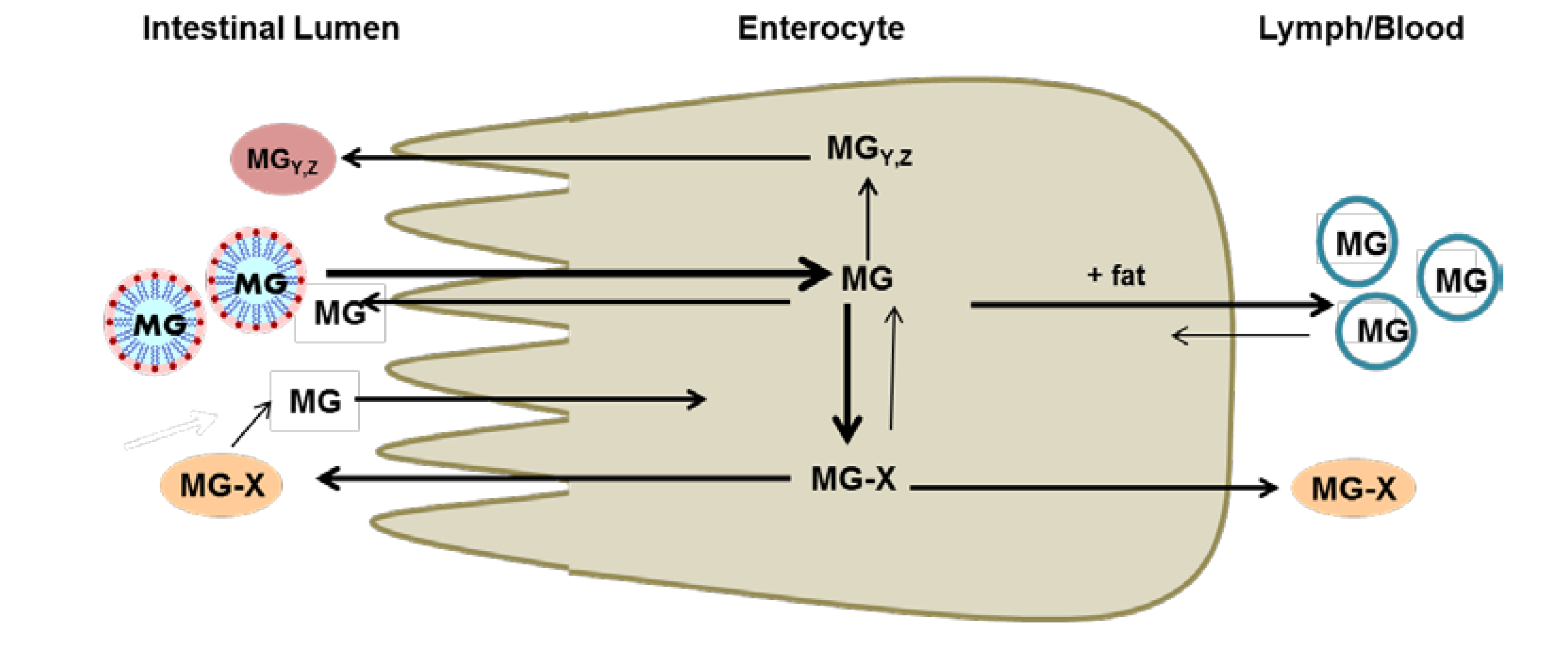
1. Introduction
Juice blends and other products containing exotic fruits, also known as
superfruits, have been aggressively marketed for their proposed health benefits. This has resulted in a steady rise in sales of superfruit juices and products to consumers interested in their personal health. Mangosteen is one such superfruit that is produced by
Garcinia mangostana L. The genus
Garcinia is native to Asia and Africa and includes more than 300 distinct species from which several families of bioactive compounds such as xanthones, flavonoids, triterpenoids, and benzophenones have been isolated and characterized [
1]. Although many
Garcinia species including
G. mangostana,
G. schomburgkiana,
G. dulcis,
G. cowa,
G. atroviridis,
G. hanburyi,
G. bancana,
G. xanthochymus,
G. thorelii,
G. hombroniana, and
G. speciosa bear edible fruits, mangosteen has captured the most attention in the market [
2]. The mangosteen tree is mainly cultivated in Indonesia, Malaysia, the Philippines, and Thailand. Mature mangosteen trees range from 6 to 25 m. Production of the fruit generally requires 10 or more years with a yield of around 400 fruits per tree that is increased in older trees. Mangosteen fruit is round, dark purple or reddish, and has a white juicy pulp possessing a slightly acidic and sweet flavor that is enjoyed by many, and has resulted in it being referred to as the “queen of fruits”. The pericarp of mangosteen fruit has been used in traditional medicine in Southeast Asia for centuries to treat infection, wounds, inflammation and diarrhea [
3].
Products containing mangosteen juice or extract are a fast growing segment of the functional beverages market. Aggressive marketing of the proposed health benefits of mangosteen has resulted in sales of mangosteen products in the US exceeding $200 million in 2008 [
4]. Oftentimes, products marketed as mangosteen juice are a blend of numerous fruit juices with mangosteen being one of the less abundant components. For example, Xango
®, one of the bestselling mangosteen products in the US, contains mangosteen puree, and a blend of juices from grape, pear, apple, blueberry, strawberry, raspberry, cranberry, and cherry.
Secondary metabolites, known as xanthones, have been isolated from the pericarp of mangosteen and are attributed to the medicinal properties of the fruit. Xanthones have a unique chemical structure composed of a tricyclic aromatic system (C
6–C
3–C
6). Isoprene, methoxyl and hydroxyl groups located at various locations on the A and B rings, resulting in a diverse array of xanthone compounds. Xanthones are found in a select few higher plant families. At least 68 distinct xanthones have been identified in different parts of the
G. mangostana plant with 50 being present in the fruit’s pericarp at higher concentrations than in the aril or edible portion of the fruit [
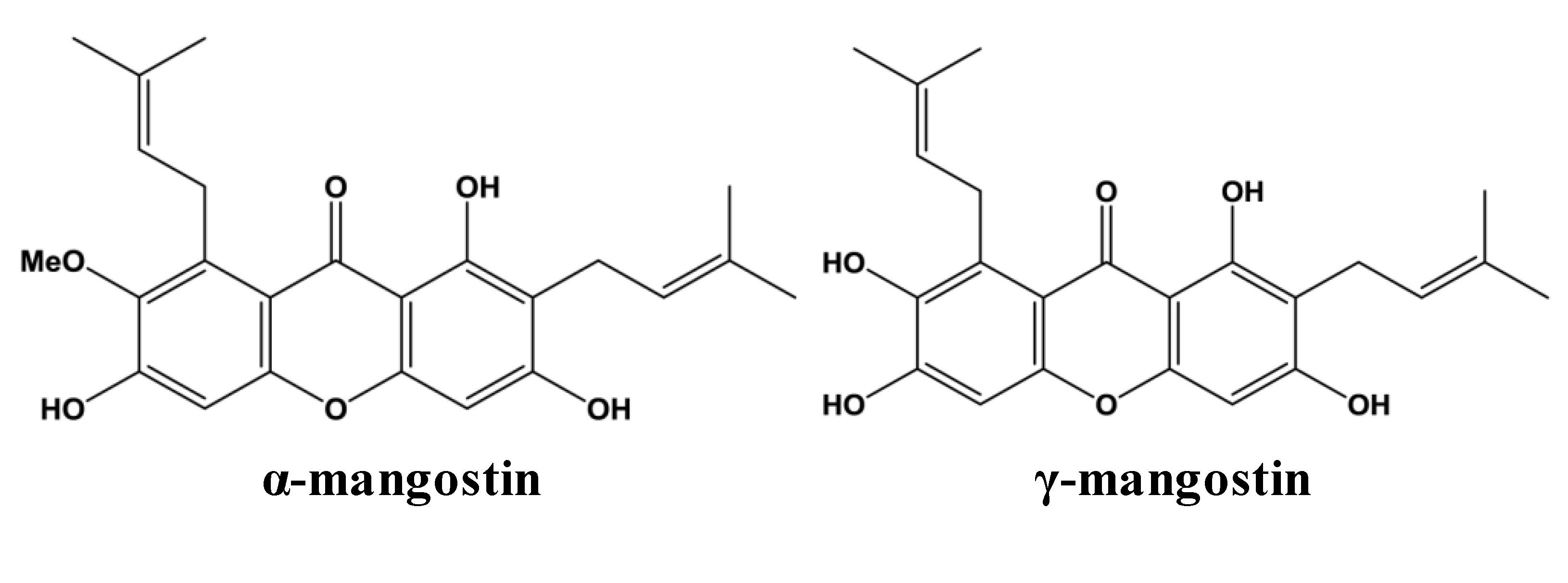
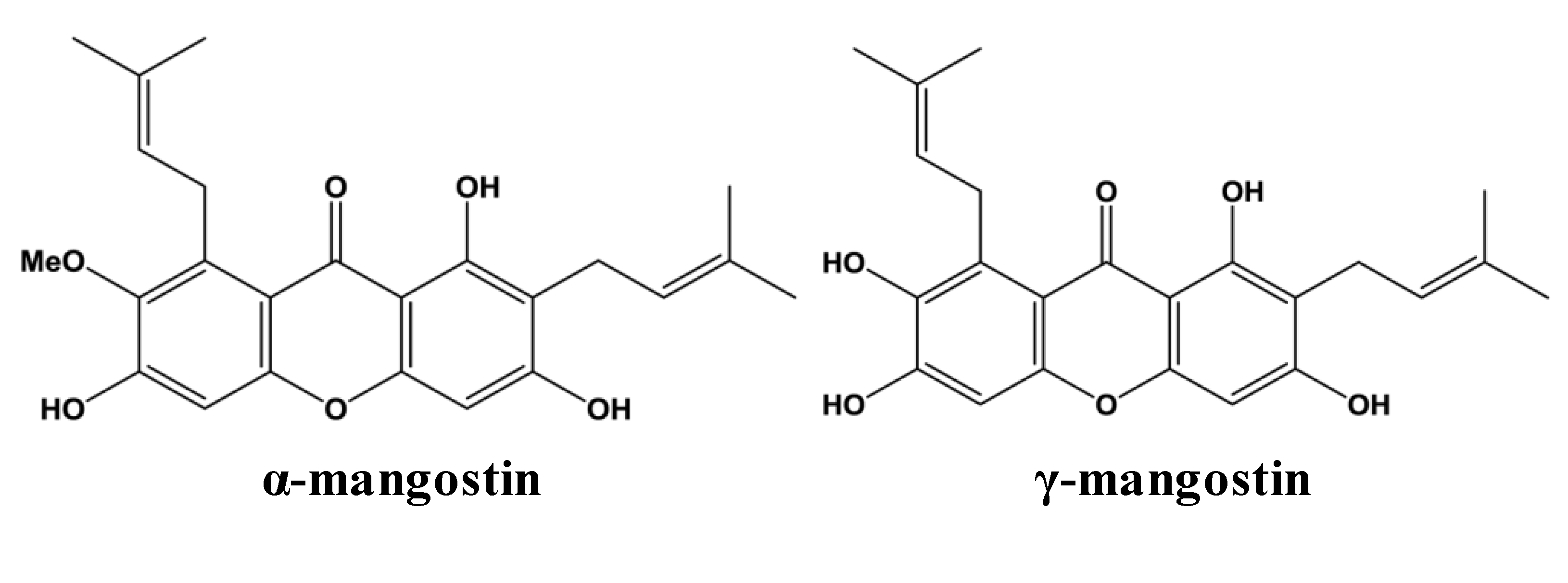
5]. The most abundant xanthones in the pericarp of mangosteen fruit are α- and γ-mangostin (
Figure 1) [
6]. Other xanthones in mangosteen pericarp include β-mangostin, gartanin, 8-deoxygartanin, garcinones A, B, C, D and E, mangostinone, 9-hydroxycalabaxanthone and isomangostin, among others. Details regarding the extraction and identification of these and other xanthones have been reviewed elsewhere [
5].
Figure 1.
Chemical structures of two most abundant xanthones in mangosteen.
Figure 1.
Chemical structures of two most abundant xanthones in mangosteen.
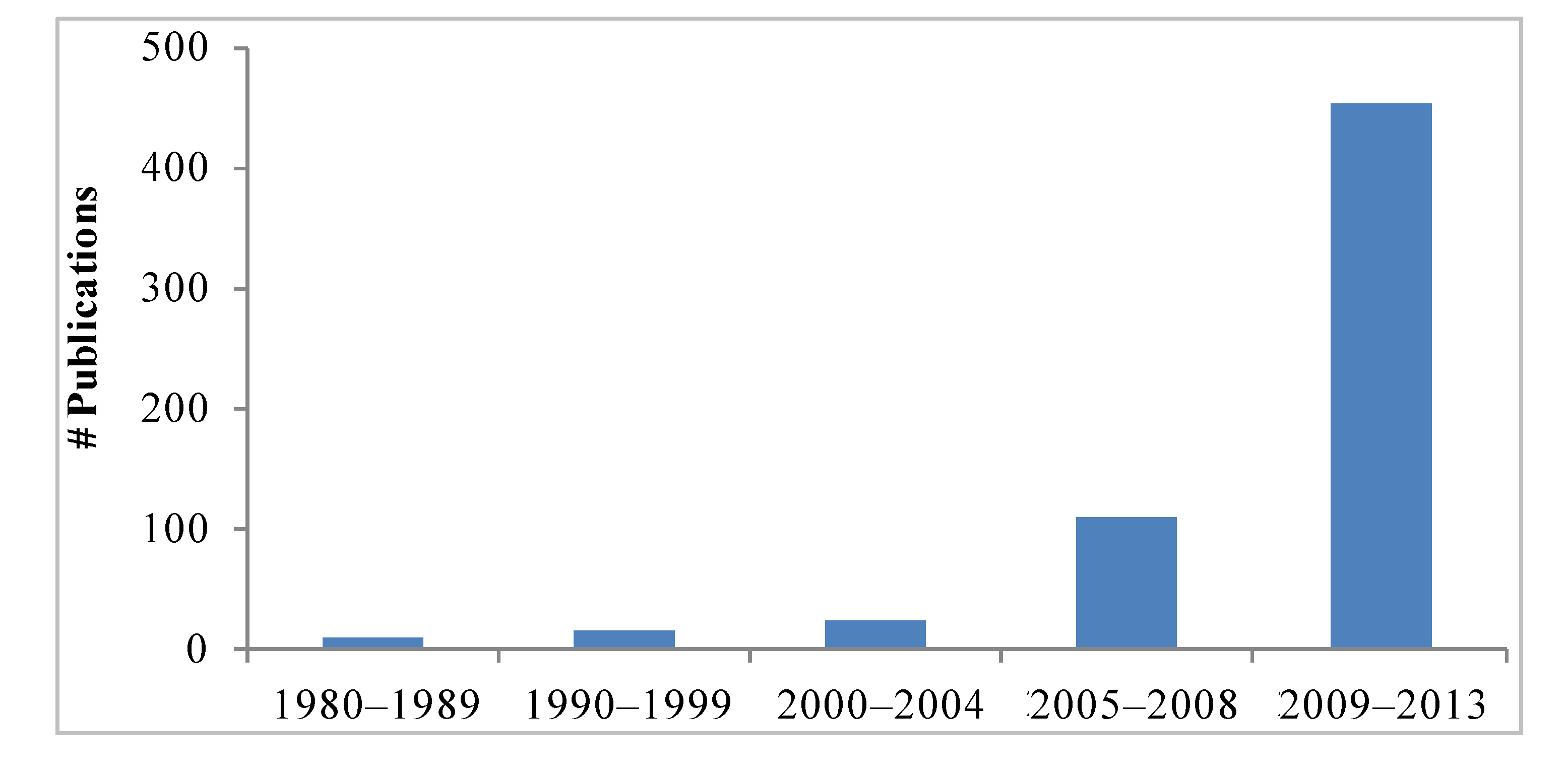
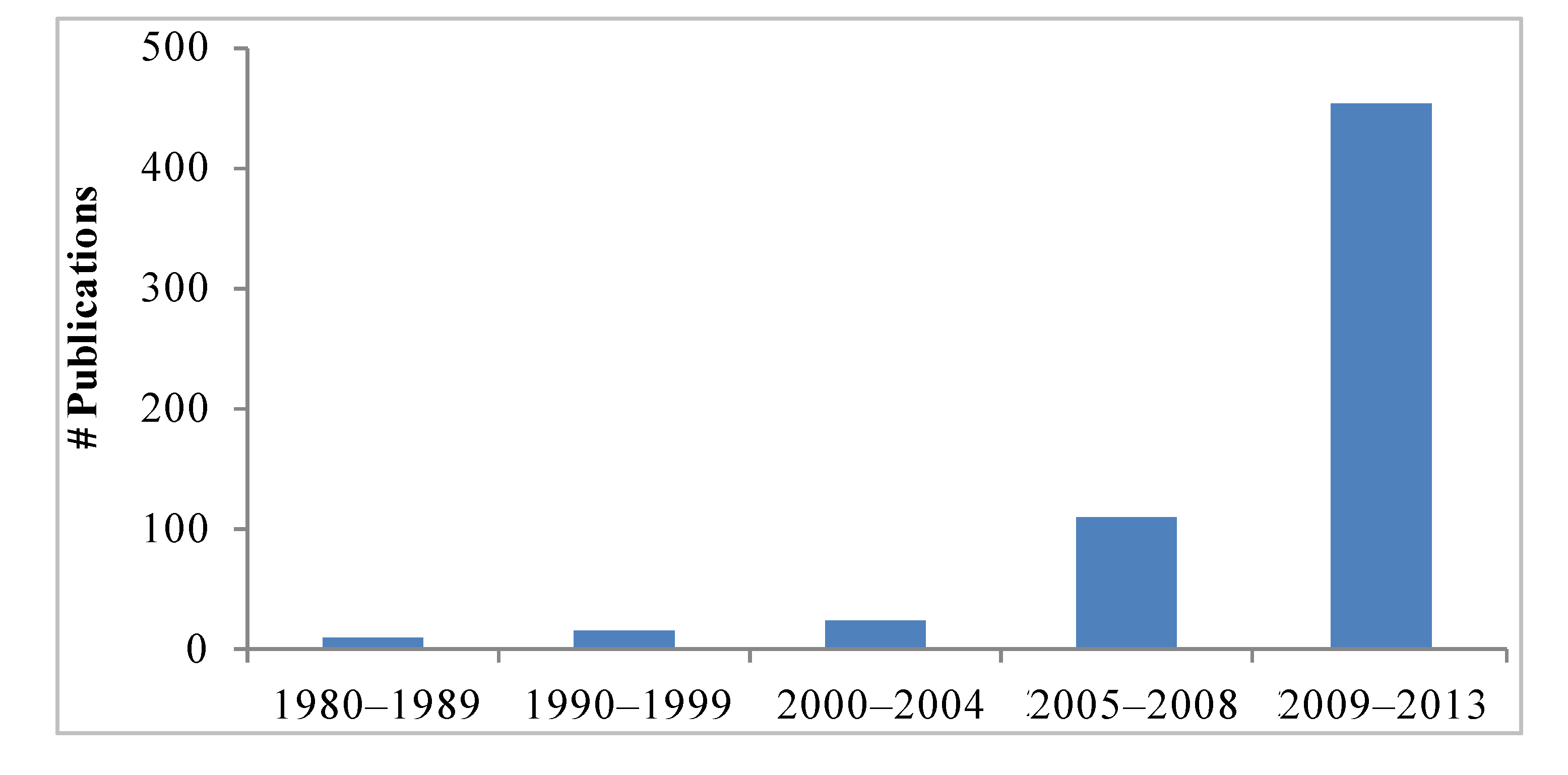
Interest in the mangosteen fruit and xanthones has greatly increased in recent years as readily demonstrated by the number of scientific reports. A search of available literature using mangosteen and xanthones as terms in Pubmed, Science Direct, Google Scholar, and Scirus, retrieved 158 reports in the period of 1980–2008. In contrast, there have been 454 published articles from 2008 through March 2013 (
Figure 2). By far, the most studied xanthone is α-mangostin (α-MG) for which anti-oxidant, anti-proliferative, pro-apoptotic, anti-inflammatory, anti-carcinogenic, and anti-microbial activities have been reported. Pertinent literature has been previously reviewed (
Table 1). In this review, we focus primarily on recent reports considering the bioavailability and cellular metabolism of xanthones, their anti-cancer and anti-inflammatory activities, and their reported effects on cellular signaling pathways.
Figure 2.
Number of publications related to mangosteen and their xanthones from 1980 to 2013. Search words: mangosteen, xanthones. Search performed on April 24 2013, including ahead of print publications. Databases: Pubmed, Science Direct, Google Scholar, Scirus.
Figure 2.
Number of publications related to mangosteen and their xanthones from 1980 to 2013. Search words: mangosteen, xanthones. Search performed on April 24 2013, including ahead of print publications. Databases: Pubmed, Science Direct, Google Scholar, Scirus.
3. Anti-Cancer Activities of Xanthones
In vivo studies examining the anti-tumorigenic activities and
in vitro anti-proliferative and pro-apoptotic activities of mangosteen xanthones with cancer cell lines are summarized in
Table 2 and
Table 3. The effects of mangosteen xanthones on mammary cancer have been examined in two studies using mammary BJMC3879 cancer cells xenografted into Balb/c mice. Subcutaneous α-MG [
17] and dietary Panaxanthone (75%–85% α-MG and 5%–15% γ-MG) [
18] significantly suppressed tumor volumes and metastastic expansion in this cancer model.
In vitro, α-MG induced apoptosis, cell cycle arrest, activation of caspases-3 and -9, cytochrome c release and the loss of mitochondrial potential in BJMC3879 cells [
17,
18].
Two recent reports addressed the anti-tumorigenic effects of α-MG in glioblastoma and prostate xenograft mouse models. Intraperitoneal treatment with α-MG inhibited tumor growth by 50% in a GBM8401 glioblastoma xenograft model and this effect was associated with increased phosphorylation of AMPK (AMP-activated protein kinase) and induction of autophagy [
19]. Oral administration of α-MG to athymic mice bearing 22Rv1 prostate tumors five times a week following cancer cell implantation significantly decreased tumor volume.
In vitro, α-MG induced cell cycle arrest and apoptosis in 22Rv1 prostate cancer cells through activation of caspase-3. By using a cell free assay, α-MG also was shown to inhibit cyclin/cyclin-dependent kinase 4, which is involved in cell cycle progression [
20].
The majority of
in vivo studies examining the anti-cancer activity of mangosteen xanthones have focused on colon cancer. Dietary administration of α-MG significantly inhibited the induction and development of aberrant crypt foci (ACF) in a chemically-induced rat model of colon carcinogenesis. Less dysplasia, fewer lesions and decreased cell proliferation were also detected in α-MG-treated rats [
21]. The growth of COLO 205 xenografts was completely suppressed when mice were injected intratumorally with 3 mg of a mangosteen extract containing α- and γ-MG. Caspase-mediated apoptosis was detected in the tumor cells. Lower doses of the extract also reduced tumor volume. Induction of COLO 205 cell apoptosis was also confirmed
in vitro [
22]. Dietary administration of an extract from mangosteen pericarp containing α- and γ-MG inhibited the growth of colorectal HCT116 xenografts in mice.
In vitro, α-MG reduced HCT116 cell viability and induced caspase activation and loss of mitochondrial potential. In addition, mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK), Myc/Max and p53 signaling was enhanced by 71%, 48% and 30%, respectively, after treatment of cells with α-MG. An increase in Jun
N-terminal kinase (JNK) pathway was observed, although the change failed to achieve statistical significance. Nuclear factor kappa-B (NF-κB) activity also was reduced by 30% [
23]. Balb/c mice bearing colon cancer NL-17 xenografts showed 50%–70% reduction in tumor size when intraperitoneally treated with an extract from mangosteen pericarp containing 25% α-MG. Anti-proliferative activity of the extract on NL-17 cells was also confirmed
in vitro [
24]. Oral administration of α-MG also reduced growth of colon cancer Her2/CT26 xenografts in mice. The anti-tumor effect of α-MG was ascribed to autophagic activation rather than induction of endoplasmic reticulum stress as the xanthone was found to activate autophagy in the small intestine [
25]. Finally, dietary α-MG reduced tumor mass of colon cancer HT-29 xenografts. In this study, xanthones and their metabolites were detected in serum, tumor, liver, and feces of these mice.
In vitro analysis confirmed that α-MG inhibited HT-29 proliferation and decreased BcL-2 and β-catenin expression [
13].
Table 2.
In vivo anti-tumorigenic activities of mangosteen xanthones.
Table 2.
In vivo anti-tumorigenic activities of mangosteen xanthones.
| Cancer cell type | Animal model | Tested compound | Delivery route | Dose | Outcomes | Reference |
|---|
| BJMC3879 (murine mammary adenocarcinoma) | Balb/c | panaxanthone (75%–85% α-MG, 5%–15% γ-MG) | diet | 5000 ppm | suppression of tumor volume and lung metastasis; decreased microvessel density | [18] |
| BJMC3879 (murine mammary adenocarcinoma) | Balb/c | α-MG | subcutaneous | 20 mg/kg/day | decreased tumor growth and metastatic expansion; increased apoptosis; activation of caspase-3; decreased microvessel density; cytochrome c release from mitochondria; cell cycle arrest | [17] |
| GBM8401 (human malignant glioblastoma) | nude Balb/cA-ν (ν/ν) | α-MG | intraperitoneal | 2 mg/kg/day | inhibition of tumor growth by 50%; increased phosphorylation of AMPK; induction of autophagy | [19] |
| 22Rv1 (human prostate carcinoma) | Athymic nu/nu mice | α-MG | oral gavage | 100 mg/kg 5x/week | decreased tumor growth | [20] |
| COLO205 (human colorectal adenocarcinoma) | Athymic NCr nu/nu mice | mangosteen pericarp extract containing 48 mg α-MG and 6.4 mg γ-MG per gram of extract | intratumorally | 0.024–3.0 mg per tumor | complete suppression of tumor growth at 3 mg extract/tumor; apoptotic cells, nuclear fragmentation and chromatin condensation; activation of caspases-3 and -8 | [22] |
| HCT116 (human colorectal carcinoma) | Athymic NCR nu/nu nude mice | extract of mangosteen pericarp (81% α-MG and 16% γ-MG) | diet | 0.25% and 0.5% extract: food ratio (wt/wt) | inhibition of tumor growth; fewer blood vessels in tumor | [23] |
| NL-17 (murine colon adenocarcinoma) | Balb/c | pericarp methanolic extract (25% α-MG) | intraperitoneal | 100–200 mg/kg | reduced tumor mass by 50%–70% | [24] |
| Her2/CT26 cells (murine colon carcinoma) | Balb/c | α-MG | oral | 20 mg/kg | reduced subcutaneous growth | [25] |
| HT-29 (human colon adenocarcinoma) | Athymic Balb/c | α-MG | diet | 900 mg/kg | 40% reduction in tumor mass; decreased Bcl-2 and β-catenin | [13] |
| Chemically induced cancer model |
| Chemically-induced (1,2dimethylhydra-zine) colon cancer | F344 rats | α-MG | diet | 0.02% and 0.05% in CE-2 basal diet | inhibition of induction and development of ACF; decreased dysplastic foci and β-catenin accumulated crypts; lower proliferating cell nuclear antigen in colon | [21] |
Table 3.
In vitro pro-apoptotic and anti-proliferative activities of mangosteen xanthones.
Table 3.
In vitro pro-apoptotic and anti-proliferative activities of mangosteen xanthones.
| Cell type | Tested compound | Dose | Outcomes | Reference |
|---|
| BJMC3879 (murine mammary adenocarcinoma) | α-MG | 8 µM | induction of apoptosis; cell cycle arrest; activation of caspase-3 and -9; loss of mitochondrial potential | [18] |
| PC3, and 22Rv1 (human prostate carcinoma) | α-MG | 2.5–15 µM | suppressed cell viability and colony formation; cell cycle arrest; activation of caspase-3 | [20] |
| COLO205 (human colorectal adenocarcinoma) | mangosteen extract: 48 mg α-MG and 6.40 mg γ-MG/g extract | 30 µg/mL | induction of apoptosis; activation of caspase-3 and -8; release of mitochondrial cytochrome c | [22] |
| HCT116 (human colorectal carcinoma) | extract of mangosteen pericarp (81% α-MG and 16% γ-MG) | 10–20 µg/mL | reduced cell viability; increased activities of caspase-3/7 and-9; loss of mitochondrial potential; enhanced activity of MAPK/ERK, Myc/Max and p53 signaling; increased JNK; decreased NF-κB | [23] |
| NL-17 (murine colon adenocarcinoma) | pericarp methanol extract (25% α-MG) | >25 µg/mL | anti-proliferative activity | [24] |
| HT-29 (human colon adenocarcinoma) | α-MG | 6–12 µM | anti-proliferative activity; decreased Bcl2 and β-catenin | [13] |
4. Anti-Inflammatory Activity of Xanthones
The reported
in vitro anti-inflammatory activities of mangosteen xanthones are summarized in
Table 4. α-MG attenuated lipopolysaccharide (LPS)-induced expression of inflammatory mediators such as tumor necrosis factor α (TNF-α) and interleukin (IL-) 6 in human U937macrophage-like cells. α-MG also decreased activation of several signaling pathways including IL-1, mitogen-activated protein kinase kinase (MEK), JNK, ERK, signal transducer and activator of transcription 1 (STAT-1), and activator protein 1 (AP-1) in these cells [
26,
27]. Concentrations of α-MG used in these studies ranged from 6 to 12 nM [
27] to 10–30 µM [
26] and the LPS insult also differed in these reports. Inhibition of activation of MAPK, NF-κB, and AP-1 and attenuation of expression of pro-inflammatory cytokine genes also was observed in LPS-stimulated primary human adipocytes in response to α-MG treatment [
7,
28].
We recently examined the inhibitory effects of α-MG on the secretion of pro-inflammatory mediators by transformed and primary human cells. α-MG inhibited the secretion of IL-8 or TNF-α by human cell lines from various tissue origins challenged with a pro-inflammatory insult. Surprisingly, α-MG further stimulated the basal and LPS-stimulated secretion of TNF-α in primary cultures of human monocyte-derived macrophages cells [
8].
α- and γ-MG inhibited nitric oxide (NO) and prostaglandin E
2 (PGE
2) production in murine RAW 264.7 macrophages. These effects were associated with reduced amounts of iNOS inducible NO synthase (iNOS) and cyclooxygenase-2 (COX-2) mRNA [
29,
30]. Suppression of histamine release by α-, β- and γ-MG was observed in IgE-sensitized rat basophilic leukemia RBL-2H3 cells [
31]. γ-MG also dose dependently inhibited basal and A23187-induced release of PGE
2 in C6 rat glioma cells [
32]. These effects were associated with reduced COX-2 mRNA and protein expression, and NF-κB activation. Garcinone B also had similar effects in C6 cells by interfering with activation of NF-κB [
33]. Contrary to these reports, an aqueous extract containing polyphenolic compounds from mangosteen pericarp stimulated the inflammatory response in cultures of Caco-2 cells treated with IL-1β [
34]. This difference may be due to the absence of the hydrophobic xanthones in the extract.
The
in vivo anti-inflammatory activities of mangosteen xanthones are summarized in
Table 5. Early studies showed that both intraperitoneal and oral administration of α-MG, 1-isomangostin, or mangostin triacetate had anti-inflammatory activities in several rat models of inflammation [
35]. The
in vivo anti-inflammatory activity of γ-MG has been confirmed in the carrageenan-induced hind paw edema model in rats when the xanthone was administered intraperitoneally 30 minutes prior to inflammatory insult [
36]. α-MG exhibited similar anti-inflammatory effects with this model [
37]. Oral administration of α-MG also inhibited paw edema formation in mice [
30]. Orally administered α- and γ-MG also exhibited anti-inflammatory activity in a mouse model of ovalbumin (OVA)-induced allergic asthma. Both xanthones had similar efficacy [
38].
Information about the anti-inflammatory activity of mangosteen xanthones in humans is limited to three reports. Topical application of a gel containing extract of mangosteen pericarp decreased periodontal inflammation suggesting that the formulation may be useful as an adjuvant. However, the xanthone content and composition in the gel was not reported for this study [
39]. Ingestion of a blended mangosteen juice decreased serum C-reactive protein (CRP) levels. However, other markers of inflammation were increased in subjects consuming the mangosteen product compared to placebo [
40]. It was also reported that CRP levels in obese subjects consuming 18 oz of a mangosteen juice blend per day for 8 weeks were lower than those in the placebo group. However, levels of the pro-inflammatory interferon-inducible protein 10 (IP-10) and macrophage inflammatory protein-1 β (MIP-1 β) were increased in subjects consuming the high volumes of mangosteen juice blend [
41].
6. Future Research
Two human studies have reported that ingestion of a mangosteen juice blend or a xanthone-rich mangosteen product decreased serum CRP levels and increased ORAC values, respectively [
15,
41]. However, increased levels of several pro-inflammatory mediators were also observed. Results from these studies should be considered with caution as there is no way to discriminate the effects of other components in these products. Although no adverse events were reported in these trials, the potential long term toxicity of products containing mangosteen xanthones requires assessment.
With the exception of our study on primary human monocyte-derived macrophages [
8], the effect of mangosteen xanthones on cultured normal cells has not been addressed. Our preliminary data suggest that primary and transformed human cells respond differently to α-MG with the xanthone promoting the secretion of pro-inflammatory mediators by normal cells. It is expected that cell phenotype will affect the response to bioactive compounds but whether this difference is limited to the inflammatory response in human cells is not clear. α-MG has been shown to promote apoptosis in cancer cells but this activity has not been evaluated in cells with a non-cancerous phenotype. Differences between normal and cancer cells may also be explained by the degree of metabolism and types of metabolic products. Our results also showed that while transformed human cell lines convert α-MG to phase II metabolites and other xanthones, only phase II metabolites are detected in primary cell cultures.
The vast majority of studies have focused on the anti-cancer and anti-inflammatory activities of mangosteen xanthones, and particularly α-MG. Our work and that of others has shown that α-MG and other xanthones are metabolized by human and animal cells. Thus, the possibility of metabolites of xanthones exerting some of the observed effects cannot be ruled out. It has been reported that metabolites of various phytochemicals possess the observed bioactivities [
54,
55,
56,
57]. Further investigation is needed to elucidate whether individual metabolites and/or their combinations possess some of the bioactivities primarily attributed to mangosteen xanthones. Similarly, more pharmacokinetic studies are needed to assess the bioavailability of xanthones from mangosteen containing beverages and food products.
One aspect of
in vitro studies that is often neglected is the stability of the compound in cell culture conditions. Many polyphenolic compounds readily react with components of cell culture media to generate H
2O
2, quinones and semiquinonescapable of inducing alterations in cellular activities [
58,
59]. Studies on our laboratory showed that α-MG was readily degraded in different types of serum-free media (
i.e., RPMI, DMEM, MEM and McCoy’s 5A) when added in a dimethyl sulfoxide (DMSO) stock solution [
8]. We stabilized the xanthone by delivering DMSO-solubilized α-MG to media containing serum or by delivering the pure compound in Tween 40 micelles. Previous studies of the biological activities of α-MG have generally used DMSO as delivery vehicle [
29,
30], although it is unclear if serum was present during experiment.
Although the bioavailability of mangosteen xanthones is limited as it is for many phytochemicals, the gastrointestinal (GI) tract is exposed to high concentrations of these compounds and their metabolites. Consequently, we are currently examining the effects of dietary α-MG in a mouse model of ulcerative colitis. It is possible that the bioactivities of xanthones and/or metabolites may differ depending on their concentrations in biofluids and cells. If activities are mediated by inducing an adaptive stress or “hormetic” response [
60,
61], higher concentrations of these compounds (such as those found in the GI tract) may not necessarily be beneficial. Finally, consideration of the reported anti-microbial activity of xanthones on the gut microbiome merits consideration.
Despite the numerous in vitro and in vivo studies on the bioactivities of mangosteen xanthones, more research is needed to evaluate their safety and health benefits before they can be recommended for preventive or therapeutic purposes.



{kind=link}
{kind=link}
{kind=link}
{kind=link}