Ovarian Cancer Stroma: Pathophysiology and the Roles in Cancer Development

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
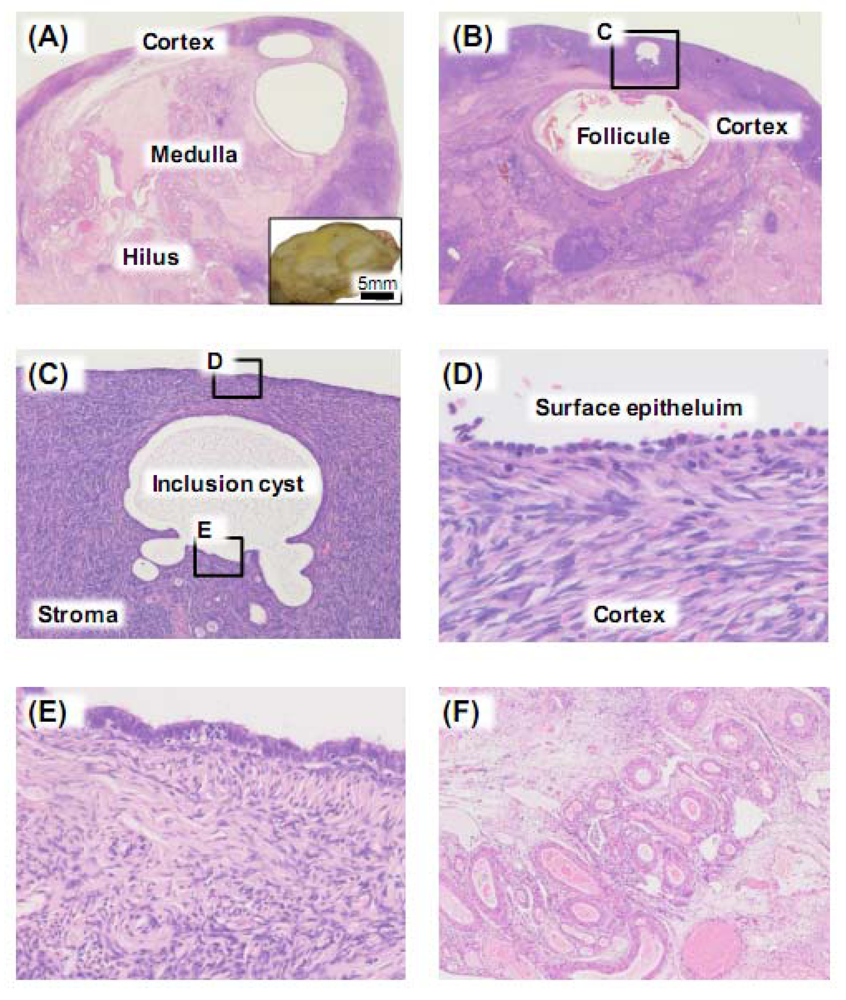
2. Constituent Cells of the Normal Adult Ovary

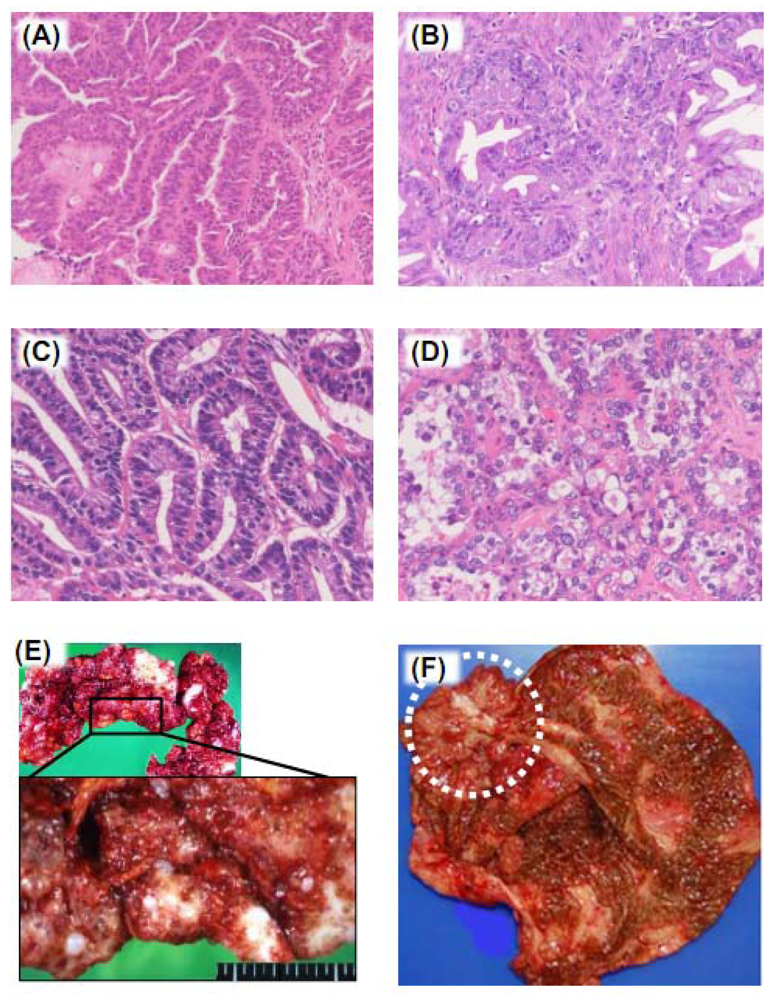
3. Pathogenesis of Ovarian Cancers

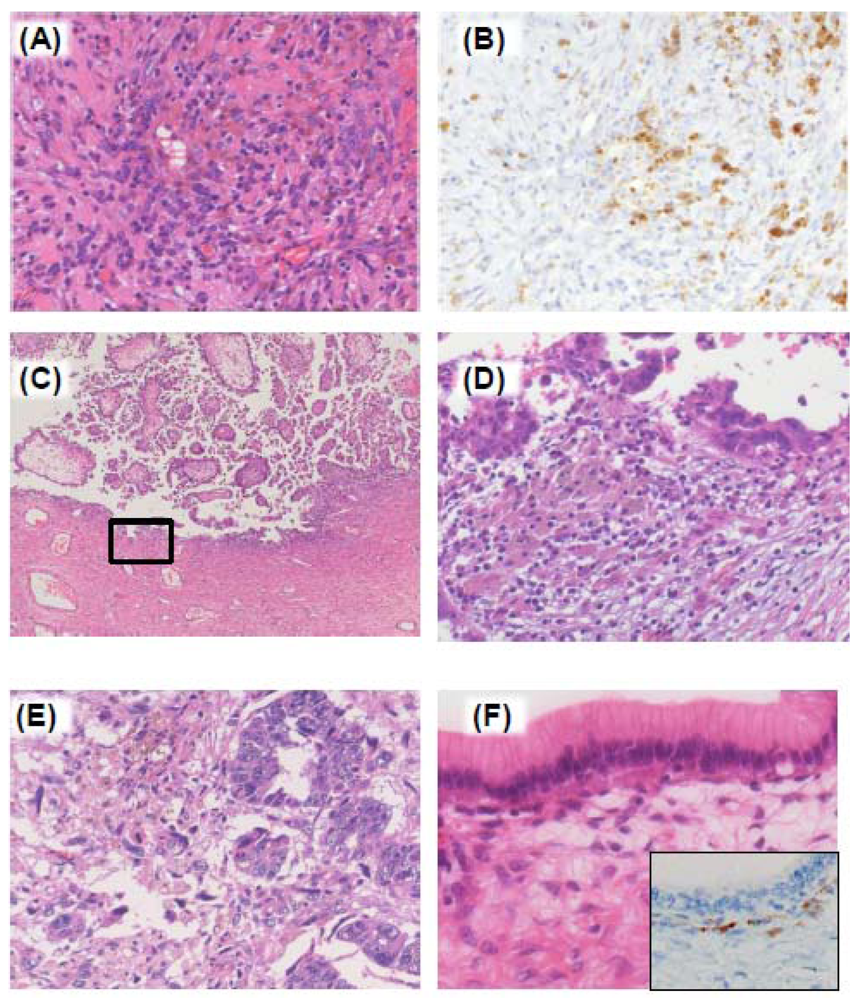
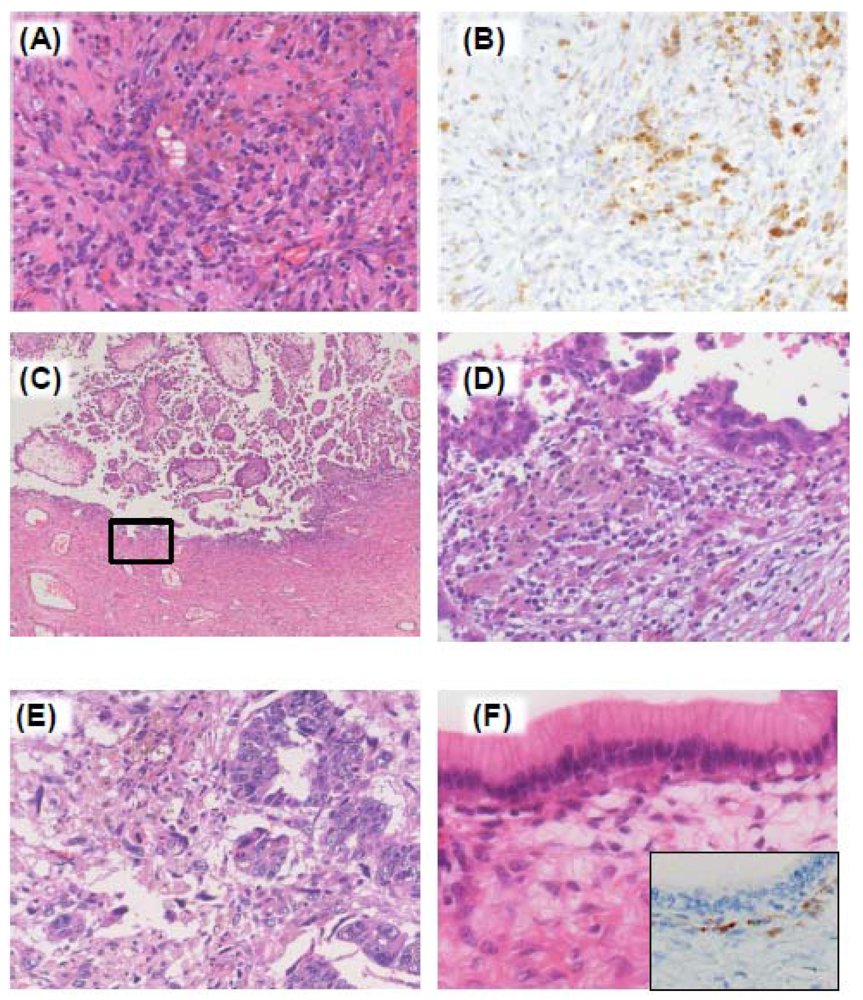
4. Endometriosis-Associated Ovarian Cancer (EAOC)
5. Roles of Stroma for the Development of Ovarian Cancers
5.1. Extracellular Matrix (ECM)
5.2. Mesothelium
5.3. Omental Adipose Tissue
5.4. Malignant Stroma and Functioning Stroma
6. Key Signaling Molecules Involved in Proinflammatory Microenvironment of Ovarian Cancers
6.1. CXC Chemokines
6.2. CC Chemokines
6.3. Other Molecules
7. Conclusions
Acknowledgments
References
- Cannistra, S.A. Cancer of the ovary. N. Engl. J. Med. 1993, 329, 1550–1559. [Google Scholar] [CrossRef]
- Berkenblit, A.; Cannistra, S.A. Advances in the management of epithelial ovarian cancer. J. Reprod. Med. 2005, 50, 426–438. [Google Scholar]
- Shimizu, M.; Toki, T.; Takagi, Y.; Konishi, I.; Fujii, S. Immunohistochemical detection of the Wilms’ tumor gene (WT1) in epithelial ovarian tumors. Int. J. Gynecol. Pathol. 2000, 19, 158–163. [Google Scholar] [CrossRef]
- Auersperg, N.; Wong, A.S.; Choi, K.C.; Kang, S.K.; Leung, P.C. Ovarian surface epithelium: Biology, endocrinology, and pathology. Endocr. Rev. 2001, 22, 255–288. [Google Scholar] [CrossRef]
- Cai, K.Q.; Caslini, C.; Capo-chichi, C.D.; Slater, C.; Smith, E.R.; Wu, H.; Klein-Szanto, A.J.; Godwin, A.K.; Xu, X.X. Loss of GATA4 and GATA6 expression specifies ovarian cancer histological subtypes and precedes neoplastic transformation of ovarian surface epithelia. PLoS One 2009, 4, e6454. [Google Scholar]
- Sundfeldt, K.; Ivarsson, K.; Carlsson, M.; Enerback, S.; Janson, P.O.; Brannstrom, M.; Hedin, L. The expression of CCAAT/enhancer binding protein (C/EBP) in the human ovary in vivo: Specific increase in C/EBPbeta during epithelial tumour progression. Br. J. Cancer 1999, 79, 1240–1248. [Google Scholar] [CrossRef]
- Gendronneau, G.; Boucherat, O.; Aubin, J.; Lemieux, M.; Jeannotte, L. The loss of Hoxa5 function causes estrous acyclicity and ovarian epithelial inclusion cysts. Endocrinology 2012, 153, 1484–1497. [Google Scholar] [CrossRef]
- Bell, D.A. Origins and molecular pathology of ovarian cancer. Mod. Pathol. 2005, 18, S19–S32. [Google Scholar] [CrossRef]
- Kurman, R.J.; Shih Ie, M. Pathogenesis of ovarian cancer: Lessons from morphology and molecular biology and their clinical implications. Int. J. Gynecol. Pathol. 2008, 27, 151–160. [Google Scholar]
- Smith Sehdev, A.E.; Sehdev, P.S.; Kurman, R.J. Noninvasive and invasive micropapillary (low-grade) serous carcinoma of the ovary: A clinicopathologic analysis of 135 cases. Am. J. Surg. Pathol. 2003, 27, 725–736. [Google Scholar] [CrossRef]
- Singer, G.; Stohr, R.; Cope, L.; Dehari, R.; Hartmann, A.; Cao, D.F.; Wang, T.L.; Kurman, R.J.; Shih Ie, M. Patterns of p53 mutations separate ovarian serous borderline tumors and low- and high-grade carcinomas and provide support for a new model of ovarian carcinogenesis: A mutational analysis with immunohistochemical correlation. Am. J. Surg. Pathol. 2005, 29, 218–224. [Google Scholar] [CrossRef]
- Singer, G.; Oldt, R., 3rd.; Cohen, Y.; Wang, B.G.; Sidransky, D.; Kurman, R.J.; Shih Ie, M. Mutations in BRAF and KRAS characterize the development of low-grade ovarian serous carcinoma. J. Natl. Cancer Inst. 2003, 95, 484–486. [Google Scholar] [CrossRef]
- Dehari, R.; Kurman, R.J.; Logani, S.; Shih Ie, M. The development of high-grade serous carcinoma from atypical proliferative (borderline) serous tumors and low-grade micropapillary serous carcinoma: A morphologic and molecular genetic analysis. Am. J. Surg. Pathol. 2007, 31, 1007–1012. [Google Scholar] [CrossRef]
- Medeiros, F.; Muto, M.G.; Lee, Y.; Elvin, J.A.; Callahan, M.J.; Feltmate, C.; Garber, J.E.; Cramer, D.W.; Crum, C.P. The tubal fimbria is a preferred site for early adenocarcinoma in women with familial ovarian cancer syndrome. Am. J. Surg. Pathol. 2006, 30, 230–236. [Google Scholar] [CrossRef]
- Kindelberger, D.W.; Lee, Y.; Miron, A.; Hirsch, M.S.; Feltmate, C.; Medeiros, F.; Callahan, M.J.; Garner, E.O.; Gordon, R.W.; Birch, C.; et al. Intraepithelial carcinoma of the fimbria and pelvic serous carcinoma: Evidence for a causal relationship. Am. J. Surg. Pathol. 2007, 31, 161–169. [Google Scholar] [CrossRef]
- Przybycin, C.G.; Kurman, R.J.; Ronnett, B.M.; Shih Ie, M.; Vang, R. Are all pelvic (nonuterine) serous carcinomas of tubal origin? Am. J. Surg. Pathol. 2010, 34, 1407–1416. [Google Scholar] [CrossRef]
- Kurman, R.J.; Vang, R.; Junge, J.; Hannibal, C.G.; Kjaer, S.K.; Shih Ie, M. Papillary tubal hyperplasia: The putative precursor of ovarian atypical proliferative (borderline) serous tumors, noninvasive implants, and endosalpingiosis. Am. J. Surg. Pathol. 2011, 35, 1605–1614. [Google Scholar] [CrossRef]
- Prat, J.; Ribe, A.; Gallardo, A. Hereditary ovarian cancer. Hum. Pathol. 2005, 36, 861–870. [Google Scholar] [CrossRef]
- Press, J.Z.; de Luca, A.; Boyd, N.; Young, S.; Troussard, A.; Ridge, Y.; Kaurah, P.; Kalloger, S.E.; Blood, K.A.; Smith, M.; et al. Ovarian carcinomas with genetic and epigenetic BRCA1 loss have distinct molecular abnormalities. BMC Cancer 2008, 8, 17. [Google Scholar] [CrossRef]
- Carcangiu, M.L.; Peissel, B.; Pasini, B.; Spatti, G.; Radice, P.; Manoukian, S. Incidental carcinomas in prophylactic specimens in BRCA1 and BRCA2 germ-line mutation carriers, with emphasis on fallopian tube lesions: Report of 6 cases and review of the literature. Am. J. Surg. Pathol. 2006, 30, 1222–1230. [Google Scholar] [CrossRef]
- Young, R.H. From Krukenberg to today: The ever present problems posed by metastatic tumors in the ovary. Part II. Adv. Anat. Pathol. 2007, 14, 149–177. [Google Scholar] [CrossRef]
- Ronnett, B.M.; Kurman, R.J.; Shmookler, B.M.; Sugarbaker, P.H.; Young, R.H. The morphologic spectrum of ovarian metastases of appendiceal adenocarcinomas: A clinicopathologic and immunohistochemical analysis of tumors often misinterpreted as primary ovarian tumors or metastatic tumors from other gastrointestinal sites. Am. J. Surg. Pathol. 1997, 21, 1144–1155. [Google Scholar] [CrossRef]
- Logani, S.; Oliva, E.; Arnell, P.M.; Amin, M.B.; Young, R.H. Use of novel immunohistochemical markers expressed in colonic adenocarcinoma to distinguish primary ovarian tumors from metastatic colorectal carcinoma. Mod. Pathol. 2005, 18, 19–25. [Google Scholar] [CrossRef]
- Vang, R.; Gown, A.M.; Barry, T.S.; Wheeler, D.T.; Yemelyanova, A.; Seidman, J.D.; Ronnett, B.M. Cytokeratins 7 and 20 in primary and secondary mucinous tumors of the ovary: Analysis of coordinate immunohistochemical expression profiles and staining distribution in 179 cases. Am. J. Surg. Pathol. 2006, 30, 1130–1139. [Google Scholar]
- Vang, R.; Gown, A.M.; Wu, L.S.; Barry, T.S.; Wheeler, D.T.; Yemelyanova, A.; Seidman, J.D.; Ronnett, B.M. Immunohistochemical expression of CDX2 in primary ovarian mucinous tumors and metastatic mucinous carcinomas involving the ovary: Comparison with CK20 and correlation with coordinate expression of CK7. Mod. Pathol. 2006, 19, 1421–1428. [Google Scholar]
- Enomoto, T.; Weghorst, C.M.; Inoue, M.; Tanizawa, O.; Rice, J.M. K-ras activation occurs frequently in mucinous adenocarcinomas and rarely in other common epithelial tumors of the human ovary. Am. J. Pathol. 1991, 139, 777–785. [Google Scholar]
- Ichikawa, Y.; Nishida, M.; Suzuki, H.; Yoshida, S.; Tsunoda, H.; Kubo, T.; Uchida, K.; Miwa, M. Mutation of K-ras protooncogene is associated with histological subtypes in human mucinous ovarian tumors. Cancer Res. 1994, 54, 33–35. [Google Scholar]
- Gemignani, M.L.; Schlaerth, A.C.; Bogomolniy, F.; Barakat, R.R.; Lin, O.; Soslow, R.; Venkatraman, E.; Boyd, J. Role of KRAS and BRAF gene mutations in mucinous ovarian carcinoma. Gynecol. Oncol. 2003, 90, 378–381. [Google Scholar] [CrossRef]
- Catasus, L.; Bussaglia, E.; Rodrguez, I.; Gallardo, A.; Pons, C.; Irving, J.A.; Prat, J. Molecular genetic alterations in endometrioid carcinomas of the ovary: Similar frequency of beta-catenin abnormalities but lower rate of microsatellite instability and PTEN alterations than in uterine endometrioid carcinomas. Hum. Pathol. 2004, 35, 1360–1368. [Google Scholar] [CrossRef]
- Obata, K.; Morland, S.J.; Watson, R.H.; Hitchcock, A.; Chenevix-Trench, G.; Thomas, E.J.; Campbell, I.G. Frequent PTEN/MMAC mutations in endometrioid but not serous or mucinous epithelial ovarian tumors. Cancer Res. 1998, 58, 2095–2097. [Google Scholar]
- Gamallo, C.; Palacios, J.; Moreno, G.; Calvo de Mora, J.; Suarez, A.; Armas, A. beta-Catenin expression pattern in stage I and II ovarian carcinomas : Relationship with beta-catenin gene mutations, clinicopathological features, and clinical outcome. Am. J. Pathol. 1999, 155, 527–536. [Google Scholar] [CrossRef]
- Moreno-Bueno, G.; Gamallo, C.; Perez-Gallego, L.; de Mora, J.C.; Suarez, A.; Palacios, J. beta-Catenin expression pattern, beta-catenin gene mutations, and microsatellite instability in endometrioid ovarian carcinomas and synchronous endometrial carcinomas. Diagn. Mol. Pathol. 2001, 10, 116–122. [Google Scholar] [CrossRef]
- Jones, S.; Wang, T.L.; Shih Ie, M.; Mao, T.L.; Nakayama, K.; Roden, R.; Glas, R.; Slamon, D.; Diaz, L.A., Jr.; Vogelstein, B.; et al. Frequent mutations of chromatin remodeling gene ARID1A in ovarian clear cell carcinoma. Science 2010, 330, 228–231. [Google Scholar]
- Wiegand, K.C.; Shah, S.P.; Al-Agha, O.M.; Zhao, Y.; Tse, K.; Zeng, T.; Senz, J.; McConechy, M.K.; Anglesio, M.S.; Kalloger, S.E.; et al. ARID1A mutations in endometriosis-associated ovarian carcinomas. N. Engl. J. Med. 2010, 363, 1532–1543. [Google Scholar]
- Roth, L.M.; Talerman, A. Recent advances in the pathology and classification of ovarian germ cell tumors. Int. J. Gynecol. Pathol. 2006, 25, 305–320. [Google Scholar] [CrossRef]
- Baker, P.M.; Oliva, E. Immunohistochemistry as a tool in the differential diagnosis of ovarian tumors: An update. Int. J. Gynecol. Pathol. 2005, 24, 39–55. [Google Scholar]
- Shah, S.P.; Kobel, M.; Senz, J.; Morin, R.D.; Clarke, B.A.; Wiegand, K.C.; Leung, G.; Zayed, A.; Mehl, E.; Kalloger, S.E.; et al. Mutation of FOXL2 in granulosa-cell tumors of the ovary. N. Engl. J. Med. 2009, 360, 2719–2729. [Google Scholar] [CrossRef]
- Kim, M.S.; Hur, S.Y.; Yoo, N.J.; Lee, S.H. Mutational analysis of FOXL2 codon 134 in granulosa cell tumour of ovary and other human cancers. J. Pathol. 2010, 221, 147–152. [Google Scholar] [CrossRef]
- Modesitt, S.C.; Tortolero-Luna, G.; Robinson, J.B.; Gershenson, D.M.; Wolf, J.K. Ovarian and extraovarian endometriosis-associated cancer. Obstet. Gynecol. 2002, 100, 788–795. [Google Scholar] [CrossRef]
- Koninckx, P.R.; Barlow, D.; Kennedy, S. Implantation versus infiltration: The Sampson versus the endometriotic disease theory. Gynecol. Obstet. Invest. 1999, 47, 3–10. [Google Scholar] [CrossRef]
- Suginami, H. A reappraisal of the coelomic metaplasia theory by reviewing endometriosis occurring in unusual sites and instances. Am. J. Obstet. Gynecol. 1991, 165, 214–218. [Google Scholar]
- Fujii, S. Secondary mullerian system and endometriosis. Am. J. Obstet. Gynecol. 1991, 165, 219–225. [Google Scholar]
- Bergqvist, A.; Bruse, C.; Carlberg, M.; Carlstrom, K. Interleukin 1beta, interleukin-6, and tumor necrosis factor-alpha in endometriotic tissue and in endometrium. Fertil. Steril. 2001, 75, 489–495. [Google Scholar] [CrossRef]
- Ota, H.; Igarashi, S.; Hatazawa, J.; Tanaka, T. Distribution of heat shock proteins in eutopic and ectopic endometrium in endometriosis and adenomyosis. Fertil. Steril. 1997, 68, 23–28. [Google Scholar]
- Gallinelli, A.; Chiossi, G.; Giannella, L.; Marsella, T.; Genazzani, A.D.; Volpe, A. Different concentrations of interleukins in the peritoneal fluid of women with endometriosis: Relationships with lymphocyte subsets. Gynecol. Endocrinol. 2004, 18, 144–151. [Google Scholar] [CrossRef]
- Cakmak, H.; Guzeloglu-Kayisli, O.; Kayisli, U.A.; Arici, A. Immune-endocrine interactions in endometriosis. Front. Biosci. (Elite Ed.) 2009, 1, 429–443. [Google Scholar]
- Koninckx, P.R.; Kennedy, S.H.; Barlow, D.H. Endometriotic disease: The role of peritoneal fluid. Hum. Reprod. Update 1998, 4, 741–751. [Google Scholar] [CrossRef]
- Sikora, J.; Mielczarek-Palacz, A.; Kondera-Anasz, Z. Role of natural killer cell activity in the pathogenesis of endometriosis. Curr. Med. Chem. 2011, 18, 200–208. [Google Scholar] [CrossRef]
- Szyllo, K.; Tchorzewski, H.; Banasik, M.; Glowacka, E.; Lewkowicz, P.; Kamer-Bartosinska, A. The involvement of T lymphocytes in the pathogenesis of endometriotic tissues overgrowth in women with endometriosis. Mediators Inflamm. 2003, 12, 131–138. [Google Scholar] [CrossRef]
- Stern, R.C.; Dash, R.; Bentley, R.C.; Snyder, M.J.; Haney, A.F.; Robboy, S.J. Malignancy in endometriosis: Frequency and comparison of ovarian and extraovarian types. Int. J. Gynecol. Pathol. 2001, 20, 133–139. [Google Scholar] [CrossRef]
- Jimbo, H.; Yoshikawa, H.; Onda, T.; Yasugi, T.; Sakamoto, A.; Taketani, Y. Prevalence of ovarian endometriosis in epithelial ovarian cancer. Int. J. Gynaecol. Obstet. 1997, 59, 245–250. [Google Scholar] [CrossRef]
- Kobayashi, H.; Sumimoto, K.; Moniwa, N.; Imai, M.; Takakura, K.; Kuromaki, T.; Morioka, E.; Arisawa, K.; Terao, T. Risk of developing ovarian cancer among women with ovarian endometrioma: A cohort study in Shizuoka, Japan. Int. J. Gynecol. Cancer 2007, 17, 37–43. [Google Scholar] [CrossRef]
- Melin, A.; Sparen, P.; Persson, I.; Bergqvist, A. Endometriosis and the risk of cancer with special emphasis on ovarian cancer. Hum. Reprod. 2006, 21, 1237–1242. [Google Scholar] [CrossRef]
- Fukunaga, M.; Nomura, K.; Ishikawa, E.; Ushigome, S. Ovarian atypical endometriosis: Its close association with malignant epithelial tumours. Histopathology 1997, 30, 249–255. [Google Scholar]
- Zhao, C.; Wu, L.S.; Barner, R. Pathogenesis of ovarian clear cell adenofibroma, atypical proliferative (borderline) tumor, and carcinoma: Clinicopathologic features of tumors with endometriosis or adenofibromatous components support two related pathways of tumor development. J. Cancer 2011, 2, 94–106. [Google Scholar]
- Jiang, X.; Hitchcock, A.; Bryan, E.J.; Watson, R.H.; Englefield, P.; Thomas, E.J.; Campbell, I.G. Microsatellite analysis of endometriosis reveals loss of heterozygosity at candidate ovarian tumor suppressor gene loci. Cancer Res. 1996, 56, 3534–3539. [Google Scholar]
- Jimbo, H.; Hitomi, Y.; Yoshikawa, H.; Yano, T.; Momoeda, M.; Sakamoto, A.; Tsutsumi, O.; Taketani, Y.; Esumi, H. Evidence for monoclonal expansion of epithelial cells in ovarian endometrial cysts. Am. J. Pathol. 1997, 150, 1173–1178. [Google Scholar]
- Sato, N.; Tsunoda, H.; Nishida, M.; Morishita, Y.; Takimoto, Y.; Kubo, T.; Noguchi, M. Loss of heterozygosity on 10q23.3 and mutation of the tumor suppressor gene PTEN in benign endometrial cyst of the ovary: Possible sequence progression from benign endometrial cyst to endometrioid carcinoma and clear cell carcinoma of the ovary. Cancer Res. 2000, 60, 7052–7056. [Google Scholar]
- Xouri, G.; Christian, S. Origin and function of tumor stroma fibroblasts. Semin. Cell Dev. Biol. 2010, 21, 40–46. [Google Scholar]
- Ishiguro, K.; Yoshida, T.; Yagishita, H.; Numata, Y.; Okayasu, T. Epithelial and stromal genetic instability contributes to genesis of colorectal adenomas. Gut 2006, 55, 695–702. [Google Scholar] [CrossRef]
- Kurose, K.; Gilley, K.; Matsumoto, S.; Watson, P.H.; Zhou, X.P.; Eng, C. Frequent somatic mutations in PTEN and TP53 are mutually exclusive in the stroma of breast carcinomas. Nat. Genet. 2002, 32, 355–357. [Google Scholar] [CrossRef]
- Moinfar, F.; Man, Y.G.; Arnould, L.; Bratthauer, G.L.; Ratschek, M.; Tavassoli, F.A. Concurrent and independent genetic alterations in the stromal and epithelial cells of mammary carcinoma: Implications for tumorigenesis. Cancer Res. 2000, 60, 2562–2566. [Google Scholar]
- Corver, W.E.; Ter Haar, N.T.; Fleuren, G.J.; Oosting, J. Cervical carcinoma-associated fibroblasts are DNA diploid and do not show evidence for somatic genetic alterations. Cell. Oncol. (Dordr.) 2011, 34, 553–563. [Google Scholar]
- Orimo, A.; Gupta, P.B.; Sgroi, D.C.; Arenzana-Seisdedos, F.; Delaunay, T.; Naeem, R.; Carey, V.J.; Richardson, A.L.; Weinberg, R.A. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 2005, 121, 335–348. [Google Scholar] [CrossRef]
- Tuhkanen, H.; Anttila, M.; Kosma, V.M.; Yla-Herttuala, S.; Heinonen, S.; Kuronen, A.; Juhola, M.; Tammi, R.; Tammi, M.; Mannermaa, A. Genetic alterations in the peritumoral stromal cells of malignant and borderline epithelial ovarian tumors as indicated by allelic imbalance on chromosome 3p. Int. J. Cancer 2004, 109, 247–252. [Google Scholar] [CrossRef]
- Tuhkanen, H.; Anttila, M.; Kosma, V.M.; Heinonen, S.; Juhola, M.; Helisalmi, S.; Kataja, V.; Mannermaa, A. Frequent gene dosage alterations in stromal cells of epithelial ovarian carcinomas. Int. J. Cancer 2006, 119, 1345–1353. [Google Scholar]
- Qiu, W.; Hu, M.; Sridhar, A.; Opeskin, K.; Fox, S.; Shipitsin, M.; Trivett, M.; Thompson, E.R.; Ramakrishna, M.; Gorringe, K.L.; et al. No evidence of clonal somatic genetic alterations in cancer-associated fibroblasts from human breast and ovarian carcinomas. Nat. Genet. 2008, 40, 650–655. [Google Scholar]
- Cai, J.; Tang, H.; Xu, L.; Wang, X.; Yang, C.; Ruan, S.; Guo, J.; Hu, S.; Wang, Z. Fibroblasts in omentum activated by tumor cells promote ovarian cancer growth, adhesion and invasiveness. Carcinogenesis 2012, 33, 20–29. [Google Scholar] [CrossRef]
- Yamamura, S.; Matsumura, N.; Mandai, M.; Huang, Z.; Oura, T.; Baba, T.; Hamanishi, J.; Yamaguchi, K.; Kang, H.S.; Okamoto, T.; et al. The activated transforming growth factor-beta signaling pathway in peritoneal metastases is a potential therapeutic target in ovarian cancer. Int. J. Cancer 2012, 130, 20–28. [Google Scholar] [CrossRef]
- Skubitz, A.P.; Bast, R.C., Jr.; Wayner, E.A.; Letourneau, P.C.; Wilke, M.S. Expression of alpha 6 and beta 4 integrins in serous ovarian carcinoma correlates with expression of the basement membrane protein laminin. Am. J. Pathol. 1996, 148, 1445–1461. [Google Scholar]
- Capo-Chichi, C.D.; Smith, E.R.; Yang, D.H.; Roland, I.H.; Vanderveer, L.; Cohen, C.; Hamilton, T.C.; Godwin, A.K.; Xu, X.X. Dynamic alterations of the extracellular environment of ovarian surface epithelial cells in premalignant transformation, tumorigenicity, and metastasis. Cancer 2002, 95, 1802–1815. [Google Scholar]
- Kato, N.; Takeda, J.; Fukase, M.; Motoyama, T. Alternate mucoid and hyalinized stroma in clear cell carcinoma of the ovary: Manifestation of serial stromal remodeling. Mod. Pathol. 2010, 23, 881–888. [Google Scholar] [CrossRef]
- Ballas, S.L.; Veltri, E.P. The efficacy of amiodarone in the treatment of refractory nonsustained ventricular tachycardia. Md. Med. J. 1991, 40, 475–478. [Google Scholar]
- Hibbs, K.; Skubitz, K.M.; Pambuccian, S.E.; Casey, R.C.; Burleson, K.M.; Oegema, T.R., Jr.; Thiele, J.J.; Grindle, S.M.; Bliss, R.L.; Skubitz, A.P. Differential gene expression in ovarian carcinoma: Identification of potential biomarkers. Am. J. Pathol. 2004, 165, 397–414. [Google Scholar] [CrossRef]
- Mercurio, A.M.; Rabinovitz, I. Towards a mechanistic understanding of tumor invasion—Lessons from the alpha6beta 4 integrin. Semin. Cancer Biol. 2001, 11, 129–141. [Google Scholar] [CrossRef]
- Natali, P.G.; Nicotra, M.R.; Bartolazzi, A.; Cavaliere, R.; Bigotti, A. Integrin expression in cutaneous malignant melanoma: Association of the alpha 3/beta 1 heterodimer with tumor progression. Int. J. Cancer 1993, 54, 68–72. [Google Scholar] [CrossRef]
- Hashida, H.; Takabayashi, A.; Tokuhara, T.; Taki, T.; Kondo, K.; Kohno, N.; Yamaoka, Y.; Miyake, M. Integrin alpha3 expression as a prognostic factor in colon cancer: Association with MRP-1/CD9 and KAI1/CD82. Int. J. Cancer 2002, 97, 518–525. [Google Scholar] [CrossRef]
- Kazarov, A.R.; Yang, X.; Stipp, C.S.; Sehgal, B.; Hemler, M.E. An extracellular site on tetraspanin CD151 determines alpha 3 and alpha 6 integrin-dependent cellular morphology. J. Cell Biol. 2002, 158, 1299–1309. [Google Scholar] [CrossRef]
- Furuya, M.; Kato, H.; Nishimura, N.; Ishiwata, I.; Ikeda, H.; Ito, R.; Yoshiki, T. Down-regulation of CD9 in human ovarian carcinoma cell might contribute to peritoneal dissemination: Morphologic alteration and reduced expression of beta1 integrin subsets. Cancer Res. 2005, 65, 2617–2625. [Google Scholar] [CrossRef]
- Hasegawa, M.; Furuya, M.; Kasuya, Y.; Nishiyama, M.; Sugiura, T.; Nikaido, T.; Momota, Y.; Ichinose, M.; Kimura, S. CD151 dynamics in carcinoma-stroma interaction: Integrin expression, adhesion strength and proteolytic activity. Lab. Invest. 2007, 87, 882–892. [Google Scholar] [CrossRef]
- Furuya, M.; Ishikura, H.; Nemori, R.; Shibata, M.; Fujimoto, S.; Yoshiki, T. Clarification of the active gelatinolytic sites in human ovarian neoplasms using in situ zymography. Hum. Pathol. 2001, 32, 163–168. [Google Scholar] [CrossRef]
- Brun, J.L.; Cortez, A.; Commo, F.; Uzan, S.; Rouzier, R.; Darai, E. Serous and mucinous ovarian tumors express different profiles of MMP-2, -7, -9, MT1-MMP, and TIMP-1 and -2. Int. J. Oncol. 2008, 33, 1239–1246. [Google Scholar]
- Adley, B.P.; Gleason, K.J.; Yang, X.J.; Stack, M.S. Expression of membrane type 1 matrix metalloproteinase (MMP-14) in epithelial ovarian cancer: High level expression in clear cell carcinoma. Gynecol. Oncol. 2009, 112, 319–324. [Google Scholar] [CrossRef]
- Manenti, L.; Paganoni, P.; Floriani, I.; Landoni, F.; Torri, V.; Buda, A.; Taraboletti, G.; Labianca, R.; Belotti, D.; Giavazzi, R. Expression levels of vascular endothelial growth factor, matrix metalloproteinases 2 and 9 and tissue inhibitor of metalloproteinases 1 and 2 in the plasma of patients with ovarian carcinoma. Eur. J. Cancer 2003, 39, 1948–1956. [Google Scholar] [CrossRef]
- Furuya, M.; Ishikura, H.; Kawarada, Y.; Ogawa, Y.; Sakuragi, N.; Fujimoto, S.; Yoshiki, T. Expression of matrix metalloproteinases and related tissue inhibitors in the cyst fluids of ovarian mucinous neoplasms. Gynecol. Oncol. 2000, 78, 106–112. [Google Scholar] [CrossRef]
- Belotti, D.; Paganoni, P.; Manenti, L.; Garofalo, A.; Marchini, S.; Taraboletti, G.; Giavazzi, R. Matrix metalloproteinases (MMP9 and MMP2) induce the release of vascular endothelial growth factor (VEGF) by ovarian carcinoma cells: Implications for ascites formation. Cancer Res. 2003, 63, 5224–5229. [Google Scholar]
- Furuya, M.; Ishikura, H.; Ogawa, Y.; Kawarada, Y.; Sakuragi, N.; Fujimoto, S.; Yoshiki, T. Analyses of matrix metalloproteinases and their inhibitors in cyst fluid of serous ovarian tumors. Pathobiology 2000, 68, 239–244. [Google Scholar] [CrossRef]
- Chang, K.; Pastan, I. Molecular cloning of mesothelin, a differentiation antigen present on mesothelium, mesotheliomas, and ovarian cancers. Proc. Natl. Acad. Sci. USA 1996, 93, 136–140. [Google Scholar] [CrossRef]
- Ho, M.; Hassan, R.; Zhang, J.; Wang, Q.C.; Onda, M.; Bera, T.; Pastan, I. Humoral immune response to mesothelin in mesothelioma and ovarian cancer patients. Clin. Cancer Res. 2005, 11, 3814–3820. [Google Scholar]
- Wang, K.; Bodempudi, V.; Liu, Z.; Borrego-Diaz, E.; Yamoutpoor, F.; Meyer, A.; Woo, R.A.; Pan, W.; Dudek, A.Z.; Olyaee, M.S.; et al. Inhibition of mesothelin as a novel strategy for targeting cancer cells. PLoS One 2012, 7, e33214. [Google Scholar]
- Hassan, R.; Bera, T.; Pastan, I. Mesothelin: A new target for immunotherapy. Clin. Cancer Res. 2004, 10, 3937–3942. [Google Scholar] [CrossRef]
- Rump, A.; Morikawa, Y.; Tanaka, M.; Minami, S.; Umesaki, N.; Takeuchi, M.; Miyajima, A. Binding of ovarian cancer antigen CA125/MUC16 to mesothelin mediates cell adhesion. J. Biol Chem 2004, 279, 9190–9198. [Google Scholar]
- Kaneko, O.; Gong, L.; Zhang, J.; Hansen, J.K.; Hassan, R.; Lee, B.; Ho, M. A binding domain on mesothelin for CA125/MUC16. J. Biol. Chem. 2009, 284, 3739–3749. [Google Scholar]
- Hassan, R.; Cohen, S.J.; Phillips, M.; Pastan, I.; Sharon, E.; Kelly, R.J.; Schweizer, C.; Weil, S.; Laheru, D. Phase I clinical trial of the chimeric anti-mesothelin monoclonal antibody MORAb-009 in patients with mesothelin-expressing cancers. Clin. Cancer Res. 2010, 16, 6132–6138. [Google Scholar]
- Nieman, K.M.; Kenny, H.A.; Penicka, C.V.; Ladanyi, A.; Buell-Gutbrod, R.; Zillhardt, M.R.; Romero, I.L.; Carey, M.S.; Mills, G.B.; Hotamisligil, G.S.; et al. Adipocytes promote ovarian cancer metastasis and provide energy for rapid tumor growth. Nat. Med. 2011, 17, 1498–1503. [Google Scholar]
- Fujii, H.; Yoshida, M.; Gong, Z.X.; Matsumoto, T.; Hamano, Y.; Fukunaga, M.; Hruban, R.H.; Gabrielson, E.; Shirai, T. Frequent genetic heterogeneity in the clonal evolution of gynecological carcinosarcoma and its influence on phenotypic diversity. Cancer Res. 2000, 60, 114–120. [Google Scholar]
- Moinfar, F.; Kremser, M.L.; Man, Y.G.; Zatloukal, K.; Tavassoli, F.A.; Denk, H. Allelic imbalances in endometrial stromal neoplasms: Frequent genetic alterations in the nontumorous normal-appearing endometrial and myometrial tissues. Gynecol. Oncol. 2004, 95, 662–671. [Google Scholar] [CrossRef]
- Wernli, K.J.; Newcomb, P.A.; Hampton, J.M.; Trentham-Dietz, A.; Egan, K.M. Hormone therapy and ovarian cancer: Incidence and survival. Cancer Causes Control 2008, 19, 605–613. [Google Scholar] [CrossRef]
- Badia, E.; Docquier, A.; Busson, M.; Lapierre, M.; Pujol, P.; Balaguer, P.; Cavailles, V. Long-term treatment with the pure anti-estrogen fulvestrant durably remodels estrogen signaling in BG-1 ovarian cancer cells. J. Steroid Biochem. Mol. Biol. 2012, 132, 176–185. [Google Scholar] [CrossRef]
- Curiel, T.J.; Coukos, G.; Zou, L.; Alvarez, X.; Cheng, P.; Mottram, P.; Evdemon-Hogan, M.; Conejo-Garcia, J.R.; Zhang, L.; Burow, M.; et al. Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 2004, 10, 942–949. [Google Scholar]
- Wolf, D.; Wolf, A.M.; Rumpold, H.; Fiegl, H.; Zeimet, A.; GMuller-Holzner, E.; Deibl, M.; Gastl, G.; Gunsilius, E.; Marth, C. The expression of the regulatory T cell-specific forkhead box transcription factor FoxP3 is associated with poor prognosis in ovarian cancer. Clin. Cancer Res. 2005, 11, 8326–8331. [Google Scholar]
- Milliken, D.; Scotton, C.; Raju, S.; Balkwill, F.; Wilson, J. Analysis of chemokines and chemokine receptor expression in ovarian cancer ascites. Clin. Cancer Res. 2002, 8, 1108–1114. [Google Scholar]
- Moradi, M.M.; Carson, L.F.; Weinberg, B.; Haney, A.F.; Twiggs, L.B.; Ramakrishnan, S. Serum and ascitic fluid levels of interleukin-1, interleukin-6, and tumor necrosis factor-alpha in patients with ovarian epithelial cancer. Cancer 1993, 72, 2433–2440. [Google Scholar]
- Mustea, A.; Pirvulescu, C.; Konsgen, D.; Braicu, E.I.; Yuan, S.; Sun, P.; Lichtenegger, W.; Sehouli, J. Decreased IL-1 RA concentration in ascites is associated with a significant improvement in overall survival in ovarian cancer. Cytokine 2008, 42, 77–84. [Google Scholar] [CrossRef]
- Zeimet, A.G.; Widschwendter, M.; Knabbe, C.; Fuchs, D.; Herold, M.; Muller-Holzner, E.; Daxenbichler, G.; Offner, F.A.; Dapunt, O.; Marth, C. Ascitic interleukin-12 is an independent prognostic factor in ovarian cancer. J. Clin. Oncol. 1998, 16, 1861–1868. [Google Scholar]
- Arya, M.; Patel, H.R.; Williamson, M. Chemokines: Key players in cancer. Curr. Med. Res. Opin. 2003, 19, 557–564. [Google Scholar] [CrossRef]
- Ben-Baruch, A. The multifaceted roles of chemokines in malignancy. Cancer Metastasis Rev. 2006, 25, 357–371. [Google Scholar] [CrossRef]
- Arenberg, D.A.; Keane, M.P.; DiGiovine, B.; Kunkel, S.L.; Morris, S.B.; Xue, Y.Y.; Burdick, M.D.; Glass, M.C.; Iannettoni, M.D.; Strieter, R.M. Epithelial-neutrophil activating peptide (ENA-78) is an important angiogenic factor in non-small cell lung cancer. J. Clin. Invest. 1998, 102, 465–472. [Google Scholar] [CrossRef]
- Vicari, A.P.; Caux, C. Chemokines in cancer. Cytokine Growth Factor Rev. 2002, 13, 143–154. [Google Scholar] [CrossRef]
- Frederick, M.J.; Clayman, G.L. Chemokines in cancer. Expert Rev. Mol. Med. 2001, 3, 1–18. [Google Scholar] [CrossRef]
- Agarwal, A.; Tressel, S.L.; Kaimal, R.; Balla, M.; Lam, F.H.; Covic, L.; Kuliopulos, A. Identification of a metalloprotease-chemokine signaling system in the ovarian cancer microenvironment: Implications for antiangiogenic therapy. Cancer Res. 2010, 70, 5880–5890. [Google Scholar]
- Schutyser, E.; Struyf, S.; Proost, P.; Opdenakker, G.; Laureys, G.; Verhasselt, B.; Peperstraete, L.; van de Putte, I.; Saccani, A.; Allavena, P.; et al. Identification of biologically active chemokine isoforms from ascitic fluid and elevated levels of CCL18/pulmonary and activation-regulated chemokine in ovarian carcinoma. J. Biol. Chem. 2002, 277, 24584–24593. [Google Scholar]
- Strieter, R.M.; Polverini, P.J.; Kunkel, S.L.; Arenberg, D.A.; Burdick, M.D.; Kasper, J.; Dzuiba, J.; van Damme, J.; Walz, A.; Marriott, D.; et al. The functional role of the ELR motif in CXC chemokine-mediated angiogenesis. J. Biol. Chem. 1995, 270, 27348–27357. [Google Scholar]
- Romagnani, P.; Annunziato, F.; Lasagni, L.; Lazzeri, E.; Beltrame, C.; Francalanci, M.; Uguccioni, M.; Galli, G.; Cosmi, L.; Maurenzig, L.; et al. Cell cycle-dependent expression of CXC chemokine receptor 3 by endothelial cells mediates angiostatic activity. J. Clin. Invest. 2001, 107, 53–63. [Google Scholar] [CrossRef]
- Bikfalvi, A. Recent developments in the inhibition of angiogenesis: Examples from studies on platelet factor-4 and the VEGF/VEGFR system. Biochem. Pharmacol. 2004, 68, 1017–1021. [Google Scholar] [CrossRef]
- Perollet, C.; Han, Z.C.; Savona, C.; Caen, J.P.; Bikfalvi, A. Platelet factor 4 modulates fibroblast growth factor 2 (FGF-2) activity and inhibits FGF-2 dimerization. Blood 1998, 91, 3289–3299. [Google Scholar]
- Jouan, V.; Canron, X.; Alemany, M.; Caen, J.P.; Quentin, G.; Plouet, J.; Bikfalvi, A. Inhibition of in vitro angiogenesis by platelet factor-4-derived peptides and mechanism of action. Blood 1999, 94, 984–993. [Google Scholar]
- Kawada, K.; Sonoshita, M.; Sakashita, H.; Takabayashi, A.; Yamaoka, Y.; Manabe, T.; Inaba, K.; Minato, N.; Oshima, M.; Taketo, M.M. Pivotal role of CXCR3 in melanoma cell metastasis to lymph nodes. CancerRes. 2004, 64, 4010–4017. [Google Scholar]
- Soejima, K.; Rollins, B.J. A functional IFN-gamma-inducible protein-10/CXCL10-specific receptor expressed by epithelial and endothelial cells that is neither CXCR3 nor glycosaminoglycan. J. Immunol. 2001, 167, 6576–6582. [Google Scholar]
- Furuya, M.; Yoneyama, T.; Miyagi, E.; Tanaka, R.; Nagahama, K.; Miyagi, Y.; Nagashima, Y.; Hirahara, F.; Inayama, Y.; Aoki, I. Differential expression patterns of CXCR3 variants and corresponding CXC chemokines in clear cell ovarian cancers and endometriosis. Gynecol. Oncol. 2011, 122, 648–655. [Google Scholar] [CrossRef]
- Furuya, M.; Tanaka, R.; Miyagi, E.; Kami, D.; Nagahama, K.; Miyagi, Y.; Nagashima, Y.; Hirahara, F.; Inayama, Y.; Aoki, I. Impaired CXCL4 Expression in Tumor-Associated Macrophages (TAMs) of Ovarian Cancers Arising in Endometriosis. Cancer Biol. Ther. 2012, 13, 10. [Google Scholar]
- Scotton, C.J.; Wilson, J.L.; Milliken, D.; Stamp, G.; Balkwill, F.R. Epithelial cancer cell migration: A role for chemokine receptors? Cancer Res. 2001, 61, 4961–4965. [Google Scholar]
- Scotton, C.J.; Wilson, J.L.; Scott, K.; Stamp, G.; Wilbanks, G.D.; Fricker, S.; Bridger, G.; Balkwill, F.R. Multiple actions of the chemokine CXCL12 on epithelial tumor cells in human ovarian cancer. Cancer Res. 2002, 62, 5930–5938. [Google Scholar]
- Kryczek, I.; Lange, A.; Mottram, P.; Alvarez, X.; Cheng, P.; Hogan, M.; Moons, L.; Wei, S.; Zou, L.; Machelon, V.; et al. CXCL12 and vascular endothelial growth factor synergistically induce neoangiogenesis in human ovarian cancers. Cancer Res. 2005, 65, 465–472. [Google Scholar]
- Sica, A.; Saccani, A.; Bottazzi, B.; Bernasconi, S.; Allavena, P.; Gaetano, B.; Fei, F.; LaRosa, G.; Scotton, C.; Balkwill, F.; et al. Defective expression of the monocyte chemotactic protein-1 receptor CCR2 in macrophages associated with human ovarian carcinoma. J. Immunol. 2000, 164, 733–738. [Google Scholar]
- Nesbeth, Y.C.; Martinez, D.G.; Toraya, S.; Scarlett, U.K.; Cubillos-Ruiz, J.R.; Rutkowski, M.R.; Conejo-Garcia, J.R. CD4+ T cells elicit host immune responses to MHC class II-negative ovarian cancer through CCL5 secretion and CD40-mediated licensing of dendritic cells. J. Immunol. 2010, 184, 5654–5662. [Google Scholar]
- Paley, P.J.; Staskus, K.A.; Gebhard, K.; Mohanraj, D.; Twiggs, L.B.; Carson, L.F.; Ramakrishnan, S. Vascular endothelial growth factor expression in early stage ovarian carcinoma. Cancer 1997, 80, 98–106. [Google Scholar] [CrossRef]
- Hartenbach, E.M.; Olson, T.A.; Goswitz, J.J.; Mohanraj, D.; Twiggs, L.B.; Carson, L.F.; Ramakrishnan, S. Vascular endothelial growth factor (VEGF) expression and survival in human epithelial ovarian carcinomas. Cancer Lett. 1997, 121, 169–175. [Google Scholar] [CrossRef]
- Cooper, B.C.; Ritchie, J.M.; Broghammer, C.L.; Coffin, J.; Sorosky, J.I.; Buller, R.E.; Hendrix, M.J.; Sood, A.K. Preoperative serum vascular endothelial growth factor levels: Significance in ovarian cancer. Clin. Cancer Res. 2002, 8, 3193–3197. [Google Scholar]
- Furuya, M.; Yonemitsu, Y.; Aoki, I. Angiogenesis: Complexity of tumor vasculature and microenvironment. Curr. Pharm. Des. 2009, 15, 1854–1867. [Google Scholar] [CrossRef]
- Miyahara, Y.; Odunsi, K.; Chen, W.; Peng, G.; Matsuzaki, J.; Wang, R.F. Generation and regulation of human CD4+ IL-17-producing T cells in ovarian cancer. Proc. Natl. Acad. Sci. USA 2008, 105, 15505–15510. [Google Scholar] [CrossRef]
- Kryczek, I.; Banerjee, M.; Cheng, P.; Vatan, L.; Szeliga, W.; Wei, S.; Huang, E.; Finlayson, E.; Simeone, D.; Welling, T.H.; et al. Phenotype, distribution, generation, and functional and clinical relevance of Th17 cells in the human tumor environments. Blood 2009, 114, 1141–1149. [Google Scholar] [CrossRef]
- Leveque, L.; Deknuydt, F.; Bioley, G.; Old, L.J.; Matsuzaki, J.; Odunsi, K.; Ayyoub, M.; Valmori, D. Interleukin 2-mediated conversion of ovarian cancer-associated CD4+ regulatory T cells into proinflammatory interleukin 17-producing helper T cells. J. Immunother. 2009, 32, 101–108. [Google Scholar] [CrossRef]
- Zhu, X.; Mulcahy, L.A.; Mohammed, R.A.; Lee, A.H.; Franks, H.A.; Kilpatrick, L.; Yilmazer, A.; Paish, E.C.; Ellis, I.O.; Patel, P.M.; et al. IL-17 expression by breast-cancer-associated macrophages: IL-17 promotes invasiveness of breast cancer cell lines. Breast Cancer Res. 2008, 10, R95. [Google Scholar]
- Wang, L.; Yi, T.; Kortylewski, M.; Pardoll, D.M.; Zeng, D.; Yu, H. IL-17 can promote tumor growth through an IL-6-Stat3 signaling pathway. J. Exp Med. 2009, 206, 1457–1464. [Google Scholar] [CrossRef]
- Nam, J.S.; Terabe, M.; Kang, M.J.; Chae, H.; Voong, N.; Yang, Y.A.; Laurence, A.; Michalowska, A.; Mamura, M.; Lonning, S.; et al. Transforming growth factor beta subverts the immune system into directly promoting tumor growth through interleukin-17. Cancer Res. 2008, 68, 3915–3923. [Google Scholar]
- Martin-Orozco, N.; Muranski, P.; Chung, Y.; Yang, X.O.; Yamazaki, T.; Lu, S.; Hwu, P.; Restifo, N.; P.Overwijk, W.W.; Dong, C. T helper 17 cells promote cytotoxic T cell activation in tumor immunity. Immunity 2009, 31, 787–798. [Google Scholar] [CrossRef]
- Buckanovich, R.J.; Facciabene, A.; Kim, S.; Benencia, F.; Sasaroli, D.; Balint, K.; Katsaros, D.; O'Brien-Jenkins, A.; Gimotty, P.A.; Coukos, G. Endothelin B receptor mediates the endothelial barrier to T cell homing to tumors and disables immune therapy. Nat. Med. 2008, 14, 28–36. [Google Scholar] [CrossRef]
- Ben-Baruch, G.; Sivan, E.; Moran, O.; Rizel, S.; Menczer, J.; Seidman, D.S. Primary peritoneal serous papillary carcinoma: A study of 25 cases and comparison with stage III-IV ovarian papillary serous carcinoma. Gynecol. Oncol. 1996, 60, 393–396. [Google Scholar] [CrossRef]
- Euscher, E.D.; Malpica, A.; Deavers, M.T.; Silva, E.G. Differential expression of WT-1 in serous carcinomas in the peritoneum with or without associated serous carcinoma in endometrial polyps. Am. J. Surg. Pathol. 2005, 29, 1074–1078. [Google Scholar]
- Ordonez, N.G. Value of immunohistochemistry in distinguishing peritoneal mesothelioma from serous carcinoma of the ovary and peritoneum: A review and update. Adv. Anat. Pathol. 2006, 13, 16–25. [Google Scholar] [CrossRef]
- Muto, M.G.; Welch, W.R.; Mok, S.C.; Bandera, C.A.; Fishbaugh, P.M.; Tsao, S.W.; Lau, C.C.; Goodman, H.M.; Knapp, R.C.; Berkowitz, R.S. Evidence for a multifocal origin of papillary serous carcinoma of the peritoneum. Cancer Res. 1995, 55, 490–492. [Google Scholar]
- Bandera, C.A.; Muto, M.G.; Schorge, J.O.; Berkowitz, R.S.; Rubin, S.C.; Mok, S.C. BRCA1 gene mutations in women with papillary serous carcinoma of the peritoneum. Obstet. Gynecol. 1998, 92, 596–600. [Google Scholar] [CrossRef]
- Bakker, R.F.; Stoot, J.H.; Blok, P.; Merkus, J.W. Primary retroperitoneal mucinous cystadenoma with sarcoma-like mural nodule : A case report and review of the literature. Virchows Arch. 2007, 451, 853–857. [Google Scholar] [CrossRef]
- Pearl, M.L.; Valea, F.; Chumas, J.; Chalas, E. Primary retroperitoneal mucinous cystadenocarcinoma of low malignant potential: a case report and literature review. Gynecol. Oncol. 1996, 61, 150–152. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Furuya, M. Ovarian Cancer Stroma: Pathophysiology and the Roles in Cancer Development. Cancers 2012, 4, 701-724. https://doi.org/10.3390/cancers4030701
Furuya M. Ovarian Cancer Stroma: Pathophysiology and the Roles in Cancer Development. Cancers. 2012; 4(3):701-724. https://doi.org/10.3390/cancers4030701
Chicago/Turabian StyleFuruya, Mitsuko. 2012. "Ovarian Cancer Stroma: Pathophysiology and the Roles in Cancer Development" Cancers 4, no. 3: 701-724. https://doi.org/10.3390/cancers4030701