Ecology of the African Maize Stalk Borer, Busseola fusca (Lepidoptera: Noctuidae) with Special Reference to Insect-Plant Interactions



Abstract
:
1. Introduction
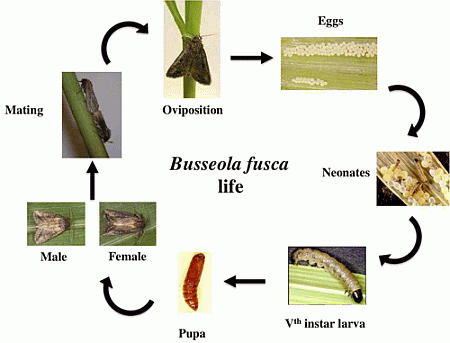
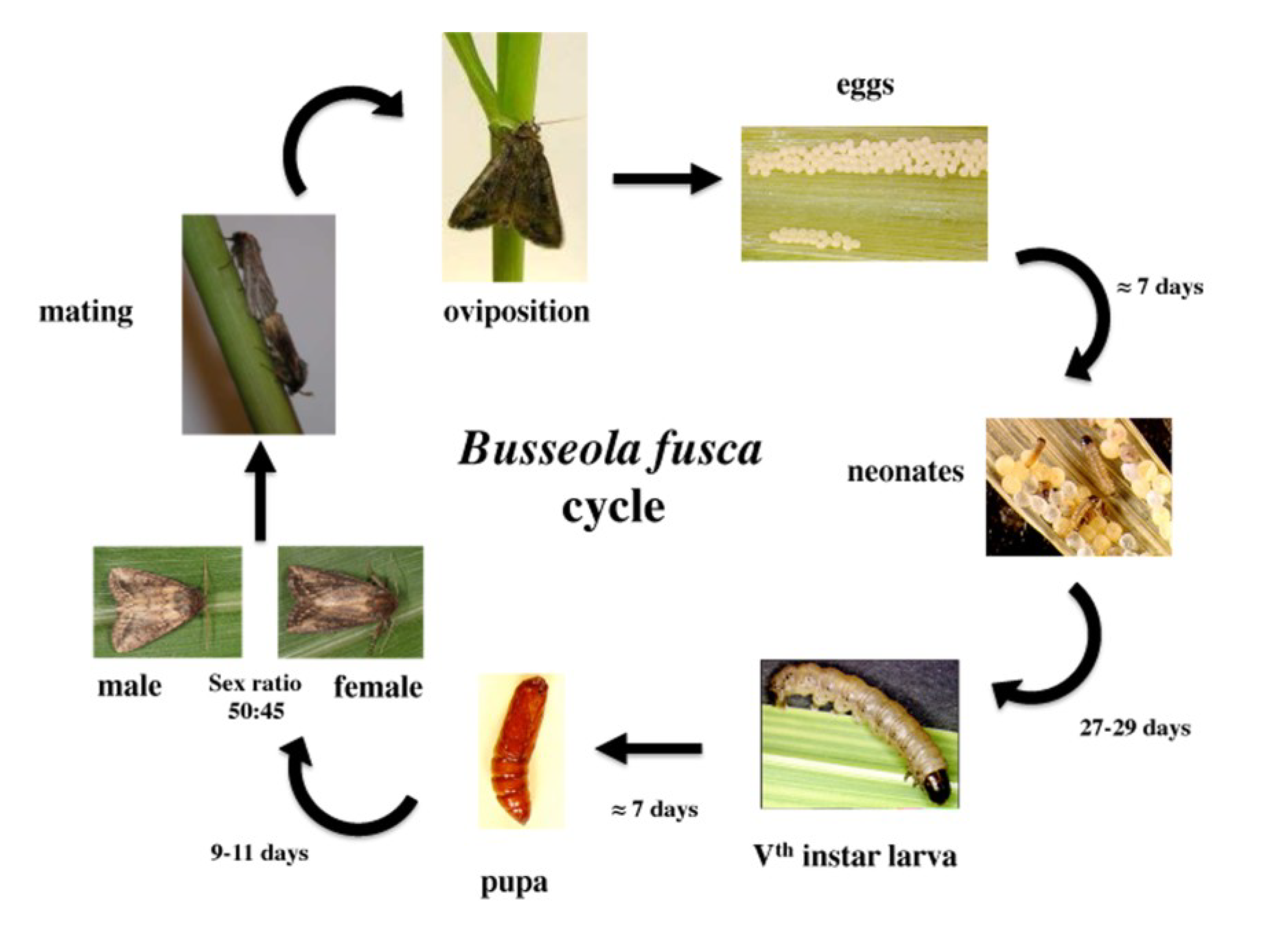
2. Biology and Reproduction
2.1. Eggs
2.2. Larvae
2.3. Adults
2.3.1. Emergence and Life Duration
2.3.2. Pheromones
2.3.3. Mating
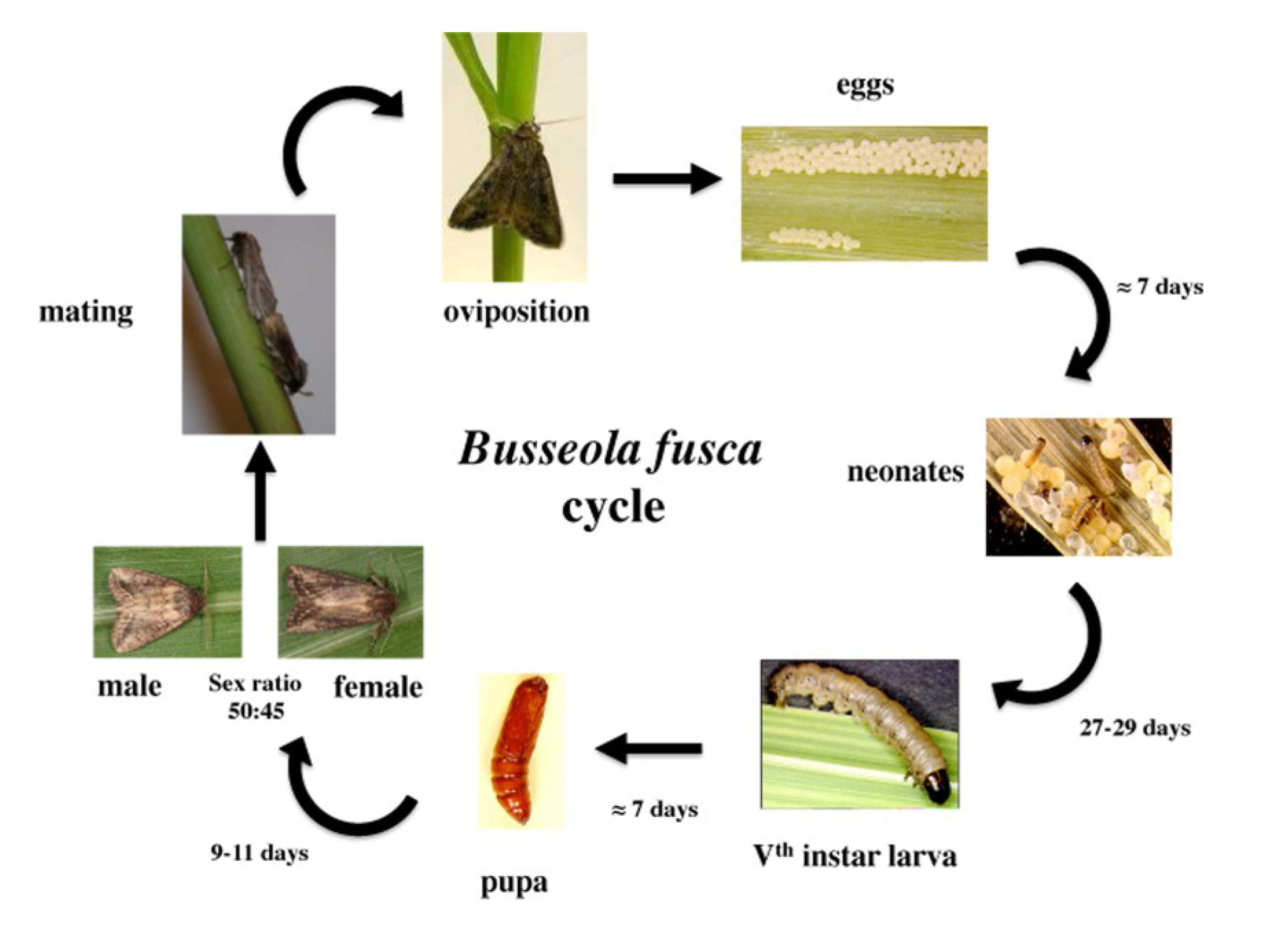
3. Damage Symptoms
4. Geographical Distribution
5. Genetics
6. Host Plant Range

{kind=link}
{kind=link}
{kind=link}
| Host plants | Number of Busseola fusca | Number of Busseola nairobica | Number of Busseola phaia | Number of Busseola segeta | Countries | Total number of individuals collected |
|---|---|---|---|---|---|---|
| Andropogon gayanus Kunth | 0 | 0 | 16 | 0 | TZ | 16 |
| Arundo donax L. | 500 | 0 | 0 | 0 | ER, ET, SA | 500 |
| Cymbopogon giganteus Chiov. | 8 | 0 | 0 | 0 | ZB | 8 |
| Cymbopogon nardus (L.) Rendle | 66 | 0 | 37 | 17 | ET, KE, SA, TZ | 120 |
| Cynodon dactylon (L.) Pers. | 0 | 0 | 0 | 7 | KE | 7 |
| Echinochloa pyramidalis (Lam.) Hitchc. & Chase | 0 | 0 | 4 | 74 | KE, UG | 78 |
| Euchlaena mexicana Schrader | 0 | 0 | 25 | 2 | KE, UG | 27 |
| Heteropogon contortus (L.) P. Beauv. ex Roem. & Schult] | 0 | 0 | 1 | 0 | TZ | 1 |
| Hyparrhenia diplandra (Hack.) Stapf | 0 | 0 | 3 | 0 | TZ | 3 |
| Hyparrhenia dregeana (Nees) Stapf ex Stent | 0 | 0 | 1 | 0 | TZ | 1 |
| Hyparrhenia papillides (A. Rich.) Stapf | 0 | 0 | 0 | 5 | KE, UG | 5 |
| Hyparrhenia schimperi (Hochst. ex A. Rich.) | 0 | 0 | 7 | 0 | MO | 7 |
| Hyparrhenia sp. | 0 | 0 | 30 | 0 | TZ | 30 |
| Melinis minutiflora P. Beauv. | 0 | 0 | 0 | 2 | CA | 2 |
| Panicum atrosanguineum Hochst. ex A. Rich. | 0 | 0 | 45 | 0 | TZ | 45 |
| Panicum deustum Thunb. | 0 | 2,559 | 0 | 199 | ET, KE, UG | 2,758 |
| Panicum mapalende Pilg. | 0 | 0 | 0 | 10 | JE, TZ | 10 |
| Panicum maximum Jacq. | 47 | 0 | 72 | 221 | ET, KE, MO, RC, DRC, RW, UG | 340 |
| Pennisetum clandestinum Chiov. | 0 | 0 | 0 | 38 | CA, ET, KE | 38 |
| Pennisetum macrourum Trin. | 0 | 0 | 7 | 18 | KE, TZ | 25 |
| Pennisetum purpureum Shumach. | 59 | 25 | 507 | 1,184 | CA, ER, ET, KE, MO, UG, DRC, RW, TZ, ZB | 1,775 |
| Pennisetum unisetum (Nees) Benth | 0 | 0 | 1 | 17 | KE, ZB | 18 |
| Phragmites mauritianus Kunth. | 0 | 0 | 11 | 0 | TZ | 11 |
| Setaria megaphylla (Steud.) T. Duran & Schinz | 3 | 0 | 10 | 2 | ET, RC, TZ | 15 |
| Setaria plicatillis (Hochst.) Engl. | 0 | 3 | 0 | 0 | KE | 3 |
| Setaria sphacelata (Schumach.) Moss | 0 | 0 | 3 | 12 | CA, TZ | 15 |
| Snowdenia polystachya (Fresen.) Pilg. | 0 | 0 | 2 | 0 | TZ | 2 |
| Sorghum arundinaceum (Desv.) Stapf | 135 | 0 | 0 | 10 | KE, SA, TZ, UG | 145 |
| Sporobolus macranthelus Chiov. | 0 | 0 | 2 | 0 | TZ | 2 |
| Total number of individuals | 818 | 2,587 | 784 | 1,818 | 6,007 |
7. Host Plant Preference and Selection
7.1. Adults Behaviour and Preferences
7.1.1. Moth Flight Patterns
7.1.2. Behavioural Bases of Host Plant Selection
7.2. Larval Behaviour and Preferences
7.2.1. Larval Migration Patterns
7.2.2. Behavioural Bases of Host Plant Selection
8. Integrated Management of Busseola fusca
8.1. Mixed Cropping
8.2. The Role of Wild Habitats
8.3. The Effect of Soil Fertility on Pest Infestation and Yields
8.4. Host-Plant Resistance
8.5. Genetically Modified Maize
8.6. Plant-Derived Pesticides
9. Conclusions
- (i)
- Physical characteristics: Stem circumference, plant pubescence and the tightness of the leaf sheath strongly influence host plant acceptance by gravid females for oviposition. High levels of Si in plant epidermal cells provide a physical barrier by increasing leaf abrasion, which subsequently increases wearing off of B. fusca larvae mandibles, which physically deter larval feeding.
- (ii)
- Chemical characteristics: Plant volatiles are used by the gravid females for host plant finding. After landing, the cuticular chemical composition of the plant surface conditions the host plant acceptance by the ovipositing females. In addition, the balance of sucrose and turanose in leaves influences the host plant choice made by the larvae.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fuller, C. First Report of the Government Entomologist; Department of Agriculture: Pietermaritzburg, Natal, South Africa; pp. 1899–1900.
- Hampson, G.F. Sesamia fusca, n.sp. Ann. S. Afr. Museum 1902, 2, 296. [Google Scholar]
- Tams, W.H.; Bowden, J. A revision of the african species of Sesamia Guenée and related genera (Agrotidae-Lepidoptera). Bull. Entomol. Res. 1953, 43, 645–678. [Google Scholar] [CrossRef]
- Mally, C.W. The mealie-stalk borer. Sesamia fusca, Hampson. Agric. J. 1905, 15, 1–12. [Google Scholar]
- Mally, C.W. The Maize Stalk Borer, Busseola Fusca, Fuller. In Bulletin; Department of Agriculture of Union of South Africa: Pretoria, South Africa, 1920; p. 3. [Google Scholar]
- Kfir, R.; Overholt, W.A.; Khan, Z.R.; Polaszek, A. Biology and management of economically important lepidopteran cereal stem borers in Africa. Annu. Rev. Entomol. 2002, 47, 701–731. [Google Scholar] [CrossRef]
- Van den Berg, J.; van Rensburg, J.B.J; Pringle, K.L. Comparative injuriousness of Busseola fusca (Lepidoptera: Noctuidae) and Chilo partellus (Lepidoptera: Pyralidae) on grain sorghum. Bull. Entomol. Res. 1991, 82, 137–143. [Google Scholar]
- De Groote, H. Maize yield losses from stemborers in Kenya. Insect Sci. Appl. 2002, 22, 89–96. [Google Scholar]
- Chabi-Olaye, A.; Nolte, C.; Schulthess, F.; Borgemeister, C. Relationships of intercropped maize, stem borer damage to maize yield and land-use efficiency in the humid forest of Cameroon. Bull. Entomol. Res. 2005, 95, 417–427. [Google Scholar]
- Cardwell, K.; Schulthess, F.; Ndemah, R.; Ngoko, Z. A systems approach to assess crop health and maize yield losses due to pests and diseases in Cameroon. Agric. Ecosyst. Environ. 1997, 65, 33–47. [Google Scholar] [CrossRef]
- Chabi-Olaye, A.; Nolte, C.; Schulthess, F.; Borgemeister, C. Effects of grain legumes and cover crops on maize yield and plant damage by Busseola fusca (Fuller) (Lepidoptera: Noctuidae) in the humid forest of southern Cameroon. Agr. Ecosyst. Environ. 2005, 108, 17–28. [Google Scholar] [CrossRef]
- Khan, Z.R.; Chiliswa, P.; Ampong-Nyarko, K.; Smart, L.E.; Polaszek, A.; Wandera, J.; Mulaa, M.A. Utilization of wild gramineous plants for management of cereal stemborers in Africa. Insect Sci. Appl. 1997, 1, 143–150. [Google Scholar]
- Muyekho, F.N.; Barrion, A.T.; Khan, Z.R. Host range for stemborers and associated natural enemies in different farming systems of Kenya. Insect Sci. Appl. 2005, 3, 173–183. [Google Scholar]
- Calatayud, P.-A.; Ahuya, P.O.; Wanjoya, A.; Le Ru, B.; Silvain, J.-F.; Frérot, B. Importance of plant physical cues in host acceptance for oviposition by Busseola fusca. Entomol. Exp. Appl. 2008, 126, 233–243. [Google Scholar] [CrossRef]
- Calatayud, P.-A.; Guénégo, H.; Ahuya, P.; Wanjoya, A.; Le Ru, B.; Silvain, J.-F.; Frérot, B. Flight and oviposition behaviour of the African stem borer, Busseola fusca, on various host plant species. Entomol. Exp. Appl. 2008, 129, 348–355. [Google Scholar] [CrossRef]
- Kaufmann, T. Behavioural biology, feeding habits and ecology of three species of maize stemborers: Eldana saccharina (Lepidoptera: Pyralidae), Sesamia calamistis and Busseola fusca (Noctuidae) in Ibadan, Nigeria, West Africa. J. Georgia Entomol. Soc. 1983, 18, 259–272. [Google Scholar]
- Calatayud, P.-A.; Gitau, C.; Calatayud, S.; Dupas, S.; Le Ru, B.; Silvain, J.-F. Variability in the reproductive biology and in resistance against Cotesia sesamiae among two Busseola fusca populations. J. Appl. Biol. 2011, 135, 423–429. [Google Scholar]
- Kruger, M.; van Rensburg, J.B.J.; van den Berg, J. Reproductive biology of Bt-resistant and susceptible field-collected larvae of the maize stem borer, Busseola fusca (Lepidoptera: Noctuidae). Afr. Entomol. 2012, 20, 35–43. [Google Scholar]
- Kruger, M.; van Rensburg, J.B.J.; van den Berg, J. No fitness costs associated with resistance of Busseola fusca (Lepidoptera: Noctuidae) to genetically modified Bt maize. Crop Prot. 2014, 55, 1–6. [Google Scholar]
- Sezonlin, M.; Dupas, S.; Le Ru, B.; Le Gall, P.; Moyal, P.; Calatayud, P.-A.; Faure, N.; Silvain, J.-F. Phylogeography and population genetics of the maize stalk borer Busseola fusca (Lepidoptera, Noctuidae) in sub-Saharan Africa. Mol. Ecol. 2006, 15, 407–420. [Google Scholar] [CrossRef]
- Harris, K.M. Lepidopterous stem borers of cereals in Nigeria. Bull. Entomol. Res. 1962, 53, 139–171. [Google Scholar] [CrossRef]
- Schulthess, F.; Bosque-Pérez, N.A.; Chabi-Olaye, A.; Gounou, S.; Ndemah, R.; Goergen, G. Exchanging natural enemies species of lepidopterous cereal stemborers between African regions. Insect Sci. Appl. 1997, 17, 97–108. [Google Scholar]
- Unnithan, G.C. Development and reproductive biology of the maize stemborer Busseola fusca Fuller (Lepid., Noctuidae). J. Appl. Entomol. 1987, 104, 172–179. [Google Scholar]
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. Ecology of the maize stalk borer, Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Bull. Entomol. Res. 1987, 77, 255–269. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. The influence of rainfall on seasonal abundance and flight activity of the maize stalk borer, Busseola fusca in South Africa. S. Afr. J. Plant Soil 1987, 4, 183–187. [Google Scholar]
- Usua, E.J. Observations on diapausing larvae of Busseola fusca. J. Econ. Entomol. 1967, 60, 1466–1467. [Google Scholar]
- Ingram, W.R. The lepidopterous stalk borers associated with Gramineae in Uganda. Bull. Entomol. Res. 1958, 49, 367–383. [Google Scholar]
- Van Rensburg, J.B.J. Seasonal moth flight activity of the maize stalk borer, Busseola fusca (Fuller) (Lepidoptera: Noctuidae) in small farming areas of South Africa. Appl. Plant Sci. 1997, 11, 20–23. [Google Scholar]
- Onyango, F.O.; Ochieng’-Odero, J.P.R. Continuous rearing of the maize stem borer Busseola fusca on an artificial diet. Entomol. Exp. Appl. 1994, 73, 139–144. [Google Scholar]
- Ratnadass, A.; Traore, T.; Sylla, M.; Diarra, D. Improved techniques for mass-rearing Busseola fusca (Lepidoptera: Noctuidae) on an artificial diet. Afr. Entomol. 2001, 9, 1–9. [Google Scholar]
- Busseola fusca (African Stem Borer, Maize Stalk Borer). Available online: http://www.biodiversityexplorer.org/lepidoptera/noctuidae/busseola_fusca.htm (accessed on 20 March 2014).
- Malusi, P.; Okuku, G.; ICIPE, Nairobi, Kenya. Personal observations. 2013.
- Walters, M.C. The influence of the weather on maize pests. In The value of phenology in increasing the productivity of crops. Tech. Bull. Department Agric. Serv. 1979, 150, 64–72. [Google Scholar]
- Usua, E.J. Some notes on maize stemborers in Nigeria. J. Econ. Entomol. 1970, 63, 776–778. [Google Scholar]
- Okuda, T. Significance of water contact as a factor terminating larval diapause in a stem borer, Busseola fusca. Entomol. Exp. Appl. 1990, 57, 151–155. [Google Scholar] [CrossRef]
- Unnithan, G.C.; Paye, S.O. Factors involved in mating, longevity, fecundity and egg fertility in the maize stem-borer, Busseola fusca (Fuller) (Lep., Noctuidae). J. Appl. Entomol. 1990, 109, 295–301. [Google Scholar] [CrossRef]
- Calatayud, P.-A.; Guénégo, H.; Le Ru, B.; Silvain, J.-F.; Frérot, B. Temporal patterns of emergence, calling behaviour and oviposition period of the maize stem borer, Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Ann. Société Entomologique Fr. 2007, 43, 63–68. [Google Scholar]
- Frérot, B.; Félix, A.-E.; Ene, S.; Calatayud, P.-A.; Le Ru, B.; Guénégo, H. Courtship behaviour of the African maize stem borer: Busseola fusca (Fuller) (Lepidoptera: Noctuidae) under laboratory conditions. Ann. Société Entomologique Fr. 2006, 42, 413–416. [Google Scholar]
- Nesbitt, B.F.; Beevor, P.S.; Cork, A.; Hall, D.R.; Lester, R.; Blair, B.W.; Tannock, J. Identification of the female sex pheromone of the maize stalk borer, Busseola fusca: A preliminary note. Trop. Pest Manage. 1980, 26, 327. [Google Scholar]
- Félix, A.-E.; Genestier, G.; Malosse, C.; Calatayud, P.-A.; Le Ru, B.; Silvain, J.-F.; Frérot, B. Variability in pheromone communication among different haplotype populations of Busseola fusca. J. Chem. Ecol. 2009, 35, 618–623. [Google Scholar] [CrossRef]
- Félix, A.-E.; Smaïl, T.; Frérot, B. Host plants and reproductive behaviour in the African maize stemborer, Busseola fusca (Fuller 1901) (Lepidoptera: Noctuidae). Ann. Société Entomologique Fr. 2013, 49, 68–72. [Google Scholar] [CrossRef]
- Félix, A.-E. Chemical ecology and phylogenetic approaches in three African Lepidoptera species of genus Busseola (Noctuidae). PhD Thesis, Physiology and Organisms Biology of Paris XI University, Paris, France, 2008. [Google Scholar]
- Ong’amo, G.O.; Le Ru, B.; Dupas, S.; Moyal, P.; Ngala, L.; Muchugu, E.; Calatayud, P.-A.; Silvain, J.-F. The role of wild host plants in the abundance of lepidopteran stem borers along altitudinal gradient in Kenya. Ann. Société Entomologique Fr. 2006, 42, 363–370. [Google Scholar] [CrossRef]
- Bate, R.; van Rensburg, G.D.J.; Giliomee, J.H. Distribution and occurrence of the stem-borer Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae) on maize in South Africa. J. Afr. Zool. 1991, 105, 11–15. [Google Scholar]
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. The comparative abundance and in-season distribution of larval infestations of Busseola fusca (Fuller) (Lepidoptera : Noctuidae) and Chilo partellus (Swinhoe)(Lepidoptera : Noctuidae). J. Entomol. Soc. S. Afr. 1988, 51, 138–150. [Google Scholar]
- Kfir, R. Competitive displacement of Busseola fusca (Lepidoptera: Noctuidae) by Chilo partellus (Lepidoptera: Pyralidae). Ann. Entomol. Soc. Am. 1997, 5, 619–624. [Google Scholar]
- Chabi-Olaye, A.; Schulthess, F.; Borgemeister, C. Effects of nitrogen and potassium combinations on yields and infestations of maize by Busseola fusca (Fuller) (Lepidoptera: Noctuidae) in the humid forest of Cameroon. J. Econ. Entomol. 2008, 101, 90–98. [Google Scholar] [CrossRef]
- Le Ru, B.P.; Ong’amo, G.O.; Moyal, P.; Ngala, L.; Musyoka, B.; Abdullah, Z.; Cugala, D.; Defabachew, B.; Haile, T.A.; Matama-Kauma, T.; et al. Diversity of lepidopteran stem borers on monocotyledonous plants in eastern Africa and the islands of Madagascar and Zanzibar revisited. Bull. Entomol. Res. 2006, 96, 555–563. [Google Scholar] [CrossRef]
- Harris, C.W.; Nwanze, K.E. Busseola fusca (Fuller), the African maize stalk borer: A handbook of information. In Information Bulletin no. 33; ICRISAT, CAB International: Oxon, UK, 1992; p. 85. [Google Scholar]
- Ong’amo, G.O.; Le Ru, B.P.; Dupas, S.; Moyal, P.; Calatayud, P.-A.; Silvain, J.-F. Distribution, pest status and agro-climatic preferences of lepidopteran stem borers of maize in Kenya. Ann. Société Entomologique Fr. 2006, 42, 171–177. [Google Scholar] [CrossRef]
- Ndemah, R.; Schulthess, F.; Nolte, C. The effect of grassy field margins and fertilizer on soil water, plant nutrient levels, stemborer attacks and yield of maize in the humid forest of Cameroon. Ann. Société Entomologique Fr. 2006, 42, 461–470. [Google Scholar] [CrossRef]
- Ndemah, R.; Schulthess, F.; Korie, S.; Borgemeister, C.; Cardwell, K.F. Distribution, relative importance and effect of lepidopterous borers on maize yields in the forest zone and mid-altitude of Cameroon. J. Econ. Entomol. 2001, 94, 1434–1444. [Google Scholar] [CrossRef]
- Krüger, W.; van den Berg, J.; van Hamburg, H. The relative abundance of maize stem borers and their parasitoids at the Tshiombo irrigation scheme in Venda, South Africa. S. Afr. J. Plant Soil 2008, 25, 144–151. [Google Scholar] [CrossRef]
- Waladde, S.M.; van den Berg, J.; Botlohle, P.M.; Mlanjeni, N. Survey of the abundance and distribution of maize stemborers in the Eastern Cape province of South Africa. In Proceedings of the Southern African Stem Borer Management Project, Pietermaritzburg, Natal, South Africa, 1–5 July 2002; van den Berg, J., Uys, V.M., Eds.; pp. 13–16.
- Ebenebe, A.A.; van den Berg, J.; van der Linde, T.C. Distribution and relative abundance of stalk borers of maize and sorghum in Lesotho. Afr. Plant Prot. 1999, 5, 77–82. [Google Scholar]
- Khan, Z.R.; Lwande, K.; Ampong-Nyarko, K.V. Use of wild habitat in the management of cereal stemborers. In ICIPE Scientific Report; ICIPE: Nairobi, Kenya, 1995–1997; pp. 59–67. [Google Scholar]
- Sezonlin, M.; Dupas, S.; Le Ru, B.; Faure, N.; Le Gall, P.; Silvain, J.-F. Phylogeographic pattern and regional evolutionary history of the maize stalk borer Busseola fusca (Fuller) (Lepidoptera: Noctuidae) in sub-Saharan Africa. Ann. Société Entomologique Fr. 2006, 42, 399–351. [Google Scholar]
- Sezonlin, M.; Ndemah, R.; Georgen, G.; Le Ru, B.; Dupas, S.; Silvain, J.-F. Genetic structure and origin of Busseola fusca populations in Cameroon. Entomol. Exp. Appl. 2012, 145, 143–152. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. Plant population and cultivar effects caused by the maize stalk borer, Busseola fusa (Lepidoptera: Noctuidae). S. Afr. J. Plant Soil 1988, 5, 215–218. [Google Scholar] [CrossRef]
- Faure, N.; Silvain, J.-F. Characterization of eight microsatellite loci in the maize stalk borer Busseola fusca Fuller, (Lepidoptera: Noctuidae). Mol. Ecol. 2005, 5, 846–848. [Google Scholar] [CrossRef]
- Dupas, S.; Le Ru, B.; Branca, A.; Faure, N.; Gigot, G.; Campagne, P.; Sezonlin, M.; Ndemah, R.; Calatayud, P.-A.; Silvain, J.-F. Phyloeography in continuous space: Coupling species distribution models and circuit theory to assess the effect of contiguous migration at different climatic periods on genetic differentiation in Busseola fusca (Lepidoptera: Noctuidae). Mol. Ecol. 2014, 23, 2313–2325. [Google Scholar] [CrossRef]
- Ndemah, R.; Gounou, S.; Schulthess, F. The role of wild grasses in the management of lepidopterous cereal stemborers on maize in western Africa. Bull. Entomol. Res. 2002, 92, 507–519. [Google Scholar]
- Le Ru, B.P.; Ong’amo, G.O.; Moyal, P.; Muchugu, E.; Ngala, L.; Musyoka, B.; Abdullah, Z.; Matama-Kauma, T.; Lada, V.Y.; Pallangyo, B.; et al. Geographic distribution and host plant ranges of East African noctuid stem borers. Ann. Société Entomologique Fr. 2006, 42, 353–361. [Google Scholar] [CrossRef]
- Ndemah, R.; Schulthess, F.; Le Ru, B.; Bame, I. Lepidopteran cereal stemborers and associated natural enemies on maize and wild grass hosts in Cameroon. J. Appl. Entomol. 2007, 131, 658–668. [Google Scholar] [CrossRef]
- Ong’amo, G.O.; Le Ru, B.; Calatayud, P.-A.; Silvain, J.-F. Composition of stem borer communities in selected vegetation mosaics in Kenya. Arthropod-Plant Interact. 2013, 7, 267–275. [Google Scholar]
- Otieno, N.A.; Le Ru, B.; Ong’amo, G.O.; Dupas, S.; Calatayud, P.-A.; Makobe, M.; Ochora, J.; Silvain, J.-F. Diversity and abundance of wild host plants of lepidopteran stem borers in two different agroecological zones of Kenya. Ann. Société Entomologique Fr. 2006, 42, 371–380. [Google Scholar] [CrossRef]
- Otieno, N.A.; Le Ru, B.; Ong’amo, G.O.; Moyal, P.; Dupas, S.; Calatayud, P.-A.; Silvain, J.-F. Diversity and abundance of wild host plants of lepidopteran stem borers in two agro-ecological zones of Kenya. Int. J. Biodivers. Sci. Manage. 2008, 4, 1–12. [Google Scholar] [CrossRef]
- Moolman, H.J.; van den Berg, J.; Conlong, D.; Cugala, D.; Siebert, S.J.; Le Ru, B.P. Species diversity and distribution of Lepidopteran stem borers in South Africa and Mozambique. J. Appl. Entomol. 2014, 138, 152–166. [Google Scholar]
- Rebe, M.; van den Berg, J.; McGeogh, M.A. Colonization of cultivated and wild graminaceous host plants by Busseola fusca (Fuller) (Lepidoptera: Noctuidae) and Chilo partellus (Swinhoe) (Lepidoptera: Crambidae) under field conditions. Afr. Entomol. 2004, 12, 187–199. [Google Scholar]
- Félix, A.-E.; Calatayud, P.-A.; Le Ru, B.; Capdevielle-Dulac, C.; Ong’amo, G.; Silvain, J.-F.; Frérot, B. To be or not to be a species: use of reproductive isolation experiments and genetic analysis to clarify the taxonomic status of two Busseola (Lepidoptera: Noctuidae) species in Kenya. Ann. Société Entomologique Fr. 2014, 49, 345–354. [Google Scholar]
- Ong’amo, G.O.; Le Ru, B.; Campagne, P.; Branca, A.; Calatayud, P.-A.; Capdevielle-Dulac, C.; Silvain, J.-F. Genetic diversity and population structure of Busseola segeta Bowden (Lepidoptera; Noctuidae): A case study of host use diversification in Guineo-Congolian rainforest relic area in Kenya. Insects 2012, 3, 1156–1170. [Google Scholar] [CrossRef]
- Nye, I.W.B. The insect pests of graminaceous crops in East Africa. Colonial Res. Stud. 1960, 31, 1–48. [Google Scholar]
- Le Ru, B.; IRD c/o ICIPE, Nairobi, Kenya. Unpublished data. 2014.
- Van den Berg, J.; de Bruyn, A.J.M; van Hamburg, H. Oviposition preference and survival of the maize stem borer, Busseola fusca (Lepidoptera: Noctuidae) on Napier grasses (Pennisetum spp.) and maize. Afr. Entomol. 2006, 14, 211–218. [Google Scholar]
- Bernays, E.A.; Chapman, R.F. Host-Plant Selection by Phytophagous Insects; Chapman & Hall: London, UK, 1994. [Google Scholar]
- Schoonhoven, L.M.; van Loon, J.J.A.; Dicke, P. Insect-Plant Biology, 2nd ed.; Oxford University Press: Oxford, UK, 2005. [Google Scholar]
- Van Rensburg, J.B.J.; van Rensburg, G.D.J.; Giliomee, J.H.; Walters, M.C. The influence of rainfall on the seasonal abundance and flight activity of the maize stalk borer, Busseola fusca in South Africa. S. Afr. J. Plant Soil 1997, 4, 183–187. [Google Scholar]
- Du Plessis, C.; Lea, M.C. The maize stalk borer Calamistis fusca (Hmpsn). In Bulletin of the Department of Agriculture and Forestry of the Union of South Africa; Government Printers: Pretoria, South Africa, 1943; 238, p. 51. [Google Scholar]
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. Geographical variation in the seasonal moth flight activity of the maize stem borer, Busseola fusca (Fuller) in South Africa. S. Afr. J. Plant Soil 1985, 2, 123–126. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. Selective oviposition by the maize stalk borer, Busseola fusca (Fuller). J. Entomol. Soc. S. Afr. 1989, 52, 105–108. [Google Scholar]
- Revington, J.; van Rensburg, J.B.J.; Burkhardt, G.; Knauf, W. Preliminary field trials with a pheromone based monitoring system for the maize stalk borer, Busseola fusca (Fuller) (Lep. Noctuidae). J. Entomol. Soc. S. Afr. 1984, 47, 107–113. [Google Scholar]
- Van Rensburg, J.B.J. Evaluation of pheromone trapping systems in relation to light trap captures of the maize stalk borer, Busseola fusca (Fuller) (Lep. Noctuidae). S. Afr. J. Plant Soil 1992, 9, 144–149. [Google Scholar] [CrossRef]
- Calatayud, P.-A.; Ahuya, P.; Le Ru, B. Importance of the experimental set-up used to measure the attractiveness of odours in moth: An example with Busseola fusca. Entomol. Exp. Appl. 2014, in press. [Google Scholar]
- Calatayud, P.-A.; Chimtawi, M.; Tauban, D.; Marion-Poll, F.; Le Ru, B.; Silvain, J.-F.; Frérot, B. Sexual dimorphism of antennal, tarsal and ovipositor chemosensilla in the African stemborer, Busseola fusca (Fuller)(Lepidoptera: Noctuidae). Ann. Société Entomologique Fr. 2006, 42, 403–412. [Google Scholar] [CrossRef]
- Juma, G. Role of plants volatile and surface metabolites in mediating host selection and oviposition of Busseola fusca (Fuller) (Lepidoptera: Noctuidae). M.Sc. Thesis, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya, 2005. [Google Scholar]
- Haile, A.; Hofsvang, T. Host plant preference of the stem borer Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Crop Prot. 2002, 21, 227–233. [Google Scholar] [CrossRef]
- Khan, Z.R.; Pickett, J.A.; van den Berg, J.; Wadhams, L.J.; Woodcock, C.M. Exploiting chemical ecology and species diversity: Stem borer and striga control for maize and sorghum, in Africa. Pest Manage. Sci. 2000, 56, 957–962. [Google Scholar] [CrossRef]
- Le Ru, B.; IRD c/o ICIPE, Nairobi, Kenya. Personal observations. 2013.
- Van Rensburg, J.B.J.; van den Berg, J. Stem borers in grain sorghum: I. injury patterns with time after crop emergence. S. Afr. J. Plant Soil 1992, 9, 73–80. [Google Scholar]
- Van den Berg, J.; van Rensburg, J.B.J. Effect of various directional insecticide sprays against Busseola fusca (Lepidoptera: Noctuidae) and Chilo partellus (Lepidoptera: Pyralidae) in maize and sorghum. S. Afr. J. Plant Soil 1996, 13, 51–54. [Google Scholar] [CrossRef]
- Van den Berg, J.; Le Ru, B.; North West University & IRD c/o ICIPE, Potchefstroom & Nairobi, South Africa & Kenya. Personal observations. 2013.
- Van Rensburg, J.B.J.; Walters, M.C.; Giliomee, J.H. Mortality in natural populations of the maize stalk borer, Busseola fusca (Fuller) (Lepidoptera: Noctuidae) in South Africa. Phytophylactica 1988, 20, 17–19. [Google Scholar]
- Smithers, C.N. Some recent observations on Busseola fusca (Fuller) (Lepidoptera, Noctuidae) in Southern Rhodesia. Bull. Entomol. Res. 1960, 50, 809–819. [Google Scholar] [CrossRef]
- Berdegué, M.; Reitz, S.; Trumble, J.T. Host plant selection and development in Spodoptera exigua: do mother and offspring know best? Entomol. Exp. Appl. 1998, 89, 57–64. [Google Scholar]
- Schoonhoven, L.M.; van Loon, J.J.A. An inventory of taste in caterpillars: each species its own key. Acta Zool. Academiae Scientiarum Hungaricae 2002, 48, 215–263. [Google Scholar]
- Juma, G.; Chimtawi, M.; Ahuya, P.O.; Njagi, P.G.N.; Le Ru, B.; Magoma, G.; Silvain, J.-F.; Calatayud, P.-A. Distribution of chemo- and mechanoreceptors on the antennae and maxillae of Busseola fusca larvae. Entomol. Exp. Appl. 2008, 128, 93–98. [Google Scholar] [CrossRef]
- Calatayud, P.-A.; IRD c/o ICIPE, Nairobi, Kenya. Unpublished data. 2013.
- Juma, G.; Thiongo, M.; Dutaur, L.; Rharrabe, K.; Marion-Poll, F.; Le Ru, B.; Magoma, G.; Silvain, J.-F.; Calatayud, P.-A. Two sugar isomers influence host plant acceptance by a cereal caterpillar pest. Bull. Entomol. Res. 2013, 103, 20–28. [Google Scholar]
- Epstein, E. Silicon. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon-augmented resistance of plants to herbivorous insects. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Juma, G. Basis of host recognition and acceptance of Busseola fusca (Fuller) (Lepidoptera: Noctuidae) larvae. Ph.D. Thesis, Jomo Kenyatta University of Agriculture and Technology, Nairobi, Kenya, 2010. [Google Scholar]
- Van den Berg, J.; Nur, A.F.; Polaszek, A. Cultural control. In Cereal Stem Borers in Africa: Economic importance, Taxonomy, Natural enemies and Control; Polaszek, A., Ed.; International Institute of Entomology, CAB International: Wallingford, UK, 1998; pp. 333–347. [Google Scholar]
- Ndemah, R.; Schulthess, F.; Korie, S.; Borgemeister, C.; Poehling, M.; Cardwell, K.F. Factors affecting infestations of the stalk borer Busseola fusca (Fuller) on maize in the forest Zone of Cameroon with special reference to scelionid egg parasitoids. Environ. Entomol. 2003, 32, 61–70. [Google Scholar] [CrossRef]
- Schulthess, F.; Chabi-Olaye, A.; Gounou, S. Multi-trophic level interactions in a cassava-maize relay cropping system in the humid tropics of West Africa. Bull. Entomol. Res. 2004, 94, 261–272. [Google Scholar]
- Chabi-Olaye, A.; Nolte, C.; Schulthess, F.; Borgemeister, C. Abundance, dispersion and parasitism of the noctuid stem borer Busseola fusca (Fuller) in mono- and intercropped maize in the humid forest zone of southern Cameroon. Bull. Entomol. Res. 2005, 95, 169–177. [Google Scholar]
- Aiyer, A. Mixed cropping in India. Indian J. Agric. Sci. 1949, 19, 439–453. [Google Scholar]
- Finch, S.; Collier, R.H. The influence of host and non-host companion plants on the behaviour of pest insects in field crops. Entomol. Exp. Appl. 2012, 142, 87–96. [Google Scholar] [CrossRef]
- Vandermeer, J. The Ecology of Intercropping. Cambridge University Press: Cambridge, UK, 1989. [Google Scholar]
- Finch, S.; Collier, R.H. Host-plant selection by insects—A theory based on “appropriate/inappropriate landings” by pest insects of cruciferous plants. Entomol. Exp. Appl. 2000, 96, 91–102. [Google Scholar]
- Ndemah, R.; IITA, Yaoundé, Cameroon. Unpublished data. 2006.
- Matama-Kauma, T.; Schulthess, F.; Mueke, J.M.; Omwega, C.O.; Ogwang, J.A. Impact of wild grasses planted as border rows on stemborer infestations in maize in Uganda. Ann. Société Entomologique Fr. 2006, 42, 455–460. [Google Scholar] [CrossRef]
- Mailafiya, D.M.; Le Ru, B.P.; Kairu, E.W.; Calatayud, P.-A.; Dupas, S. Species diversity of lepidopteran stem borer parasitoids in cultivated and natural habitats in Kenya. J. Appl. Entomol. 2009, 133, 416–429. [Google Scholar] [CrossRef]
- Mailafiya, D.M.; Le Ru, B.P.; Kairu, E.W.; Calatayud, P.-A.; Dupas, S. Geographic distribution, host range and perennation of Cotesia sesamiae and Cotesia flavipes Cameron in cultivated and natural habitats in Kenya. Biol. Control 2009, 54, 1–8. [Google Scholar]
- Mailafiya, D.M.; Le Ru, B.P.; Kairu, E.W.; Calatayud, P.-A.; Dupas, S. Factors affecting stem borer parasitoid diversity and parasitism. Environ. Entomol. 2010, 39, 57–67. [Google Scholar] [CrossRef]
- Moolman, H.J.; van den Berg, J.; Conlong, D.; Cugala, D.; Siebert, S.J.; Le Ru, B.P. Diversity of stem borer parasitoids and their associated wild host plants in South Africa and Mozambique. Phytoparasitica 2013, 41, 89–104. [Google Scholar] [CrossRef]
- Sétamou, M.; Schulthess, F.; Bosque-Pérez, N.A.; Thomas-Odjo, A. The effect of stem and cob borers on maize subjected to different nitrogen treatments with special reference to Sesamia calamistis Hampson (Lepidoptera: Noctuidae). Entomogia Exp. Appl. 1995, 77, 205–210. [Google Scholar] [CrossRef]
- Mgoo, V.H.; Makundi, R.H.; Pallangyo, B.; Schulthess, F.; Jiang, N.; Omwega, C.O. Yield loss due to the stemborer Chilo partellus (Swinhoe) (Lepidoptera: Crambidae) at different nitrogen application rates to maize. Ann. Société Entomologique Fr. 2006, 42, 487–494. [Google Scholar] [CrossRef]
- Jiang, N.; Schulthess, F. The effect of nitrogen fertilizer to maize on the bionomics of Chilo partellus (Swinhoe) (Lepidoptera: Crambidae) and performance of its larval parasitoid Cotesia flavipes Cameron (Hymenoptera: Braconidae). Bull. Entomol. Res. 2005, 95, 495–504. [Google Scholar]
- Ali, A.I.; Jiang, N.; Schulthess, F.; Ogol, C.K.P.O.; Omwega, C.O. Effect of nitrogen fertilizer level on infestations of lepidopterous stemborers and yields of maize in Zanzibar. Ann. Société Entomol. Fr. 2006, 42, 481–486. [Google Scholar] [CrossRef]
- Wale, M.; Schulthess, F.; Kairu, E.W.; Omwega, C.O. Cereal yield losses due to lepidopterous stemborers at different nitrogen fertilizer rates in Ethiopia. J. Appl. Entomol. 2006, 130, 220–229. [Google Scholar] [CrossRef]
- Van den Berg, J.; van Rensburg, J.B.J. Infestation and injury levels of stem borers in relation to yield potential of grain sorghum. S. Afr. J. Plant Soil 1991, 8, 127–131. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J.; van Rensburg, G.D.J. Laboratory production of Busseola fusca (Fuller) (Lep. Noctuidae) and techniques for the detection of resistance in maize plants. Afr. Entomol. 1993, 1, 25–28. [Google Scholar]
- Bosque-Perez, N.A.; Schulthess, F. Maize: West and Central Africa. In African Cereal Stemborers: Economic Importance, Taxonomy, Natural Enemies and Control; Polaszek, A., Ed.; CABI: Wallingford, Oxon, UK, 1998; pp. 11–27. [Google Scholar]
- Van Rensburg, J.B.J.; Malan, C. Resistance of maize genotypes to the maize stalk borer, Busseola fusca (Fuller) (Lep. Noctuidae). J. Entomol. Soc. S. Afr. 1990, 53, 49–55. [Google Scholar]
- Van Rensburg, J.B.J.; van den Berg, J. New sources of resistance to the stalk borers Busseola fusca (Fuller) and Chilo partellus Swinhoe in maize. S. Afr. J. Plant Soil 1995, 12, 91–93. [Google Scholar] [CrossRef]
- Van Rensburg, J.B.J.; Klopper, J. Recurrent selection for resistance in maize to the African stalk borer, Busseola fusca (Fuller). S. Afr. J. Plant Soil 2004, 21, 59–62. [Google Scholar] [CrossRef]
- Van den Berg, J. The potential value of stem borer resistance in the management of Busseola fusca (Lepidoptera: Noctuidae) and Chilo partellus (Lepidoptera: Pyralidae) populations in sorghum. Insect Sci. Appl. 1995, 18, 31–36. [Google Scholar]
- Van den Berg, J.; Wenzel, W.G.; van der Westhuizen, M.C. Tolerance and recovery resistance of grain sorghum genotypes artificially infested with Busseola fusca (Fuller) (Lepidoptera: Noctuidae). Insect Sci. Appl. 1994, 15, 61–65. [Google Scholar]
- Wenzel, W.G.; Mohammed, J.; van den Berg, J. Evaluation of accessions of South African sorghum germplasm for use in the development of improved varieties. Afr. Crop Sci. J. 1997, 5, 9–14. [Google Scholar]
- Muturi, P.W.; Rubaihayo, P.; Mgonja, M.; Kyamanywa, S.; Sharma, H.C.; Hash, C.T. Novel source of sorghum tolerance to the African stem borer, Busseola fusca. Afr. J. Plant Sci. 2012, 11, 295–302. [Google Scholar]
- Van Rensburg, J.B.J. First report of field resistance by the stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. S. Afr. J. Plant Soil 2007, 24, 147–151. [Google Scholar] [CrossRef]
- Kruger, M.; van Rensburg, J.B.J.; van den Berg, J. Transgenic Bt maize: Farmers’ perceptions, refuge compliance and reports of stem borer resistance in South Africa. J. Appl. Entomol. 2012, 136, 38–50. [Google Scholar] [CrossRef]
- Campagne, P.; Kruger, M.; Pasquet, R.; Le Ru, B.; van den Berg, J. Dominant Inheritance of field-evolved resistance to Bt corn in Busseola fusca. PLoS One 2013, 8, e69675. [Google Scholar]
- Assefa, Y.; van den Berg, J. Genetically modified maize: Adoption practices of small-scale farmers in South Africa and implications for resource-poor farmers on the continent. Aspects Appl. Biol. 2010, 96, 215–223. [Google Scholar]
- Valencia, A.; Frérot, B.; Guénego, H.; Munera, D.F.; Grossi De Sa, M.F.; Calatayud, P.-A. Effect of Jatropha gossypiifolia leaf extracts on three Lepidoptera species. Rev. Colomb. Entomol. 2006, 32, 45–48. [Google Scholar]
- Schmutterer, H. Properties and potentials of natural pesticides from the neem tree, Azadirachta indica. Ann. Rev. Entomol. 1990, 35, 271–297. [Google Scholar] [CrossRef]
- Assefa, G.A.; Ferdu, A. Insecticidal activity of chinaberry, endod and peper tree against the maize stalk borer Busseola fusca (Lepidoptera: Noctuidae) in southern Ethiopia. Int. J. Pest Manage. 1999, 45, 9–13. [Google Scholar] [CrossRef]
- Ratnadass, A.; Wink, M. The phorbol ester fraction from Jatropha curcas seed oil: Potential and limits for crop protection against insect pests. Int. J. Mol. Sci. 2012, 13, 16157–16171. [Google Scholar] [CrossRef] [Green Version]
- Shiberu, T. In vitro evaluation of aqua extracts of some botanicals against maize stem borer, Busseola fusca F. (Lepidoptera: Noctuidae). J. Plant Pathol. Microbiol. 2013, 4, 179. [Google Scholar]
- Leppik, E. Reproductive isolation and host plant specialization in European corn borer pheromone strains. PhD Thesis, Physiology and Organisms Biology of Paris XI University, Paris, France, 2011. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Calatayud, P.-A.; Le Ru, B.P.; Van den Berg, J.; Schulthess, F. Ecology of the African Maize Stalk Borer, Busseola fusca (Lepidoptera: Noctuidae) with Special Reference to Insect-Plant Interactions. Insects 2014, 5, 539-563. https://doi.org/10.3390/insects5030539
Calatayud P-A, Le Ru BP, Van den Berg J, Schulthess F. Ecology of the African Maize Stalk Borer, Busseola fusca (Lepidoptera: Noctuidae) with Special Reference to Insect-Plant Interactions. Insects. 2014; 5(3):539-563. https://doi.org/10.3390/insects5030539
Chicago/Turabian StyleCalatayud, Paul-André, Bruno P. Le Ru, Johnnie Van den Berg, and Fritz Schulthess. 2014. "Ecology of the African Maize Stalk Borer, Busseola fusca (Lepidoptera: Noctuidae) with Special Reference to Insect-Plant Interactions" Insects 5, no. 3: 539-563. https://doi.org/10.3390/insects5030539