
Entirely Carbohydrate-Based Vaccines: An Emerging Field for Specific and Selective Immune Responses


Abstract
:
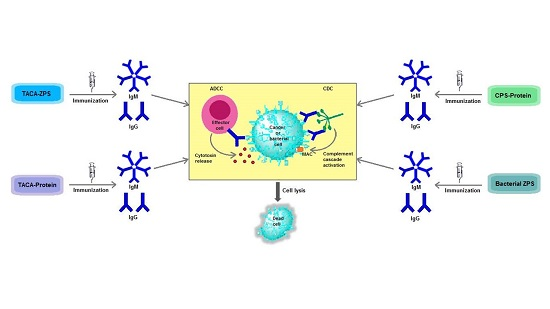{kind=link}
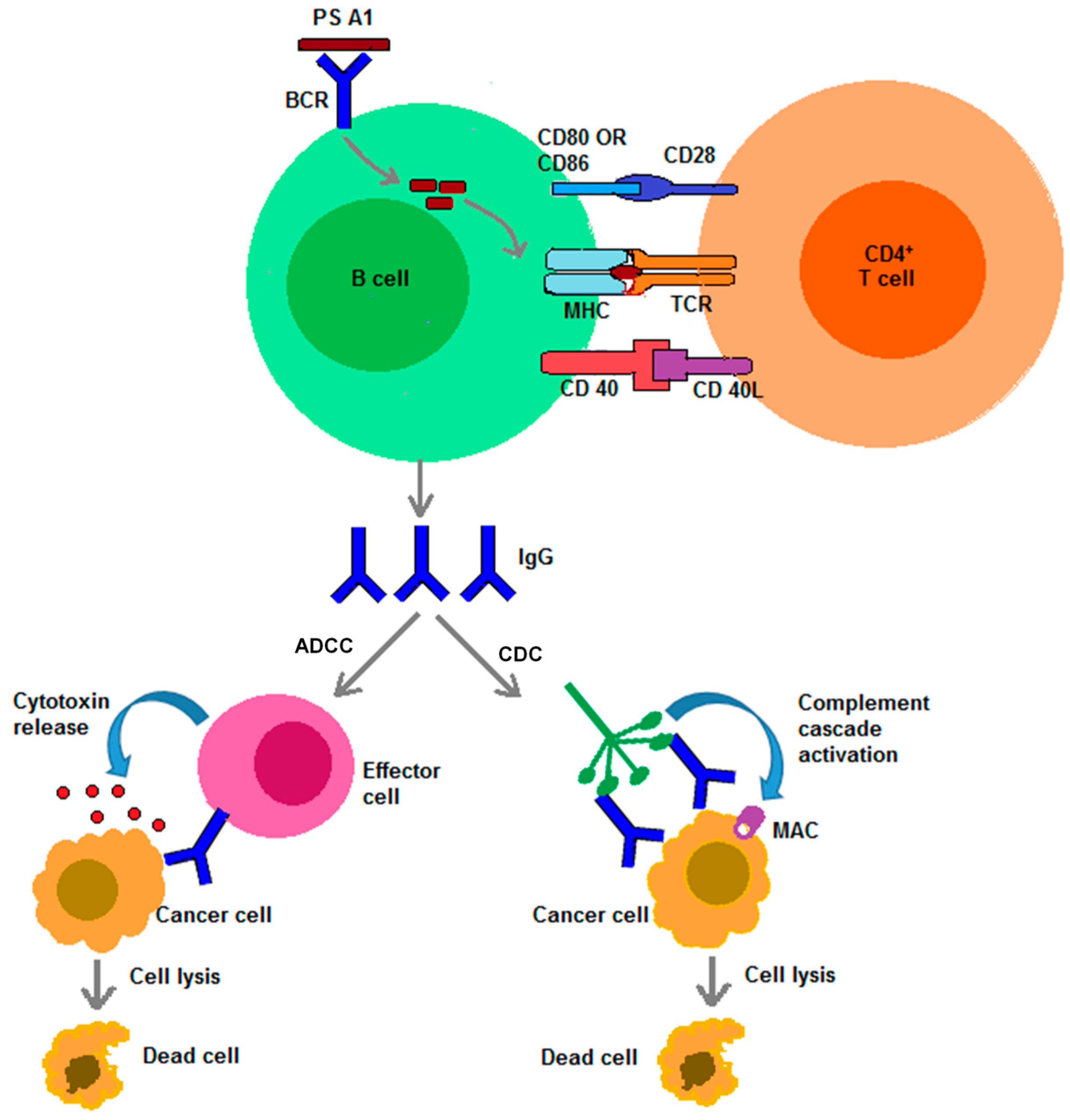{kind=link}
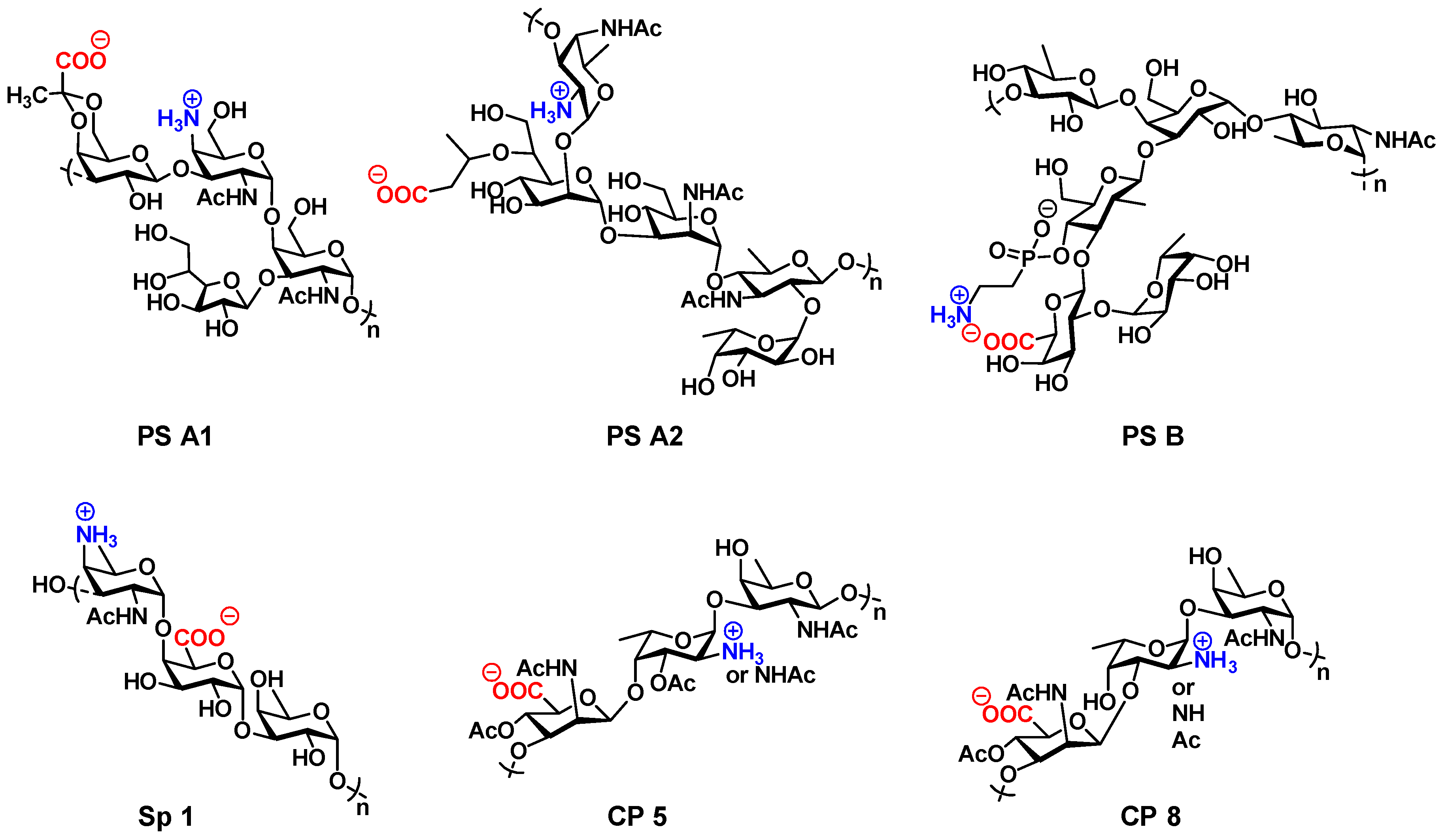{kind=link}
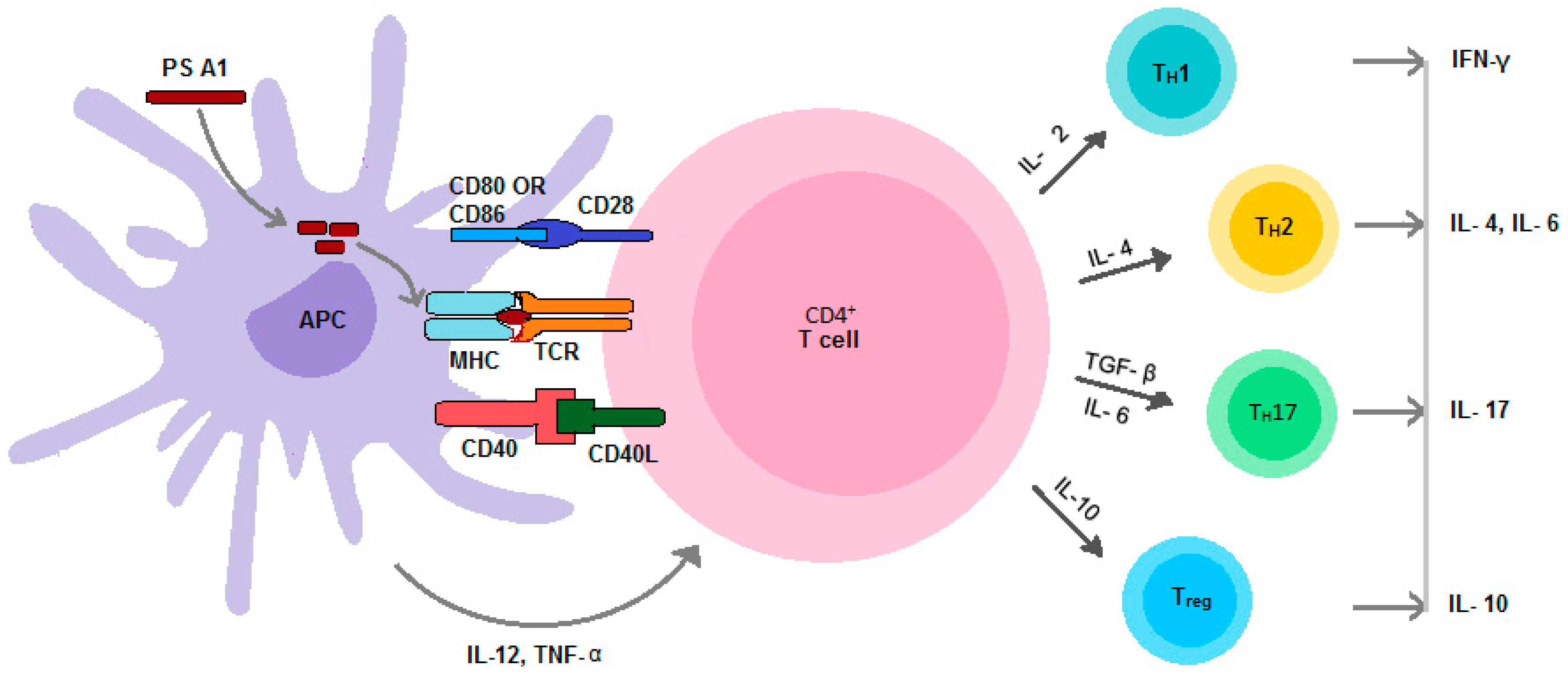{kind=link}
1. Introduction
2. T-cell Independent Immune Responses
3. Entirely Carbohydrate Vaccine Conjugates
3.1. Vi Polysaccharide Vaccine
3.2. Neisseria Meningitides Polysaccharide Vaccine
3.3. Tumor Associated Carbohydrate Vaccine
4. Zwitterionic Polysaccharides: A Means to T-cell Activation
4.1. Zwitterionic Capsular Polysaccharides from B. fragilis (PS A1, PS B, PS A2)
4.2. Zwitterionic Capsular Polysaccharide Sp 1 from S. pneumoniae
4.3. Zwitterionic CP5 and CP8 from S. aureus
5. Zwitterionic Polysaccharides in the Development of Cancer Vaccines
5.1. PS A1
5.2. PS B
6. Zwitterionic Polysaccharides in Bacterial Vaccines
6.1. Sp 1
6.2. CP5 and CP8
7. Conclusions
Acknowledgments
Conflicts of Interest
References
- Coley, W.B. The treatment of malignant tumors by repeated inoculations of erysipelas: With a report of ten original cases. Am. J. Med. Sci. 1893, 105, 487–510. [Google Scholar] [CrossRef]
- Mazmanian, S.K.; Kasper, D.L. The love-hate relationship between bacterial polysaccharides and the host immune system. Nat. Rev. Immunol. 2006, 6, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Dziadek, S.; Hobel, A.; Schmitt, E.; Kunz, H. A fully synthetic vaccine consisting of a tumor-associated glycopeptide antigen and a T-cell epitope for the induction of a highly specific humoral immune response. Angew. Chem. Int. Ed. Engl. 2005, 44, 7630–7635. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Synthetic glycopeptide vaccines combining β-mannan and peptide epitopes induce protection against candidiasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13526–13531. [Google Scholar] [CrossRef] [PubMed]
- Astronomo, R.D.; Burton, D.R. Carbohydrate vaccines: Developing sweet solutions to sticky situations? Nat. Rev. Drug. Discov. 2010, 9, 308–324. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Aryal, R.P.; Stowell, C.J.; Cummings, R.D. Regulation of protein O-glycosylation by the endoplasmic reticulum–localized molecular chaperone Cosmc. J. Cell Biol. 2008, 182, 531–542. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Lanneau, G.S.; Gautam, T.; Wang, Y.; Xia, B.; Stowell, S.R.; Willard, M.T.; Wang, W.; Xia, J.Y.; Zuna, R.E.; et al. Human tumor antigens Tn and sialyl Tn arise from mutations in Cosmc. Cancer Res. 2008, 68, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Adams, E.W.; Ratner, D.M.; Seeberger, P.H.; Hacohen, N. Carbohydrate-mediated targeting of antigen to dendritic cells leads to enhanced presentation of antigen to T cells. ChemBioChem 2008, 9, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Duan, J.; Avci, F.Y.; Kasper, D.L. Microbial carbohydrate depolymerization by antigen-presenting cells: Deamination prior to presentation by the MHCII pathway. Proc. Natl. Acad. Sci. USA 2008, 105, 5183–5188. [Google Scholar] [CrossRef] [PubMed]
- Jacques, I.; Olivier-Bernardin, V.; Dubray, G. Induction of antibody and protective responses in mice by Brucella O-polysaccharide-BSA conjugate. Vaccine 1991, 9, 896–900. [Google Scholar] [CrossRef]
- Ibrahim, N.K.; Murray, J.L. Clinical development of the STn-KLH vaccine (Theratope). Clin. Breast Cancer 2003, 3, 139–143. [Google Scholar] [CrossRef]
- Finn, A. Bacterial polysaccharide–protein conjugate vaccines. Br. Med. Bull. 2004, 70, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Eskola, J.; Kayhty, H.; Takala, A.K.; Peltola, H.; Ronnberg, P.R.; Kela, E.; Pekkanen, E.; McVerry, P.H.; Makela, P.H. A randomized, prospective field trial of a conjugate vaccine in the protection of infants and young children against invasive Haemophilus influenzae type B disease. N. Engl. J. Med. 1990, 323, 1381–1387. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, F.B.; Poutrel, B.; Sutra, L. Immunogenicity in cows of Staphylococcus aureus type 5 capsular polysaccharide—Ovalbumin conjugate. Vaccine 1994, 12, 369–374. [Google Scholar] [CrossRef]
- Pozsgay, V.; Chu, C.; Pannell, L.; Wolfe, J.; Robbins, J.B.; Schneerson, R. Protein conjugates of synthetic saccharides elicit higher levels of serum IgG lipopolysaccharide antibodies in mice than do those of the O-specific polysaccharide from Shigella dysenteriae type 1. Proc. Natl. Acad. Sci. USA 1999, 96, 5194–5197. [Google Scholar] [CrossRef] [PubMed]
- Fattom, A.I.; Horwith, G.; Fuller, S.; Propst, M.; Naso, R. Development of StaphVAX, a polysaccharide conjugate vaccine against S. aureus infection: From the lab bench to phase III clinical trials. Vaccine 2004, 22, 880–887. [Google Scholar] [CrossRef] [PubMed]
- Kaltgrad, E.; Sen Gupta, S.; Punna, S.; Huang, C.Y.; Chang, A.; Wong, C.H.; Finn, M.G.; Blixt, O. Anti-carbohydrate antibodies elicited by polyvalent display on a viral scaffold. ChemBioChem 2007, 8, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Herzenberg, L.A.; Tokuhisa, T.; Herzenberg, L.A. Carrier-priming leads to hapten-specific suppression. Nature 1980, 285, 664–667. [Google Scholar] [CrossRef] [PubMed]
- Rossjohn, J.; Pellicci, D.G.; Patel, O.; Gapin, L.; Godfrey, D.I. Recognition of CD1d-restricted antigens by natural killer T cells. Nat. Rev. Immunol. 2012, 12, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Tzianabos, A.O.; Onderdonk, A.B.; Rosner, B.; Cisneros, R.L.; Kasper, D.L. Structural features of polysaccharides that induce intra-abdominal abscesses. Science 1993, 262, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Gibson, F.C.; Onderdonk, A.B.; Kasper, D.L.; Tzianabos, A.O. Cellular mechanism of intraabdominal abscess formation by Bacteroides fragilis. J. Immunol. 1998, 160, 5000–5006. [Google Scholar] [PubMed]
- Natsume, A.; Niwa, R.; Satoh, M. Improving effector functions of antibodies for cancer treatment: Enhancing ADCC and CDC. Drug Des. Devel. Ther. 2009, 3, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Rakoff-Nahoum, S.; Paglino, J.; Eslami-Varzaneh, F.; Edberg, S.; Medzhitov, R. Recognition of commensal microflora by toll-like receptors is required for intestinal homeostasis. Cell 2004, 118, 229–241. [Google Scholar] [CrossRef] [PubMed]
- Pardoll, D.M.; Fowlkes, B.J.; Lew, A.M.; Maloy, W.L.; Weston, M.A.; Bluestone, J.A.; Schwartz, R.H.; Coligan, J.E.; Kruisbeek, A.M. Thymus-dependent and thymus-independent developmental pathways for peripheral T cell receptor-gamma delta-bearing lymphocytes. J. Immunol. 1988, 140, 4091–4096. [Google Scholar] [PubMed]
- Wang, C.L.; Lu, C.Y.; Pi, C.C.; Zhuang, Y.J.; Chu, C.L.; Liu, W.H.; Chen, C.J. Extracellular polysaccharides produced by Ganoderma formosanum stimulate macrophage activation via multiple pattern-recognition receptors. BMC Complement. Altern. Med. 2012, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Avery, O.T.; Goebel, W.F. Chemo-immunological studies on conjugated carbohydrate-proteins: V. The immunological specifity of an antigen prepared by combining the capsular polysaccharide of type III pneumococcus with foreign protein. J. Exp. Med. 1931, 54, 437–447. [Google Scholar] [CrossRef] [PubMed]
- Beuvery, E.C.; van Rossum, F.; Nagel, J. Comparison of the induction of immunoglobulin M and G antibodies in mice with purified pneumococcal type 3 and meningococcal group C polysaccharides and their protein conjugates. Infect. Immun. 1982, 37, 15–22. [Google Scholar] [PubMed]
- Schneerson, R.; Barrera, O.; Sutton, A.; Robbins, J.B. Preparation, characterization, and immunogenicity of Haemophilus influenzae type B polysaccharide-protein conjugates. J. Exp. Med. 1980, 152, 361–376. [Google Scholar] [CrossRef] [PubMed]
- Wessels, M.R.; Paoletti, L.C.; Rodewald, A.K.; Michon, F.; DiFabio, J.; Jennings, H.J.; Kasper, D.L. Stimulation of protective antibodies against type Ia and Ib group B streptococci by a type Ia polysaccharide-tetanus toxoid conjugate vaccine. Infect. Immun. 1993, 61, 4760–4766. [Google Scholar] [PubMed]
- Heidelberger, M.; Avery, O.T. The soluble specific substance of pneumococcus. J. Exp. Med. 1923, 38, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Keitel, W.A.; Bond, N.L.; Zahradnik, J.M.; Cramton, T.A.; Robbins, J.B. Clinical and serological responses following primary and booster immunization with Salmonella typhi Vi capsular polysaccharide vaccines. Vaccine 1994, 12, 195–199. [Google Scholar] [CrossRef]
- Klugman, K.P.; Gilbertson, I.T.; Koornhof, H.J.; Robbins, J.B.; Schneerson, R.; Schulz, D.; Cadoz, M.; Armand, J. Protective activity of Vi capsular polysaccharide vaccine against typhoid fever. Lancet 1987, 2, 1165–1169. [Google Scholar] [CrossRef]
- King, W.J.; MacDonald, N.E.; Wells, G.; Huang, J.; Allen, U.; Chan, F.; Ferris, W.; Diaz-Mitoma, F.; Ashton, F. Total and functional antibody response to a quadrivalent meningococcal polysaccharide vaccine among children. J. Pediatr. 1996, 128, 196–202. [Google Scholar] [CrossRef]
- Felix, A.; Margaret Pitt, R. A new antigen of B. typhosus: Its relation to virulence and to active and passive immunisation. Lancet 1934, 224, 186–191. [Google Scholar] [CrossRef]
- Garg, R.; Qadri, A. Hemoglobin transforms anti-inflammatory Salmonella typhi virulence polysaccharide into a TLR-2 agonist. J. Immunol. 2010, 184, 5980–5987. [Google Scholar] [CrossRef] [PubMed]
- Schreibelt, G.; Benitez-Ribas, D.; Schuurhuis, D.; Lambeck, A.J.; van Hout-Kuijer, M.; Schaft, N.; Punt, C.J.; Figdor, C.G.; Adema, G.J.; de Vries, I.J. Commonly used prophylactic vaccines as an alternative for synthetically produced TLR ligands to mature monocyte-derived dendritic cells. Blood 2010, 116, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.L.; Flores-Langarica, A.; Kingsley, R.A.; Hitchcock, J.R.; Ross, E.A.; Lopez-Macias, C.; Lakey, J.; Martin, L.B.; Toellner, K.M.; MacLennan, C.A.; et al. The capsular polysaccharide Vi from Salmonella typhi is a B1b antigen. J. Immunol. 2012, 189, 5527–5532. [Google Scholar] [CrossRef] [PubMed]
- Garg, R.; Akhade, A.S.; Yadav, J.; Qadri, A. MyD88-dependent pro-inflammatory activity in Vi polysaccharide vaccine against typhoid promotes Ab switching to IgG. Innate Immun. 2015, 21, 778–783. [Google Scholar] [CrossRef] [PubMed]
- Fraser, A.; Paul, M.; Goldberg, E.; Acosta, C.J.; Leibovici, L. Typhoid fever vaccines: Systematic review and meta-analysis of randomised controlled trials. Vaccine 2007, 25, 7848–7857. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.Y.; Ho, V.A.; Khiem, H.B.; Trach, D.D.; Bay, P.V.; Thanh, T.C.; Kossaczka, Z.; Bryla, D.A.; Shiloach, J.; Robbins, J.B.; et al. The efficacy of a Salmonella typhi Vi conjugate vaccine in two-to-five-year-old children. N. Engl. J. Med. 2001, 344, 1263–1269. [Google Scholar] [CrossRef] [PubMed]
- Micoli, F.; Rondini, S.; Pisoni, I.; Proietti, D.; Berti, F.; Costantino, P.; Rappuoli, R.; Szu, S.; Saul, A.; Martin, L.B. Vi-CRM(197) as a new conjugate vaccine against Salmonella Typhi. Vaccine 2011, 29, 712–720. [Google Scholar] [CrossRef] [PubMed]
- Romeu, B.; Lastre, M.; Reyes, L.; Gonzalez, E.; Borrero, Y.; Lescaille, D.; Perez, R.; Nunez, D.; Perez, O. Nasal immunization of mice with AFCo1 or AFPL1 plus capsular polysaccharide Vi from Salmonella typhi induces cellular response and memory B and T cell responses. Vaccine 2014, 32, 6971–6978. [Google Scholar] [CrossRef] [PubMed]
- Finne, J.; Leinonen, M.; Makela, P.H. Antigenic similarities between brain components and bacteria causing meningitis. Implications for vaccine development and pathogenesis. Lancet 1983, 2, 355–357. [Google Scholar] [CrossRef]
- Stephens, D.S.; Greenwood, B.; Brandtzaeg, P. Epidemic meningitis, meningococcaemia, and Neisseria meningitidis. Lancet 2007, 369, 2196–2210. [Google Scholar] [CrossRef]
- Zughaier, S.M. Neisseria meningitidis capsular polysaccharides induce inflammatory responses via TLR2 and TLR4-MD-2. J. Leukoc. Biol. 2011, 89, 469–480. [Google Scholar] [CrossRef] [PubMed]
- Gotschlich, E.C.; Goldschneider, I.; Artenstein, M.S. Human immunity to the meningococcus. IV. Immunogenicity of group A and group C meningococcal polysaccharides in human volunteers. J. Exp. Med. 1969, 129, 1367–1384. [Google Scholar] [CrossRef] [PubMed]
- Griffiss, J.M.; Brandt, B.L.; Altieri, P.L.; Pier, G.B.; Berman, S.L. Safety and immunogenicity of group Y and group W135 meningococcal capsular polysaccharide vaccines in adults. Infect. Immun. 1981, 34, 725–732. [Google Scholar] [PubMed]
- Noya, F.; McCormack, D.; Reynolds, D.L.; Neame, D.; Oster, P. Safety and immunogenicity of two doses of quadrivalent meningococcal conjugate vaccine or one dose of meningococcal group C conjugate vaccine, both administered concomitantly with routine immunization to 12- to 18-month-old children. Can. J. Infect. Dis. Med. Microbiol. 2014, 25, 211–216. [Google Scholar] [CrossRef] [PubMed]
- Pina, L.M.; Bassily, E.; Machmer, A.; Hou, V.; Reinhardt, A. Safety and immunogenicity of a quadrivalent meningococcal polysaccharide diphtheria toxoid conjugate vaccine in infants and toddlers: Three multicenter phase III studies. Pediatr. Infect. Dis. J. 2012, 31, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.M.; Danishefsky, S.J. A vision for vaccines built from fully synthetic tumor-associated antigens: From the laboratory to the clinic. J. Am. Chem. Soc. 2013, 135, 14462–14472. [Google Scholar] [CrossRef] [PubMed]
- Brinãs, R.P.; Sundgren, A.; Sahoo, P.; Morey, S.; Rittenhouse-Olson, K.; Wilding, G.E.; Deng, W.; Barchi, J.J. Design and synthesis of multifunctional gold nanoparticles bearing tumor-associated glycopeptide antigens as potential cancer vaccines. Bioconjugate Chem. 2012, 23, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Comellas-Aragones, M.; Chowdhury, S.; Bentley, P.; Kaczanowska, K.; BenMohamed, L.; Gildersleeve, J.C.; Finn, M.G.; Huang, X. Boosting immunity to small tumor-associated carbohydrates with bacteriophage Qβ capsids. ACS Chem. Biol. 2013, 8, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Lakshminarayanan, V.; Thompson, P.; Wolfert, M.A.; Buskas, T.; Bradley, J.M.; Pathangey, L.B.; Madsen, C.S.; Cohen, P.A.; Gendler, S.J.; Boons, G.J. Immune recognition of tumor-associated mucin MUC1 is achieved by a fully synthetic aberrantly glycosylated MUC1 tripartite vaccine. Proc. Natl. Acad. Sci. USA 2012, 109, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kristensen, D. Opportunities and challenges of developing thermostable vaccines. Expert Rev. Vaccines 2009, 8, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, S.; Erturk-Hasdemir, D.; Ochoa-Reparaz, J.; Reinecker, H.C.; Kasper, D.L. Plasmacytoid dendritic cells mediate anti-inflammatory responses to a gut commensal molecule via both innate and adaptive mechanisms. Cell Host Microbe 2014, 15, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, C.; Schutze, M.P.; Deriaud, E.; Przewlocki, G. The in vivo elimination of CD4+ T cells prevents the induction but not the expression of carrier-induced epitopic suppression. J. Immunol. 1990, 145, 1343–1349. [Google Scholar] [PubMed]
- Miles, D.; Papazisis, K. Rationale for the clinical development of STn-KLH (Theratope) and anti-MUC-1 vaccines in breast cancer. Clin. Breast Cancer 2003, 3, 134–138. [Google Scholar] [CrossRef]
- Slovin, S.F.; Ragupathi, G.; Musselli, C.; Fernandez, C.; Diani, M.; Verbel, D.; Danishefsky, S.; Livingston, P.; Scher, H.I. Thomsen-Friedenreich (TF) antigen as a target for prostate cancer vaccine: Clinical trial results with TF cluster (c)-KLH plus QS21 conjugate vaccine in patients with biochemically relapsed prostate cancer. Cancer Immunol. Immunother. 2005, 54, 694–702. [Google Scholar] [CrossRef] [PubMed]
- Livingston, P.O.; Adluri, S.; Helling, F.; Yao, T.J.; Kensil, C.R.; Newman, M.J.; Marciani, D. Phase 1 trial of immunological adjuvant QS-21 with a GM2 ganglioside-keyhole limpet haemocyanin conjugate vaccine in patients with malignant melanoma. Vaccine 1994, 12, 1275–1280. [Google Scholar] [CrossRef]
- Kalka-Moll, W.M.; Tzianabos, A.O.; Bryant, P.W.; Niemeyer, M.; Ploegh, H.L.; Kasper, D.L. Zwitterionic polysaccharides stimulate T cells by MHC class II-dependent interactions. J. Immunol. 2002, 169, 6149–6153. [Google Scholar] [CrossRef] [PubMed]
- Kabanova, A.; Margarit, I.; Berti, F.; Romano, M.R.; Grandi, G.; Bensi, G.; Chiarot, E.; Proietti, D.; Swennen, E.; Cappelletti, E.; et al. Evaluation of a Group A Streptococcus synthetic oligosaccharide as vaccine candidate. Vaccine 2010, 29, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Ovodov, Y.S. Bacterial capsular antigens. Structural patterns of capsular antigens. Biochemistry 2006, 71, 937–954. [Google Scholar] [CrossRef] [PubMed]
- Pantosti, A.; Tzianabos, A.O.; Onderdonk, A.B.; Kasper, D.L. Immunochemical characterization of two surface polysaccharides of Bacteroides fragilis. Infect. Immun. 1991, 59, 2075–2082. [Google Scholar] [PubMed]
- Wang, Y.; Kalka-Moll, W.M.; Roehrl, M.H.; Kasper, D.L. Structural basis of the abscess-modulating polysaccharide A2 from Bacteroides fragilis. Proc. Natl. Acad. Sci. USA 2000, 97, 13478–13483. [Google Scholar] [CrossRef] [PubMed]
- Stingele, F.; Corthésy, B.; Kusy, N.; Porcelli, S.A.; Kasper, D.L.; Tzianabos, A.O. Zwitterionic polysaccharides stimulate T cells with no preferential Vβ usage and promote anergy, resulting in protection against experimental abscess formation. J. Immunol. 2004, 172, 1483–1490. [Google Scholar] [CrossRef] [PubMed]
- Tzianabos, A.O.; Wang, J.Y.; Lee, J.C. Structural rationale for the modulation of abscess formation by Staphylococcus aureus capsular polysaccharides. Proc. Natl. Acad. Sci. USA 2001, 98, 9365–9370. [Google Scholar] [CrossRef] [PubMed]
- Gallorini, S.; Berti, F.; Parente, P.; Baronio, R.; Aprea, S.; D’Oro, U.; Pizza, M.; Telford, J.L.; Wack, A. Introduction of zwitterionic motifs into bacterial polysaccharides generates TLR2 agonists able to activate APCs. J. Immunol. 2007, 179, 8208–8215. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; McLoughlin, R.M.; Cobb, B.A.; Charrel-Dennis, M.; Zaleski, K.J.; Golenbock, D.; Tzianabos, A.O.; Kasper, D.L. A bacterial carbohydrate links innate and adaptive responses through Toll-like receptor 2. J. Exp. Med. 2006, 203, 2853–2863. [Google Scholar] [CrossRef] [PubMed]
- Polk, B.F.; Kasper, D.L. Bacteroides fragilis Subspecies in Clinical Isolates. Ann. Intern. Med. 1977, 86, 569–571. [Google Scholar] [CrossRef] [PubMed]
- Onderdonk, A.B.; Kasper, D.L.; Cisneros, R.L.; Bartlett, J.G. The capsular polysaccharide of Bacteroides fragilis as a virulence factor: Comparison of the pathogenic potential of encapsulated and unencapsulated strains. J. Infect. Dis. 1977, 136, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Kasper, D.L.; Onderdonk, A.B.; Crabb, J.; Bartlett, J.G. Protective efficacy of immunization with capsular antigen against experimental infection with Bacteroides fragilis. J. Infect. Dis. 1979, 140, 724–731. [Google Scholar] [CrossRef] [PubMed]
- Tzianabos, A.O.; Kasper, D.L.; Cisneros, R.L.; Smith, R.S.; Onderdonk, A.B. Polysaccharide-mediated protection against abscess formation in experimental intra-abdominal sepsis. J. Clin. Invest. 1995, 96, 2727–2731. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M.E.; Onderdonk, A.B.; Kasper, D.L.; Finberg, R.W. Cellular immunity to Bacteroides fragilis capsular polysaccharide. J. Exp. Med. 1982, 155, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Baumann, H.; Tzianabos, A.O.; Brisson, J.R.; Kasper, D.L.; Jennings, H.J. Structural elucidation of two capsular polysaccharides from one strain of Bacteroides fragilis using high-resolution NMR spectroscopy. Biochemistry 1992, 31, 4081–4089. [Google Scholar] [CrossRef] [PubMed]
- Cobb, B.A.; Wang, Q.; Tzianabos, A.O.; Kasper, D.L. Polysaccharide processing and presentation by the MHCII pathway. Cell 2004, 117, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Cobb, B.A.; Kasper, D.L. Zwitterionic capsular polysaccharides: The new MHCII-dependent antigens. Cell. Microbiol. 2005, 7, 1398–1403. [Google Scholar] [CrossRef] [PubMed]
- Kalka-Moll, W.M.; Tzianabos, A.O.; Wang, Y.; Carey, V.J.; Finberg, R.W.; Onderdonk, A.B.; Kasper, D.L. Effect of molecular size on the ability of zwitterionic polysaccharides to stimulate cellular immunity. J. Immunol. 2000, 164, 719–724. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Kasper, D.L. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef] [PubMed]
- Troy, E.B.; Kasper, D.L. Beneficial effects of Bacteroides fragilis polysaccharides on the immune system. Front. Biosci. 2010, 15, 25–34. [Google Scholar] [CrossRef]
- Ruiz-Perez, B.; Chung, D.R.; Sharpe, A.H.; Yagita, H.; Kalka-Moll, W.M.; Sayegh, M.H.; Kasper, D.L.; Tzianabos, A.O. Modulation of surgical fibrosis by microbial zwitterionic polysaccharides. Proc. Natl. Acad. Sci. USA 2005, 102, 16753–16758. [Google Scholar] [CrossRef] [PubMed]
- Rouleau, M.; Cottrez, F.; Bigler, M.; Antonenko, S.; Carballido, J.M.; Zlotnik, A.; Roncarolo, M.G.; Groux, H. IL-10 transgenic mice present a defect in T cell development reminiscent of SCID patients. J. Immunol. 1999, 163, 1420–1427. [Google Scholar] [PubMed]
- Surana, N.K.; Kasper, D.L. The yin yang of bacterial polysaccharides: Lessons learned from B. fragilis PSA. Immunol. Rev. 2012, 245, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Lindberg, B.; Lindqvist, B.; Lonngren, J.; Powell, D.A. Structural studies of the capsular polysaccharide from Streptococcus pneumoniae type 1. Carbohydr. Res. 1980, 78, 111–117. [Google Scholar] [CrossRef]
- Velez, C.D.; Lewis, C.J.; Kasper, D.L.; Cobb, B.A. Type I Streptococcus pneumoniae carbohydrate utilizes a nitric oxide and MHC II-dependent pathway for antigen presentation. Immunology 2009, 127, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Hochkeppel, H.K.; Braun, D.G.; Vischer, W.; Imm, A.; Sutter, S.; Staeubli, U.; Guggenheim, R.; Kaplan, E.L.; Boutonnier, A.; Fournier, J.M. Serotyping and electron microscopy studies of Staphylococcus aureus clinical isolates with monoclonal antibodies to capsular polysaccharide types 5 and 8. J. Clin. Microbiol. 1987, 25, 526–530. [Google Scholar] [PubMed]
- Arbeit, R.D.; Karakawa, W.W.; Vann, W.F.; Robbins, J.B. Predominance of two newly described capsular polysaccharide types among clinical isolates of Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 1984, 2, 85–91. [Google Scholar] [CrossRef]
- Jones, C. Revised structures for the capsular polysaccharides from Staphylococcus aureus Types 5 and 8, components of novel glycoconjugate vaccines. Carbohydr. Res. 2005, 340, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- De Silva, R.A.; Wang, Q.; Chidley, T.; Appulage, D.K.; Andreana, P.R. Immunological response from an entirely carbohydrate antigen: Design of synthetic vaccines based on Tn−PS A1 conjugates. J. Am. Chem. Soc. 2009, 131, 9622–9623. [Google Scholar] [CrossRef] [PubMed]
- Ju, T.; Otto, V.I.; Cummings, R.D. The Tn antigen-structural simplicity and biological complexity. Angew. Chem. Int. Ed. Engl. 2011, 50, 1770–1791. [Google Scholar] [CrossRef] [PubMed]
- Bourgault, J.P.; Trabbic, K.R.; Shi, M.; Andreana, P.R. Synthesis of the tumor associative alpha-aminooxy disaccharide of the TF antigen and its conjugation to a polysaccharide immune stimulant. Org. Biomol. Chem. 2014, 12, 1699–1702. [Google Scholar] [CrossRef] [PubMed]
- Trabbic, K.R.; De Silva, R.A.; Andreana, P.R. Elucidating structural features of an entirely carbohydrate cancer vaccine construct employing circular dichroism and fluorescent labeling. MedChemComm 2014, 5, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- De Silva, R.A.; Appulage, D.K.; Pietraszkiewicz, H.; Bobbitt, K.R.; Media, J.; Shaw, J.; Valeriote, F.A.; Andreana, P.R. The entirely carbohydrate immunogen Tn-PS A1 induces a cancer cell selective immune response and cytokine IL-17. Cancer Immunol. Immunother. 2012, 61, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, G.; Umemura, M. Interleukin-17 as an effector molecule of innate and acquired immunity against infections. Microbiol. Immunol. 2007, 51, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Muranski, P.; Boni, A.; Antony, P.A.; Cassard, L.; Irvine, K.R.; Kaiser, A.; Paulos, C.M.; Palmer, D.C.; Touloukian, C.E.; Ptak, K.; et al. Tumor-specific Th17-polarized cells eradicate large established melanoma. Blood 2008, 112, 362–373. [Google Scholar] [CrossRef] [PubMed]
- Benchetrit, F.; Ciree, A.; Vives, V.; Warnier, G.; Gey, A.; Sautes-Fridman, C.; Fossiez, F.; Haicheur, N.; Fridman, W.H.; Tartour, E. Interleukin-17 inhibits tumor cell growth by means of a T-cell-dependent mechanism. Blood 2002, 99, 2114–2121. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.A. The advent of the cytokine storm. Immunol. Cell. Biol. 2007, 85, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Round, J.L.; Lee, S.M.; Li, J.; Tran, G.; Jabri, B.; Chatila, T.A.; Mazmanian, S.K. The Toll-like receptor pathway establishes commensal gut colonization. Science 2011, 332, 974–977. [Google Scholar] [CrossRef] [PubMed]
- Kreisman, L.S.C.; Friedman, J.H.; Neaga, A.; Cobb, B.A. Structure and function relations with a T-cell-activating polysaccharide antigen using circular dichroism. Glycobiology 2007, 17, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Shaik, A.A.; Nishat, S.; Andreana, P.R. Stereoselective synthesis of natural and non-natural thomsen-nouveau antigens and hydrazide derivatives. Org. Lett. 2015, 17, 2582–2585. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Andreana, P.R. Synthesis of an aminooxy derivative of the trisaccharide globotriose Gb3. J. Carbohydr. Chem. 2014, 33, 381–394. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Nishat, S.; Andreana, P.R. Synthesis of an aminooxy derivative of the tetrasaccharide repeating unit of streptococcus dysgalactiae 2023 polysaccharide for a PS A1 conjugate vaccine. J. Org. Chem. 2016. [Google Scholar] [CrossRef] [PubMed]
- Buskas, T.; Li, Y.; Boons, G.J. The Immunogenicity of the tumor-associated antigen lewisy may be suppressed by a bifunctional cross-linker required for coupling to a carrier protein. Chem. Eur. J. 2004, 10, 3517–3524. [Google Scholar] [CrossRef] [PubMed]
- Heimburg-Molinaro, J.; Lum, M.; Vijay, G.; Jain, M.; Almogren, A.; Rittenhouse-Olson, K. Cancer vaccines and carbohydrate epitopes. Vaccine 2011, 29, 8802–8826. [Google Scholar] [CrossRef] [PubMed]
- Trabbic, K.R.; Bourgault, J.P.; Shi, M.; Andreana, P.R.; Department of Chemistry and Biochemistry, University of Toledo, Toledo, OH, USA. Unpublished data. 2016.
- Trabbic, K.R.; Bourgault, J.P.; Shi, M.; Clark, M.; Andreana, P.R. Immunological evaluation of the entirely carbohydrate-based Thomsen-Friedenreich—PS B conjugate. Org. Biomol. Chem. 2016, 14, 3350–3355. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, K.; Yadav, A.; Morey, S.; Abdullah, J.; Hrysenko, G.; Eng, J.Y.; Sajjad, M.; Koury, S.; Rittenhouse-Olson, K. Preclinical studies with JAA-F11 anti-Thomsen-Friedenreich monoclonal antibody for human breast cancer. Future Oncol. 2014, 10, 385–399. [Google Scholar] [CrossRef] [PubMed]
- Groneck, L.; Schrama, D.; Fabri, M.; Stephen, T.L.; Harms, F.; Meemboor, S.; Hafke, H.; Bessler, M.; Becker, J.C.; Kalka-Moll, W.M. Oligoclonal CD4+ T cells promote host memory immune responses to zwitterionic polysaccharide of streptococcus pneumoniae. Infect. Immun. 2009, 77, 3705–3712. [Google Scholar] [CrossRef] [PubMed]
- Robbins, J.B.; Austrian, R.; Lee, C.J.; Rastogi, S.C.; Schiffman, G.; Henrichsen, J.; Makela, P.H.; Broome, C.V.; Facklam, R.R.; Tiesjema, R.H.; et al. Considerations for formulating the second-generation pneumococcal capsular polysaccharide vaccine with emphasis on the cross-reactive types within groups. J. Infect. Dis. 1983, 148, 1136–1159. [Google Scholar] [CrossRef] [PubMed]
- McLoughlin, R.M.; Lee, J.C.; Kasper, D.L.; Tzianabos, A.O. IFN-gamma regulated chemokine production determines the outcome of Staphylococcus aureus infection. J. Immunol. 2008, 181, 1323–1332. [Google Scholar] [CrossRef] [PubMed]
- Bagnoli, F.; Bertholet, S.; Grandi, G. Inferring reasons for the failure of Staphylococcus aureus vaccines in clinical trials. Front. Cell. Infect. Microbiol. 2012, 2, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cui, L.; Lipinski, T.; Bundle, D.R. Synthesis of monomeric and dimeric repeating units of the zwitterionic type 1 capsular polysaccharide from Streptococcus pneumoniae. Chem. Eur. J. 2010, 16, 3476–3488. [Google Scholar] [CrossRef] [PubMed]
- Pragani, R.; Seeberger, P.H. Total synthesis of the Bacteroides fragilis zwitterionic polysaccharide A1 repeating unit. J. Am. Chem. Soc. 2011, 133, 102–107. [Google Scholar] [CrossRef] [PubMed]


© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nishat, S.; Andreana, P.R. Entirely Carbohydrate-Based Vaccines: An Emerging Field for Specific and Selective Immune Responses. Vaccines 2016, 4, 19. https://doi.org/10.3390/vaccines4020019
Nishat S, Andreana PR. Entirely Carbohydrate-Based Vaccines: An Emerging Field for Specific and Selective Immune Responses. Vaccines. 2016; 4(2):19. https://doi.org/10.3390/vaccines4020019
Chicago/Turabian StyleNishat, Sharmeen, and Peter R. Andreana. 2016. "Entirely Carbohydrate-Based Vaccines: An Emerging Field for Specific and Selective Immune Responses" Vaccines 4, no. 2: 19. https://doi.org/10.3390/vaccines4020019