Shift in Food Intake and Changes in Metabolic Regulation and Gene Expression during Simulated Night-Shift Work: A Rat Model

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Animals and Housing
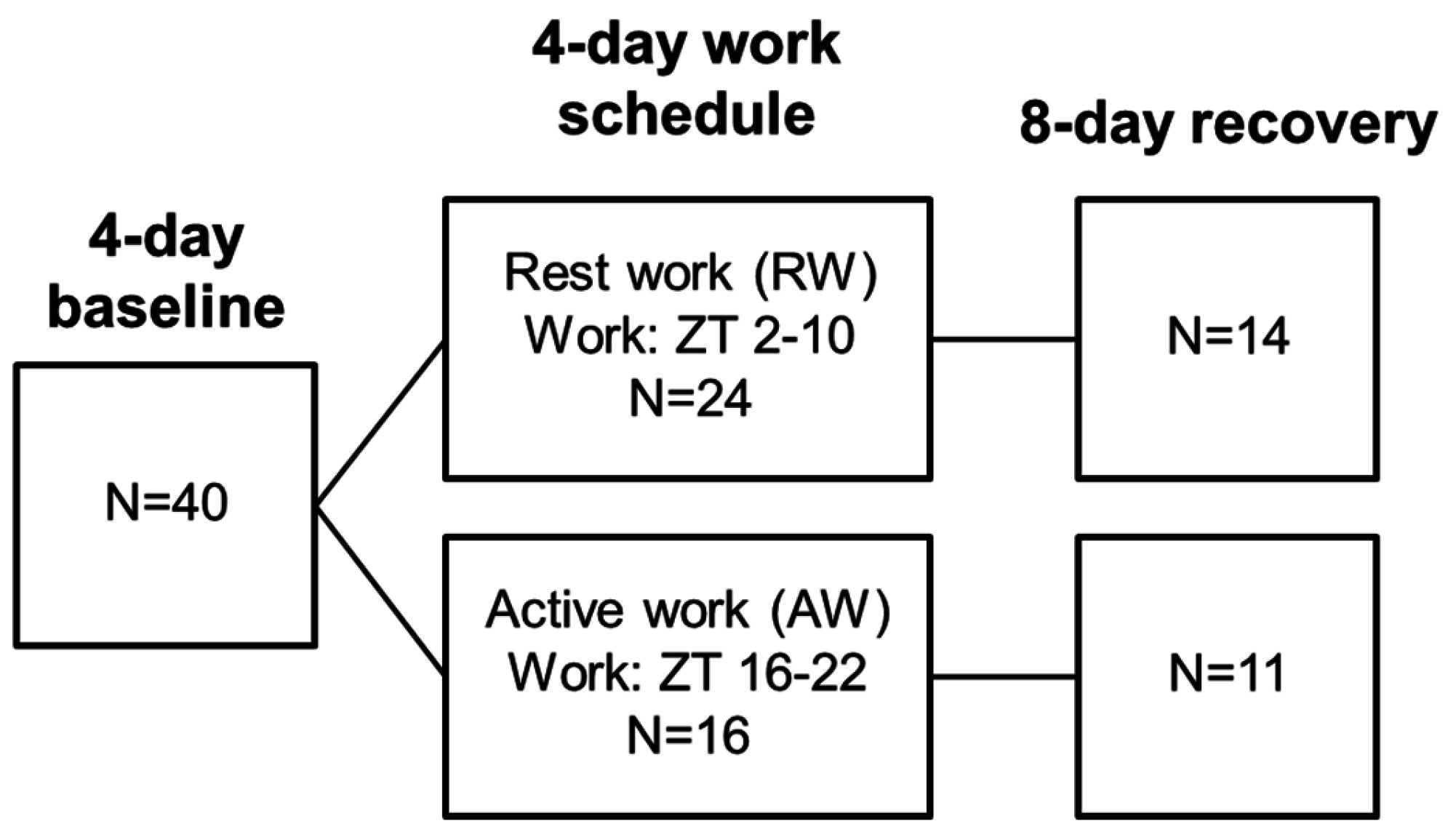
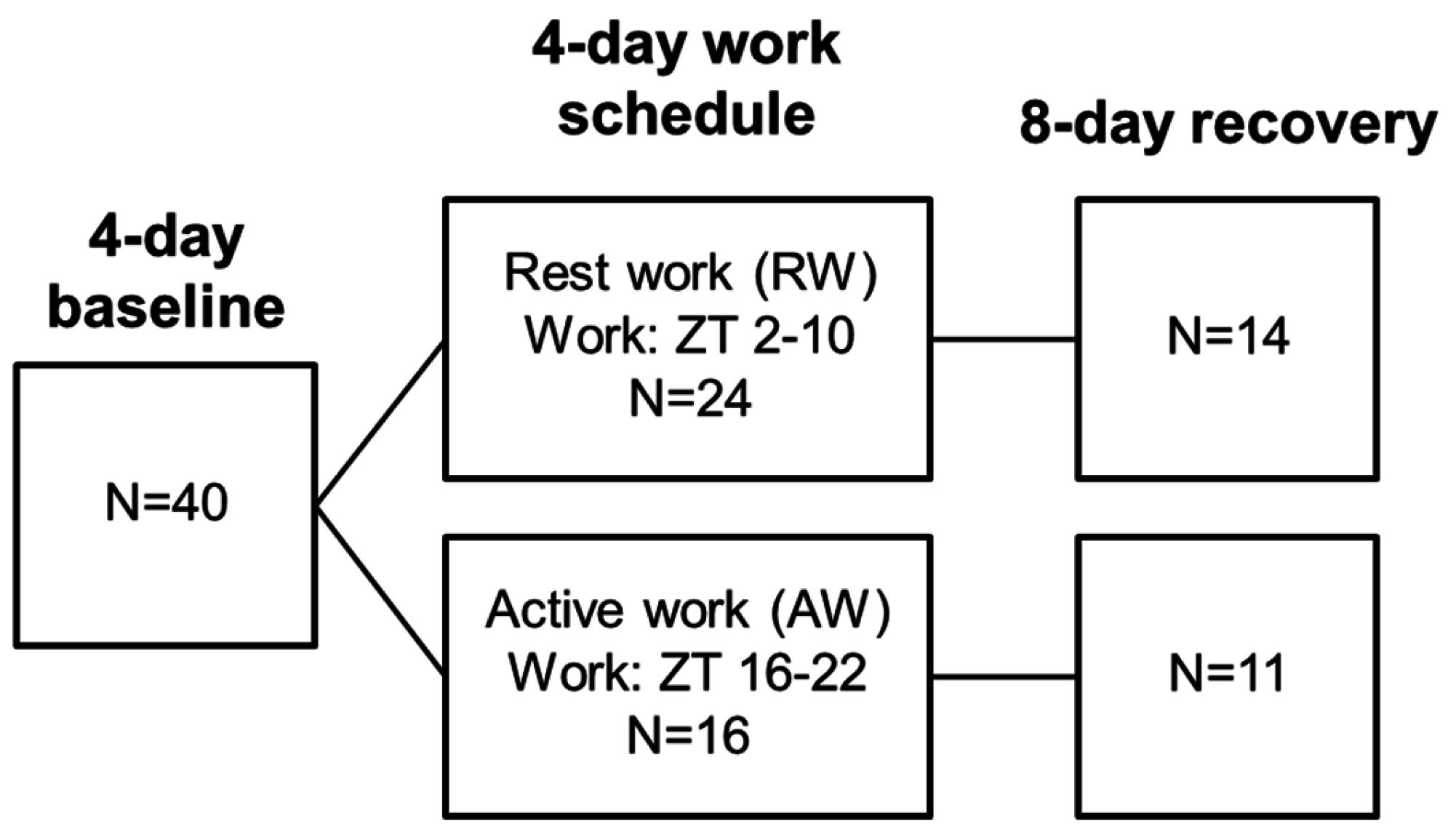
2.3. Study Design
2.4. Simulated Shift Work Procedure
2.5. Telemetric Recording and Analyses of Body Temperature
2.6. Body Weight and Food Intake Measurements
2.7. Assessment of Metabolic Gene Expression in the Liver
2.8. Statistical Analyses
3. Results
3.1. Baseline Parameters
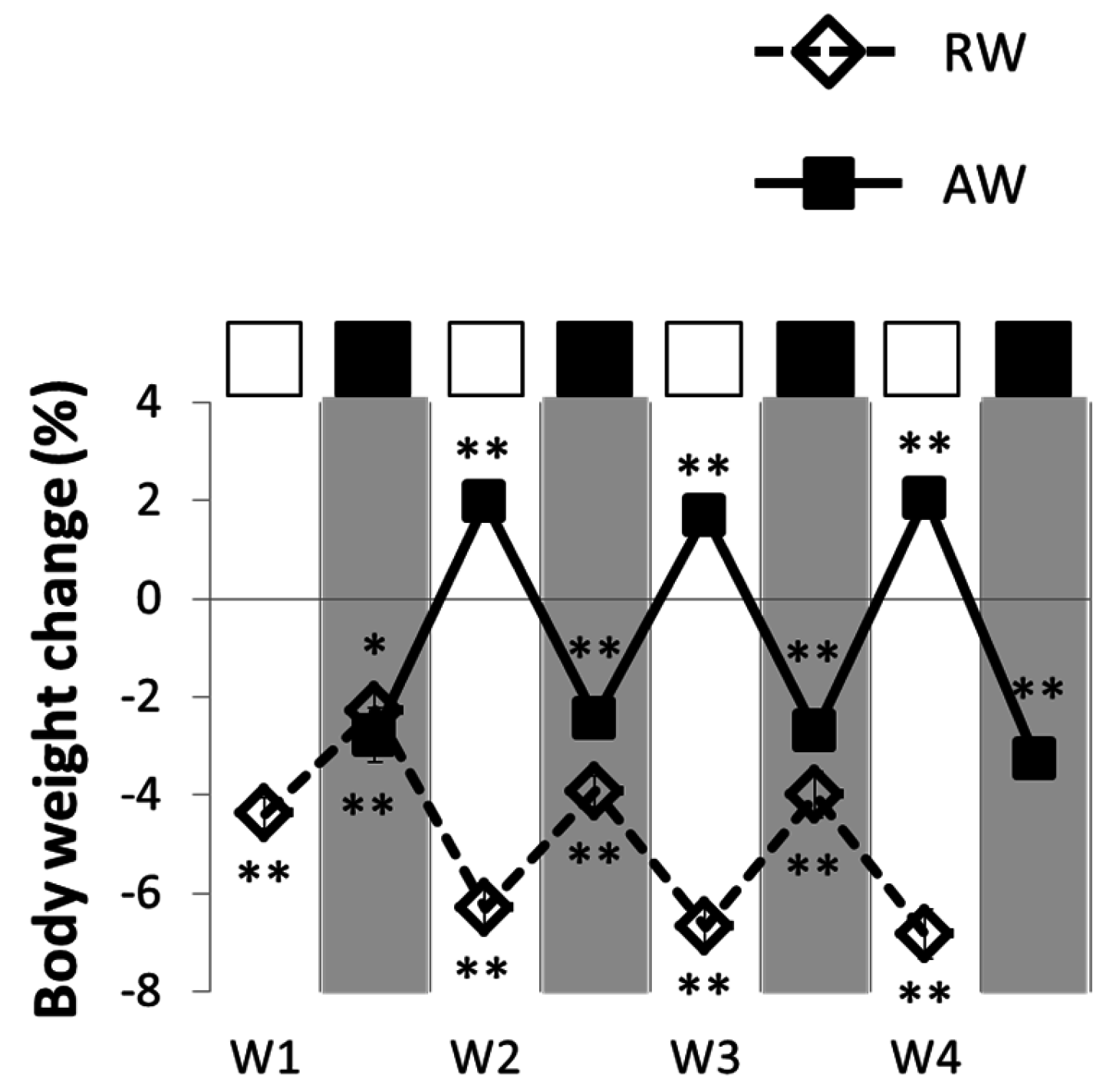
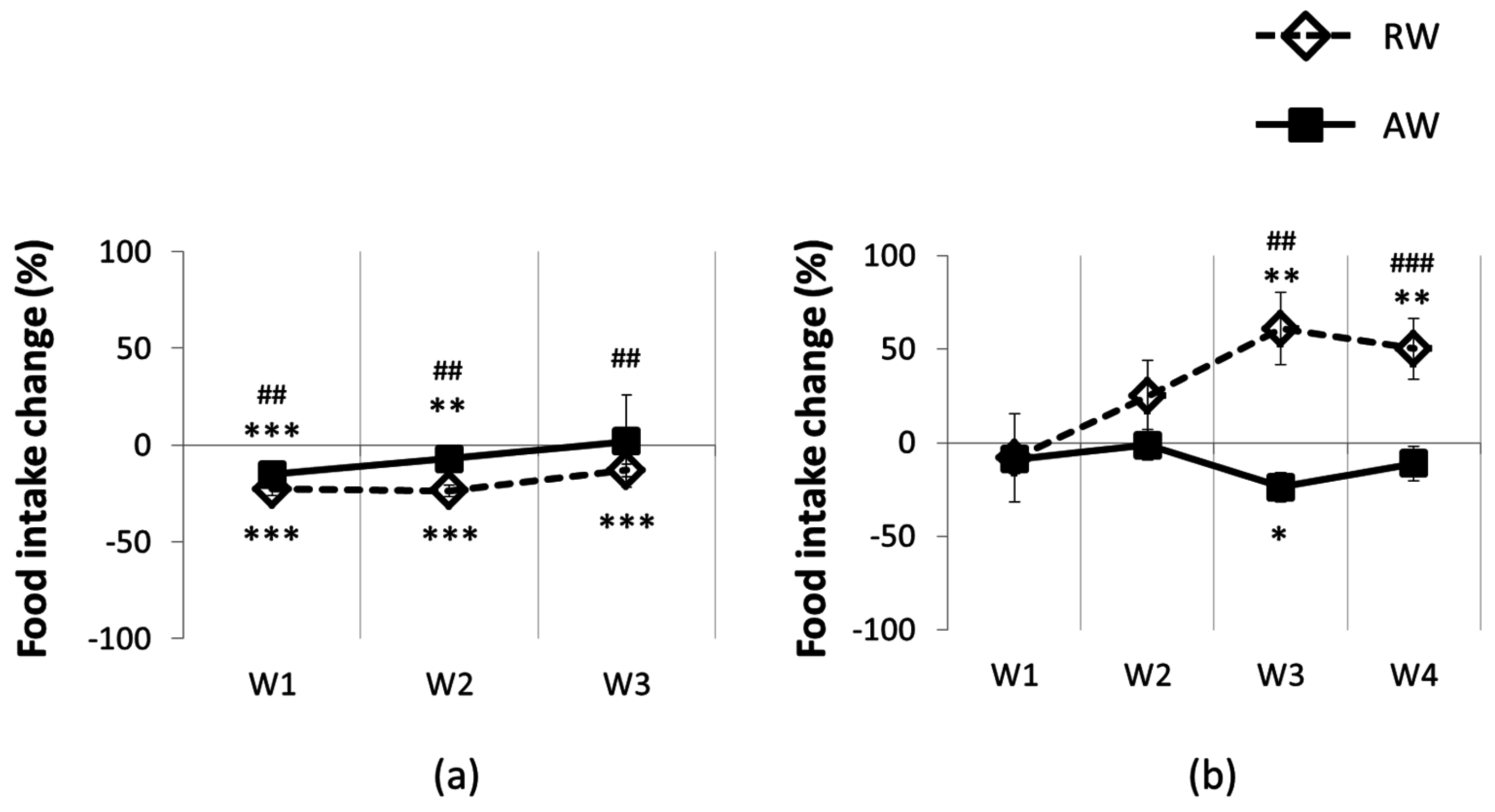
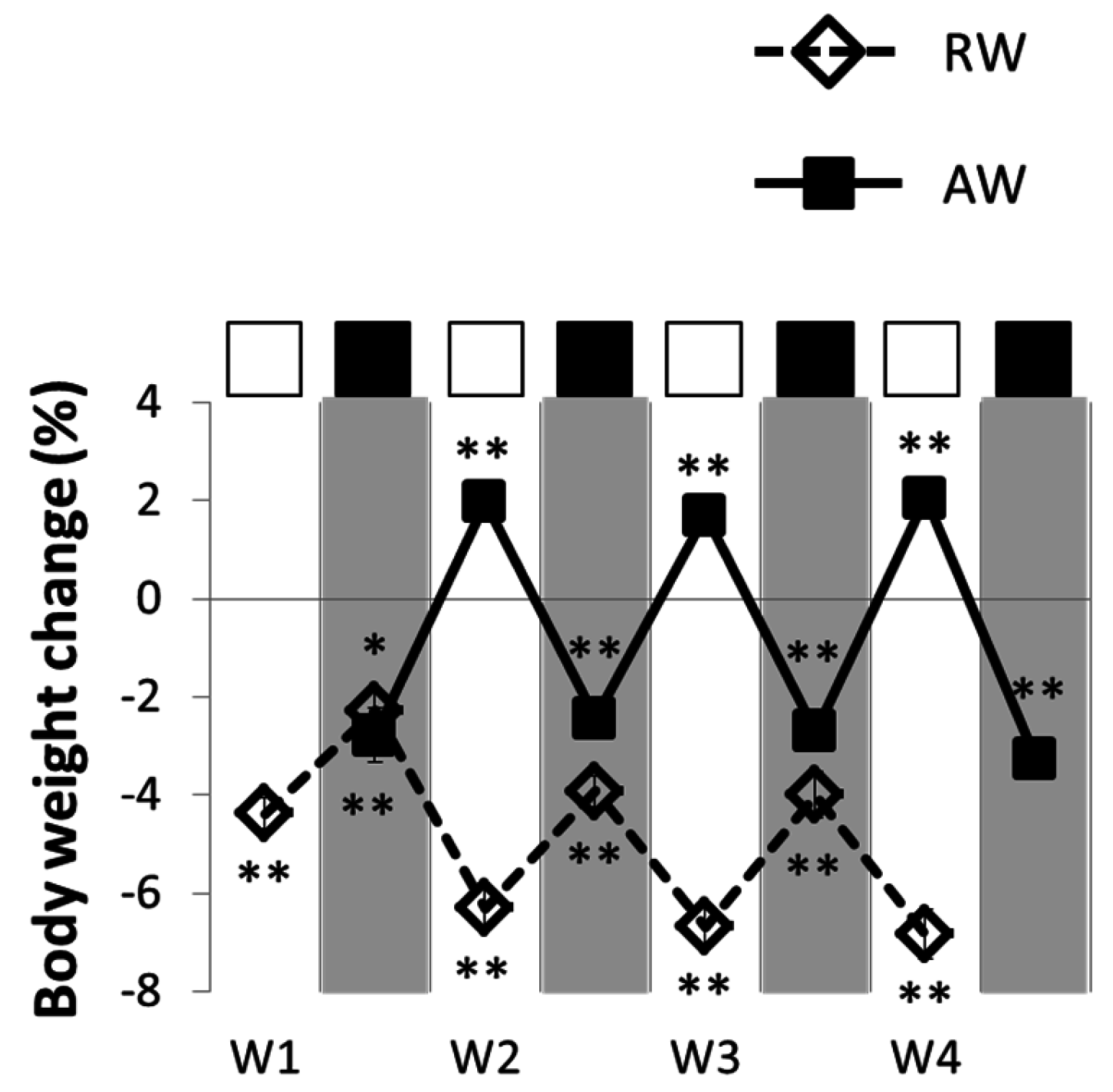
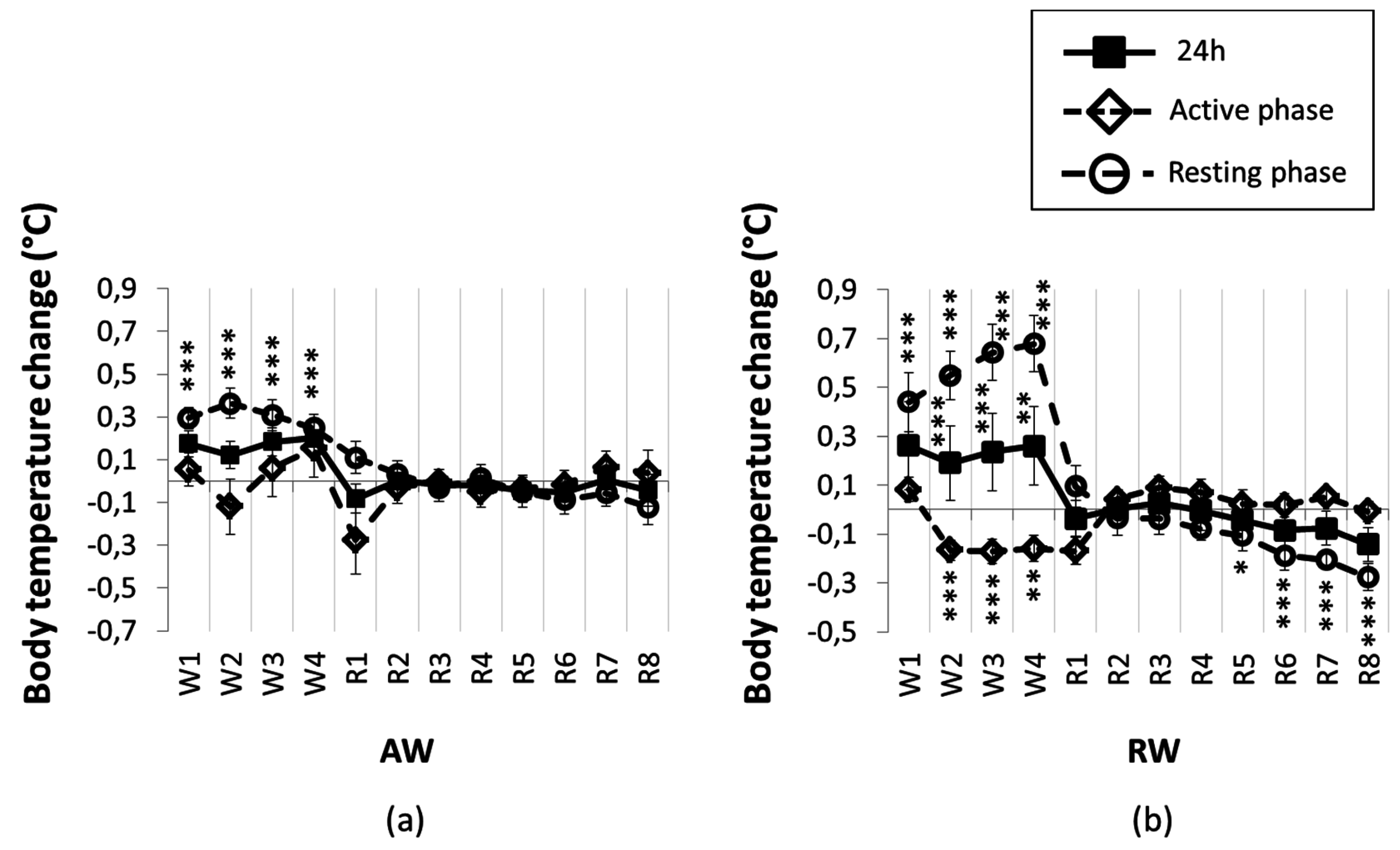
3.2. Feeding and Body Temperature Rhythms, during and after One Rest-Work Period
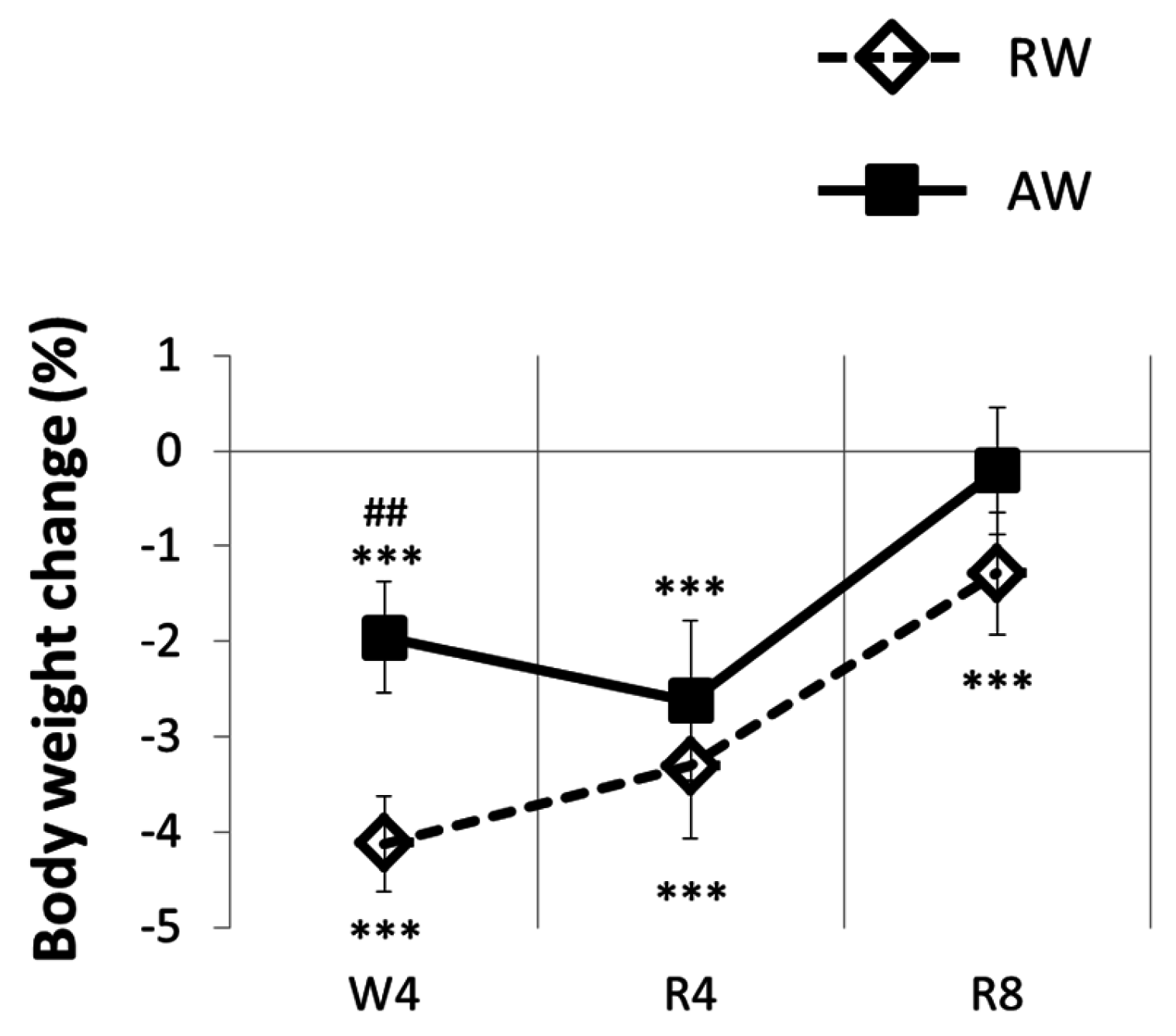
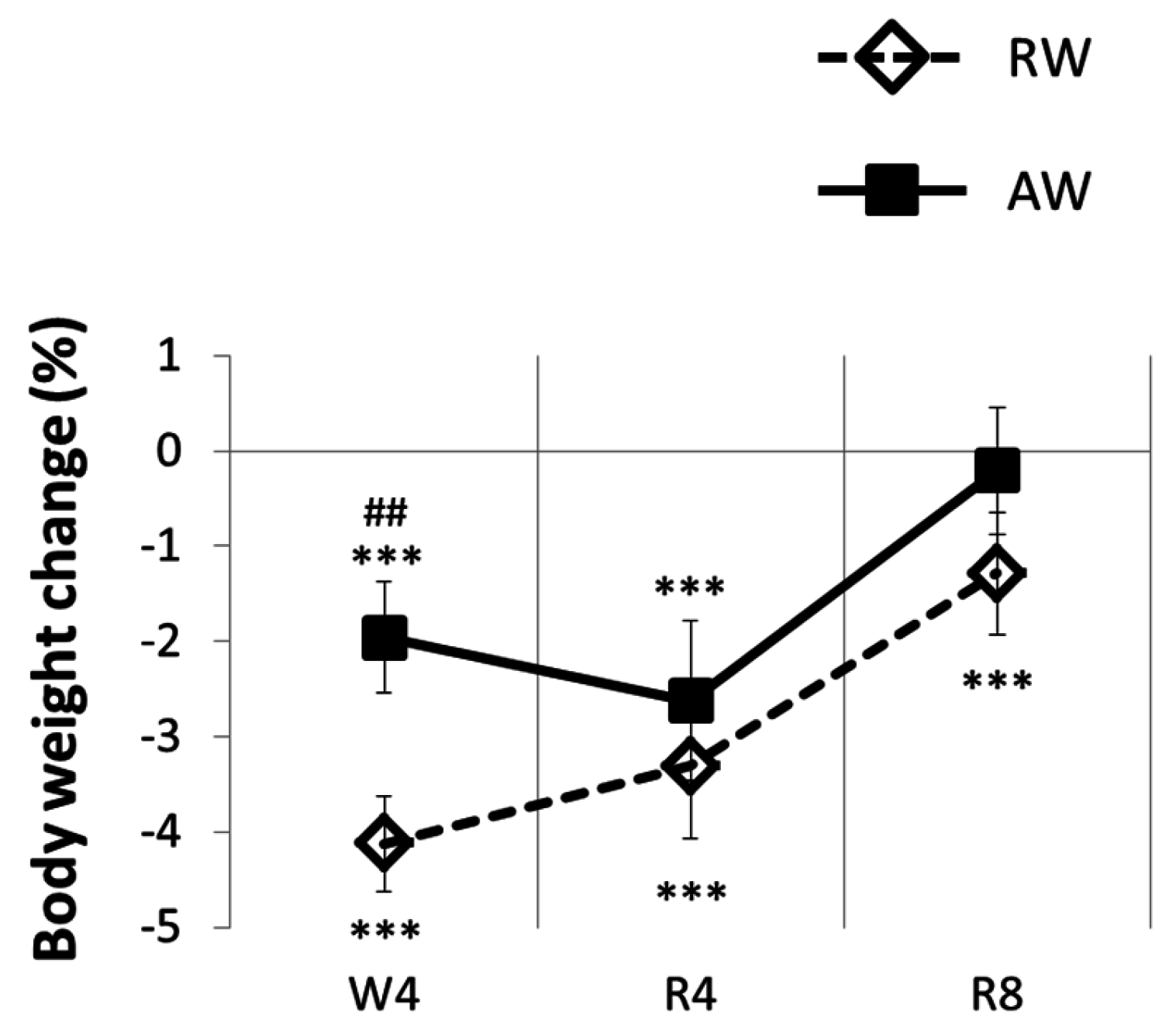
3.3. The Recovery Period
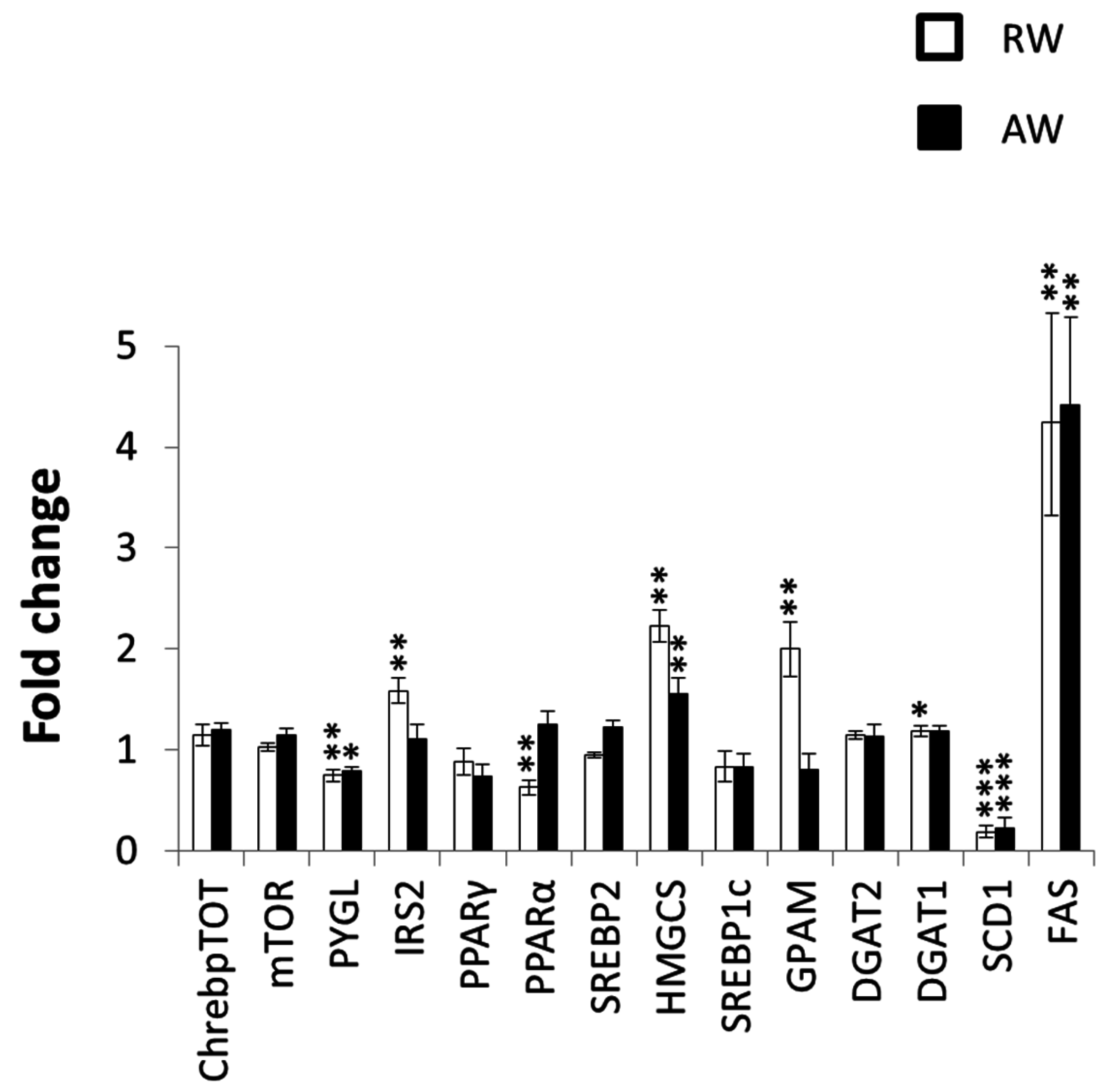
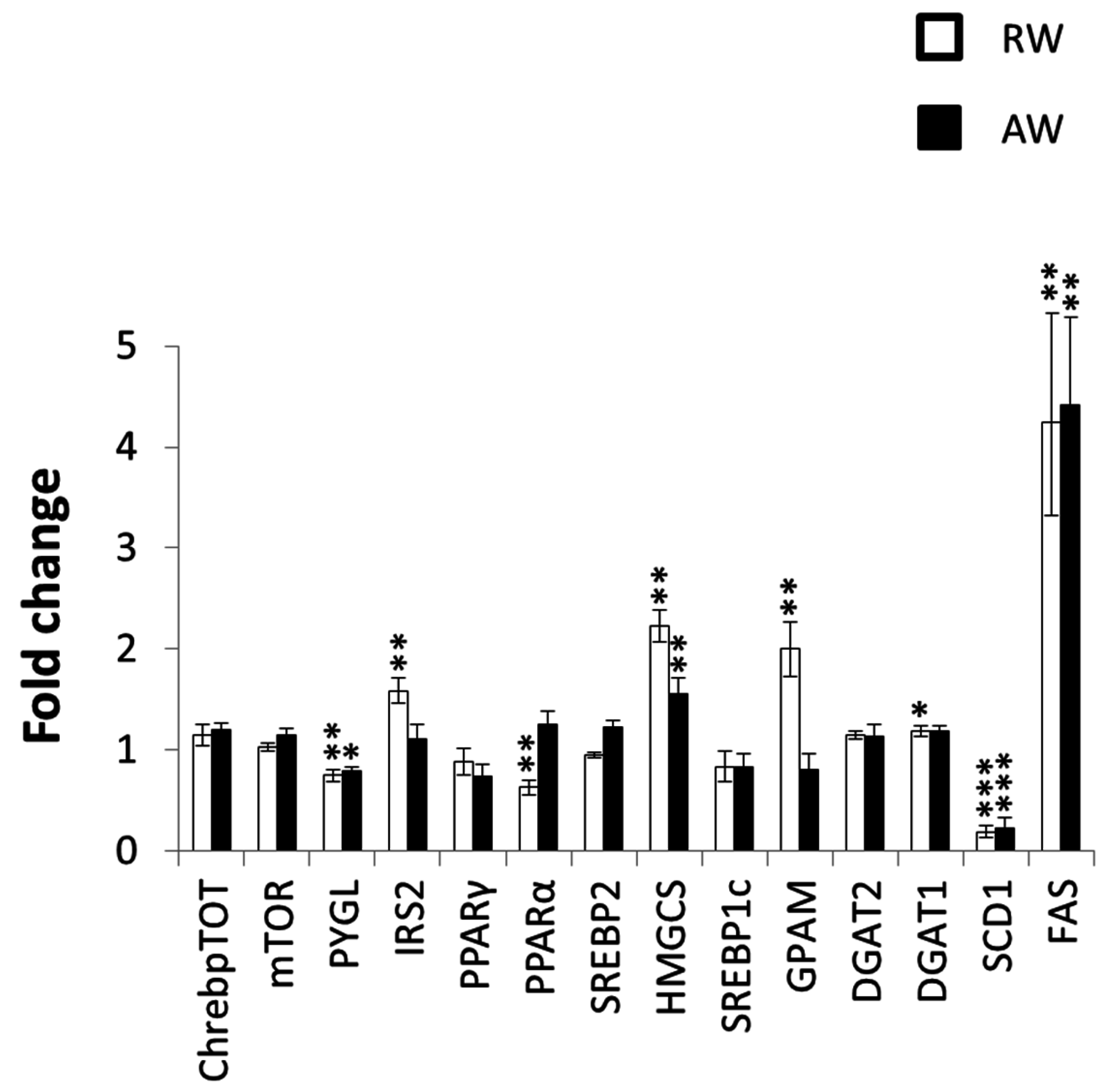
3.4. Metabolic Gene Expression in the Liver at the Shift in Feeding Rhythm of Rest-Workers
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- European Foundation for the Improvement of Living and Working Conditions. Fourth European Working Conditions Survey; Eurofound: Luxembourg, 2007. [Google Scholar]
- Karlsson, B.; Knutsson, A.; Lindahl, B. Is there an association between shift work and having a metabolic syndrome? Results from a population based study of 27 485 people. Occup. Environ. Med. 2001, 58, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Scheer, F.A.; Hilton, M.F.; Mantzoros, C.S.; Shea, S.A. Adverse metabolic and cardiovascular consequences of circadian misalignment. Proc. Natl. Acad. Sci. USA 2009, 106, 4453–4458. [Google Scholar] [CrossRef] [PubMed]
- Knutsson, A. Health disorders of shift workers. Occup. Med. (Lond.) 2003, 53, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Puttonen, S.; Harma, M.; Hublin, C. Shift work and cardiovascular disease—Pathways from circadian stress to morbidity. Scand. J. Work. Environ. Health 2010, 36, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Van Drongelen, A.; Boot, C.R.; Merkus, S.L.; Smid, T.; van der Beek, A.J. The effects of shift work on body weight change—A systematic review of longitudinal studies. Scand. J. Work. Environ. Health 2011, 37, 263–275. [Google Scholar] [CrossRef] [PubMed]
- Gan, Y.; Yang, C.; Tong, X.; Sun, H.; Cong, Y.; Yin, X.; Li, L.; Cao, S.; Dong, X.; Gong, Y.; et al. Shift work and diabetes mellitus: A meta-analysis of observational studies. Occup. Environ. Med. 2015, 72, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Patterson, R.E.; Ang, A.; Emond, J.A.; Shetty, N.; Arab, L. Timing of energy intake during the day is associated with the risk of obesity in adults. J. Hum. Nutr. Diet. 2014, 27 (Suppl. 2), 255–262. [Google Scholar] [CrossRef] [PubMed]
- Hastings, M.; O’Neill, J.S.; Maywood, E.S. Circadian clocks: Regulators of endocrine and metabolic rhythms. J. Endocrinol. 2007, 195, 187–198. [Google Scholar] [CrossRef] [PubMed]
- Dibner, C.; Schibler, U. Circadian timing of metabolism in animal models and humans. J. Intern. Med. 2015, 277, 513–527. [Google Scholar] [CrossRef] [PubMed]
- Konturek, P.C.; Brzozowski, T.; Konturek, S.J. Gut clock: Implication of circadian rhythms in the gastrointestinal tract. J. Physiol. Pharmacol. 2011, 62, 139–150. [Google Scholar] [PubMed]
- Reddy, A.B.; Karp, N.A.; Maywood, E.S.; Sage, E.A.; Deery, M.; O’Neill, J.S.; Wong, G.K.; Chesham, J.; Odell, M.; Lilley, K.S.; et al. Circadian orchestration of the hepatic proteome. Curr. Biol. 2006, 16, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, R.A.; Reddy, A.B.; Maywood, E.S.; Clayton, J.D.; King, V.M.; Smith, A.G.; Gant, T.W.; Hastings, M.H.; Kyriacou, C.P. Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr. Biol. 2002, 12, 540–550. [Google Scholar] [CrossRef]
- Sadacca, L.A.; Lamia, K.A.; deLemos, A.S.; Blum, B.; Weitz, C.J. An intrinsic circadian clock of the pancreas is required for normal insulin release and glucose homeostasis in mice. Diabetologia 2011, 54, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Damiola, F.; Le Minh, N.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [PubMed]
- Eastman, C.I.; Stewart, K.T.; Mahoney, M.P.; Liu, L.; Fogg, L.F. Dark goggles and bright light improve circadian rhythm adaptation to night-shift work. Sleep 1994, 17, 535–543. [Google Scholar] [PubMed]
- Crowley, S.J.; Eastman, C.I. Phase advancing human circadian rhythms with morning bright light, afternoon melatonin, and gradually shifted sleep: Can we reduce morning bright-light duration? Sleep Med. 2015, 16, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Wyse, C.A.; Biello, S.M.; Gill, J.M. The bright-nights and dim-days of the urban photoperiod: Implications for circadian rhythmicity, metabolism and obesity. Ann. Med. 2014, 46, 253–263. [Google Scholar] [CrossRef] [PubMed]
- Folkard, S. Do permanent night workers show circadian adjustment? A review based on the endogenous melatonin rhythm. Chronobiol. Int. 2008, 25, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, D.C.; Hampton, S.M.; Morgan, L.; Deacon, S.; Arendt, J. Altered postprandial hormone and metabolic responses in a simulated shift work environment. J. Endocrinol. 1998, 158, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Opperhuizen, A.L.; van Kerkhof, L.W.; Proper, K.I.; Rodenburg, W.; Kalsbeek, A. Rodent models to study the metabolic effects of shiftwork in humans. Front. Pharmacol. 2015, 6, 50. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P. The laboratory rat: Relating its age with human’s. Int. J. Prev. Med. 2013, 4, 624–630. [Google Scholar] [PubMed]
- Gronli, J.; Meerlo, P.; Pedersen, T.; Pallesen, S.; Skrede, S.; Marti, A.R.; Wisor, J.; Murison, R.; Henriksen, T.E.; Rempe, M.; et al. A rodent model of night-shift work induces short-term and enduring sleep and electroencephalographic disturbances. J. Biol. Rhythms 2016, in press. [Google Scholar]
- Mrdalj, J.; Pallesen, S.; Milde, A.M.; Jellestad, F.K.; Murison, R.; Ursin, R.; Bjorvatn, B.; Gronli, J. Early and later life stress alter brain activity and sleep in rats. PLoS ONE 2013, 8, e69923. [Google Scholar] [CrossRef] [PubMed]
- Zuther, P.; Gorbey, S.; Lemmer, B. Chronos-Fit 1.05. Available online: http://www.ma.uni-heidelberg.de/inst/phar/lehre/chrono.html (accessed on 1 June 2009).
- Bonham, M.P.; Bonnell, E.K.; Huggins, C.E. Energy intake of shift workers compared to fixed day workers: A systematic review and meta-analysis. Chronobiol. Int. 2016, 33, 1086–1100. [Google Scholar] [CrossRef] [PubMed]
- Lowden, A.; Moreno, C.; Holmback, U.; Lennernas, M.; Tucker, P. Eating and shift work—Effects on habits, metabolism and performance. Scand. J. Work. Environ. Health 2010, 36, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Delgado, R.C.; Saderi, N.; Basualdo Mdel, C.; Guerrero-Vargas, N.N.; Escobar, C.; Buijs, R.M. Shift work or food intake during the rest phase promotes metabolic disruption and desynchrony of liver genes in male rats. PLoS ONE 2013, 8, e60052. [Google Scholar] [CrossRef] [PubMed]
- Awazawa, M.; Ueki, K.; Inabe, K.; Yamauchi, T.; Kubota, N.; Kaneko, K.; Kobayashi, M.; Iwane, A.; Sasako, T.; Okazaki, Y.; et al. Adiponectin enhances insulin sensitivity by increasing hepatic IRS-2 expression via a macrophage-derived IL-6-dependent pathway. Cell Metab. 2011, 13, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, C.M.; Ueki, K.; Kahn, R. Complementary roles of IRS-1 and IRS-2 in the hepatic regulation of metabolism. J. Clin. Investig. 2005, 115, 718–727. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Park, S.; Lin, X.; Copps, K.; Yi, X.; White, M.F. Irs1 and Irs2 signaling is essential for hepatic glucose homeostasis and systemic growth. J. Clin. Investig. 2006, 116, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Urstad, A.P.; Semenkovich, C.F. Fatty acid synthase and liver triglyceride metabolism: Housekeeper or messenger? Biochim. Biophys. Acta 2012, 1821, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Murphy, H.M.; Wideman, C.H.; Nadzam, G.R. A laboratory animal model of human shift work. Integr. Physiol. Behav. Sci. 2003, 38, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Leenaars, C.H.; Kalsbeek, A.; Hanegraaf, M.A.; Foppen, E.; Joosten, R.N.; Post, G.; Dematteis, M.; Feenstra, M.G.; van Someren, E.J. Unaltered instrumental learning and attenuated body-weight gain in rats during non-rotating simulated shiftwork. Chronobiol. Int. 2012, 29, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Barf, R.P.; Desprez, T.; Meerlo, P.; Scheurink, A.J. Increased food intake and changes in metabolic hormones in response to chronic sleep restriction alternated with short periods of sleep allowance. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R112–R117. [Google Scholar] [CrossRef] [PubMed]
- Salgado-Delgado, R.; Angeles-Castellanos, M.; Buijs, M.R.; Escobar, C. Internal desynchronization in a model of night-work by forced activity in rats. Neuroscience 2008, 154, 922–931. [Google Scholar] [CrossRef] [PubMed]
- De Assis, M.A.; Kupek, E.; Nahas, M.V.; Bellisle, F. Food intake and circadian rhythms in shift workers with a high workload. Appetite 2003, 40, 175–183. [Google Scholar] [CrossRef]
- McHill, A.W.; Melanson, E.L.; Higgins, J.; Connick, E.; Moehlman, T.M.; Stothard, E.R.; Wright, K.P., Jr. Impact of circadian misalignment on energy metabolism during simulated nightshift work. Proc. Natl. Acad. Sci. USA 2014, 111, 17302–17307. [Google Scholar] [CrossRef] [PubMed]
- Rodnick, K.J.; Reaven, G.M.; Haskell, W.L.; Sims, C.R.; Mondon, C.E. Variations in running activity and enzymatic adaptations in voluntary running rats. J. Appl. Physiol. 1989, 66, 1250–1257. [Google Scholar] [PubMed]
- Siyamak, A.Y.; Macdonald, I.A. Sub-acute underfeeding but not acute starvation reduces noradrenaline induced thermogenesis in the conscious rat. Int. J. Obes. Relat. Metab. Disord. 1992, 16, 113–117. [Google Scholar] [PubMed]
- Mrdalj, J.; Lundegaard Mattson, A.; Murison, R.; Konow Jellestad, F.; Milde, A.M.; Pallesen, S.; Ursin, R.; Bjorvatn, B.; Gronli, J. Hypothermia after chronic mild stress exposure in rats with a history of postnatal maternal separations. Chronobiol. Int. 2014, 31, 252–264. [Google Scholar] [CrossRef] [PubMed]
- Harris, R.B.; Zhou, J.; Youngblood, B.D.; Rybkin, I.I.; Smagin, G.N.; Ryan, D.H. Effect of repeated stress on body weight and body composition of rats fed low- and high-fat diets. Am. J. Physiol. 1998, 275 6 Pt 2, R1928–R1938. [Google Scholar] [PubMed]
- Shibli-Rahhal, A.; Van Beek, M.; Schlechte, J.A. Cushing’s syndrome. Clin. Dermatol. 2006, 24, 260–265. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Main Function | Accession Number | Forward Primer | Reverse Primer |
|---|---|---|---|---|
| Fatty acid synthase (Fasn) | Fatty acid synthesis | NM_017332 | CCATCATCCCCTTGATGAAGA | GTTGATGTCGATGCCTGTGAG |
| Stearoyl-CoA 9-desaturase; Stearoyl-CoA desaturase 1 (Scd1) | Fatty acid desaturation | NM_139192 | TCAATCTCGGGAGAACATCC | CATGCAGTCGATGAAGAACG |
| Diacylglycerol O-acyltransferase 1 (Dgat1) | Triglyceride synthesis | NM_053437 | AATGCTGCGGAAAAACTACG | TTGCTGGTAACAGTGCTTGC |
| Diacylglycerol O-acyltransferase 2 (Dgat2) | Triglyceride synthesis | NM_001012345 | AATCTGTGGTGCCGCCAG | TCCCTGCAGACACAGCTTTG |
| Glycerol-3-phosphate acyltransferase 1, mitochondrial (Gpam) | Triglyceride synthesis | NM_017274 | AATGCTGCGGAAAAACTACG | TTGCTGGTAACAGTGCTTGC |
| Sterol regulatory element-binding protein 1c (Srebf1) | Key regulator of fatty acid/triglyceride synthesis | NM_001271207 | GAACCGCAAAGGCTTTGTAAA | ACCCAGATCAGCTCCATGGC |
| Hydroxymethylglutaryl-CoA synthase, cytoplasmic (Hmgcs1) | Sterol synthesis | NM_017268 | CAGCTCTTGGGATGGACGA | GGCGTTTCCTGAGGCATATATAG |
| Sterol regulatory element-binding protein 2 (Srebf2) | Key regulator of sterol synthesis | NM_001033694.1 | GCCGCAACCAGCTTTCAA | CCTGCTGCACCTGTGGTGTA |
| Peroxisome proliferator-activated receptor alpha (Ppara) | Fatty acid oxidation | NM_013196 | AATGCAATCCGTTTTGGAAGA | ACAGGTAAGGATTTCTGCCTTCAG |
| Peroxisome proliferator-activated receptor gamma (Pparg) | Adipocyte differentiation | NM_001145366 | CCACAAAAAGAGTAGAAATAAATGTCAGTAC | CAAACCTGATGGCATTGTGAGA |
| Insulin receptor substrate 2 (Irs2) | Mediation of insulin effects | NM_001168633 | GAAGCGGCTAAGTCTCATGG | CTGGCTGACTTGAAGGAAGG |
| Phosphorylase, glycogen, liver (Pygl) | Glycogen breakdown | NM_022268 | AAAAGCCTGGAACACAATGG | TCGGTCACTGGAGAACTTCC |
| Mechanistic target of rapamycin (Mtor) | Mediation of cellular metabolic stress response | NM_019906 | CATGAGATGTGGCATGAAGG | AAACATGCCTTTGACGTTCC |
| Carbohydrate responsive element binding protein (ChREBP/Mlxipl) | Triglyceride synthesis in response to carbohydrates | AB074517 | CTCTCAGGGAATACACGTCTCC | ATCTTGGTCTTTGGGTCTTCAGG |
| β-actin (Actb) | Endogenous control | NM_031144 | TACAGCTTCACCACCACAGC | CTTCTCCAGGGAGGAAGAGG |
| Rattus norvegicus ribosomal protein lateral stalk subunit P0 (Rplp0) | Endogenous control | NM_022402.2 | CATTGAAATCCTGAGCGATGT | AGATGTTCAACATGTTCAGCAG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marti, A.R.; Meerlo, P.; Grønli, J.; Van Hasselt, S.J.; Mrdalj, J.; Pallesen, S.; Pedersen, T.T.; Henriksen, T.E.G.; Skrede, S. Shift in Food Intake and Changes in Metabolic Regulation and Gene Expression during Simulated Night-Shift Work: A Rat Model. Nutrients 2016, 8, 712. https://doi.org/10.3390/nu8110712
Marti AR, Meerlo P, Grønli J, Van Hasselt SJ, Mrdalj J, Pallesen S, Pedersen TT, Henriksen TEG, Skrede S. Shift in Food Intake and Changes in Metabolic Regulation and Gene Expression during Simulated Night-Shift Work: A Rat Model. Nutrients. 2016; 8(11):712. https://doi.org/10.3390/nu8110712
Chicago/Turabian StyleMarti, Andrea Rørvik, Peter Meerlo, Janne Grønli, Sjoerd Johan Van Hasselt, Jelena Mrdalj, Ståle Pallesen, Torhild Thue Pedersen, Tone Elise Gjøtterud Henriksen, and Silje Skrede. 2016. "Shift in Food Intake and Changes in Metabolic Regulation and Gene Expression during Simulated Night-Shift Work: A Rat Model" Nutrients 8, no. 11: 712. https://doi.org/10.3390/nu8110712
APA StyleMarti, A. R., Meerlo, P., Grønli, J., Van Hasselt, S. J., Mrdalj, J., Pallesen, S., Pedersen, T. T., Henriksen, T. E. G., & Skrede, S. (2016). Shift in Food Intake and Changes in Metabolic Regulation and Gene Expression during Simulated Night-Shift Work: A Rat Model. Nutrients, 8(11), 712. https://doi.org/10.3390/nu8110712