The Role of Lutein in Eye-Related Disease

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
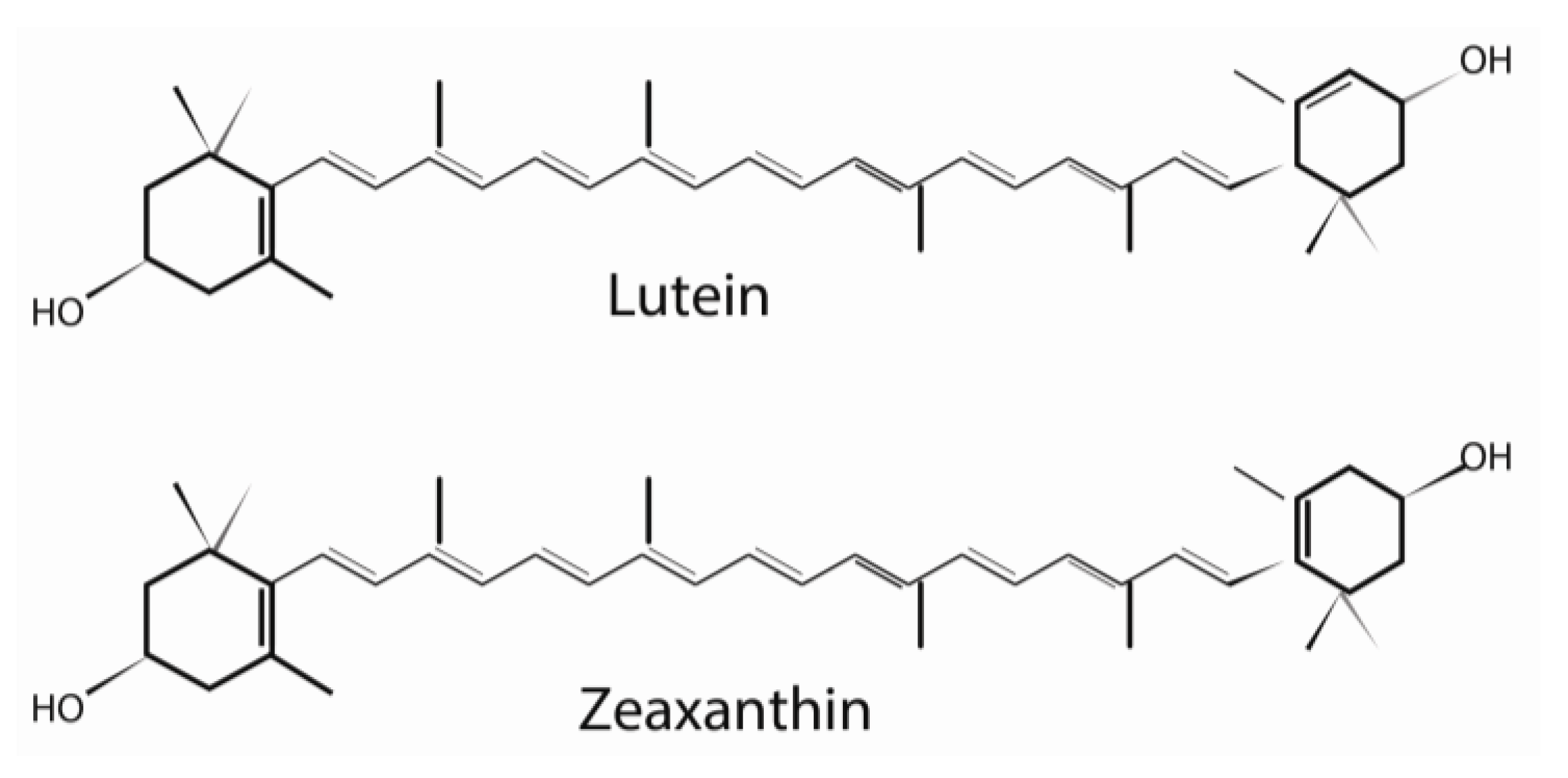
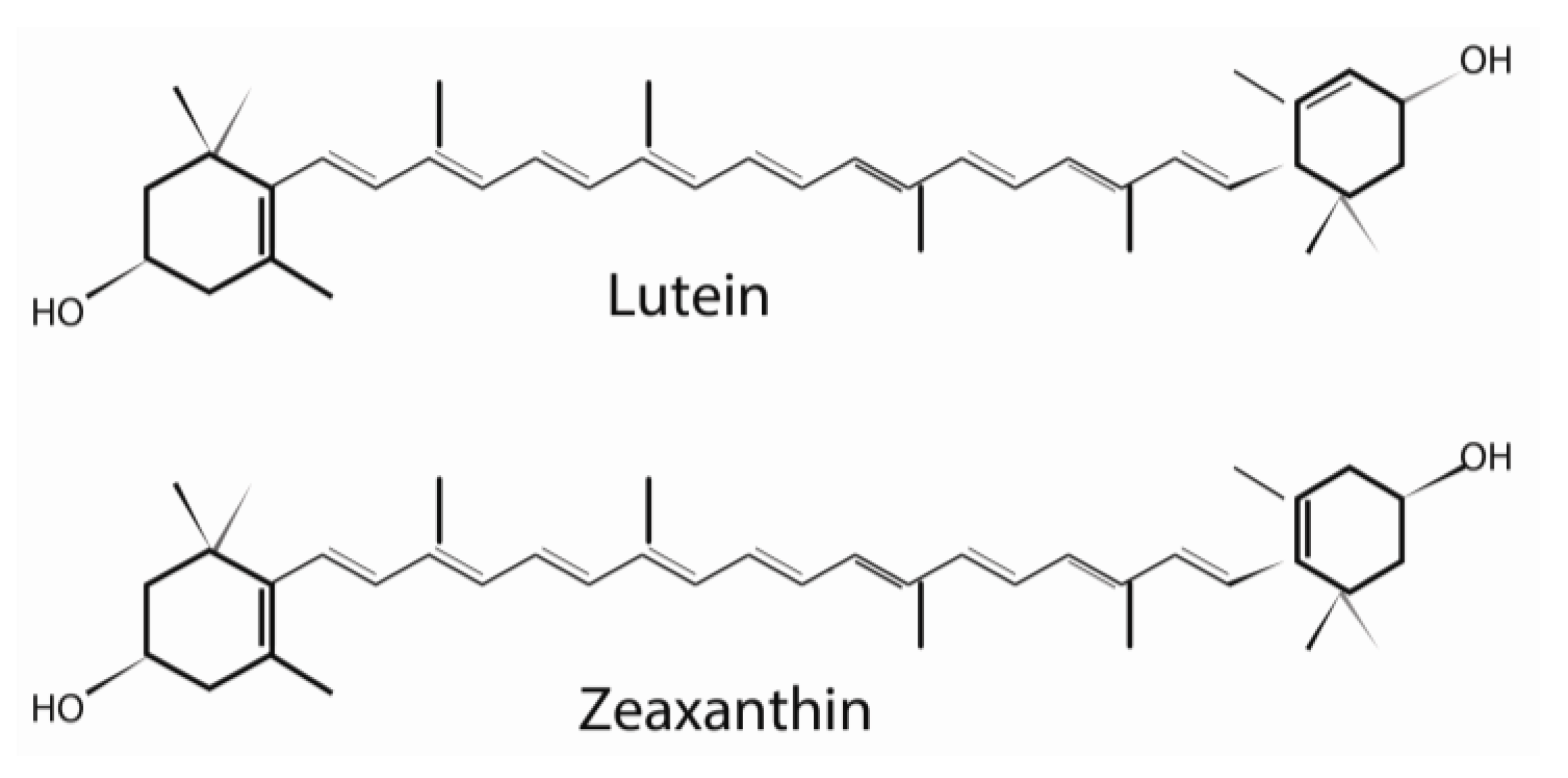
2. Structure and Biochemistry of Lutein
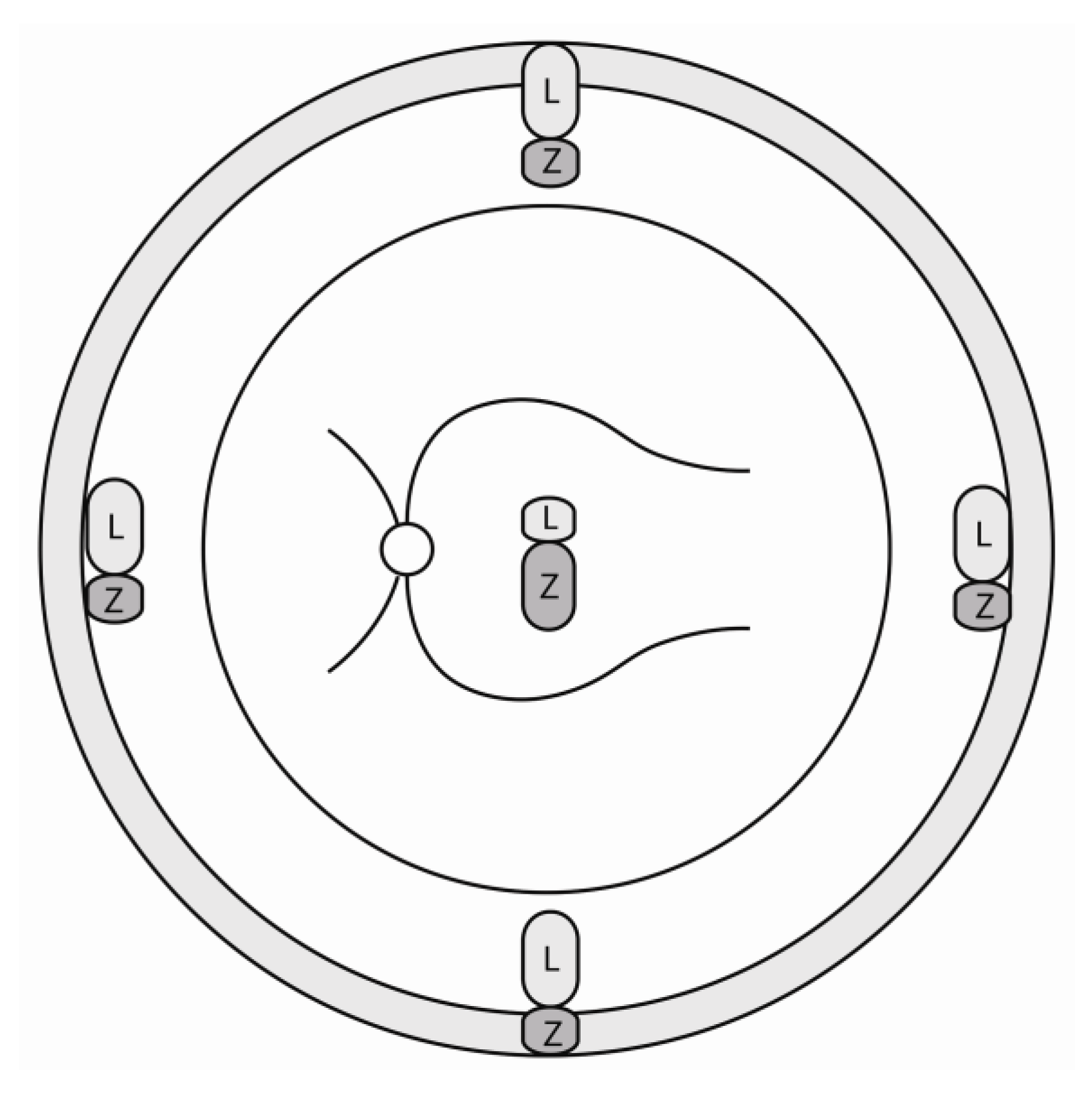
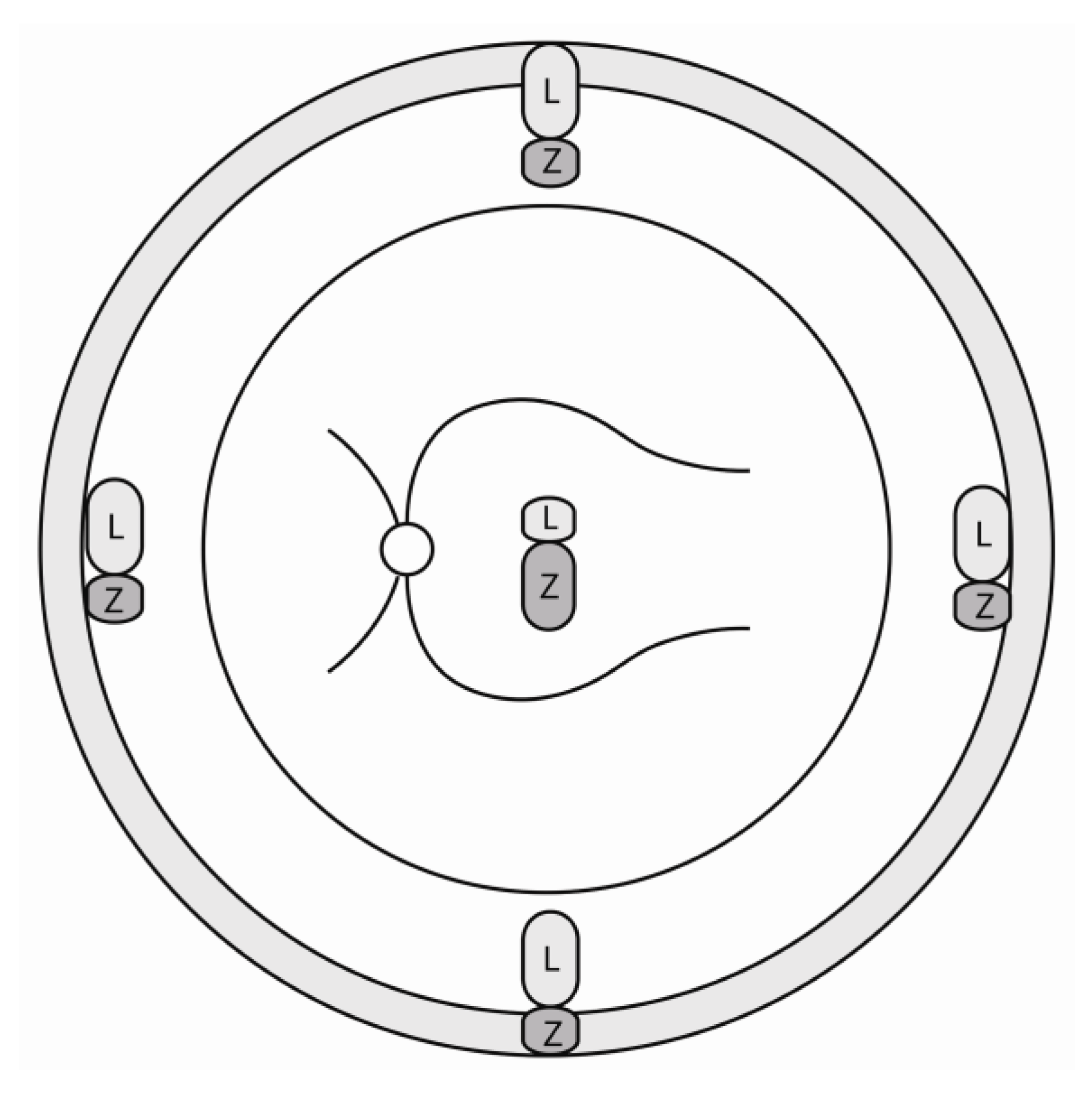
2.1. Lutein in the Retina
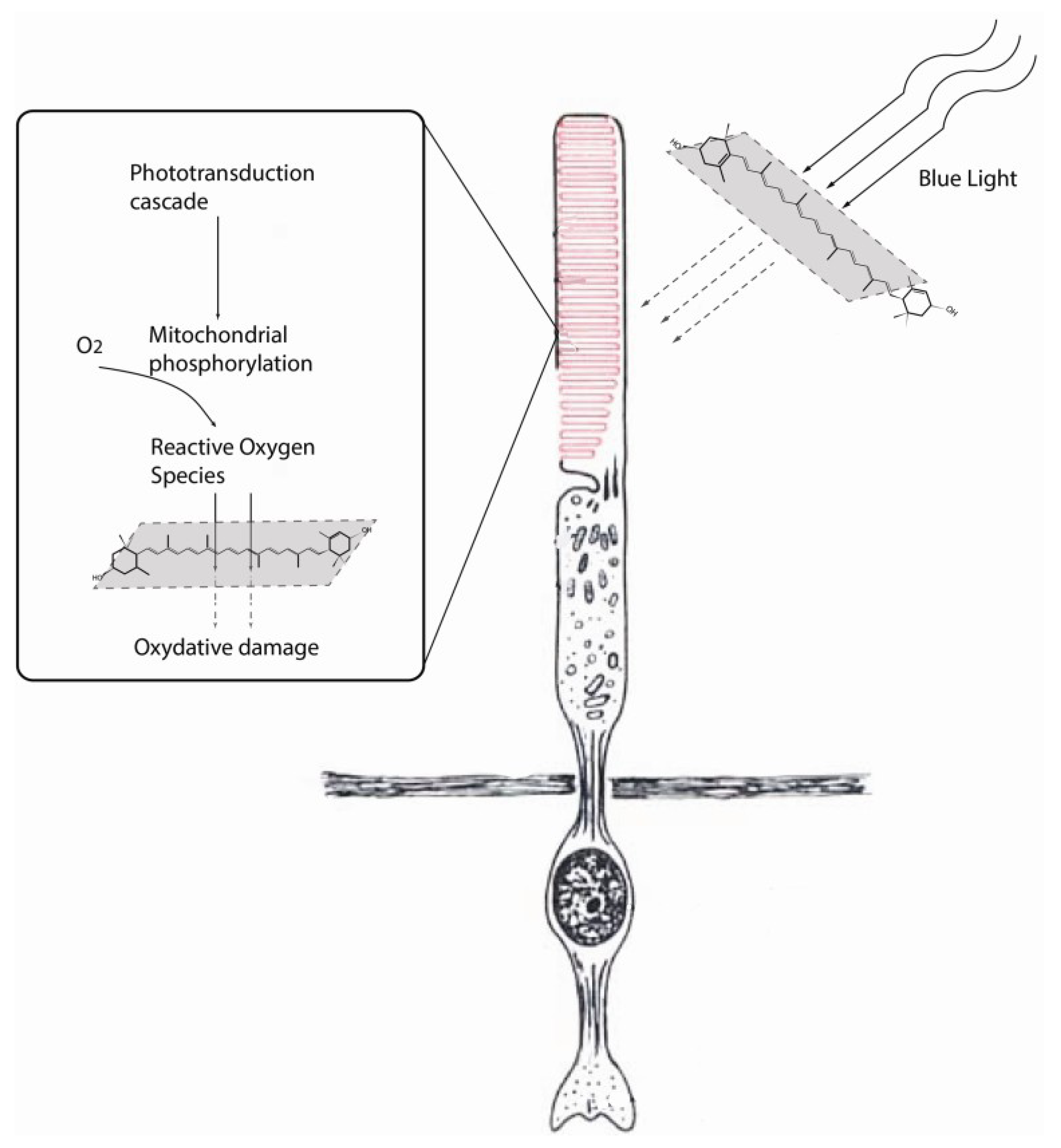
2.2. Role of Lutein in Prevention of Hypoxia-Induced Cell Damage in the Eye
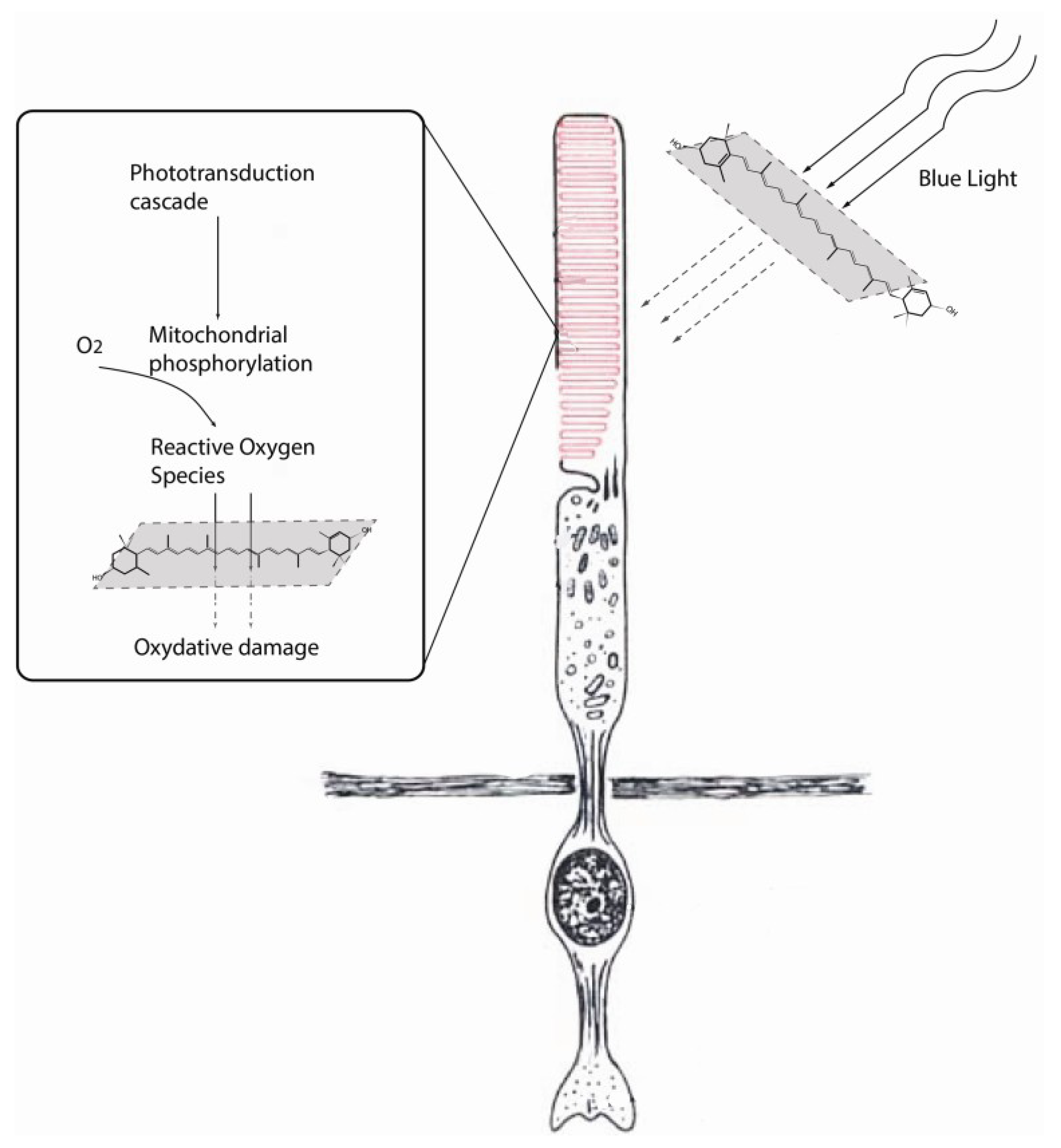
3. Role of Lutein in Age-Related Macular Degeneration (AMD)
3.1. Experimental Studies
3.2. Epidemiological and Clinical Studies
4. Lutein Role in Diabetic Retinopathy
4.1. Experimental Studies
4.2. Epidemiological and Clinical Studies
5. Lutein in Retinal Detachment
5.1. Experimental Studies
5.2. Clinical Studies
5.3. Lutein in the Lens
5.4. Experimental Studies
5.5. Epidemiological and Clinical Studies
6. Lutein and the Uvea
6.1. Experimental Studies
6.2. Epidemiological and Clinical Studies
7. Lutein Supplementation
8. Discussion and Conclusions
References
- Snodderly, D.M. Evidence for protection against age-related macular degeneration by carotenoids and antioxidant vitamins. Am. J. Clin. Nutr. 1995, 62, 1448S–1461S. [Google Scholar]
- Subczynski, W.K.; Wisniewska, A.; Widomska, J. Location of macular xanthophylls in the most vulnerable regions of photoreceptor outer-segment membranes. Arch. Biochem. Biophys. 2010, 504, 61–66. [Google Scholar]
- Trumbo, P.R.; Ellwood, K.C. Lutein and zeaxanthin intakes and risk of age-related macular degeneration and cataracts: An evaluation using the Food and Drug Administration’s evidence-based review system for health claims. Am. J. Clin. Nutr. 2006, 84, 971–974. [Google Scholar]
- Alves-Rodrigues, A.; Shao, A. The science behind lutein. Toxicol. Lett. 2004, 150, 57–83. [Google Scholar]
- Loane, E.; McKay, G.J.; Nolan, J.M.; Beatty, S. Apolipoprotein E genotype is associated with macular pigment optical density. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2636–2643. [Google Scholar] [CrossRef] [Green Version]
- qRenzi, L.M.; Hammond, B.R., Jr.; Dengler, M.; Roberts, R. The relation between serum lipids and lutein and zeaxanthin in the serum and retina: Results from cross-sectional, case-control and case study designs. Lipids Health Dis. 2012, 11, 33. [Google Scholar] [CrossRef]
- Wisniewska, A.; Draus, J.; Subczynski, W.K. Is a fluid-mosaic model of biological membranes fully relevant? Studies on lipid organization in model and biological membranes. Cell. Mol. Biol. Lett. 2003, 8, 147–159. [Google Scholar]
- Nolan, J.; O’Donovan, O.; Kavanagh, H.; Stack, J.; Harrison, M.; Muldoon, A.; Mellerio, J.; Beatty, S. Macular pigment and percentage of body fat. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3940–3950. [Google Scholar] [CrossRef]
- Riso, P.; Porrini, M. Determination of carotenoids in vegetable foods and plasma. Int. J. Vitam. Nutr. Res. 1997, 67, 47–54. [Google Scholar]
- Bhosale, P.; Bernstein, P.S. Vertebrate and invertebrate carotenoid-binding proteins. Arch. Biochem. Biophys. 2007, 458, 121–127. [Google Scholar] [CrossRef]
- Lee, B.L.; New, A.L.; Ong, C.N. Simultaneous determination of tocotrienols, tocopherols, retinol, and major carotenoids in human plasma. Clin. Chem. 2003, 49, 2056–2066. [Google Scholar] [CrossRef]
- Jenkins, M.Y.; Mitchell, G.V.; Grundel, E. Natural tocopherols in a dietary supplement of lutein affect tissue distribution of tocopherols in young rats. Nutr. Cancer 2000, 37, 207–214. [Google Scholar] [CrossRef]
- Snodderly, D.M.; Auran, J.D.; Delori, F.C. The macular pigment. II. Spatial distribution in primate retinas. Investig. Ophthalmol. Vis. Sci. 1984, 25, 674–685. [Google Scholar]
- Sommerburg, O.G.; Siems, W.G.; Hurst, J.S.; Lewis, J.W.; Kliger, D.S.; van Kuijk, F.J. Lutein and zeaxanthin are associated with photoreceptors in the human retina. Curr. Eye Res. 1999, 19, 491–495. [Google Scholar] [CrossRef]
- Rapp, L.M.; Maple, S.S.; Choi, J.H. Lutein and zeaxanthin concentrations in rod outer segment membranes from perifoveal and peripheral human retina. Investig. Ophthalmol. Vis. Sci. 2000, 41, 1200–1209. [Google Scholar]
- Bernstein, P.S.; Khachik, F.; Carvalho, L.S.; Muir, G.J.; Zhao, D.Y.; Katz, N.B. Identification and quantitation of carotenoids and their metabolites in the tissues of the human eye. Exp. Eye Res. 2001, 72, 215–223. [Google Scholar] [CrossRef]
- Khachik, F.; de Moura, F.F.; Zhao, D.Y.; Aebischer, C.P.; Bernstein, P.S. Transformations of selected carotenoids in plasma, liver, and ocular tissues of humans and in nonprimate animal models. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3383–3392. [Google Scholar]
- Seddon, J.M.; Ajani, U.A.; Sperduto, R.D.; Hiller, R.; Blair, N.; Burton, T.C.; Farber, M.D.; Gragoudas, E.S.; Haller, J.; et al. Dietary carotenoids, vitamins A, C, and E, and advanced age-related macular degeneration. Eye Disease Case-Control Study Group. JAMA 1994, 272, 1413–1420. [Google Scholar]
- Ma, L.; Yan, S.F.; Huang, Y.M.; Lu, X.R.; Qian, F.; Pang, H.L.; Xu, X.R.; Zou, Z.Y.; Dong, P.C.; Xiao, X.; et al. Effect of lutein and zeaxanthin on macular pigment and visual function in patients with early age-related macular degeneration. Ophthalmology 2012, 119, 2290–2297. [Google Scholar] [CrossRef]
- Loughman, J.; Nolan, J.M.; Howard, A.N.; Connolly, E.; Meagher, K.; Beatty, S. The impact of macular pigment augmentation on visual performance using different carotenoid formulations. Investig. Ophthalmol. Vis. Sci. 2012, 53, 7871–7880. [Google Scholar] [CrossRef]
- Ma, L.; Dou, H.L.; Huang, Y.M.; Lu, X.R.; Xu, X.R.; Qian, F.; Zou, Z.Y.; Pang, H.L.; Dong, P.C.; Xiao, X.; et al. Improvement of retinal function in early age-related macular degeneration after lutein and zeaxanthin supplementation: A randomized, double-masked, placebo-controlled trial. Am. J. Ophthalmol. 2012, 154, 625–634. [Google Scholar] [CrossRef]
- Luchsinger, J.A.; Tang, M.X.; Shea, S.; Mayeux, R. Antioxidant vitamin intake and risk of Alzheimer disease. Arch. Neurol. 2003, 60, 203–208. [Google Scholar] [CrossRef]
- Ramassamy, C.; Averill, D.; Beffert, U.; Bastianetto, S.; Theroux, L.; Lussier-Cacan, S.; Cohn, J.S.; Christen, Y.; Davignon, J.; Quirion, R.; et al. Oxidative damage and protection by antioxidants in the frontal cortex of Alzheimer’s disease is related to the apolipoprotein E genotype. Free Radic. Biol. Med. 1999, 27, 544–553. [Google Scholar] [CrossRef]
- Trieschmann, M.; Spital, G.; Lommatzsch, A.; van Kuijk, E.; Fitzke, F.; Bird, A.C.; Pauleikhoff, D. Macular pigment: Quantitative analysis on autofluorescence images. Graefes Arch. Clin. Exp. Ophthalmol. 2003, 241, 1006–1012. [Google Scholar] [CrossRef]
- Nolan, J.M.; Kenny, R.; O’Regan, C.; Cronin, H.; Loughman, J.; Connolly, E.E.; Kearney, P.; Loane, E.; Beatty, S. Macular pigment optical density in an ageing Irish population: The Irish Longitudinal Study on Ageing. Ophthalmic Res. 2010, 44, 131–139. [Google Scholar] [CrossRef]
- Ciulla, T.A.; Hammond, B.R., Jr. Macular pigment density and aging, assessed in the normal elderly and those with cataracts and age-related macular degeneration. Am. J. Ophthalmol. 2004, 138, 582–587. [Google Scholar] [CrossRef]
- Bone, R.A.; Landrum, J.T.; Mayne, S.T.; Gomez, C.M.; Tibor, S.E.; Twaroska, E.E. Macular pigment in donor eyes with and without AMD: A case-control study. Investig. Ophthalmol. Vis. Sci. 2001, 42, 235–240. [Google Scholar]
- Bone, R.A.; Landrum, J.T.; Dixon, Z.; Chen, Y.; Llerena, C.M. Lutein and zeaxanthin in the eyes, serum and diet of human subjects. Exp. Eye Res. 2000, 71, 239–245. [Google Scholar] [CrossRef]
- Bone, R.A.; Landrum, J.T.; Cao, Y.; Howard, A.N.; Alvarez-Calderon, F. Macular pigment response to a supplement containing meso-zeaxanthin, lutein and zeaxanthin. Nutr. Metab. (Lond.) 2007, 4, 12. [Google Scholar] [CrossRef]
- Bone, R.A.; Landrum, J.T.; Guerra, L.H.; Ruiz, C.A. Lutein and zeaxanthin dietary supplements raise macular pigment density and serum concentrations of these carotenoids in humans. J. Nutr. 2003, 133, 992–998. [Google Scholar]
- Stringham, J.M.; Hammond, B.R.; Nolan, J.M.; Wooten, B.R.; Mammen, A.; Smollon, W.; Snodderly, D.M. The utility of using customized heterochromatic flicker photometry (cHFP) to measure macular pigment in patients with age-related macular degeneration. Exp. Eye Res. 2008, 87, 445–453. [Google Scholar] [CrossRef]
- Zhao, L.; Sweet, B.V. Lutein and zeaxanthin for macular degeneration. Am. J. Health Syst. Pharm. 2008, 65, 1232–1238. [Google Scholar]
- Johnson, E.J.; Chung, H.Y.; Caldarella, S.M.; Snodderly, D.M. The influence of supplemental lutein and docosahexaenoic acid on serum, lipoproteins, and macular pigmentation. Am. J. Clin. Nutr. 2008, 87, 1521–1529. [Google Scholar]
- Bone, R.A.; Landrum, J.T.; Cains, A. Optical density spectra of the macular pigment in vivo and in vitro. Vis. Res. 1992, 32, 105–110. [Google Scholar] [CrossRef]
- Brindley, G.S.; Willmer, E.N. The reflexion of light from the macular and peripheral fundus oculi in man. J. Physiol. 1952, 116, 350–356. [Google Scholar]
- Bernstein, P.S.; Yoshida, M.D.; Katz, N.B.; McClane, R.W.; Gellermann, W. Raman detection of macular carotenoid pigments in intact human retina. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2003–2011. [Google Scholar]
- Bui, M.H. Simple determination of retinol, alpha-tocopherol and carotenoids (lutein, all-trans-lycopene, α- and β-carotenes) in human plasma by isocratic liquid chromatography. J. Chromatogr. B 1994, 654, 129–133. [Google Scholar] [CrossRef]
- Lee, N.; Wielaard, J.; Fawzi, A.A.; Sajda, P.; Laine, A.F.; Martin, G.; Humayun, M.S.; Smith, R.T. In vivo snapshot hyperspectral image analysis of age-related macular degeneration. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2010, 2010, 5363–5366. [Google Scholar]
- Leung, I.Y. Macular pigment: New clinical methods of detection and the role of carotenoids in age-related macular degeneration. Optometry 2008, 79, 266–272. [Google Scholar] [CrossRef]
- Bhosale, P.; Serban, B.; Zhao, da Y.; Bernstein, P.S. Identification and metabolic transformations of carotenoids in ocular tissues of the Japanese quail Coturnix japonica. Biochemistry 2007, 46, 9050–9057. [Google Scholar] [CrossRef]
- Bhosale, P.; Bernstein, P.S. Quantitative measurement of 3'-oxolutein from human retina by normal-phase high-performance liquid chromatography coupled to atmospheric pressure chemical ionization mass spectrometry. Anal. Biochem. 2005, 345, 296–301. [Google Scholar] [CrossRef]
- Khachik, F.; Carvalho, L.; Bernstein, P.S.; Muir, G.J.; Zhao, D.Y.; Katz, N.B. Chemistry, distribution, and metabolism of tomato carotenoids and their impact on human health. Exp. Biol. Med. (Maywood) 2002, 227, 845–851. [Google Scholar]
- Siems, W.G.; Sommerburg, O.; van Kuijk, F.J. Lycopene and β-carotene decompose more rapidly than lutein and zeaxanthin upon exposure to various pro-oxidants in vitro. Biofactors 1999, 10, 105–113. [Google Scholar] [CrossRef]
- Panfoli, I.; Calzia, D.; Ravera, S.; Morelli, A.M.; Traverso, C.E. Extra-mitochondrial aerobic metabolism in retinal rod outer segments: New perspectives in retinopathies. Med. Hypotheses 2012, 78, 423–427. [Google Scholar] [CrossRef]
- Panfoli, I.; Calzia, D.; Ravera, S.; Bruschi, M.; Tacchetti, C.; Candiani, S.; Morelli, A.; Candiano, G. Extramitochondrial tricarboxylic acid cycle in retinal rod outer segments. Biochimie 2011, 93, 1565–1575. [Google Scholar] [CrossRef]
- Bone, R.A.; Landrum, J.T. Macular pigment in Henle fiber membranes: A model for Haidinger’s brushes. Vis. Res. 1984, 24, 103–108. [Google Scholar] [CrossRef]
- Landrum, J.; Bone, R.; Mendez, V.; Valenciaga, A.; Babino, D. Comparison of dietary supplementation with lutein diacetate and lutein: A pilot study of the effects on serum and macular pigment. Acta Biochim. Pol. 2012, 59, 167–169. [Google Scholar]
- Qin, L.; Bartlett, H.; Griffiths, H.R.; Eperjesi, F.; Armstrong, R.A.; Gherghel, D. Macular pigment optical density is related to blood glutathione levels in healthy individuals. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5029–5033. [Google Scholar] [CrossRef]
- Conn, P.F.; Schalch, W.; Truscott, T.G. The singlet oxygen and carotenoid interaction. J. Photochem. Photobiol. B 1991, 11, 41–47. [Google Scholar] [CrossRef]
- Foote, C.S.; Chang, Y.C.; Denny, R.W. Chemistry of singlet oxygen. X. Carotenoid quenching parallels biological protection. J. Am. Chem. Soc. 1970, 92, 5216–5218. [Google Scholar] [CrossRef]
- Nilsson, S.E.; Sundelin, S.P.; Wihlmark, U.; Brunk, U.T. Aging of cultured retinal pigment epithelial cells: oxidative reactions, lipofuscin formation and blue light damage. Doc. Ophthalmol. 2003, 106, 13–16. [Google Scholar] [CrossRef]
- Snodderly, D.M.; Russett, M.D.; Land, R.I.; Krinsky, N.I. Plasma carotenoids of monkeys (Macaca fascicularis and Saimiri sciureus) fed a nonpurified diet. J. Nutr. 1990, 120, 1663–1671. [Google Scholar]
- Neuringer, M.; Sandstrom, M.M.; Johnson, E.J.; Snodderly, D.M. Nutritional manipulation of primate retinas, I: Effects of lutein or zeaxanthin supplements on serum and macular pigment in xanthophyll-free rhesus monkeys. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3234–3243. [Google Scholar] [CrossRef]
- Huang, L.L.; Coleman, H.R.; Kim, J.; de Monasterio, F.; Wong, W.T.; Schleicher, R.L.; Ferris, F.L., III; Chew, E.Y. Oral supplementation of lutein/zeaxanthin and omega-3 long chain polyunsaturated fatty acids in persons aged 60 years or older, with or without AMD. Investig. Ophthalmol. Vis. Sci. 2008, 49, 3864–3869. [Google Scholar]
- Loane, E.; Nolan, J.M.; O’Donovan, O.; Bhosale, P.; Bernstein, P.S.; Beatty, S. Transport and retinal capture of lutein and zeaxanthin with reference to age-related macular degeneration. Surv. Ophthalmol. 2008, 53, 68–81. [Google Scholar] [CrossRef]
- Rehak, M.; Fric, E.; Wiedemann, P. Lutein and antioxidants in the prevention of age-related macular degeneration. Ophthalmologe 2008, 105, 37–38, 40–45. [Google Scholar]
- Lornejad-Schafer, M.R.; Lambert, C.; Breithaupt, D.E.; Biesalski, H.K.; Frank, J. Solubility, uptake and biocompatibility of lutein and zeaxanthin delivered to cultured human retinal pigment epithelial cells in tween40 micelles. Eur. J. Nutr. 2007, 46, 79–86. [Google Scholar] [CrossRef]
- Kalariya, N.M.; Ramana, K.V.; Srivastava, S.K.; van Kuijk, F.J. Genotoxic effects of carotenoid breakdown products in human retinal pigment epithelial cells. Curr. Eye Res. 2009, 34, 737–747. [Google Scholar] [CrossRef]
- Sasaki, M.; Ozawa, Y.; Kurihara, T.; Noda, K.; Imamura, Y.; Kobayashi, S.; Ishida, S.; Tsubota, K. Neuroprotective effect of an antioxidant, lutein, during retinal inflammation. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1433–1439. [Google Scholar]
- Li, S.Y.; Lo, A.C. Lutein protects RGC-5 cells against hypoxia and oxidative stress. Int. J. Mol. Sci. 2010, 11, 2109–2117. [Google Scholar] [CrossRef] [Green Version]
- Sundelin, S.P.; Nilsson, S.E.G. Lipofuscin-formation in retinal pigment epithelial cells is reduced by antioxidants. Free Radic. Biol. Med. 2001, 31, 217–225. [Google Scholar] [CrossRef]
- Zerbib, J.; Seddon, J.M.; Richard, F.; Reynolds, R.; Leveziel, N.; Benlian, P.; Borel, P.; Feingold, J.; Munnich, A.; Soubrane, G.; et al. rs5888 variant of SCARB1 gene is a possible susceptibility factor for age-related macular degeneration. PLoS One 2009, 4, e7341. [Google Scholar] [CrossRef]
- Izumi-Nagai, K.; Nagai, N.; Ohgami, K.; Satofuka, S.; Ozawa, Y.; Tsubota, K.; Umezawa, K.; Ohno, S.; Oike, Y.; Ishida, S. Macular pigment lutein is antiinflammatory in preventing choroidal neovascularization. Arterioscler. Thromb. Vasc. Biol. 2007, 27, 2555–2562. [Google Scholar] [CrossRef]
- Goldberg, J.; Flowerdew, G.; Smith, E.; Brody, J.A.; Tso, M.O. Factors associated with age-related macular degeneration. An analysis of data from the first National Health and Nutrition Examination Survey. Am. J. Epidemiol. 1988, 128, 700–710. [Google Scholar]
- Sommerburg, O.; Keunen, J.E.; Bird, A.C.; van Kuijk, F.J. Fruits and vegetables that are sources for lutein and zeaxanthin: The macular pigment in human eyes. Br. J. Ophthalmol. 1998, 82, 907–910. [Google Scholar] [CrossRef]
- Risk factors for neovascular age-related macular degeneration. The Eye Disease Case-Control Study Group. Arch. Ophthalmol. 1992, 110, 1701–1708.
- Curran-Celentano, J.; Hammond, B.R., Jr.; Ciulla, T.A.; Cooper, D.A.; Pratt, L.M.; Danis, R.B. Relation between dietary intake, serum concentrations, and retinal concentrations of lutein and zeaxanthin in adults in a Midwest population. Am. J. Clin. Nutr. 2001, 74, 796–802. [Google Scholar]
- Bernstein, P.S.; Zhao, D.Y.; Wintch, S.W.; Ermakov, I.V.; McClane, R.W.; Gellermann, W. Resonance Raman measurement of macular carotenoids in normal subjects and in age-related macular degeneration patients. Ophthalmology 2002, 109, 1780–1787. [Google Scholar] [CrossRef]
- Weigert, G.; Kaya, S.; Pemp, B.; Sacu, S.; Lasta, M.; Werkmeister, R.M.; Dragostinoff, N.; Simader, C.; Garhofer, G.; Schmidt-Erfurth, U.; et al. Effects of lutein supplementation on macular pigment optical density and visual acuity in patients with age-related macular degeneration. InvestIG. Ophthalmol. Vis. Sci. 2011, 52, 8174–8178. [Google Scholar] [CrossRef]
- Group, A.R.; Chew, E.Y.; Clemons, T.; Sangiovanni, J.P.; Danis, R.; Domalpally, A.; McBee, W.; Sperduto, R.; Ferris, F.L. The Age-Related Eye Disease Study 2 (AREDS2): Study design and baseline characteristics (AREDS2 report number 1). Ophthalmology 2012, 119, 2282–2289. [Google Scholar] [CrossRef]
- Age-Related Eye Disease Study Research Group; SanGiovanni, J.P.; Chew, E.Y.; Clemons, T.E.; Ferris, F.L., III; Gensler, G.; Lindblad, A.S.; Milton, R.C.; Seddon, J.M.; Sperduto, R.D. The relationship of dietary carotenoid and vitamin A, E, and C intake with age-related macular degeneration in a case-control study: AREDS Report No. 22. Arch. Ophthalmol. 2007, 125, 1225–1232. [Google Scholar] [CrossRef]
- Tang, L.; Zhang, Y.; Jiang, Y.; Willard, L.; Ortiz, E.; Wark, L.; Medeiros, D.; Lin, D. Dietary wolfberry ameliorates retinal structure abnormalities in db/db mice at the early stage of diabetes. Exp. Biol. Med. (Maywood) 2011, 236, 1051–1063. [Google Scholar]
- Hu, B.J.; Hu, Y.N.; Lin, S.; Ma, W.J.; Li, X.R. Application of Lutein and Zeaxanthin in nonproliferative diabetic retinopathy. Int. J. Ophthalmol. 2011, 4, 303–306. [Google Scholar]
- Woo, T.T.; Li, S.Y.; Lai, W.W.; Wong, D.; Lo, A.C. Neuroprotective effects of lutein in a rat model of retinal detachment. Graefes Arch. Clin. Exp. Ophthalmol. 2013, 251, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Chan, C.; Leung, I.; Lam, K.W.; Tso, M.O. The occurrence of retinol and carotenoids in human subretinal fluid. Curr. Eye Res. 1998, 17, 890–895. [Google Scholar] [CrossRef]
- Gao, S.; Qin, T.; Liu, Z.; Caceres, M.A.; Ronchi, C.F.; Chen, C.Y.; Yeum, K.J.; Taylor, A.; Blumberg, J.B.; Liu, Y.; et al. Lutein and zeaxanthin supplementation reduces H2O2-induced oxidative damage in human lens epithelial cells. Mol. Vis. 2011, 17, 3180–3190. [Google Scholar]
- Zhao, B.; He, Y.Y.; Chignell, C.F.; Yin, J.J.; Andley, U.; Roberts, J.E. Difference in phototoxicity of cyclodextrin complexed fullerene [(gamma-CyD)2/C60] and its aggregated derivatives toward human lens epithelial cells. Chem. Res. Toxicol. 2009, 22, 660–667. [Google Scholar] [CrossRef]
- Roberts, J.E.; Wielgus, A.R.; Boyes, W.K.; Andley, U.; Chignell, C.F. Phototoxicity and cytotoxicity of fullerol in human lens epithelial cells. Toxicol. Appl. Pharmacol. 2008, 228, 49–58. [Google Scholar] [CrossRef]
- Nolan, J.M.; O’Reilly, P.; Loughman, J.; Stack, J.; Loane, E.; Connolly, E.; Beatty, S. Augmentation of macular pigment following implantation of blue light-filtering intraocular lenses at the time of cataract surgery. Investig. Ophthalmol. Vis. Sci. 2009, 50, 4777–4785. [Google Scholar] [CrossRef]
- Barker, F.M., II. Dietary supplementation: Effects on visual performance and occurrence of AMD and cataracts. Curr. Med. Res. Opin. 2010, 26, 2011–2023. [Google Scholar] [CrossRef]
- Moeller, S.M.; Voland, R.; Tinker, L.; Blodi, B.A.; Klein, M.L.; Gehrs, K.M.; Johnson, E.J.; Snodderly, D.M.; Wallace, R.B.; Chappell, R.J.; et al. Associations between age-related nuclear cataract and lutein and zeaxanthin in the diet and serum in the Carotenoids in the Age-Related Eye Disease Study, an Ancillary Study of the Women’s Health Initiative. Arch. Ophthalmol. 2008, 126, 354–364. [Google Scholar] [CrossRef]
- Berendschot, T.T.; Broekmans, W.M.; Klopping-Ketelaars, I.A.; Kardinaal, A.F.; van Poppel, G.; van Norren, D. Lens aging in relation to nutritional determinants and possible risk factors for age-related cataract. Arch. Ophthalmol. 2002, 120, 1732–1737. [Google Scholar] [CrossRef]
- Jacques, P.F.; Chylack, L.T., Jr.; Hankinson, S.E.; Khu, P.M.; Rogers, G.; Friend, J.; Tung, W.; Wolfe, J.K.; Padhye, N.; Willett, W.C.; et al. Long-term nutrient intake and early age-related nuclear lens opacities. Arch. Ophthalmol. 2001, 119, 1009–1019. [Google Scholar]
- Suhler, E.B.; Lloyd, M.J.; Choi, D.; Rosenbaum, J.T.; Austin, D.F. Incidence and prevalence of uveitis in Veterans Affairs Medical Centers of the Pacific Northwest. Am. J. Ophthalmol. 2008, 146, 890–896. [Google Scholar] [CrossRef]
- Gritz, D.C.; Wong, I.G. Incidence and prevalence of uveitis in Northern California; the Northern California Epidemiology of Uveitis Study. Ophthalmology 2004, 111, 491–500; discussion 500. [Google Scholar]
- Yao, N.; Lan, F.; He, R.R.; Kurihara, H. Protective effects of bilberry (Vaccinium myrtillus L.) extract against endotoxin-induced uveitis in mice. J. Agric. Food Chem. 2010, 58, 4731–4736. [Google Scholar]
- He, R.R.; Tsoi, B.; Lan, F.; Yao, N.; Yao, X.S.; Kurihara, H. Antioxidant properties of lutein contribute to the protection against lipopolysaccharide-induced uveitis in mice. Chin. Med. 2011, 6, 38. [Google Scholar] [CrossRef]
- Jin, X.H.; Ohgami, K.; Shiratori, K.; Suzuki, Y.; Hirano, T.; Koyama, Y.; Yoshida, K.; Ilieva, I.; Iseki, K.; Ohno, S. Inhibitory effects of lutein on endotoxin-induced uveitis in Lewis rats. Investig. Ophthalmol. Vis. Sci. 2006, 47, 2562–2568. [Google Scholar] [CrossRef]
- WebMD L: Lutein. Edited by WebMd Editorial Team, 2013. Available online: http://www.webmd.com/vitamins-supplements/ingredientmono-754-lutein.aspx?activeIngredientId=754&activeIngredientName=lutein (accessed on 9 May 2013).
- Harikumar, K.B.; Nimita, C.V.; Preethi, K.C.; Kuttan, R.; Shankaranarayana, M.L.; Deshpande, J. Toxicity profile of lutein and lutein ester isolated from marigold flowers (Tagetes erecta). Int. J. Toxicol. 2008, 27, 1–9. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Koushan, K.; Rusovici, R.; Li, W.; Ferguson, L.R.; Chalam, K.V. The Role of Lutein in Eye-Related Disease. Nutrients 2013, 5, 1823-1839. https://doi.org/10.3390/nu5051823
Koushan K, Rusovici R, Li W, Ferguson LR, Chalam KV. The Role of Lutein in Eye-Related Disease. Nutrients. 2013; 5(5):1823-1839. https://doi.org/10.3390/nu5051823
Chicago/Turabian StyleKoushan, Keyvan, Raluca Rusovici, Wenhua Li, Lee R. Ferguson, and Kakarla V. Chalam. 2013. "The Role of Lutein in Eye-Related Disease" Nutrients 5, no. 5: 1823-1839. https://doi.org/10.3390/nu5051823