorice, M.; Mattei, V. Isolation, Propagation, and Prion Protein Expression During Neuronal Differentiation of Human Dental Pulp Stem Cells. J. Vis. Exp. JoVE 2019, 145, e59282. [Google Scholar] [CrossRef]
- Mattei, V.; Martellucci, S.; Pulcini, F.; Santilli, F.; Sorice, M.; Delle Monache, S. Regenerative Potential of DPSCs and Revascularization: Direct, Paracrine or Autocrine Effect? Stem Cell Rev. Rep. 2021, 17, 1635–1646. [Google Scholar] [CrossRef]
- Delle Monache, S.; Martellucci, S.; Clementi, L.; Pulcini, F.; Santilli, F.; Mei, C.; Piccoli, L.; Angelucci, A.; Mattei, V. In Vitro Conditioning Determines the Capacity of Dental Pulp Stem Cells to Function as Pericyte-Like Cells. Stem Cells Dev. 2019, 28, 695–706. [Google Scholar] [CrossRef]
- Kobolak, J.; Dinnyes, A.; Memic, A.; Khademhosseini, A.; Mobasheri, A. Mesenchymal stem cells: Identification, phenotypic characterization, biological properties, and potential for regenerative medicine through biomaterial micro-engineering of their niche. Methods 2016, 99, 62–68. [Google Scholar] [CrossRef]
- Wang, Y.; Pan, J.; Wang, D.; Liu, J. The Use of Stem Cells in Neural Regeneration: A Review of Current Opinion. Curr. Stem Cell Res. Ther. 2018, 13, 608–617. [Google Scholar] [CrossRef]
- Can, A.; Karahuseyinoglu, S. Concise review: Human umbilical cord stroma with regard to the source of fetus-derived stem cells. Stem Cells 2007, 25, 2886–2895. [Google Scholar] [CrossRef]
- Xu, J.; Liao, W.; Gu, D.; Liang, L.; Liu, M.; Du, W.; Liu, P.; Zhang, L.; Lu, S.; Dong, C.; et al. Neural ganglioside GD2 identifies a subpopulation of mesenchymal stem cells in umbilical cord. Cell. Physiol. Biochem. 2009, 23, 415–424. [Google Scholar] [CrossRef]
- Xie, Q.; Liu, R.; Jiang, J.; Peng, J.; Yang, C.; Zhang, W.; Wang, S.; Song, J. What is the impact of human umbilical cord mesenchymal stem cell transplantation on clinical treatment? Stem Cell Res. Ther. 2020, 11, 519. [Google Scholar] [CrossRef] [PubMed]
- Filip, A.; Dabrowski, F.A.; Burdzinska, A.; Kulesza, A.; Sladowska, A.; Zolocinska, A.; Gala, K.; Paczek, L.; Wielgos, M. Comparison of the paracrine activity of mesenchymal stem cells derived from human umbilical cord, amniotic membrane, and adipose tissue. J. Obstet. Gynaecol. Res. 2017, 43, 1758–1768. [Google Scholar] [CrossRef]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.J.; Nam, H.Y.; Bae, Y.K.; Kim, S.Y.; Im, I.R.; Oh, W.; Yang, Y.S.; Choi, S.J.; Kim, S.W. GD2 expression is closely associated with neuronal differentiation of human umbilical cord blood-derived mesenchymal stem cells. Cell Mol. Life Sci. 2010, 67, 1845–1858. [Google Scholar] [CrossRef]
- Nan, C.; Shi, Y.; Zhao, Z.; Ma, S.; Liu, J.; Yan, D.; Song, G.; Liu, H. Monosialoteterahexosyl ganglioside induces the differentiation of human umbilical cord-derived mesenchymal stem cells into neuron-like cells. Int. J. Mol. Med. 2015, 36, 1057–1062. [Google Scholar] [CrossRef][Green Version]
- Brenner, M.; Messing, A. Regulation of GFAP Expression. ASN Neuro 2021, 13, 1759091420981206. [Google Scholar] [CrossRef]
- Xu, L.; Hanamatsu, H.; Homan, K.; Onodera, T.; Miyazaki, T.; Furukawa, J.I.; Hontani, K.; Tian, Y.; Baba, R.; Iwasaki, N. Alterations of Glycosphingolipid Glycans and Chondrogenic Markers during Differentiation of Human Induced Pluripotent Stem Cells into Chondrocytes. Biomolecules 2020, 10, 1622. [Google Scholar] [CrossRef]
- Jin, U.H.; Chung, T.W.; Song, K.H.; Kwak, C.H.; Choi, H.J.; Ha, K.T.; Chang, Y.C.; Lee, Y.C.; Kim, C.H. Ganglioside GM3 is required for caffeic acid phenethyl ester-induced megakaryocytic differentiation of human chronic myelogenous leukemia K562 cells. Biochem. Cell Biol. 2014, 92, 243–249. [Google Scholar] [CrossRef]
- Nagafuku, M.; Okuyama, K.; Onimaru, Y.; Suzuki, A.; Odagiri, Y.; Yamashita, T.; Iwasaki, K.; Fujiwara, M.; Takayanagi, M.; Ohno, I.; et al. CD4 and CD8 T cells require different membrane gangliosides for activation. Proc. Natl. Acad. Sci. USA 2012, 109, E336–E342. [Google Scholar] [CrossRef]
- Cho, J.H.; Ju, W.S.; Seo, S.Y.; Kim, B.H.; Kim, J.S.; Kim, J.G.; Park, S.J.; Choo, Y.K. The Potential Role of Human NME1 in Neuronal Differentiation of Porcine Mesenchymal Stem Cells: Application of NB-hNME1 as a Human NME1 Suppressor. Int. J. Mol. Sci. 2021, 22, 12194. [Google Scholar] [CrossRef]
- Nagafuku, M.; Sato, T.; Sato, S.; Shimizu, K.; Taira, T.; Inokuchi, J. Control of homeostatic and pathogenic balance in adipose tissue by ganglioside GM3. Glycobiology 2015, 25, 303–318. [Google Scholar] [CrossRef] [PubMed]
- Rampler, E.; Egger, D.; Schoeny, H.; Rusz, M.; Pacheco, M.P.; Marino, G.; Kasper, C.; Naegele, T.; Koellensperger, G. The Power of LC-MS Based Multiomics: Exploring Adipogenic Differentiation of Human Mesenchymal Stem/Stromal Cells. Molecules 2019, 24, 3615. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.D.; Casali, K.R.; Ghem, C.; da Silva, M.K.; Sausen, G.; Palma, P.B.; Covas, D.T.; Kalil, R.A.K.; Schaan, B.D.; Nardi, N.B.; et al. Mesenchymal stem cells from sternum: The type of heart disease, ischemic or valvular, does not influence the cell culture establishment and growth kinetics. J. Transl. Med. 2017, 15, 161. [Google Scholar] [CrossRef] [PubMed]
- Drela, K.; Stanaszek, L.; Snioch, K.; Kuczynska, Z.; Wrobel, M.; Sarzynska, S.; Legosz, P.; Maldyk, P.; Lukomska, B. Bone marrow-derived from the human femoral shaft as a new source of mesenchymal stem/stromal cells: An alternative cell material for banking and clinical transplantation. Stem Cell Res. Ther. 2020, 11, 262. [Google Scholar] [CrossRef]
- Fragkakis, E.M.; El-Jawhari, J.J.; Dunsmuir, R.A.; Millner, P.A.; Rao, A.S.; Henshaw, K.T.; Pountos, I.; Jones, E.; Giannoudis, P.V. Vertebral body versus iliac crest bone marrow as a source of multipotential stromal cells: Comparison of processing techniques, tri-lineage differentiation, and application on a scaffold for spine fusion. PLoS ONE 2018, 13, e0197969. [Google Scholar] [CrossRef]
- Liu, R.; Chang, W.; Wei, H.; Zhang, K. Comparison of the Biological Characteristics of Mesenchymal Stem Cells Derived from Bone Marrow and Skin. Stem Cells Int. 2016, 2016, 3658798. [Google Scholar] [CrossRef]
- Liu, C.; Tsai, A.L.; Li, P.C.; Huang, C.W.; Wu, C.C. Endothelial differentiation of bone marrow mesenchymal stem cells applicable to hypoxia and increased migration through Akt and NFκB signals. Stem Cell Res. Ther. 2017, 8, 29. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.J.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Wang, C.; Meng, H.; Wang, X.; Zhao, C.; Peng, J.; Wang, Y. Differentiation of Bone Marrow Mesenchymal Stem Cells in Osteoblasts and Adipocytes and its Role in Treatment of Osteoporosis. Med. Sci. Monit. 2016, 22, 226–233. [Google Scholar] [CrossRef]
- Sudo, K.; Kanno, M.; Miharada, K.; Ogawa, S.; Hiroyama, T.; Saijo, K.; Nakamura, Y. Mesenchymal progenitors able to differentiate into osteogenic, chondrogenic, and/or adipogenic cells in vitro are present in most primary fibroblast-like cell populations. Stem Cells 2007, 25, 1610–1617. [Google Scholar] [CrossRef]
- Moghadam, F.H.; Tayebi, T.; Dehghan, M.; Eslami, G.; Nadri, H.; Moradi, A.; Vahedian-Ardakani, H.; Barzegar, K. Differentiation of bone marrow mesenchymal stem cells into chondrocytes after short term culture in alkaline medium. Int. J. Hematol. Oncol. Stem Cell Res. 2014, 8, 12–19. [Google Scholar] [PubMed]
- Yang, H.; Xia, Y.; Lu, S.Q.; Soong, T.W.; Feng, Z.W. Basic fibroblast growth factor-induced neuronal differentiation of mouse bone marrow stromal cells requires FGFR-1, MAPK/ERK, and transcription factor AP-1. J. Biol. Chem. 2008, 283, 5287–5295. [Google Scholar] [CrossRef] [PubMed]
- Arthur, A.; Gronthos, S. Clinical Application of Bone Marrow Mesenchymal Stem/Stromal Cells to Repair Skeletal Tissue. Int. J. Mol. Sci. 2020, 21, 9759. [Google Scholar] [CrossRef]
- Fu, X.; Liu, G.; Halim, A.; Ju, Y.; Luo, Q.; Song, A.G. Mesenchymal Stem Cell Migration and Tissue Repair. Cells 2019, 8, 784. [Google Scholar] [CrossRef] [PubMed]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef]
- Purwaningrum, M.; Jamilah, N.S.; Purbantoro, S.D.; Sawangmake, C.; Nantavisai, S. Comparative characteristic study from bone marrow-derived mesenchymal stem cells. J. Vet. Sci. 2021, 22, e74. [Google Scholar] [CrossRef] [PubMed]
- Funari, A.; Alimandi, M.; Pierelli, L.; Pino, V.; Gentileschi, S.; Sacchetti, B. Human Sinusoidal Subendothelial Cells Regulate Homing, and Invasion of Circulating Metastatic Prostate Cancer Cells to Bone Marrow. Cancers 2019, 11, 763. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Lei, T.; Liu, Y.; Yang, Y.; Bi, W.; Du, H. The potential therapy with dental tissue-derived mesenchymal stem cells in Parkinson’s disease. Stem Cell Res. Ther. 2021, 12, 5. [Google Scholar] [CrossRef]
- Delle Monache, S.; Pulcini, F.; Santilli, F.; Martellucci, S.; Santacroce, C.; Fabrizi, J.; Angelucci, A.; Sorice, M.; Mattei, V. Hypoxia Induces DPSC Differentiation versus a Neurogenic Phenotype by the Paracrine Mechanism. Biomedicines 2022, 10, 1056. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Nguyen, A.L.; Yu, W.H.; Le, A.D. Human oral mucosa and gingiva: A unique reservoir for mesenchymal stem cells. J. Dent. Res. 2012, 91, 1011–1018. [Google Scholar] [CrossRef]
- Gronthos, S.; Arthur, A.; Bartold, P.M.; Shi, S. A method to isolate and culture expand human dental pulp stem cells. Methods Mol. Biol. 2011, 698, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Al Madhoun, A.; Sindhu, S.; Haddad, D.; Atari, M.; Ahmad, R.; Al-Mulla, F. Dental Pulp Stem Cells Derived from Adult Human Third Molar Tooth: A Brief Review. Front. Cell Dev. Biol. 2021, 9, 717624. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Han, X.; Kato, H.; Sato, H.; Zhang, Y.; Sun, X.; Hirofuji, Y.; Yamaza, H.; Yamada, A.; Fukumoto, S. Dental Pulp-Derived Mesenchymal Stem Cells for Modeling Genetic Disorders. Int. J. Mol. Sci. 2021, 22, 2269. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed]
- Luzuriaga, J.; Polo, Y.; Pastor-Alonso, O.; Pardo-Rodríguez, B.; Larrañaga, A.; Unda, F.; Sarasua, J.R.; Pineda, J.R.; Ibarretxe, G. Advances and Perspectives in Dental Pulp Stem Cell Based Neuroregeneration Therapies. Int. J. Mol. Sci. 2021, 22, 3546. [Google Scholar] [CrossRef] [PubMed]
- Tirino, V.; Paino, F.; De Rosa, A.; Papaccio, G. Identification, isolation, characterization, and banking of human dental pulp stem cells. Methods Mol. Biol. 2012, 879, 443–463. [Google Scholar] [CrossRef] [PubMed]
- Rodas-Junco, B.A.; Villicaña, C. Dental Pulp Stem Cells: Current Advances in Isolation, Expansion and Preservation. Tissue Eng. Regen. Med. 2017, 14, 333–347. [Google Scholar] [CrossRef]
- Aghajani, F.; Hooshmand, T.; Khanmohammadi, M.; Khanjani, S.; Edalatkhah, H.; Zarnani, A.H.; Kazemnejad, S. Comparative Immunophenotypic Characteristics, Proliferative Features, and Osteogenic Differentiation of Stem Cells Isolated from Human Permanent and Deciduous Teeth with Bone Marrow. Mol. Biotechnol. 2016, 58, 415–427. [Google Scholar] [CrossRef]
- Yamada, Y.; Nakamura-Yamada, S.; Kusano, K.; Baba, S. Clinical Potential and Current Progress of Dental Pulp Stem Cells for Various Systemic Diseases in Regenerative Medicine: A Concise Review. Int. J. Mol. Sci. 2019, 20, 1132. [Google Scholar] [CrossRef]
- Delle Monache, S.; Pulcini, F.; Frosini, R.; Mattei, V.; Talesa, V.N.; Antognelli, C. Methylglyoxal-Dependent Glycative Stress Is Prevented by the Natural Antioxidant Oleuropein in Human Dental Pulp Stem Cells through Nrf2/Glo1 Pathway. Antioxidants 2021, 10, 716. [Google Scholar] [CrossRef]
- Mortada, I.; Mortada, R.; Al Bazzal, M. Dental Pulp Stem Cells and Neurogenesis. Adv. Exp. Med. Biol. 2018, 1083, 63–75. [Google Scholar] [CrossRef] [PubMed]
- Ching, H.S.; Luddin, N.; Rahman, I.A.; Ponnuraj, K.T. Expression of Odontogenic and Osteogenic Markers in DPSCs and SHED: A Review. Curr. Stem Cell Res. Ther. 2017, 12, 71–79. [Google Scholar] [CrossRef]
- Zhai, Q.; Dong, Z.; Wang, W.; Li, B.; Jin, Y. Dental stem cell and dental tissue regeneration. Front. Med. 2019, 13, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Koenen, M.; Hill, M.A.; Cohen, P.; Sowers, J.R. Obesity, Adipose Tissue and Vascular Dysfunction. Circ. Res. 2021, 128, 951–968. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Cubedo, J. Adipose tissue depots and inflammation: Effects on plasticity and resident mesenchymal stem cell function. Cardiovasc. Res. 2017, 113, 1064–1073. [Google Scholar] [CrossRef]
- Dai, R.; Wang, Z.; Samanipour, R.; Koo, K.I.; Kim, K. Adipose-Derived Stem Cells for Tissue Engineering and Regenerative Medicine Applications. Stem Cells Int. 2016, 2016, 6737345. [Google Scholar] [CrossRef]
- Minteer, D.; Marra, K.G.; Rubin, J.P. Adipose-derived mesenchymal stem cells: Biology and potential applications. Adv. Biochem. Eng. Biotechnol. 2013, 129, 59–71. [Google Scholar] [CrossRef]
- Bunnell, B.A.; Flaat, M.; Gagliardi, C.; Patel, B.; Ripoll, C. Adipose-derived stem cells: Isolation, expansion and differentiation. Methods 2008, 45, 115–120. [Google Scholar] [CrossRef]
- Wilson, A.; Chee, M.; Butler, P.; Boyd, A.S. Isolation and Characterisation of Human Adipose-Derived Stem Cells. Methods Mol. Biol. 2019, 1899, 3–13. [Google Scholar] [CrossRef]
- Delle Monache, S.; Calgani, A.; Sanità, P.; Zazzeroni, F.; Gentile Warschauer, E.; Giuliani, A.; Amicucci, G.; Angelucci, A. Adipose-derived stem cells sustain prolonged angiogenesis through leptin secretion. Growth Factors 2016, 34, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Bacakova, L.; Zarubova, J.; Travnickova, M.; Musilkova, J.; Pajorova, J.; Slepicka, P.; Kasalkova, N.S.; Svorcik, V.; Kolska, Z.; Motarjemi, H.; et al. Stem cells: Their source, potency and use in regenerative therapies with focus on adipose-derived stem cells-a review. Biotechnol. Adv. 2018, 36, 1111–1126. [Google Scholar] [CrossRef] [PubMed]
- Zuk, P.A.; Zhu, M.; Mizuno, H.; Huang, J.; Futrell, J.W.; Katz, A.J.; Benhaim, P.; Lorenz, H.P.; Hedrick, M.H. Multilineage cells from human adipose tissue: Implications for cell-based therapies. Tissue Eng. 2001, 7, 211–228. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, A.; Futerman, A.H. Determination of the localization of gangliosides using anti-ganglioside antibodies: Comparison of fixation methods. J. Histochem. Cytochem. 1997, 45, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Kannagi, R.; Cai, B.H.; Huang, H.C.; Chao, C.C.; Sakuma, K. Gangliosides and Tumors. Methods Mol. Biol. 2018, 1804, 143–171. [Google Scholar] [CrossRef]
- Ngamukote, S.; Yanagisawa, M.; Ariga, T.; Ando, S.; Yu, R.K. Developmental changes of glycosphingolipids and expression of glycogenes in mouse brains. J. Neurochem. 2007, 103, 2327–2341. [Google Scholar] [CrossRef]
- Xu, J.; Fan, W.; Tu, X.X.; Zhang, T.; Hou, Z.J.; Guo, T.; Shu, X.; Luo, X.; Liu, Y.; Peng, F.; et al. Neural ganglioside GD2(+) cells define a subpopulation of mesenchymal stem cells in adult murine bone marrow. Cell. Physiol. Biochem. 2013, 32, 889–898. [Google Scholar] [CrossRef]
- Wang, Q.; Song, Y.H.; Tang, Z.; Wang, Z.P.; Xu, Q.; Bao, N. Effects of ganglioside GM1 and neural growth factor on neural stem cell proliferation and differentiation. Genet. Mol. Res. 2016, 15, 1–10. [Google Scholar] [CrossRef]
- Rossdam, C.; Konze, S.A.; Oberbeck, A.; Rapp, E.; Gerardy-Schahn, R.; von Itzstein, M.; Buettner, F.F.R. Approach for Profiling of Glycosphingolipid Glycosylation by Multiplexed Capillary Gel Electrophoresis Coupled to Laser-Induced Fluorescence Detection to Identify Cell-Surface Markers of Human Pluripotent Stem Cells and Derived Cardiomyocytes. Anal. Chem. 2019, 91, 6413–6418. [Google Scholar] [CrossRef]
- Battula, V.L.; Evans, K.W.; Hollier, B.G.; Shi, Y.; Marini, F.C.; Ayyanan, A.; Wang, R.Y.; Brisken, C.; Guerra, R.; Andreeff, M.; et al. Epithelial-mesenchymal transition-derived cells exhibit multilineage differentiation potential similar to mesenchymal stem cells. Stem Cells 2010, 28, 1435–1445. [Google Scholar] [CrossRef]
- Battula, V.L.; Shi, Y.; Evans, K.W.; Wang, R.Y.; Spaeth, E.L.; Jacamo, R.O.; Guerra, R.; Sahin, A.A.; Marini, F.C.; Hortobagyi, G.; et al. Ganglioside GD2 identifies breast cancer stem cells and promotes tumorigenesis. J. Clin. Investig. 2012, 122, 2066–2078. [Google Scholar] [CrossRef] [PubMed]
- Ogden, A.T.; Waziri, A.E.; Lochhead, R.A.; Fusco, D.; Lopez, K.; Ellis, J.A.; Kang, J.; Assanah, M.; McKhann, G.M.; Sisti, M.B.; et al. Identification of A2B5+CD133- tumor- initiating cells in adult human gliomas. Neurosurgery 2008, 62, 505–514. [Google Scholar] [CrossRef] [PubMed]
- Tchoghandjian, A.; Baeza, N.; Colin, C.; Cayre, M.; Metellus, P.; Beclin, C.; Ouafik, L.; Figarella-Branger, D. A2B5 cells from human glioblastoma have cancer stem cell properties. Brain Pathol. 2010, 20, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.K.; Tsai, Y.T.; Ariga, T.; Yanagisawa, M. Structures, biosynthesis, and functions of gangliosides an overview. J. Oleo Sci. 2011, 60, 537–544. [Google Scholar] [CrossRef]
- Shao, C.; Anand, V.; Andreeff, M.; Battula, V.L. Ganglioside GD2: A novel therapeutic target in triple-negative breast cancer. Ann. N. Y. Acad. Sci. 2022, 1508, 35–53. [Google Scholar] [CrossRef]
- Priester, C.; MacDonald, A.; Dhar, M.; Bow, A. Examining the Characteristics and Applications of Mesenchymal, Induced Pluripotent, and Embryonic Stem Cells for Tissue Engineering Approaches across the Germ Layers. Pharmaceuticals 2020, 13, 344. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).