Mineralization Content Alters Osteogenic Responses of Bone Marrow Stromal Cells on Hydroxyapatite/Polycaprolactone Composite Nanofiber Scaffolds



Abstract
:1. Introduction
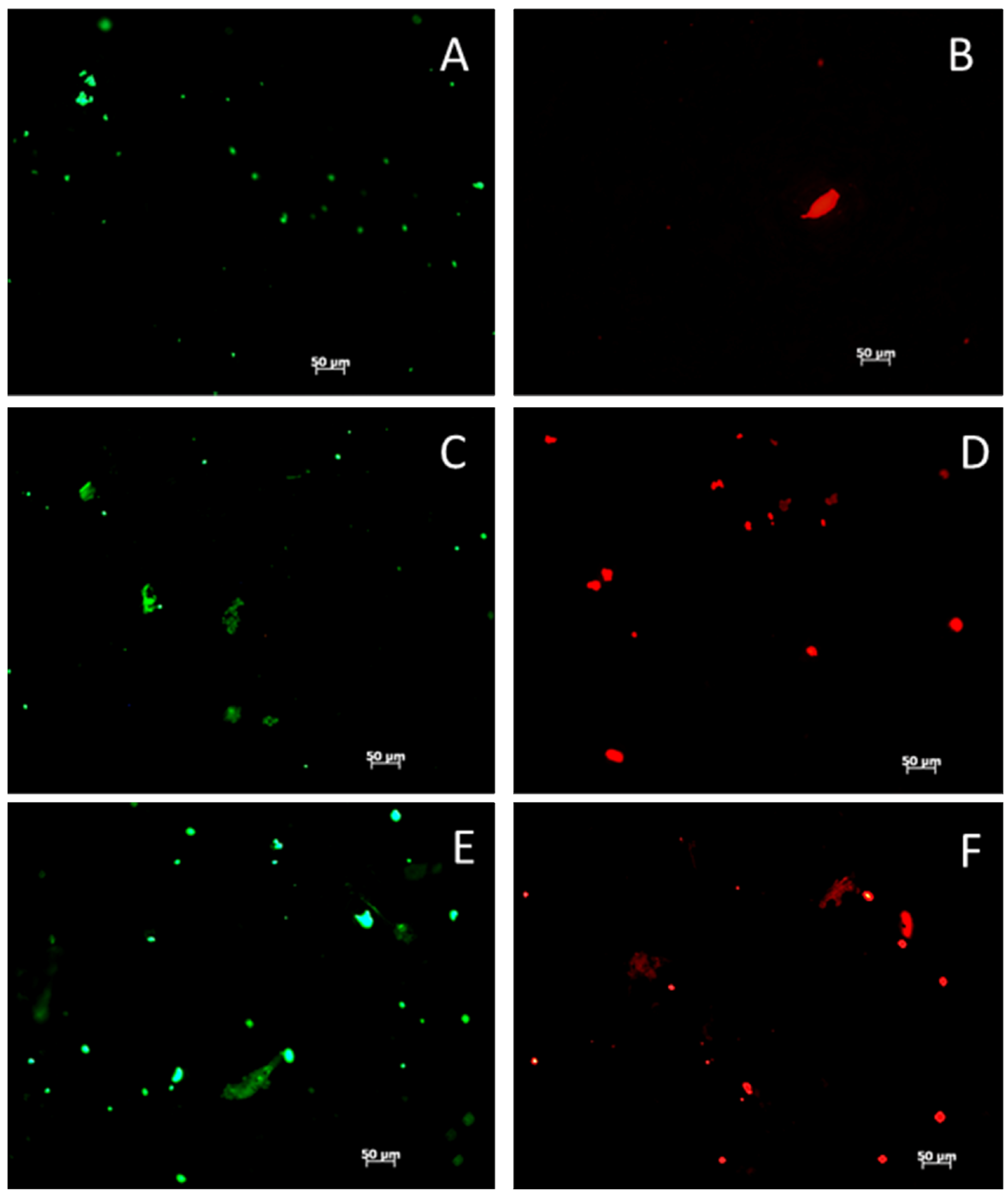
2. Results and Discussion
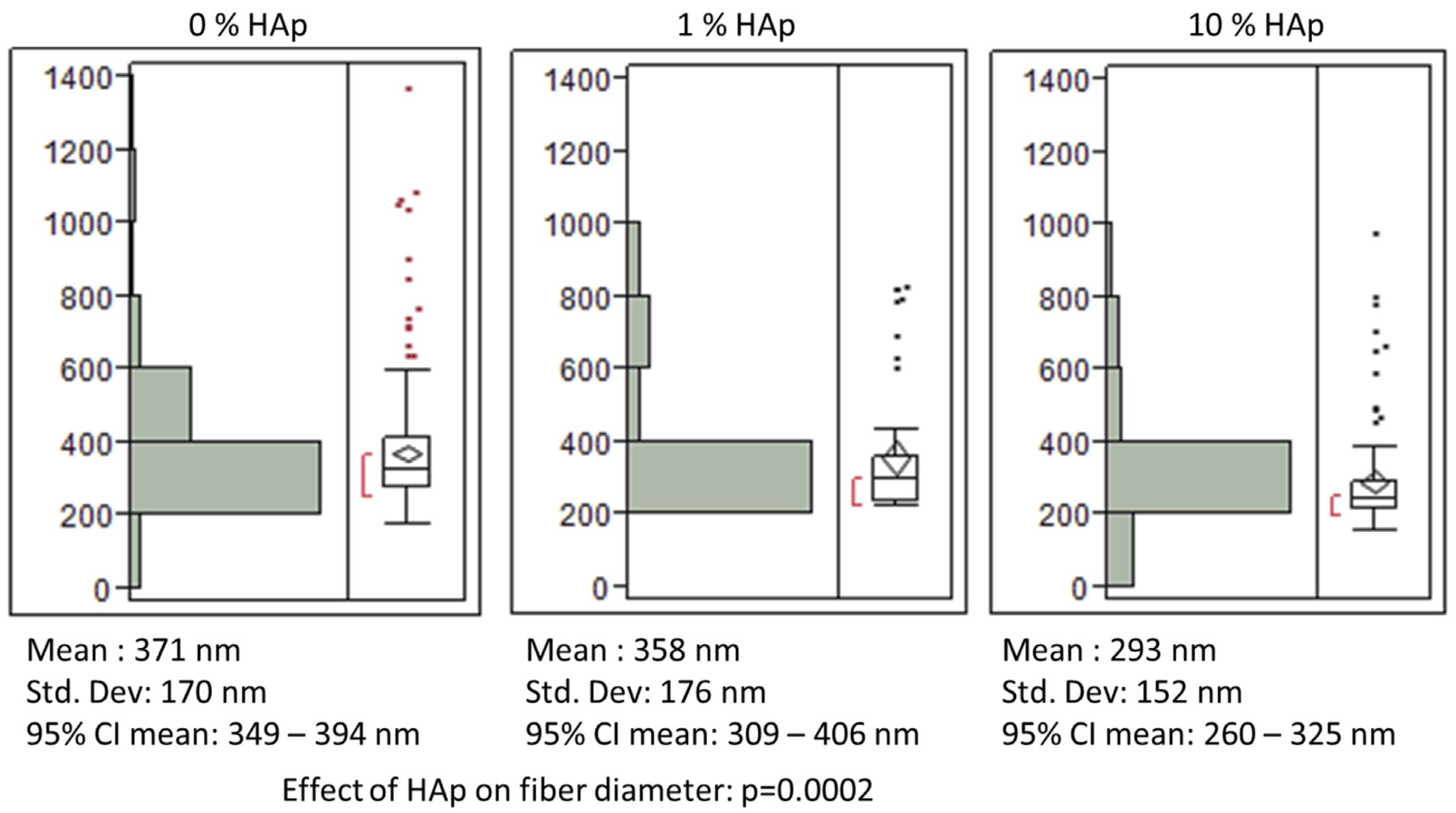
2.1. Fabrication and Characterization of HAp/PCL Composite Nanofiber Scaffolds

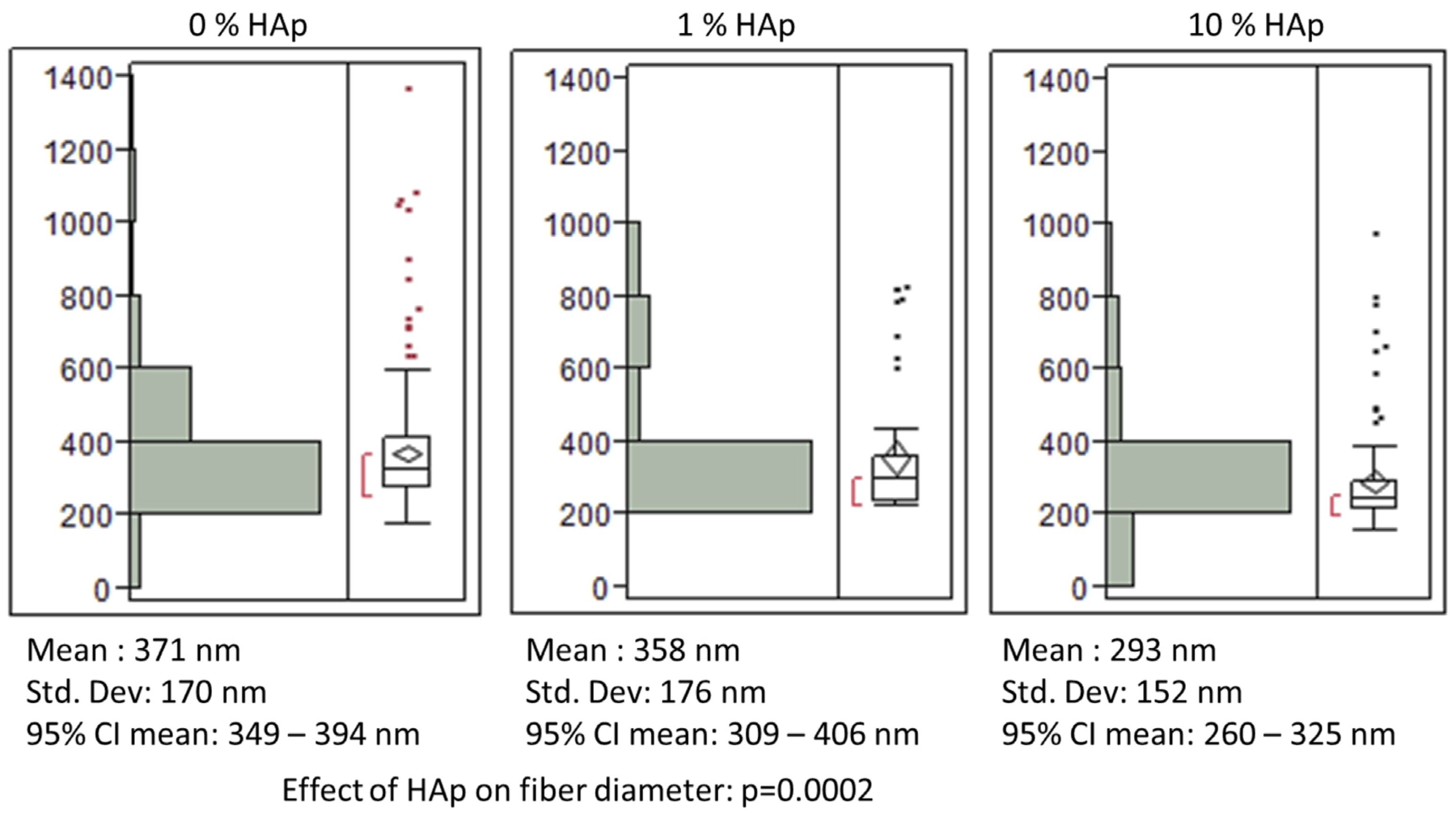
 indicates statistically significant difference (pcrit = 0.05).
indicates statistically significant difference (pcrit = 0.05).
indicates statistically significant difference (pcrit = 0.05).
indicates statistically significant difference (pcrit = 0.05).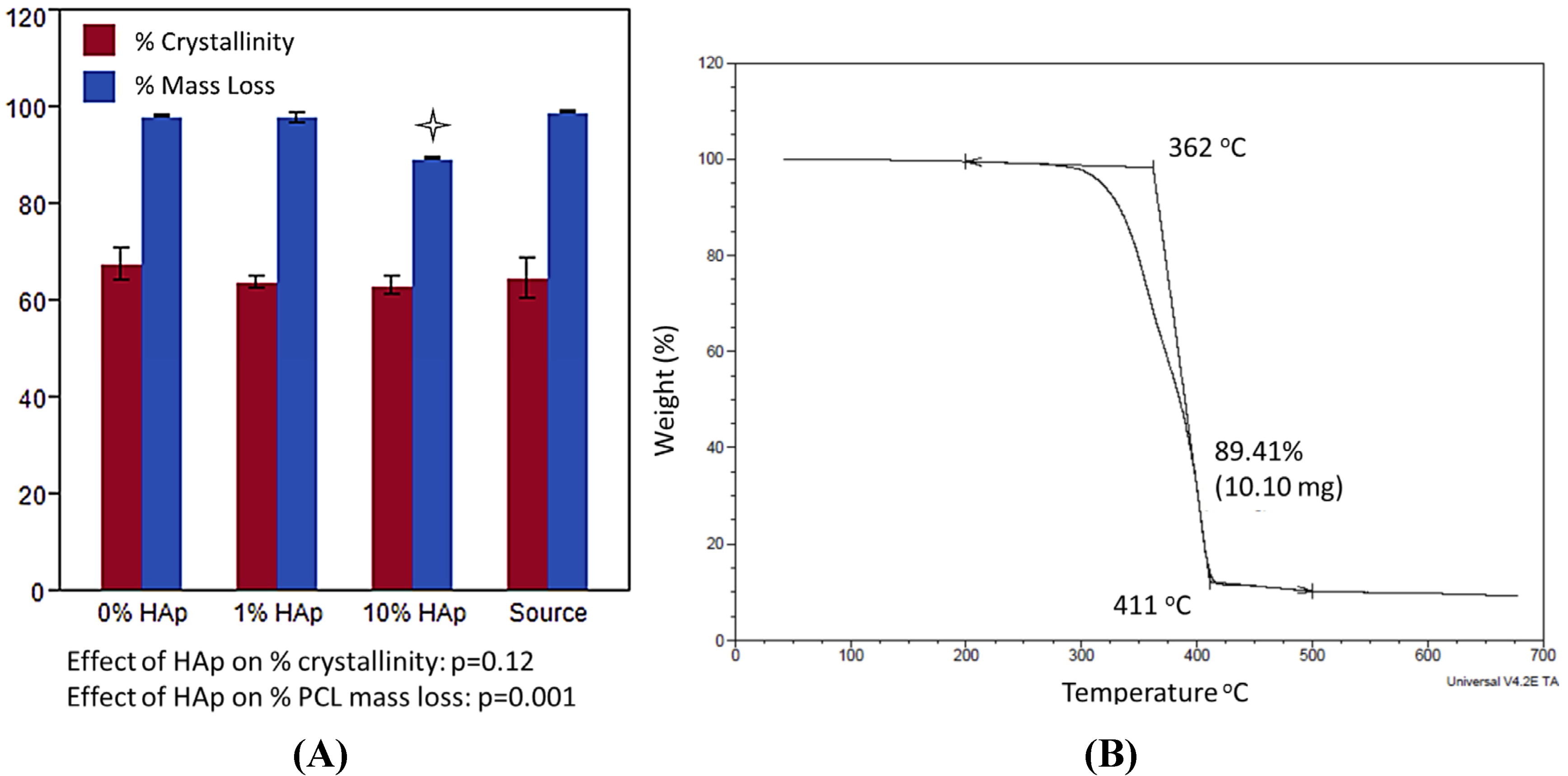
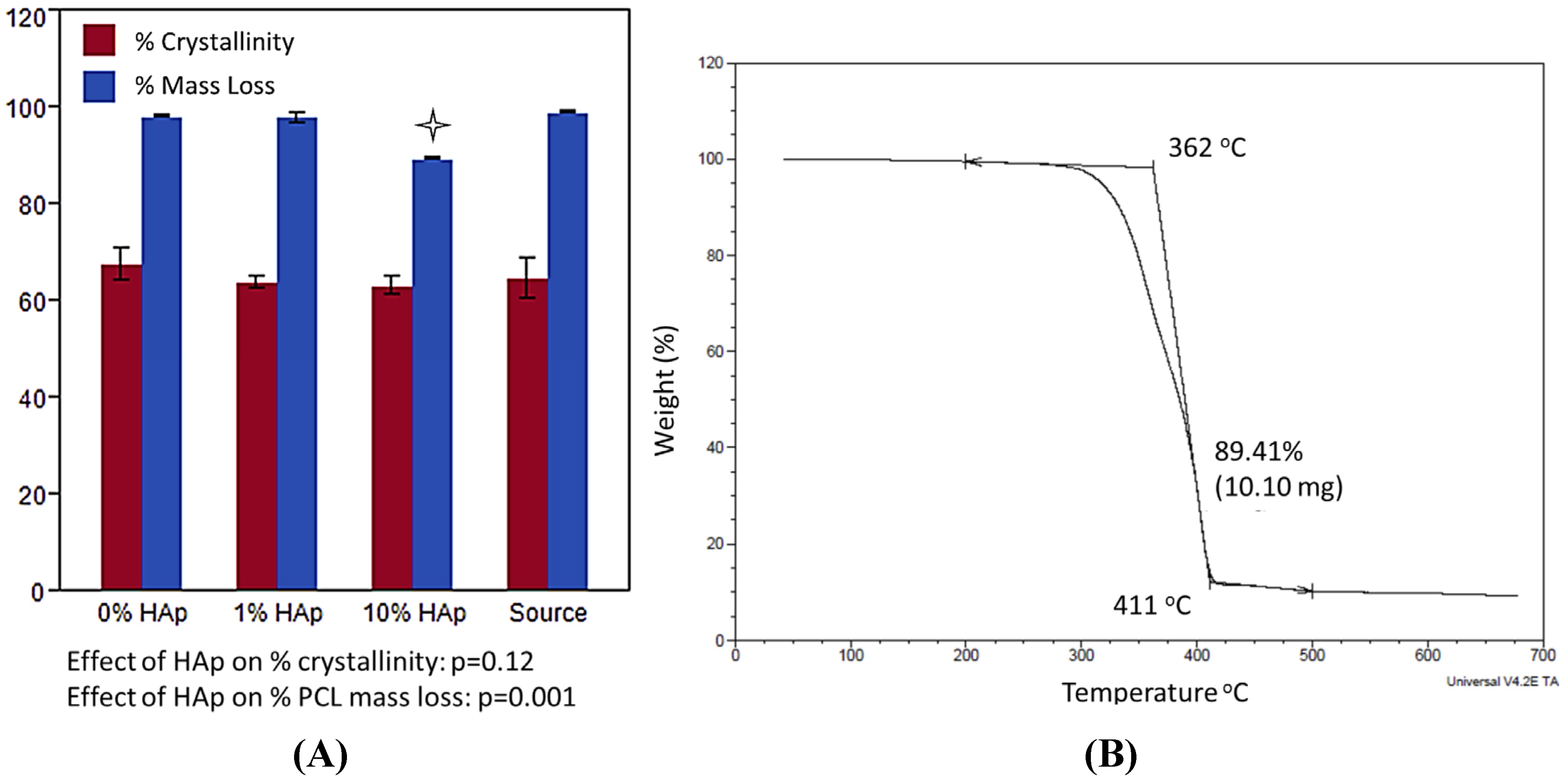
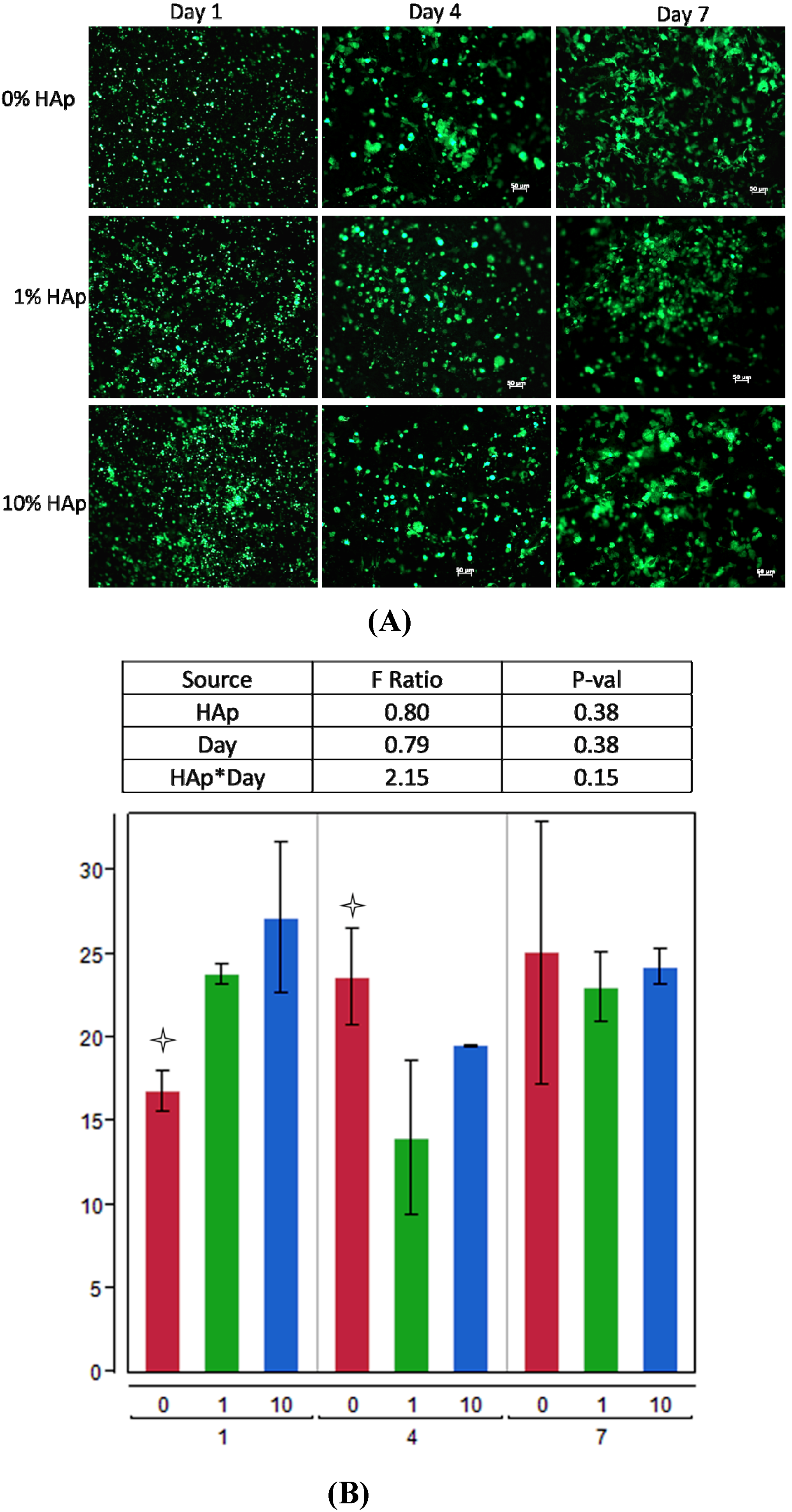
2.2. MSC Response to HAp/PCL Composite Nanofiber Scaffolds in Maintenance Conditions
indicating a statistically significant difference (pcrit = 0.05) from other treatments within the timepoint. An ANOVA analysis is summarized with F-statistics and p-values for the effects of HAp, day, and the interaction of HAp and day.
indicating a statistically significant difference (pcrit = 0.05) from other treatments within the timepoint. An ANOVA analysis is summarized with F-statistics and p-values for the effects of HAp, day, and the interaction of HAp and day. 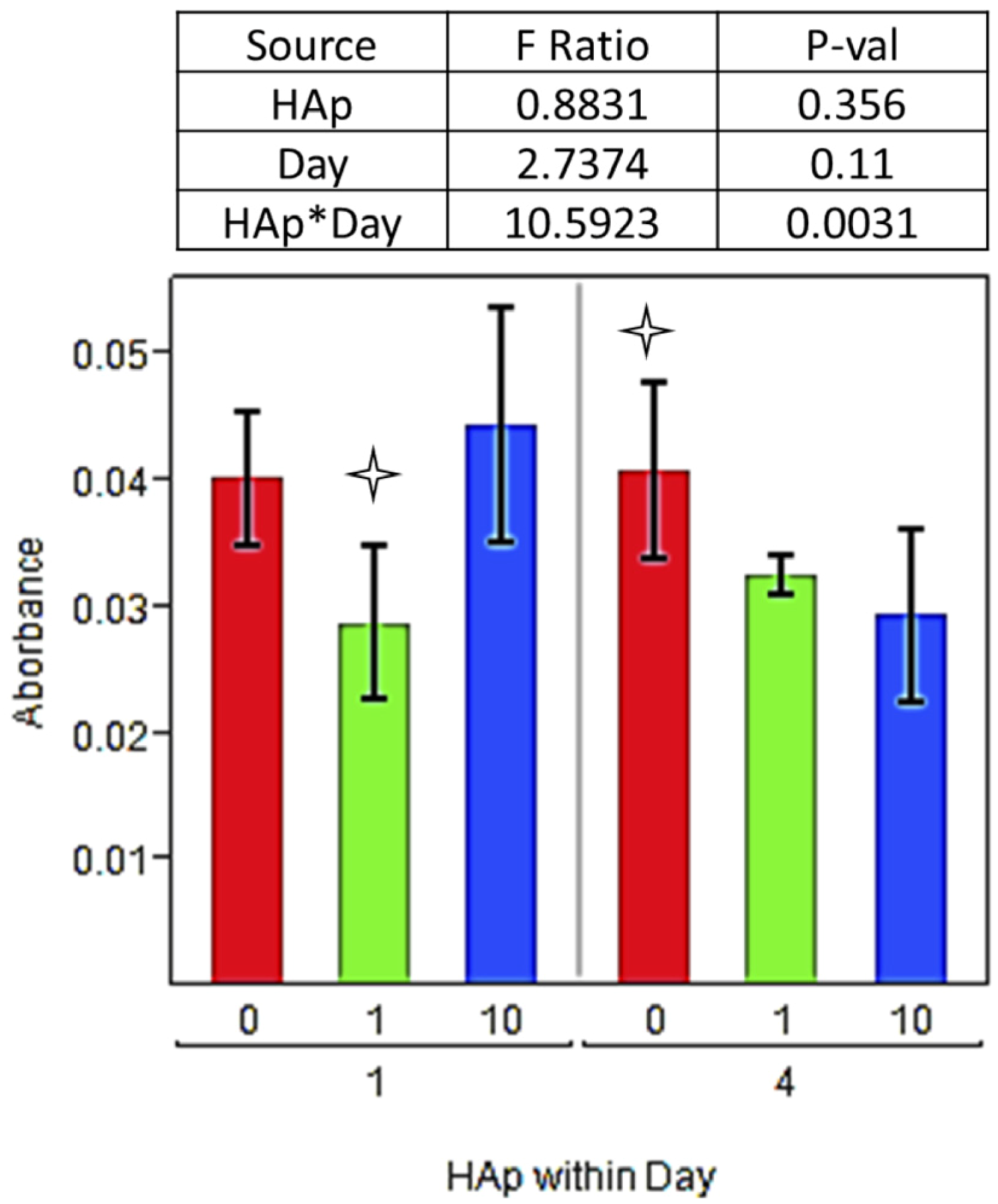 indicating statistically significant difference (pcrit = 0.05) from other treatments. An ANOVA summary is presented with F-statistics and p-values for the effects of HAp, time (day), and the interaction of HAp and time.
indicating statistically significant difference (pcrit = 0.05) from other treatments. An ANOVA summary is presented with F-statistics and p-values for the effects of HAp, time (day), and the interaction of HAp and time.
indicating statistically significant difference (pcrit = 0.05) from other treatments. An ANOVA summary is presented with F-statistics and p-values for the effects of HAp, time (day), and the interaction of HAp and time.
indicating statistically significant difference (pcrit = 0.05) from other treatments. An ANOVA summary is presented with F-statistics and p-values for the effects of HAp, time (day), and the interaction of HAp and time. 
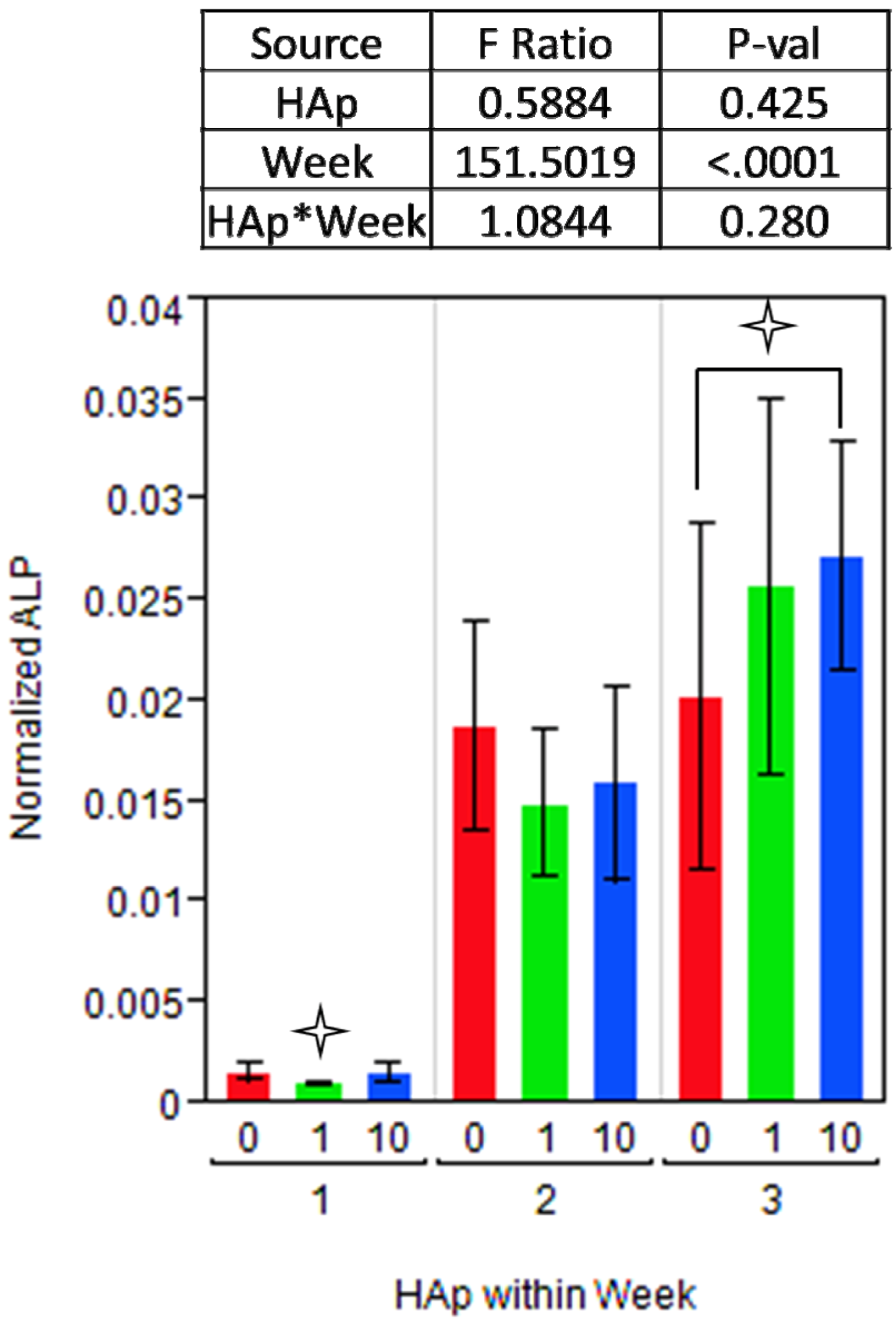
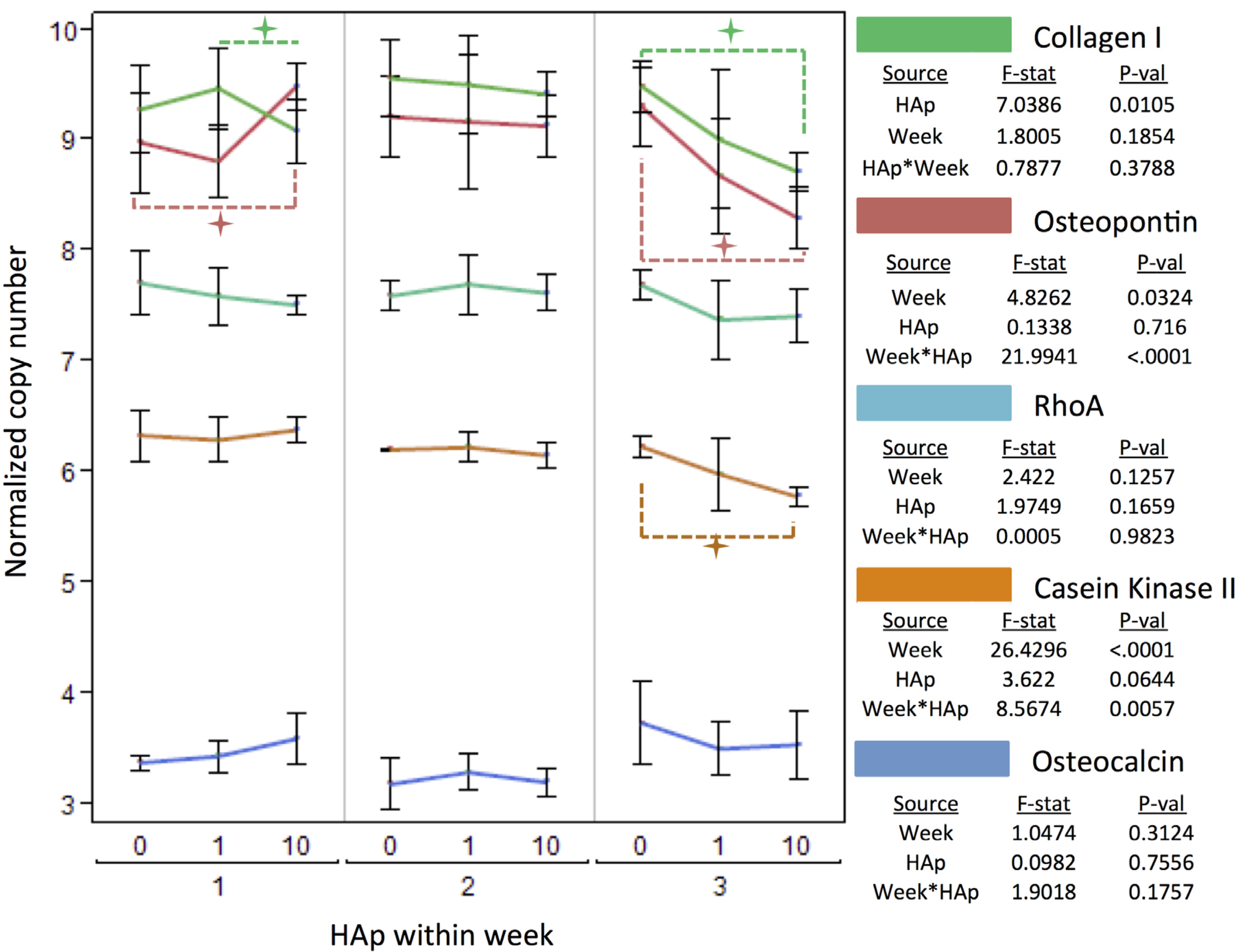
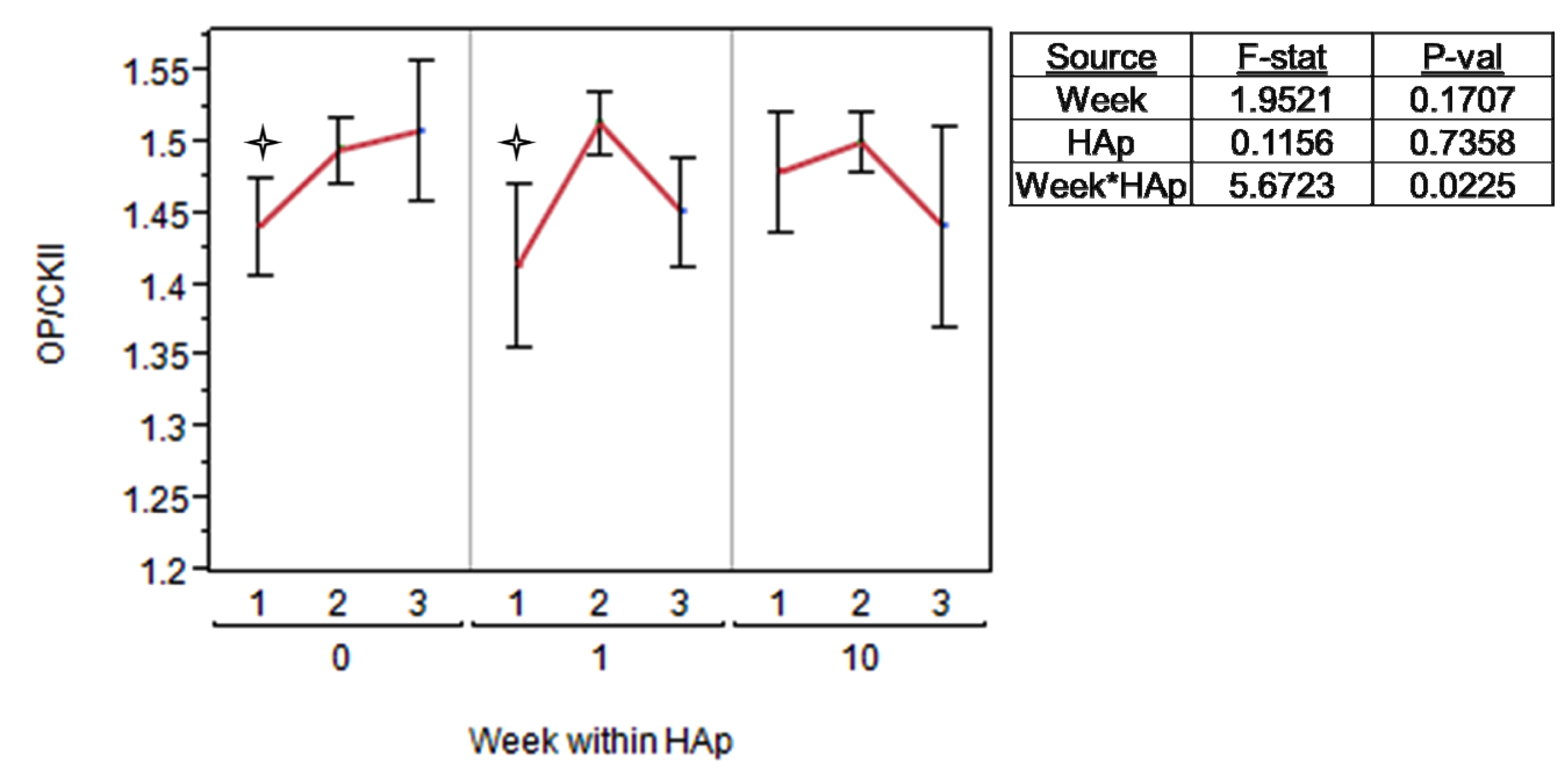
2.3. MSC Differentiation on HAp/PCL Composite Nanofiber Scaffolds
indicates a statistically significant (pcrit=0.05) difference from other treatments. An ANOVA analysis is presented with F-statistics and p-values for the effects of HAp, week, and the interaction of HAp and week.
indicates a statistically significant (pcrit=0.05) difference from other treatments. An ANOVA analysis is presented with F-statistics and p-values for the effects of HAp, week, and the interaction of HAp and week. 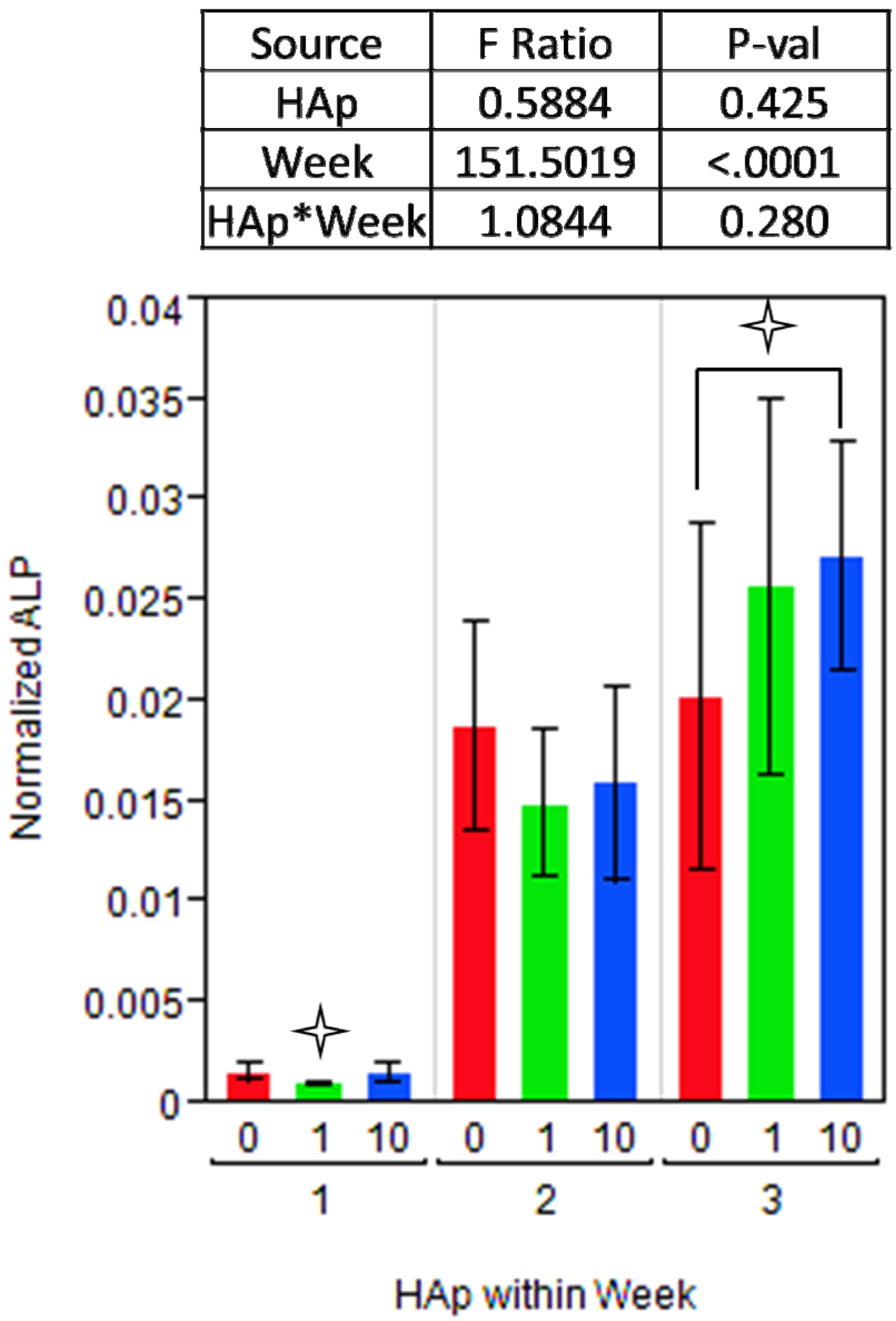 indicates a statistically significance (pcrit =0.05) difference from another treatment within the time point. An ANOVA adjusted for multiple comparisons analyzed the effects of HAp, week, and the interaction of HAp by week, and the F-statistics and p-values are summarized for each gene separately.
indicates a statistically significance (pcrit =0.05) difference from another treatment within the time point. An ANOVA adjusted for multiple comparisons analyzed the effects of HAp, week, and the interaction of HAp by week, and the F-statistics and p-values are summarized for each gene separately.
indicates a statistically significance (pcrit =0.05) difference from another treatment within the time point. An ANOVA adjusted for multiple comparisons analyzed the effects of HAp, week, and the interaction of HAp by week, and the F-statistics and p-values are summarized for each gene separately.
indicates a statistically significance (pcrit =0.05) difference from another treatment within the time point. An ANOVA adjusted for multiple comparisons analyzed the effects of HAp, week, and the interaction of HAp by week, and the F-statistics and p-values are summarized for each gene separately. indicates a statistically significance (pcrit = 0.05) difference from another treatment within the time point.
indicates a statistically significance (pcrit = 0.05) difference from another treatment within the time point.
indicates a statistically significance (pcrit = 0.05) difference from another treatment within the time point.
indicates a statistically significance (pcrit = 0.05) difference from another treatment within the time point. 

3. Experimental Section
3.1. Fabrication and Characterization of HAp/PCL Composite Nanofiber Scaffolds
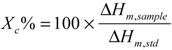
3.2. Rat Marrow Stromal Cell Culture
3.3. MSC Adhesion and Proliferation on HAp/PCL Composite Nanofiber Scaffolds
3.4. Osteogenic Differentiation of MSCs on HAp/PCL Composite Nanofiber Scaffolds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward primer | Reverse primer | Amplicon length |
|---|---|---|---|
| Osteopontin | atcaggacagcaacgggaagac | gagttccaaagccagcctggaa | 224 bp |
| Collagen I | acagaggcataaagggtcatcg | cctggcaaagatggactcaacg | 159 bp |
3.5. Statistical Analysis
4. Conclusions
Acknowledgements
References
- Drosse, I.; Volkmer, E.; Capanna, R.; de Biase, P.; Mutschler, W.; Schieker, M. Tissue engineering for bone defect healing: an update on a multi-component approach. Injury 2008, 39, 9–20. [Google Scholar]
- Heino, T.J.; Hentunen, T.A. Differentiation of osteoblasts and osteocytes from mesenchymal stem cells. Curr. Stem. Cell Res. Ther. 2008, 3, 131–145. [Google Scholar] [CrossRef]
- Haynesworth, S.E.; Goshima, J.; Goldberg, V.M.; Caplan, A.L. Characterization of cells with osteogenic potential from human marrow. Bone 1992, 13, 81–88. [Google Scholar]
- Prockop, D. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef]
- Jorgensen, N.R.; Henriksen, Z.; Sorensen, O.H.; Civitelli, R. Dexamethasone, BMP-2, and 1,25-dihydroxyvitamin D enhance a more differentiated osteoblast phenotype: Validation of an in vitro model for human bone marrow-derived primary osteoblasts. Steroids 2004, 69, 219–226. [Google Scholar] [CrossRef]
- Johnson, M.R.; Lee, H.J.; Bellamkonda, R.V.; Guldberg, R.E. Sustained release of BMP-2 in a lipid-based microtube vehicle. Acta Biomater. 2009, 5, 23–28. [Google Scholar] [CrossRef]
- Laflamme, C.; Rouabhia, M. Effect of BMP-2 and BMP-7 homodimers and a mixture of BMP-2/BMP-7 homodimers on osteoblast adhesion and growth following culture on a collagen scaffold. Biomed. Mater. 2008, 3. [Google Scholar] [CrossRef]
- Laurencin, C.T.; Attawia, M.A.; Lu, L.Q.; Borden, M.D.; Lu, H.H.; Gorum, W.J.; Lieberman, J.R. Poly(lactide-co-glycolide)/hydroxyapatite delivery of BMP-2-producing cells: A regional gene therapy approach to bone regeneration. Biomaterials 2001, 22, 1271–1277. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choo, J.E.; Choi, Y.S.; Suh, J.S.; Lee, S.J.; Chung, C.P.; Park, Y.J. Osteoblastic differentiation of human bone marrow stromal cells in self-assembled BMP-2 receptor-binding peptide-amphiphiles. Biomaterials 2009, 30, 3532–3541. [Google Scholar] [CrossRef]
- Olivares-Navarrete, R.; Raz, P.; Zhao, G.; Chen, J.; Wieland, M.; Cochran, D.L.; Chaudhri, R.A.; Ornoy, A.; Boyan, B.D.; Schwartz, Z. Integrin alpha2beta1 plays a critical role in osteoblast response to micron-scale surface structure and surface energy of titanium substrates. Proc. Natl. Acad. Sci. USA 2008, 105, 15767–15772. [Google Scholar]
- Marletta, G.; Ciapetti, G.; Satriano, C.; Pagani, S.; Baldini, N. The effect of irradiation modification and RGD sequence adsorption on the response of human osteoblasts to polycaprolactone. Biomaterials 2005, 26, 4793–4804. [Google Scholar] [CrossRef]
- Dalby, M.J.; Gadegaard, N.; Tare, R.; Andar, A.; Riehle, M.O.; Herzyk, P.; Wilkinson, C.D.; Oreffo, R.O. The control of human mesenchymal cell differentiation using nanoscale symmetry and disorder. Nat. Mater. 2007, 6, 997–1003. [Google Scholar] [CrossRef]
- Biggs, M.J.; Richards, R.G.; Gadegaard, N.; Wilkinson, C.D.; Oreffo, R.O.; Dalby, M.J. The use of nanoscale topography to modulate the dynamics of adhesion formation in primary osteoblasts and ERK/MAPK signalling in STRO-1+ enriched skeletal stem cells. Biomaterials 2009, 30, 5094–5103. [Google Scholar] [CrossRef]
- Olivares-Navarrete, R.; Hyzy, S.L.; Hutton, D.L.; Erdman, C.P.; Wieland, M.; Boyan, B.D.; Schwartz, Z. Direct and indirect effects of microstructured titanium substrates on the induction of mesenchymal stem cell differentiation towards the osteoblast lineage. Biomaterials 2010, 31, 2728–2735. [Google Scholar]
- Ruckh, T.T.; Kumar, K.; Kipper, M.J.; Popat, K.C. Osteogenic differentiation of bone marrow stromal cells on poly(epsilon-caprolactone) nanofiber scaffolds. Acta Biomater. 2010. [Google Scholar]
- Wutticharoenmongkol, P.; Pavasant, P.; Supaphol, P. Osteoblastic phenotype expression of MC3T3-E1 cultured on electrospun polycaprolactone fiber mats filled with hydroxyapatite nanoparticles. Biomacromolecules 2007, 8, 2602–2610. [Google Scholar] [CrossRef]
- Zhang, P.; Hong, Z.; Yu, T.; Chen, X.; Jing, X. In vivo mineralization and osteogenesis of nanocomposite scaffold of poly(lactide-co-glycolide) and hydroxyapatite surface-grafted with poly(l-lactide). Biomaterials 2008. [Google Scholar]
- Lee, J.H.; Rim, N.G.; Jung, H.S.; Shin, H. Control of osteogenic differentiation and mineralization of human mesenchymal stem cells on composite nanofibers containing poly[lactic-co-(glycolic acid)] and hydroxyapatite. Macromol. Biosci. 2010, 10, 173–182. [Google Scholar]
- Gupta, D.; Venugopal, J.; Mitra, S.; Giri Dev, V.R.; Ramakrishna, S. Nanostructured biocomposite substrates by electrospinning and electrospraying for the mineralization of osteoblasts. Biomaterials 2009, 30, 2085–2094. [Google Scholar]
- Ramakrishna, S.; Fujihara, K.; Teo, W.-E.; Lim, T.-C.; Ma, Z. An Introduction to Electrospinning and Nanofibers; World Scientific: Singapore, 2005; p. 382. [Google Scholar]
- Rutledge, G.C.; Fridrikh, S.V. Formation of fibers by electrospinning. Adv. Drug Deliv. Rev. 2007, 59, 1384–1391. [Google Scholar] [CrossRef]
- Theron, S.A.; Zussman, E.; Yarin, A.L. Experimental investigation of the governing parameters in the electrospinning of polymer solutions. Polymer 2004, 45, 2017–2030. [Google Scholar]
- Sinha, V.R.; Bansal, K.; Kaushik, R.; Kumria, R.; Trehan, A. Poly-e-caprolactone microspheres and nanospheres: An overview. Int. J. Pharm. 2004, 278, 1–23. [Google Scholar]
- Zeng, J.; Xu, X.; Chen, X.; Liang, Q.; Bian, X.; Yang, L.; Jing, X. Biodegradable electrospun fibers for drug delivery. J. Control. Release 2003, 92, 227–231. [Google Scholar] [CrossRef]
- Pulkkinen, M.; Malin, M.; Tarvainen, T.; Saarimäki, T.; Seppälä, J.; Järvinen, K. Effects of block length on the enzymatic degradation and erosion of oxazoline linked poly-e-caprolactone. Eur. J. Pharm. Sci. 2007, 31, 119–128. [Google Scholar] [CrossRef]
- Sangsanoh, P.; Waleetorncheepsawat, S.; Suwantong, O.; Wutticharoenmongkol, P.; Weeranantanapan, O.; Chuenjitbuntaworn, B.; Cheepsunthorn, P.; Pavasant, P.; Supaphol, P. In vitro biocompatibility of schwann cells on surfaces of biocompatible polymeric electrospun fibrous and solution-cast film scaffolds. Biomacromolecules 2007, 8, 1587–1594. [Google Scholar] [CrossRef]
- Palmer, L.C.; Newcomb, C.J.; Kaltz, S.R.; Spoerke, E.D.; Stupp, S.I. Biomimetic systems for hydroxyapatite mineralization inspired by bone and enamel. Chem. Rev. 2008, 108, 4754–4783. [Google Scholar]
- Anderson, H.C.; Sipe, J.B.; Hessle, L.; Dhanyamraju, R.; Atti, E.; Camacho, N.P.; Millan, J.L. Impaired calcification around matrix vesicles of growth plate and bone in alkaline phosphatase-deficient mice. Am. J. Pathol. 2004, 164, 841–847. [Google Scholar]
- Genge, B.R.; Wu, L.N.; Wuthier, R.E. In vitro modeling of matrix vesicle nucleation: Synergistic stimulation of mineral formation by annexin A5 and phosphatidylserine. J. Biol. Chem. 2007, 282, 26035–26045. [Google Scholar]
- Nikolov, S.; Raabe, D. Hierarchical modeling of the elastic properties of bone at submicron scales: The role of extrafibrillar mineralization. Biophys. J. 2008, 94, 4220–4232. [Google Scholar] [CrossRef]
- Norman, J.; Shapter, J.G.; Short, K.; Smith, L.J.; Fazzalari, N.L. Micromechanical properties of human trabecular bone: A hierarchical investigation using nanoindentation. J. Biomed. Mater. Res. A 2008, 87, 196–202. [Google Scholar]
- McCreadie, B.R.; Goulet, R.W.; Feldkamp, L.A.; Goldstein, S.A. Hierarchical structure of bone and micro-computed tomography. Adv. Exp. Med. Biol. 2001, 496, 67–83. [Google Scholar]
- White, D.J.; Puranen, S.; Johnson, M.S.; Heino, J. The collagen receptor subfamily of the integrins. Int. J. Biochem. Cell Biol. 2004, 36, 1405–1410. [Google Scholar] [CrossRef]
- Denhardt, D.T.; Noda, M. Osteopontin expression and function: Role in bone remodeling. J. Cell Biochem. Suppl. 1998, 30-31, 92–102. [Google Scholar] [CrossRef]
- Kazanecki, C.C.; Uzwiak, D.J.; Denhardt, D.T. Control of osteopontin signaling and function by post-translational phosphorylation and protein folding. J. Cell Biochem. 2007, 102, 912–924. [Google Scholar] [CrossRef]
- McKee, M.D.; Nanci, A. Osteopontin: An interfacial extracellular matrix protein in mineralized tissues. Connect. Tissue Res. 1996, 35, 197–205. [Google Scholar] [CrossRef]
- Cui, W.; al, e. In situ growth of hydroxyapatite within electrospun poly(DL-lactide) fibers. J. Biomed. Mater. Res. Part A 2007, 82, 831–841. [Google Scholar]
- Fang, B.; Wan, Y.Z.; Tang, T.T.; Gao, C.; Dai, K.R. Proliferation and osteoblastic differentiation of human bone marrow stromal cells on hydroxyapatite/bacterial cellulose nanocomposite scaffolds. Tissue Eng. Part A 2009, 15, 1091–1098. [Google Scholar] [CrossRef]
- Guarino, V.; Taddei, P.; Di Foggia, M.; Fagnano, C.; Ciapetti, G.; Ambrosio, L. The influence of hydroxyapatite particles on in vitro degradation behaviour of pcl based composite scaffolds. Tissue Eng. Part A 2009, 15, 3655–3668. [Google Scholar] [CrossRef]
- Mei, F.; Zhong, J.; Yang, X.; Ouyang, X.; Zhang, S.; Hu, X.; Ma, Q.; Lu, J.; Ryu, S.; Deng, X. Improved biological characteristics of poly(L-lactic acid) electrospun membrane by incorporation of multiwalled carbon nanotubes/hydroxyapatite nanoparticles. Biomacromolecules 2007, 8, 3729–3735. [Google Scholar] [CrossRef]
- Venugopal, J.R.; Low, S.; Choon, A.T.; Kumar, A.B.; Ramakrishna, S. Nanobioengineered electrospun composite nanofibers and osteoblasts for bone regeneration. Artif Organs 2008, 32, 388–397. [Google Scholar] [CrossRef]
- Kim, H.-W.; Lee, H.-H.; Knowles, J.C. Electrospinning biomedical nanocomposite fibers of hydroxyapaite/poly(lactic acid) for bone regeneration. J. Biomed. Mater. Res. Part A 2006, 79, 643–649. [Google Scholar]
- Jackson, S.M.; Demer, L.L. Peroxisome proliferator-activated receptor activators modulate the osteoblastic maturation of MC3T3-E1 preosteoblasts. FEBS Lett. 2000, 471, 119–124. [Google Scholar] [CrossRef]
- Still, K.; Grabowski, P.; Mackie, I.; Perry, M.; Bishop, N. The peroxisome proliferator activator receptor alpha/delta agonists linoleic acid and bezafibrate upregulate osteoblast differentiation and induce periosteal bone formation in vivo. Calcif. Tissue Int. 2008, 83, 285–292. [Google Scholar] [CrossRef]
- Xu, H.E.; Lambert, M.H.; Montana, V.G.; Parks, D.J.; Blanchard, S.G.; Brown, P.J.; Sternbach, D.D.; Lehmann, J.M.; Wisely, G.B.; Willson, T.M.; Kliewer, S.A.; Milburn, M.V. Molecular recognition of fatty acids by peroxisome proliferator-activated receptors. Mol. Cell 1999, 3, 397–403. [Google Scholar] [CrossRef]
- Berger, J.; Moller, D. The mechanisms of action of PPARs. Annu. Rev. Med. 2002, 53, 409–435. [Google Scholar] [CrossRef]
- Lecka-Czernik, B.; Suva, L.J. Resolving the two “bony” faces of PPAR-gamma. PPAR Res. 2006. [Google Scholar] [CrossRef]
- Lin, T.-H.; Yang, R.-S.; Tang, C.-H.; Lin, C.-P.; Fu, W.-M. PPARg inhibits osteogenesis via the down-regulation of the expression of COX-2 and iNOS in rats. Bone 2007, 41, 562–574. [Google Scholar]
- Syversen, U.; Bakke, I.; Aune, G.; Thommesen, L. PPAR-Alpha agonists increase bone mineral density in female rats. J. Bone Miner. Res. 2003, 18, S2–S56. [Google Scholar]
- De Jong, D.S.; Vaes, B.L.; Dechering, K.J.; Feijen, A.; Hendriks, J.M.; Wehrens, R.; Mummery, C.L.; van Zoelen, E.J.; Olijve, W.; Steegenga, W.T. Identification of novel regulators associated with early-phase osteoblast differentiation. J. Bone Miner. Res. 2004, 19, 947–958. [Google Scholar]
- Badami, A.S.; Kreke, M.R.; Thompson, M.S.; Riffle, J.S.; Goldstein, A.S. Effect of fiber diameter on spreading, proliferation, and differentiation of osteoblastic cells on electrospun poly(lactic acid) substrates. Biomaterials 2006, 27, 596–606. [Google Scholar] [CrossRef]
- Bashur, C.A.; Dahlgren, L.A.; Goldstein, A.S. Effect of fiber diameter and orientation on fibroblast morphology and proliferation on electrospun poly(D,L-lactic-co-glycolic acid) meshes. Biomaterials 2006, 27, 5681–5688. [Google Scholar] [CrossRef]
- Tsiridis, E.; Upadhyay, N.; Giannoudis, P. Molecular aspects of fracture healing: Which are the important molecules? Injury 2007, 38, 11–25. [Google Scholar]
- Chen, L.; Jiang, W.; Huang, J.; He, B.C.; Zuo, G.W.; Zhang, W.; Luo, Q.; Shi, Q.; Zhang, B.Q.; Wagner, E.R.; et al. Insulin-like growth factor 2 (IGF2) potentiates BMP9-induced osteogenic differentiation and bone formation. J. Bone Miner. Res. 2010, 25, 2447–2459. [Google Scholar] [CrossRef]
- Sundelacruz, S.; Kaplan, D.L. Stem cell- and scaffold-based tissue engineering approaches to osteochondral regenerative medicine. Semin. Cell Dev. Biol. 2009, 20, 646–655. [Google Scholar] [CrossRef]
- Ekholm, E.C.; Ravanti, L.; Kahari, V.; Paavolainen, P.; Penttinen, R.P. Expression of extracellular matrix genes: transforming growth factor (TGF)-beta1 and ras in tibial fracture healing of lathyritic rats. Bone 2000, 27, 551–557. [Google Scholar] [CrossRef]
- Hirakawa, K.; Hirota, S.; Ikeda, T.; Yamaguchi, A.; Takemura, T.; Nagoshi, J.; Yoshiki, S.; Suda, T.; Kitamura, Y.; Nomura, S. Localization of the mRNA for bone matrix proteins during fracture healing as determined by in situ hybridization. J. Bone Miner. Res. 1994, 9, 1551–1557. [Google Scholar]
- Schuckert, K.H.; Jopp, S.; Teoh, S.H. Mandibular defect reconstruction using three-dimensional polycaprolactone scaffold in combination with platelet-rich plasma and recombinant human bone morphogenetic protein-2: De novo synthesis of bone in a single case. Tissue Eng. Part A 2009, 15, 493–499. [Google Scholar]
- Thorwarth, M.; Rupprecht, S.; Falk, S.; Felszeghy, E.; Wiltfang, J.; Schlegel, K.A. Expression of bone matrix proteins during de novo bone formation using a bovine collagen and platelet-rich plasma (prp)—an immunohistochemical analysis. Biomaterials 2005, 26, 2575–2584. [Google Scholar]
- Kosaki, N.; Takaishi, H.; Kamekura, S.; Kimura, T.; Okada, Y.; Minqi, L.; Amizuka, N.; Chung, U.I.; Nakamura, K.; Kawaguchi, H.; et al. Impaired bone fracture healing in matrix metalloproteinase-13 deficient mice. Biochem. Biophys. Res. Commun. 2007, 354, 846–851. [Google Scholar] [CrossRef]
- Weiss, S.; Zimmermann, G.; Baumgart, R.; Kasten, P.; Bidlingmaier, M.; Henle, P. Systemic regulation of angiogenesis and matrix degradation in bone regeneration—distraction osteogenesis compared to rigid fracture healing. Bone 2005, 37, 781–790. [Google Scholar] [CrossRef]
- Henle, P.; Zimmermann, G.; Weiss, S. Matrix metalloproteinases and failed fracture healing. Bone 2005, 37, 791–798. [Google Scholar] [CrossRef]
- Coleman, J.E. Structure and mechanism of alkaline phosphatase. Annu. Rev. Biophys. Biomol. Struct. 1992, 21, 441–483. [Google Scholar] [CrossRef]
- Orimo, H.; Shimada, T. The role of tissue-nonspecific alkaline phosphatase in the phosphate-induced activation of alkaline phosphatase and mineralization in SaOS-2 human osteoblast-like cells. Mol. Cell. Biochem. 2008, 315, 51–60. [Google Scholar] [CrossRef]
- Kulterer, B.; Friedl, G.; Jandrositz, A.; Sanchez-Cabo, F.; Prokesch, A.; Paar, C.; Scheideler, M.; Windhager, R.; Preisegger, K.H.; Trajanoski, Z. Gene expression profiling of human mesenchymal stem cells derived from bone marrow during expansion and osteoblast differentiation. BMC Genomics. 2007, 8. [Google Scholar] [CrossRef]
- Park, S.-H.; Kim, T.-L.; Ku, Y.; Chung, C.-P.; Han, S.-B.; Yu, J.-H.; Lee, S.-P.; Kim, H.-W.; Lee, H.-H. Effect of hydroxyapatite-coated nanofibrous membrane on the responses of human periodontal ligament fibroblast. J. Ceram. Soc. Jpn. 2008, 116, 31–35. [Google Scholar] [CrossRef]
- Venugopal, J.; Low, S.; Choon, A.; Kumar, A.; Ramakrishna, S. Electrospun-modified nanofibrous scaffolds for the mineralization of osteoblast cells. J. Biomed. Mater. Res. Part A 2007, 85, 408–417. [Google Scholar]
- Salih, E.; Ashkar, S.; Gerstenfeld, L.C.; Glimcher, M.J. Protein kinases of cultured osteoblasts: Selectivity for the extracellular matrix proteins of bone and their catalytic competence for osteopontin. J. Bone Miner. Res. 1996, 11, 1461–1473. [Google Scholar]
- Katayama, Y.; House, C.M.; Udagawa, N.; Kazama, J.J.; McFarland, R.J.; Martin, T.J.; Findlay, D.M. Casein kinase 2 phosphorylation of recombinant rat osteopontin enhances adhesion of osteoclasts but not osteoblasts. J. Cell Physiol. 1998, 176, 179–187. [Google Scholar] [CrossRef]
- Wang, J.; Glimcher, M.J.; Mah, J.; Zhou, H.Y.; Salih, E. Expression of bone microsomal casein kinase II, bone sialoprotein, and osteopontin during the repair of calvarial defects. Bone 1998, 22, 621–628. [Google Scholar] [CrossRef]
- Liu, F.; Malaval, L.; Aubin, J.E. Global amplification polymerase chain reaction reveals novel transitional stages during osteoprogenitor differentiation. J. Cell Sci. 2003, 116, 1787–1796. [Google Scholar] [CrossRef]
- Hamada, K.; Hirose, M.; Yamashita, T.; Ohgushi, H. Spatial distribution of mineralized bone matrix produced by marrow mesenchymal stem cells in self-assembling peptide hydrogel scaffold. J. Biomed. Mater. Res. A 2008, 84, 128–136. [Google Scholar]
- Robey, P.G.; Fedarko, N.S.; Hefferan, T.E.; Bianco, P.; Vetter, U.K.; Grzesik, W.; Friedenstein, A.; van der Pluijm, G.; Mintz, K.P.; Young, M.F.; et al. Structure and molecular regulation of bone matrix proteins. J. Bone Miner. Res. 1993, 8, 483–487. [Google Scholar]
- Young, M.F. Bone matrix proteins: Their function, regulation, and relationship to osteoporosis. Osteoporos. Int. 2003, 14, 35–42. [Google Scholar]
- Qin, C.; Baba, O.; Butler, W.T. Post-translational modifications of sibling proteins and their roles in osteogenesis and dentinogenesis. Crit. Rev. Oral Biol. Med. 2004, 15, 126–136. [Google Scholar] [CrossRef]
- Keykhosravani, M.; Doherty-Kirby, A.; Zhang, C.; Brewer, D.; Goldberg, H.A.; Hunter, G.K.; Lajoie, G. Comprehensive identification of post-translational modifications of rat bone osteopontin by mass spectrometry. Biochemistry 2005, 44, 6990–7003. [Google Scholar]
- Goldberg, H.A.; Hunter, G.K. The inhibitory activity of osteopontin on hydroxyapatite formation in vitro. Ann. NY. Acad. Sci. 1995, 760, 305–308. [Google Scholar] [CrossRef]
- Pampena, D.A.; Robertson, K.A.; Litvinova, O.; Lajoie, G.; Goldberg, H.A.; Hunter, G.K. Inhibition of hydroxyapatite formation by osteopontin phosphopeptides. Biochem. J. 2004, 378, 1083–1087. [Google Scholar]
- Rajachar, R.M.; Truong, A.Q.; Giachelli, C.M. The influence of surface mineral and osteopontin on the formation and function of murine bone marrow-derived osteoclasts. J. Mater. Sci. Mater. Med. 2008, 19, 3279–3285. [Google Scholar] [CrossRef]
- Sodek, J.; Zhu, B.; Huynh, M.H.; Brown, T.J.; Ringuette, M. Novel functions of the matricellular proteins osteopontin and osteonectin/SPARC. Connect. Tissue Res. 2002, 43, 308–319. [Google Scholar]
- Taller, A.; Grohe, B.; Rogers, K.A.; Goldberg, H.A.; Hunter, G.K. Specific adsorption of osteopontin and synthetic polypeptides to calcium oxalate monohydrate crystals. Biophys. J. 2007, 93, 1768–1777. [Google Scholar] [CrossRef]
- McBeath, R.; Pirone, D.M.; Nelson, C.M.; Bhadriraju, K.; Chen, C.S. Cell shape, cytoskeletal tension, and RhoA regulate stem cell lineage commitment. Dev. Cell 2004, 6, 483–495. [Google Scholar] [CrossRef]
- Meyers, V.E.; Zayzafoon, M.; Douglas, J.T.; McDonald, J.M. RhoA and cytoskeletal disruption mediate reduced osteoblastogenesis and enhanced adipogenesis of human mesenchymal stem cells in modeled microgravity. J. Bone Miner. Res. 2005, 20, 1858–1866. [Google Scholar]
- Khatiwala, C.B.; Kim, P.D.; Peyton, S.R.; Putnam, A.J. ECM compliance regulates osteogenesis by influencing MAPK signaling downstream of RhoA and ROCK. J. Bone Miner. Res. 2009, 24, 886–898. [Google Scholar]
- Yoshida, T.; Clark, M.F.; Stern, P.H. The small GTPase RhoA is crucial for MC3T3-E1 osteoblastic cell survival. J. Cell Biochem. 2009, 106, 896–902. [Google Scholar] [CrossRef]
- Chan, M.C.; Nguyen, P.H.; Davis, B.N.; Ohoka, N.; Hayashi, H.; Du, K.; Lagna, G.; Hata, A. A novel regulatory mechanism of the bone morphogenetic protein (BMP) signaling pathway involving the carboxyl-terminal tail domain of BMP type II receptor. Mol. Cell. Biol. 2007, 27, 5776–5789. [Google Scholar]
- Zambuzzi, W.F.; Bruni-Cardoso, A.; Granjeiro, J.M.; Peppelenbosch, M.P.; de Carvalho, H.F.; Aoyama, H.; Ferreira, C.V. On the road to understanding of the osteoblast adhesion: Cytoskeleton organization is rearranged by distinct signaling pathways. J. Cell Biochem. 2009, 108, 134–144. [Google Scholar] [CrossRef]
- Robling, A.G.; Castillo, A.B.; Turner, C.H. Biomechanical and molecular regulation of bone remodeling. Annu. Rev. Biomed. Eng. 2006, 8, 455–498. [Google Scholar] [CrossRef]
- Fregel, R.; Gonzalez, A.; Cabrera, V.M. Improved ethanol precipitation of DNA. Electrophoresis 2010, 31, 1350–1352. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ruckh, T.T.; Carroll, D.A.; Weaver, J.R.; Popat, K.C. Mineralization Content Alters Osteogenic Responses of Bone Marrow Stromal Cells on Hydroxyapatite/Polycaprolactone Composite Nanofiber Scaffolds. J. Funct. Biomater. 2012, 3, 776-798. https://doi.org/10.3390/jfb3040776
Ruckh TT, Carroll DA, Weaver JR, Popat KC. Mineralization Content Alters Osteogenic Responses of Bone Marrow Stromal Cells on Hydroxyapatite/Polycaprolactone Composite Nanofiber Scaffolds. Journal of Functional Biomaterials. 2012; 3(4):776-798. https://doi.org/10.3390/jfb3040776
Chicago/Turabian StyleRuckh, Timothy T., Derek A. Carroll, Justin R. Weaver, and Ketul C. Popat. 2012. "Mineralization Content Alters Osteogenic Responses of Bone Marrow Stromal Cells on Hydroxyapatite/Polycaprolactone Composite Nanofiber Scaffolds" Journal of Functional Biomaterials 3, no. 4: 776-798. https://doi.org/10.3390/jfb3040776