Light Influences How the Fungal Toxin Deoxynivalenol Affects Plant Cell Death and Defense Responses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
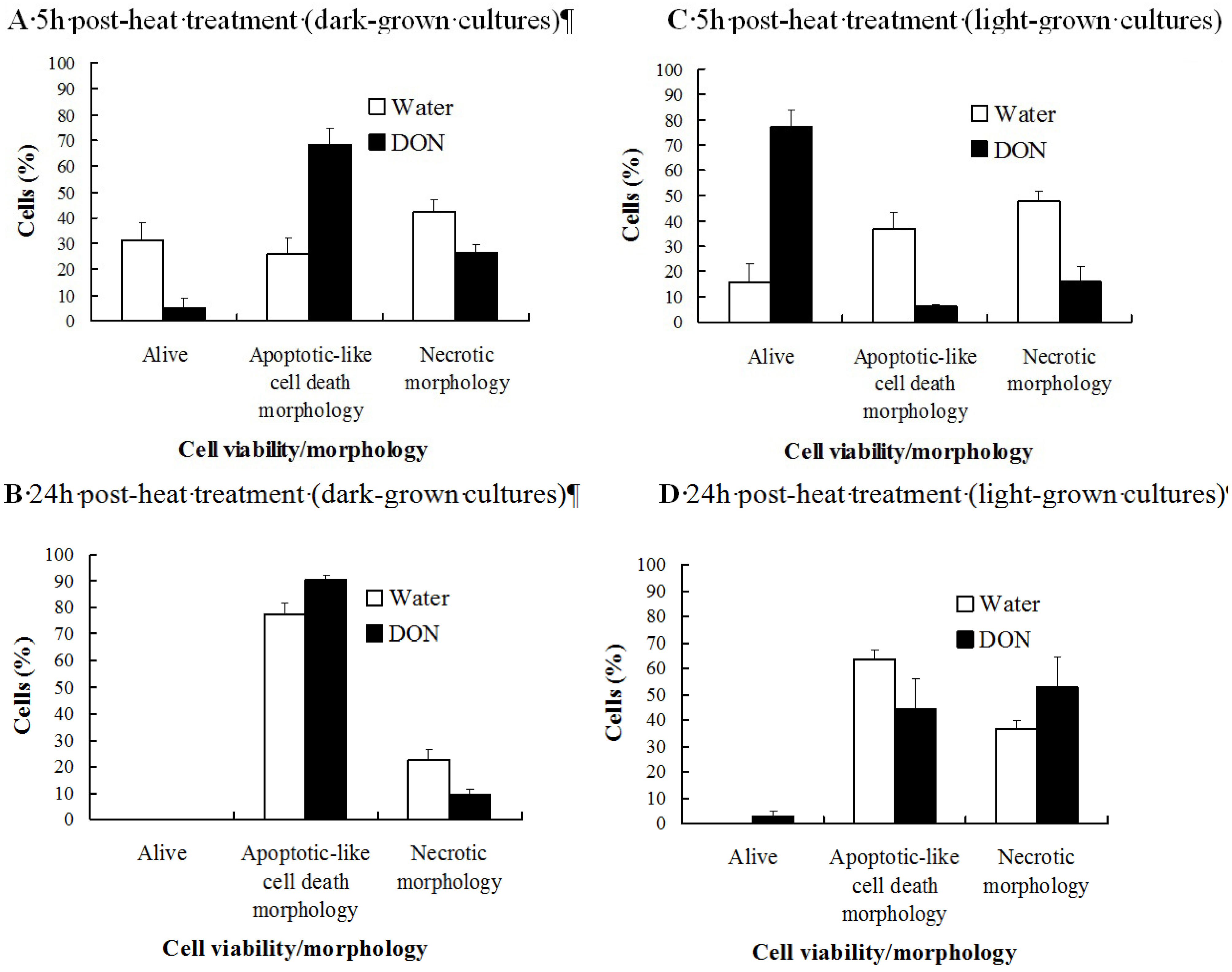
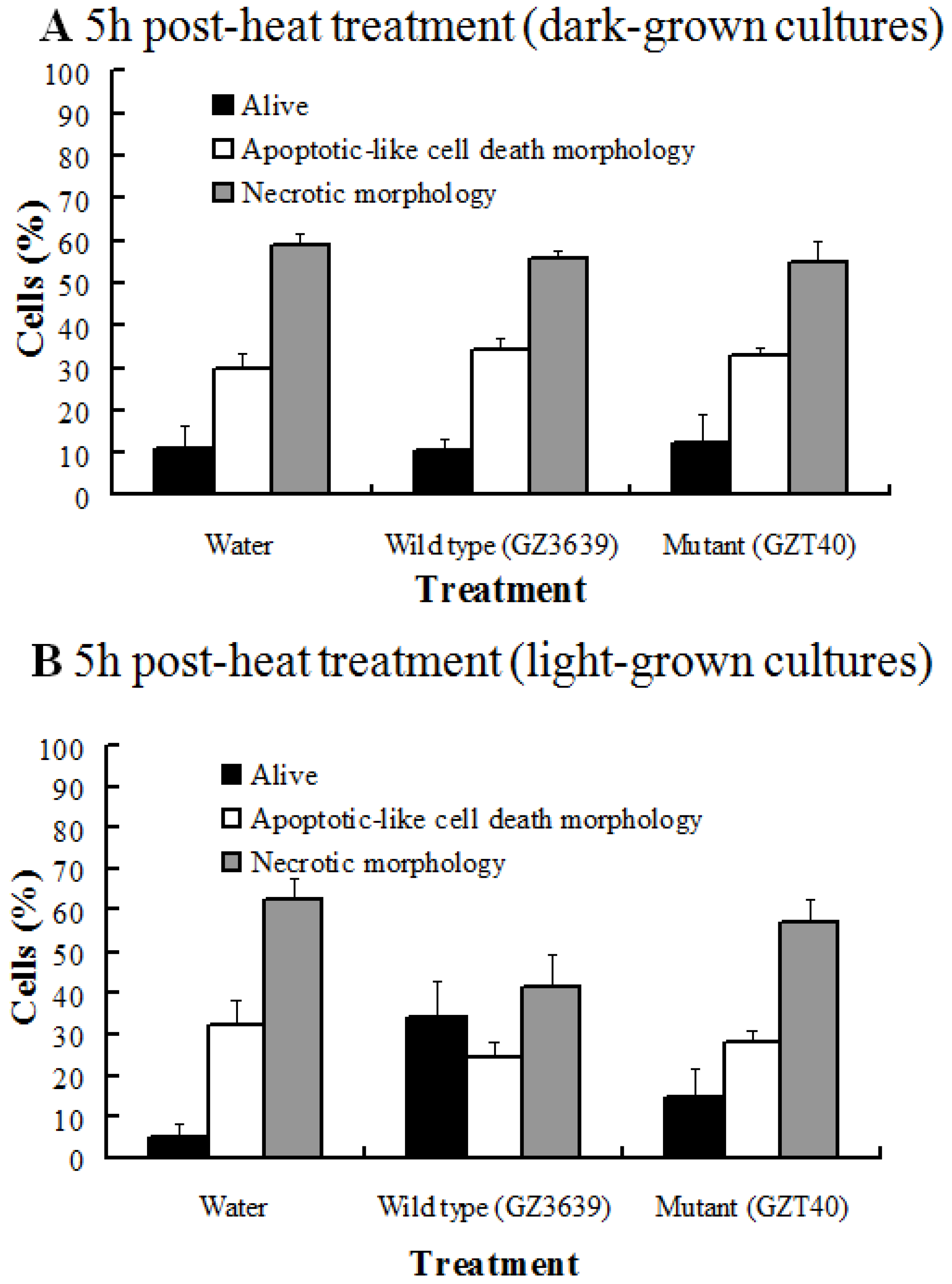
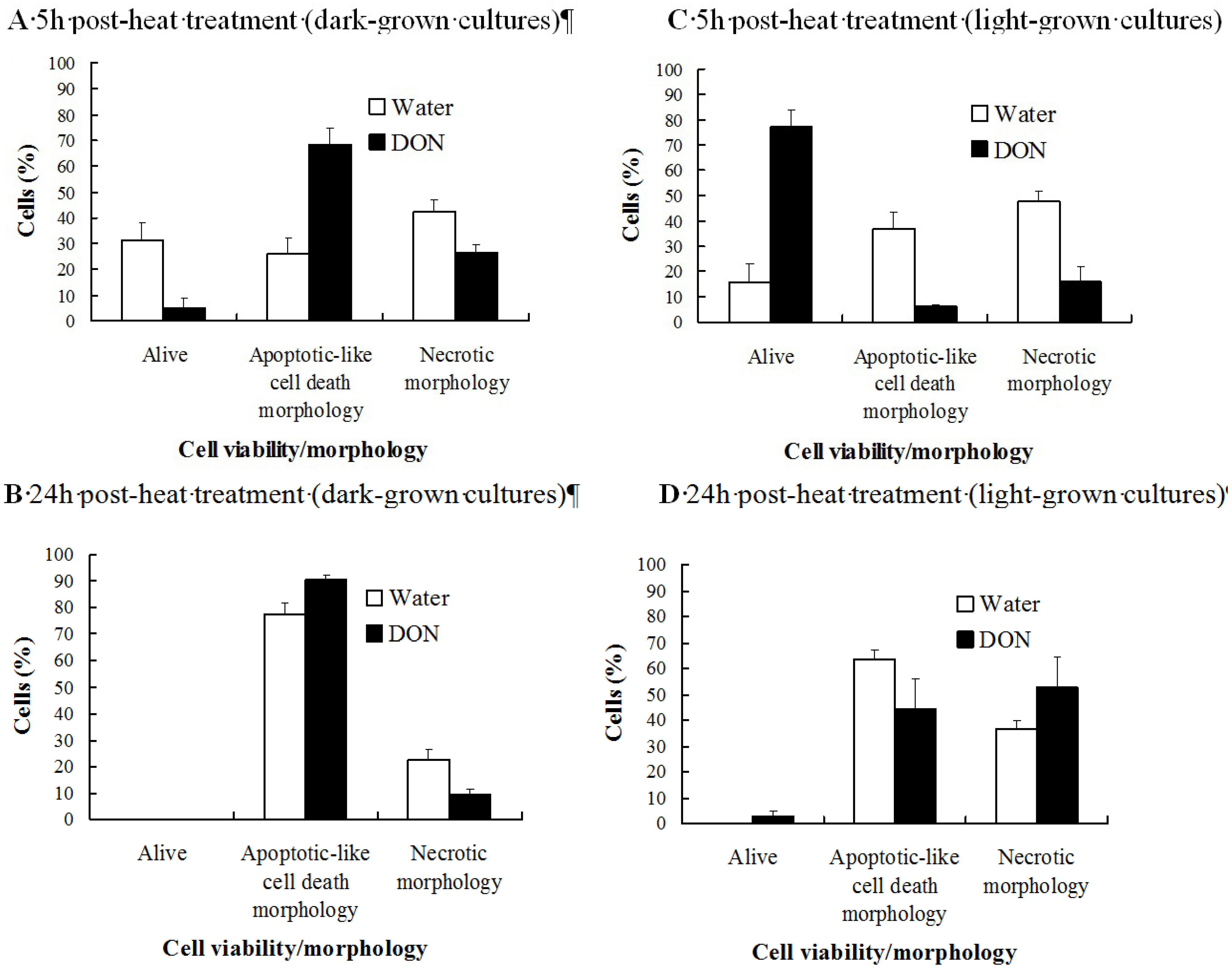
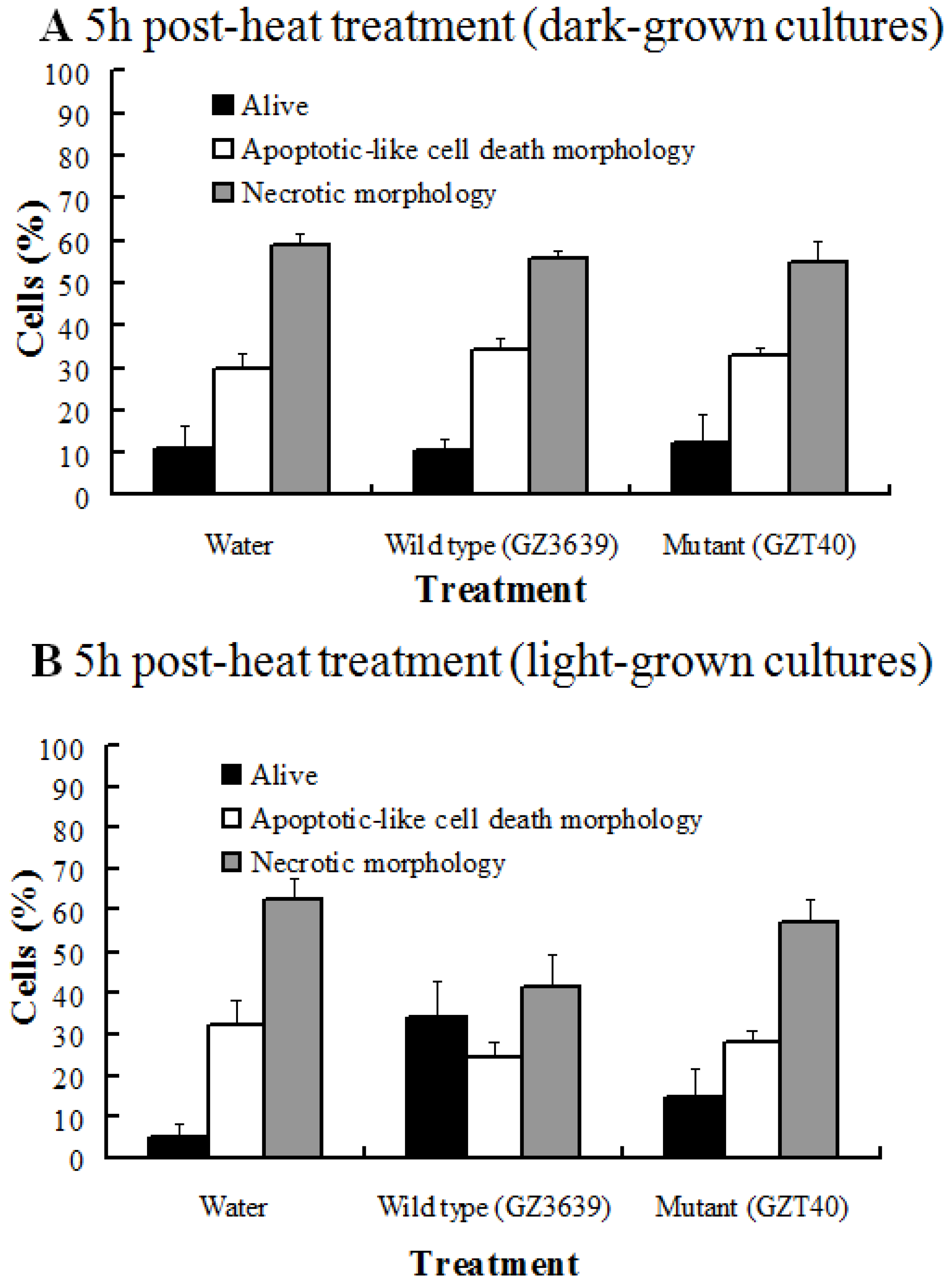
2.1. The Effect of DON on the Viability of Heat-Shocked Arabidopsis Cell Cultures Is Light Dependent
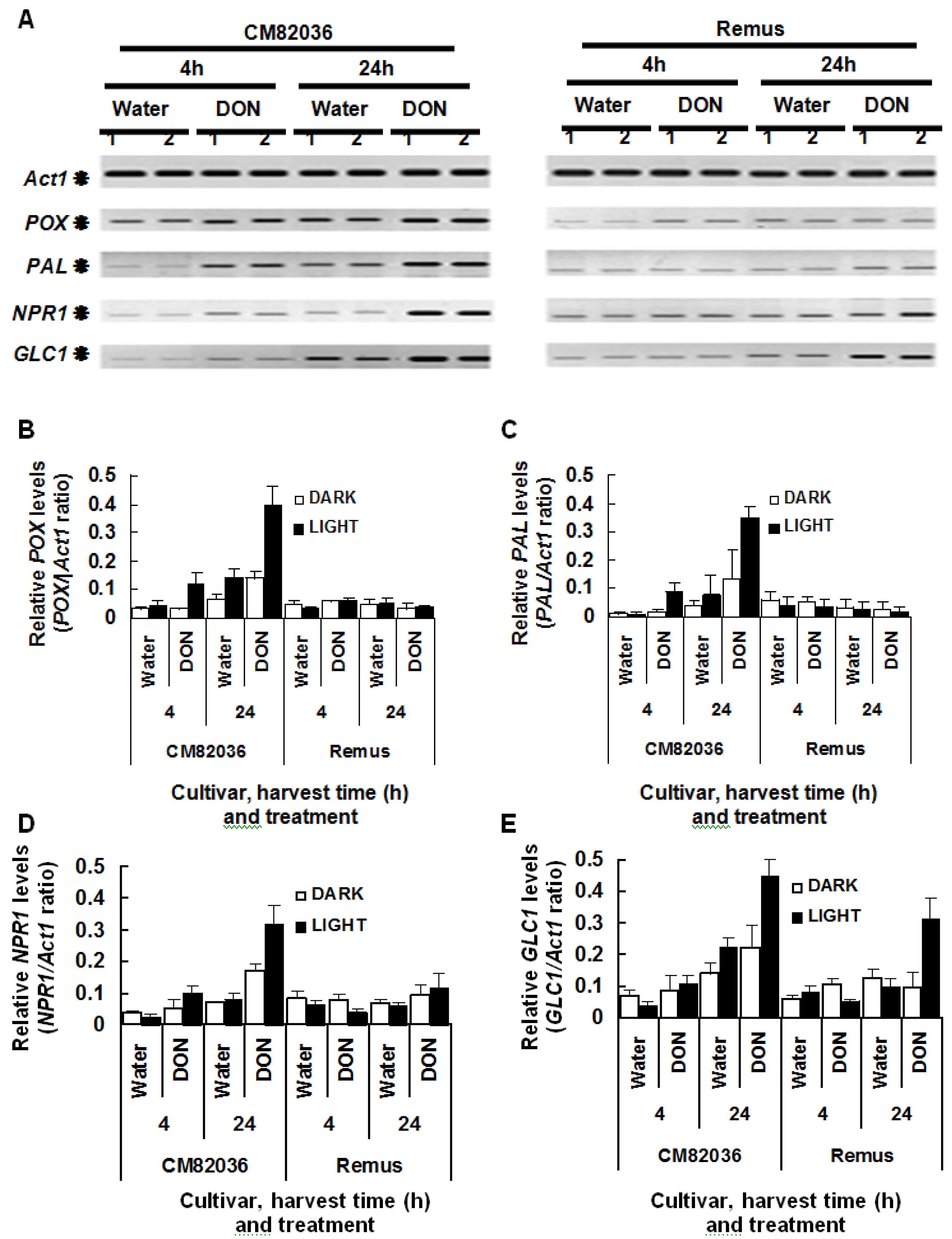
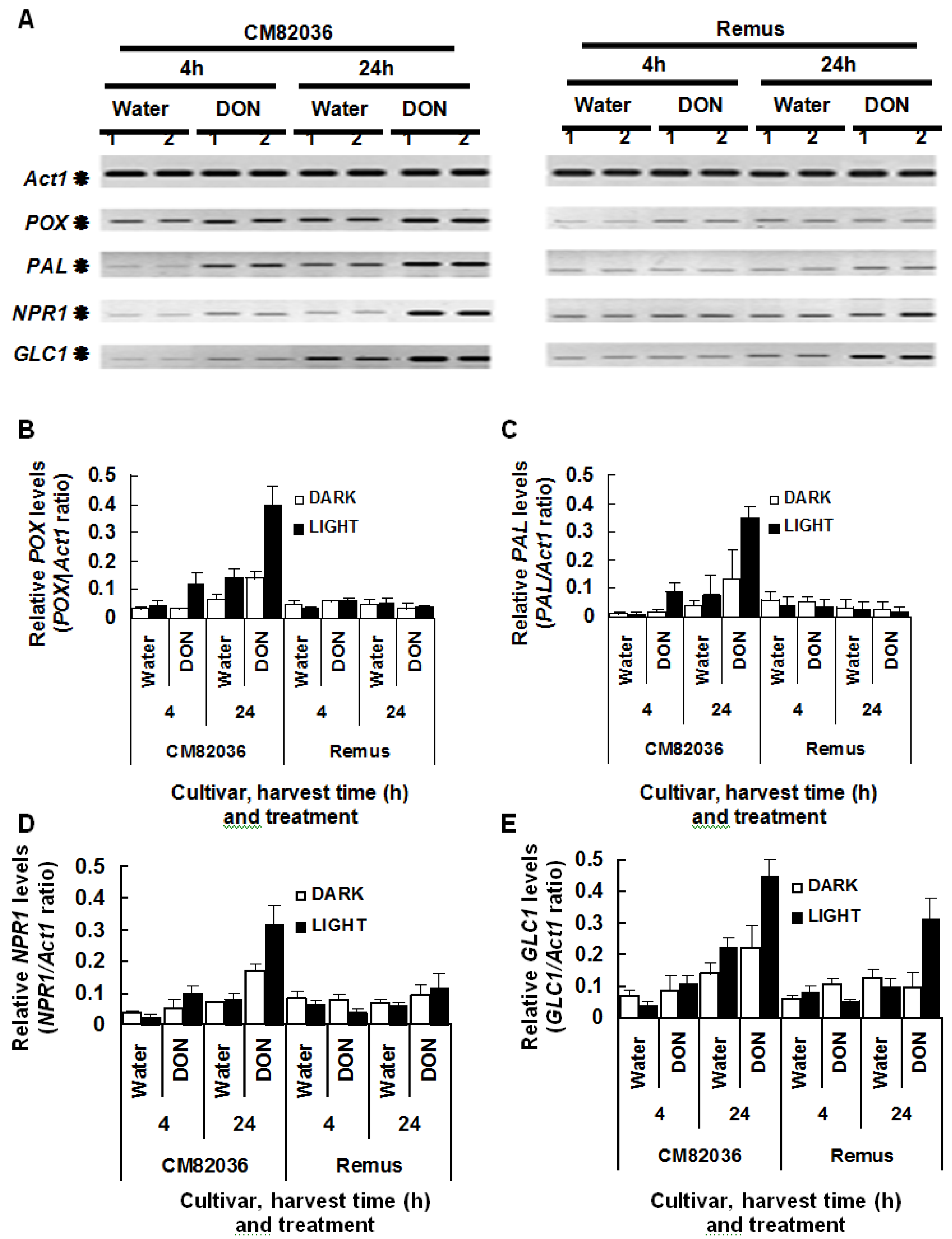
2.2. DON Induction of Defense Gene Expression in Wheat Is Light Enhanced and Genotype Dependent
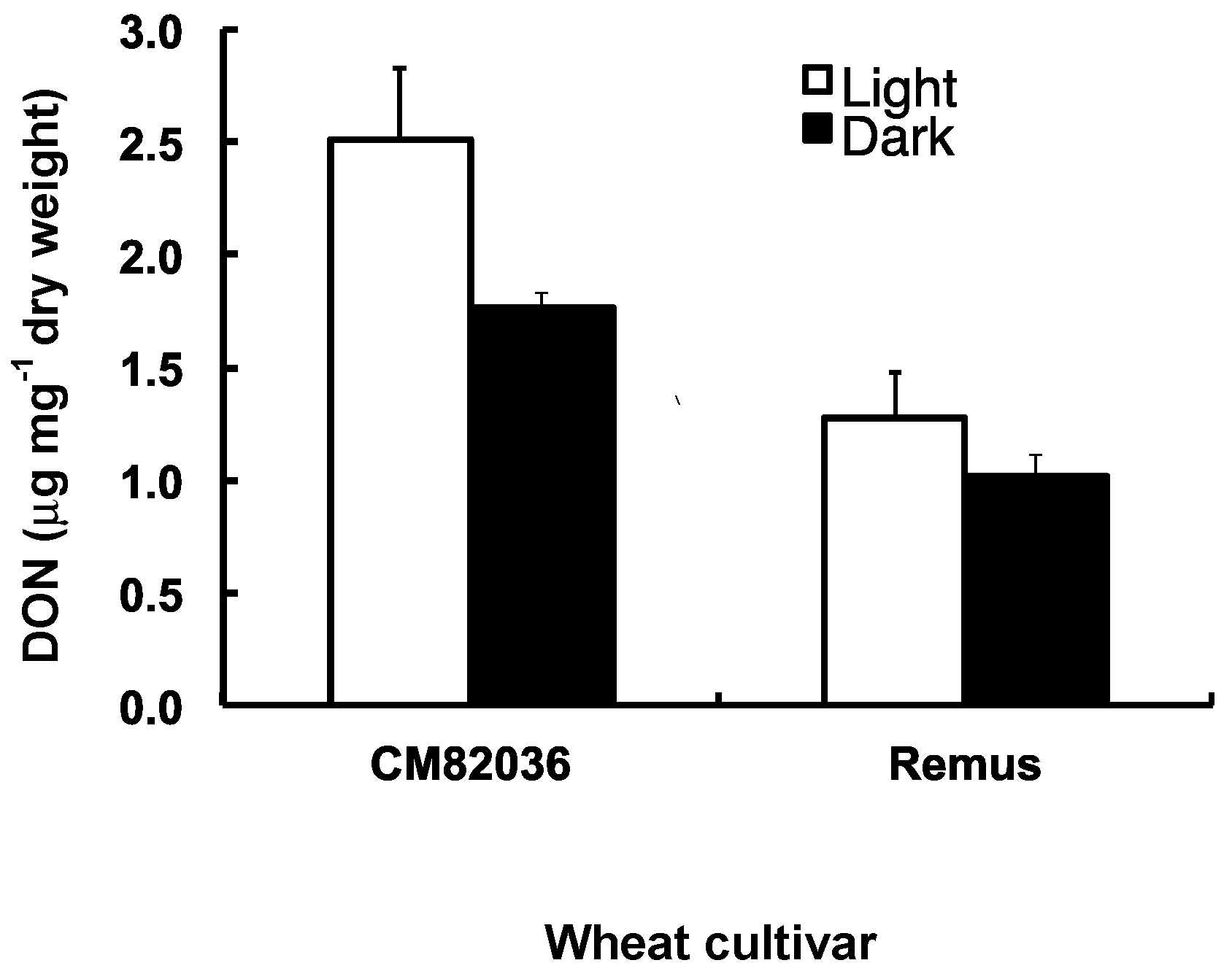
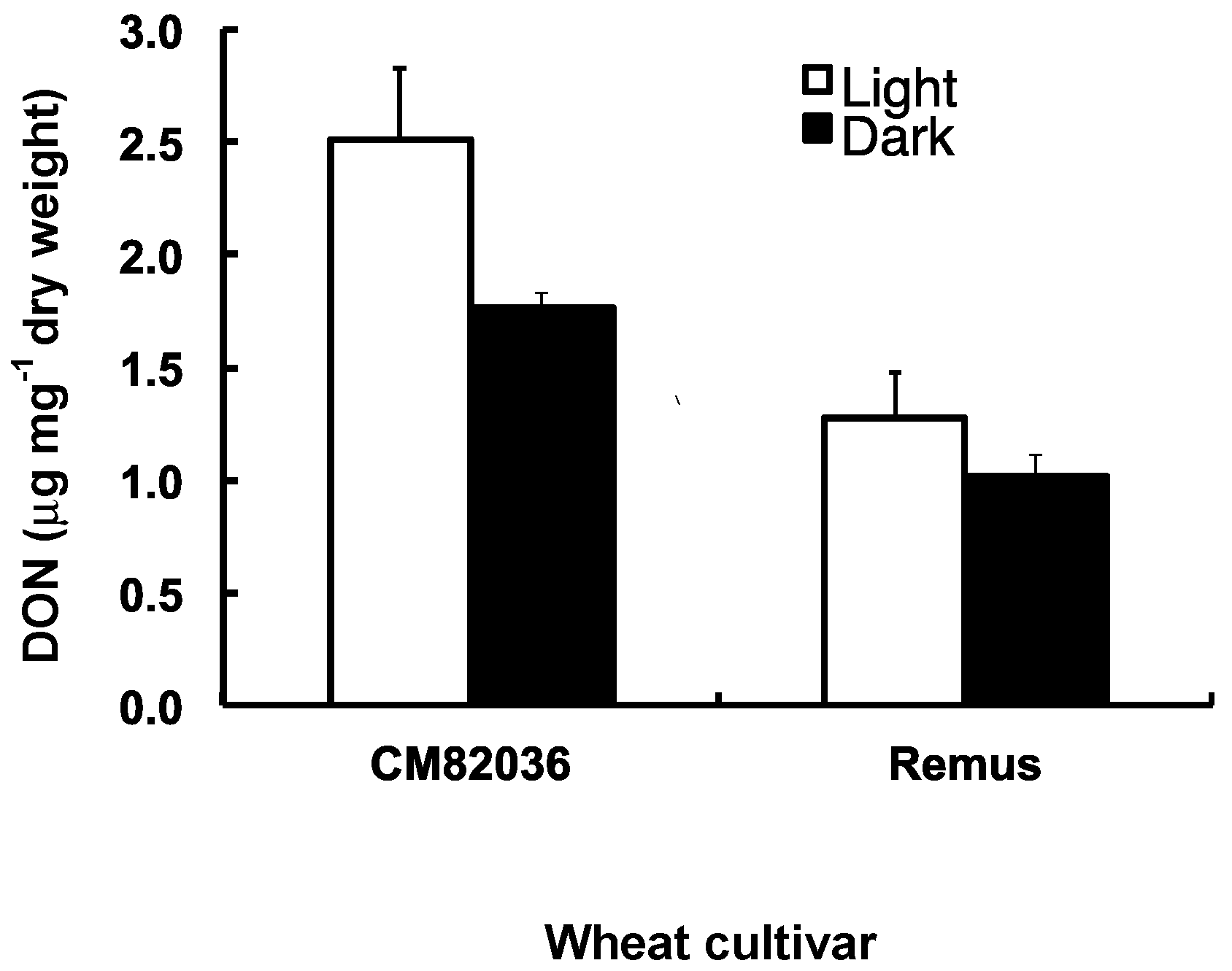
2.3. Both Genotype and Light Affect the Movement of DON Metabolites within Wheat Seedlings
3. Discussion
4. Experimental Section
4.1. Maintenance, Growth and Treatment of Arabidopsis Cells
4.2. Morphological Analysis of Arabidopsis Cells
4.3. Growth and Treatment of Wheat Seedlings
4.4. Quantification of DON
4.5. RNA Extraction and Gene-Specific RT-PCR Analyses
4.6. Data Analysis
5. Conclusions
Supplementary Files
Acknowledgements
Conflicts of Interest
References
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium ear blight (scab) in small-grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Langevin, F.; Eudes, F.; Comeau, A. Effect of trichothecenes produced by Fusarium graminearum during Fusarium head blight development in six cereal species. Eur. J. Plant Pathol. 2004, 110, 735–746. [Google Scholar] [CrossRef]
- Wang, H.; Hwang, S.F.; Eudes, F.; Chang, K.F.; Howard, R.J.; Turnbull, G.D. Trichothecenes and aggressiveness of Fusarium graminearum causing seedling blight and root rot in cereals. Plant Pathol. 2006, 55, 224–230. [Google Scholar] [CrossRef]
- Lemmens, M.; Scholz, U.; Berthiller, F.; Dall’Asta, C.; Koutnik, A.; Schuhmacher, R.; Adam, G.; Buerstmayr, H.; Mesterhazy, A.; Krska, R.; et al. The ability to detoxify the mycotoxin deoxynivalenol colocalizes with a major quantitative trait locus for Fusarium head blight resistance in wheat. Mol. Plant. Microbe Interact. 2005, 18, 1318–1324. [Google Scholar] [CrossRef]
- Ansari, K.I.; Walter, S.; Brennan, J.M.; Lemmens, M.; Kessans, S.; McGahern, A.; Egan, D.; Doohan, F.M. Retrotransposon and gene activation in wheat in response to mycotoxigenic and non-mycotoxigenic-associated Fusarium stress. Theor. Appl. Genet. 2007, 114, 927–937. [Google Scholar] [CrossRef]
- Desmond, O.J.; Manners, J.M.; Stephens, A.E.; MaClean, D.J.; Schenk, P.M.; Gardiner, D.M.; Munn, A.L.; Kazan, K. The Fusarium mycotoxin deoxynivalenol elicits hydrogen peroxide production, programmed cell death and defence responses in wheat. Mol. Plant Pathol. 2008, 9, 435–445. [Google Scholar] [CrossRef]
- Walter, S.; Brennan, J.M.; Arunachalam, C.; Ansari, K.I.; Hu, X.; Khan, M.R.; Trognit, Z.F.; Trognitz, B.; Leonard, G.; Egan, D.; et al. Components of the gene network associated with genotype-dependent response of wheat to the Fusarium mycotoxin deoxynivalenol. Funct. Integr. Genomics 2008, 8, 421–427. [Google Scholar] [CrossRef]
- Diamond, M.; Reape, T.J.; Rocha, O.; Doyle, S.M.; Doohan, F.M.; McCabe, P.F. The mycotoxin Deoxynivalenol produced by necrotrophic Fusarium species can inhibit plant apoptotic-like programmed cell death. PLoS ONE 2013. [Google Scholar] [CrossRef]
- Bushnell, W.R.; Seeland, T.M.; Perkins-Veazie, P.M.; Krueger, D.E.; Collins, J.K.; Russo, V.M. The effects of deoxynivalenol on barley leaf tissues. In Genomic and Genetic Analysis of Plant Parasitism and Defence; Tsuyumu, S., Leach, J.E., Shiraishi, T., Wolpert, T., Eds.; APS Press: St. Paul, MN, USA, 2004; pp. 270–284. [Google Scholar]
- Poppenberger, B.; Berthiller, F.; Lucyshyn, D.; Sieberer, T.; Schuhmacher, R.; Krska, R.; Kuchler, K.; Glössl, J.; Luschnig, C.; Adam, G. Detoxification of the Fusarium mycotoxin Deoxynivalenol by a UDP-glucosyltransferase from Arabidopsis thaliana. J. Biol. Chem. 2003, 278, 47905–47914. [Google Scholar] [CrossRef]
- Doyle, S.M.; Diamond, M.; McCabe, P.F. Chloroplast and reactive oxygen species involvement in apoptotic-like programmed cell death in Arabidopsis suspension cultures. J. Exp. Bot. 2010, 61, 473–482. [Google Scholar] [CrossRef]
- Doohan, F.M. University College Dublin: Dublin, Ireland, Unpublished data. 2008.
- Steiner, B.; Kurz, H.; Lemmens, M.; Buerstmayr, H. Differential gene expression of related wheat lines with contrasting levels of head blight resistance after Fusarium graminearum inoculation. Theor. Appl. Genet. 2009. [Google Scholar] [CrossRef]
- Chamnongpol, S.; Willekens, H.; Langebartels, C.; VanMontagu, M.; Inze, D.; VanCamp, W. Transgenic tobacco with a reduced catalase activity develops necrotic lesions and induces pathogenesis-related expression under high light. Plant J. 1996, 10, 491–503. [Google Scholar]
- Genoud, T.; Buchala, A.J.; Chua, N.H.; Metraux, J.P. Phytochrome signalling modulates the SA-perceptive pathway in Arabidopsis. Plant J. 2002, 31, 87–95. [Google Scholar] [CrossRef]
- Metraux, J.P.; Nawrath, C.; Genoud, T. Systemic acquired resistance. Euphytica 2002, 124, 237–243. [Google Scholar] [CrossRef]
- Zeier, J.; Pink, B.; Mueller, M.J.; Berger, S. Light conditions influence specific defence responses in incompatible plant-pathogen interactions: Uncoupling systemic resistance from salicylic acid and PR-1 accumulation. Planta 2004, 219, 673–683. [Google Scholar]
- Griebel, T.; Zeier, J. Light regulation and daytime dependency of inducible plant defenses in arabidopsis: Phytochrome signaling controls systemic acquired resistance rather than local defense. Plant Physiol. 2008, 147, 790–801. [Google Scholar] [CrossRef]
- Roberts, M.R.; Paul, N.D. Seduced by the dark side: Integrating molecular and ecological perspectives on the influence of light on plant defence against pests and pathogens. New Phytol. 2006, 170, 677–699. [Google Scholar] [CrossRef] [Green Version]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant Biol. 2004, 55, 373–399. [Google Scholar] [CrossRef]
- Babaeizad, V.; Imani, J.; Kogel, K.H.; Eichmann, R.; Hückelhoven, R. Over-expression of the cell death regulator BAX inhibitor-1 in barley confers reduced or enhanced susceptibility to distinct fungal pathogens. Theor. Appl. Genet. 2008, 118, 455–463. [Google Scholar]
- Durrant, W.E.; Dong, X. Systemic acquired resistance. Annu. Rev. Phytopathol. 2004, 42, 185–209. [Google Scholar] [CrossRef]
- Cuzick, A.; Lee, S.; Gezan, S.; Hammond-Kosack, K.E. NPR1 and EDS11 contribute to host resistance against Fusarium. culmorum in Arabidopsis buds and flowers. Mol. Plant Pathol. 2008, 9, 697–704. [Google Scholar] [CrossRef]
- Makandar, R.; Essig, J.S.; Schapaugh, M.A.; Trick, H.N.; Shah, J. Genetically engineered resistance to Fusarium head blight in wheat by expression of Arabidopsis NPR1. Mol. Plant Microbe Interact. 2006, 19, 123–129. [Google Scholar] [CrossRef]
- Chern, M.; Fitzgerald, H.A.; Canlas, P.E.; Navarre, D.A.; Ronald, P.C. Overexpression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Mol. Plant-Microbe Interact. 2005, 18, 511–520. [Google Scholar] [CrossRef]
- Hamzehzarghani, H.; Kushalappa, A.C.; Dion, Y.; Rioux, S.; Comeau, A.; Yaylayan, V.; Marshall, W.D.; Mather, D.E. Metabolic profiling and factor analysis to discriminate quantitative resistance in wheat cultivars against Fusarium head blight. Physiol. Mol. Plant Pathol. 2005, 66, 119–133. [Google Scholar] [CrossRef]
- Gunnaiah, R.; Kushalappa, A.C.; Duggavathi, R.; Fox, S.; Somers, D.J. Integrated metabolo-proteomic approach to decipher the mechanisms by which wheat QTL (Fhb1) contributes to resistance against Fusarium graminearum. PloS One 2012, 7, e40695. [Google Scholar]
- Ruprich, J.; Ostry, J. Immunochemical methods in health risk assessment: Cross reactivity of antibodies against mycotoxin deoxynivalenol with deoxynivalenol-3-glucoside. Cent. Eur. J. Public Health 2008, 16, 34–37. [Google Scholar]
- May, M.J.; Leaver, C.J. Oxidative stimulation of glutathione synthesis in Arabidopsis thaliana suspension cultures. Plant Physiol. 1993, 103, 621–627. [Google Scholar]
- McCabe, P.F.; Leaver, C.J. Programmed cell death in cell cultures. Plant Mol. Biol. 2000, 44, 359–368. [Google Scholar] [CrossRef]
- Proctor, R.H.; Hohn, T.M.; McCormick, S.P. Reduced virulence of Gibberella zeae caused by disruption of a trichothecene toxin biosynthetic gene. Mol. Plant-Microbe Interact. 1995, 8, 593–601. [Google Scholar] [CrossRef]
- Li, W.L.; Faris, J.D.; Muthukrishnan, S.; Liu, D.J.; Chen, P.D.; Gill, B.S. Isolation and characterization of novel cDNA clones of acidic chitinases and beta-1,3-glucanases from wheat spikes infected by Fusarium graminearum. Theor. Appl. Genet. 2001, 102, 353–362. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Steiner, B.; Hartl, L.; Griesser, M.; Angerer, N.; Lengauer, D.; Miedaner, T.; Schneider, B.; Lemmens, M. Molecular mapping of QTLs for Fusarium head blight resistance in spring wheat. II. Resistance to fungal penetration and spread. Theor. Appl. Genet. 2003, 107, 503–508. [Google Scholar] [CrossRef]
- Doohan, F.M. University College Dublin: Dublin, Ireland, Unpublished work. 2010.
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ansari, K.I.; Doyle, S.M.; Kacprzyk, J.; Khan, M.R.; Walter, S.; Brennan, J.M.; Arunachalam, C.S.; McCabe, P.F.; Doohan, F.M. Light Influences How the Fungal Toxin Deoxynivalenol Affects Plant Cell Death and Defense Responses. Toxins 2014, 6, 679-692. https://doi.org/10.3390/toxins6020679
Ansari KI, Doyle SM, Kacprzyk J, Khan MR, Walter S, Brennan JM, Arunachalam CS, McCabe PF, Doohan FM. Light Influences How the Fungal Toxin Deoxynivalenol Affects Plant Cell Death and Defense Responses. Toxins. 2014; 6(2):679-692. https://doi.org/10.3390/toxins6020679
Chicago/Turabian StyleAnsari, Khairul I., Siamsa M. Doyle, Joanna Kacprzyk, Mojibur R. Khan, Stephanie Walter, Josephine M. Brennan, Chanemouga Soundharam Arunachalam, Paul F. McCabe, and Fiona M. Doohan. 2014. "Light Influences How the Fungal Toxin Deoxynivalenol Affects Plant Cell Death and Defense Responses" Toxins 6, no. 2: 679-692. https://doi.org/10.3390/toxins6020679