Selenium Supranutrition: Are the Potential Benefits of Chemoprevention Outweighed by the Promotion of Diabetes and Insulin Resistance?

Abstract
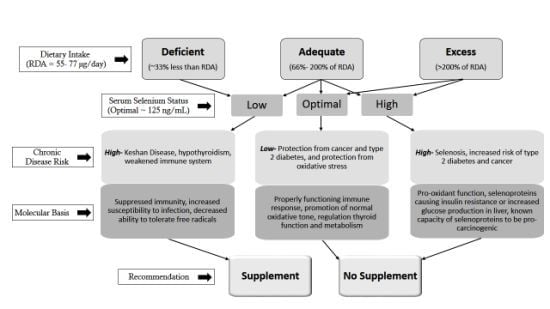
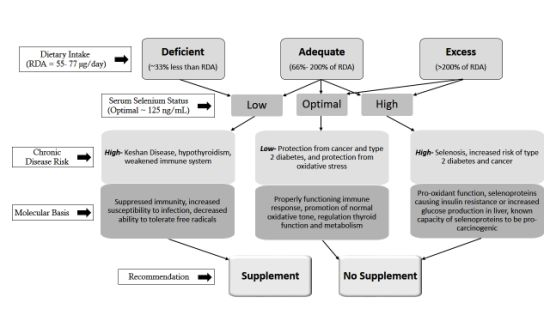
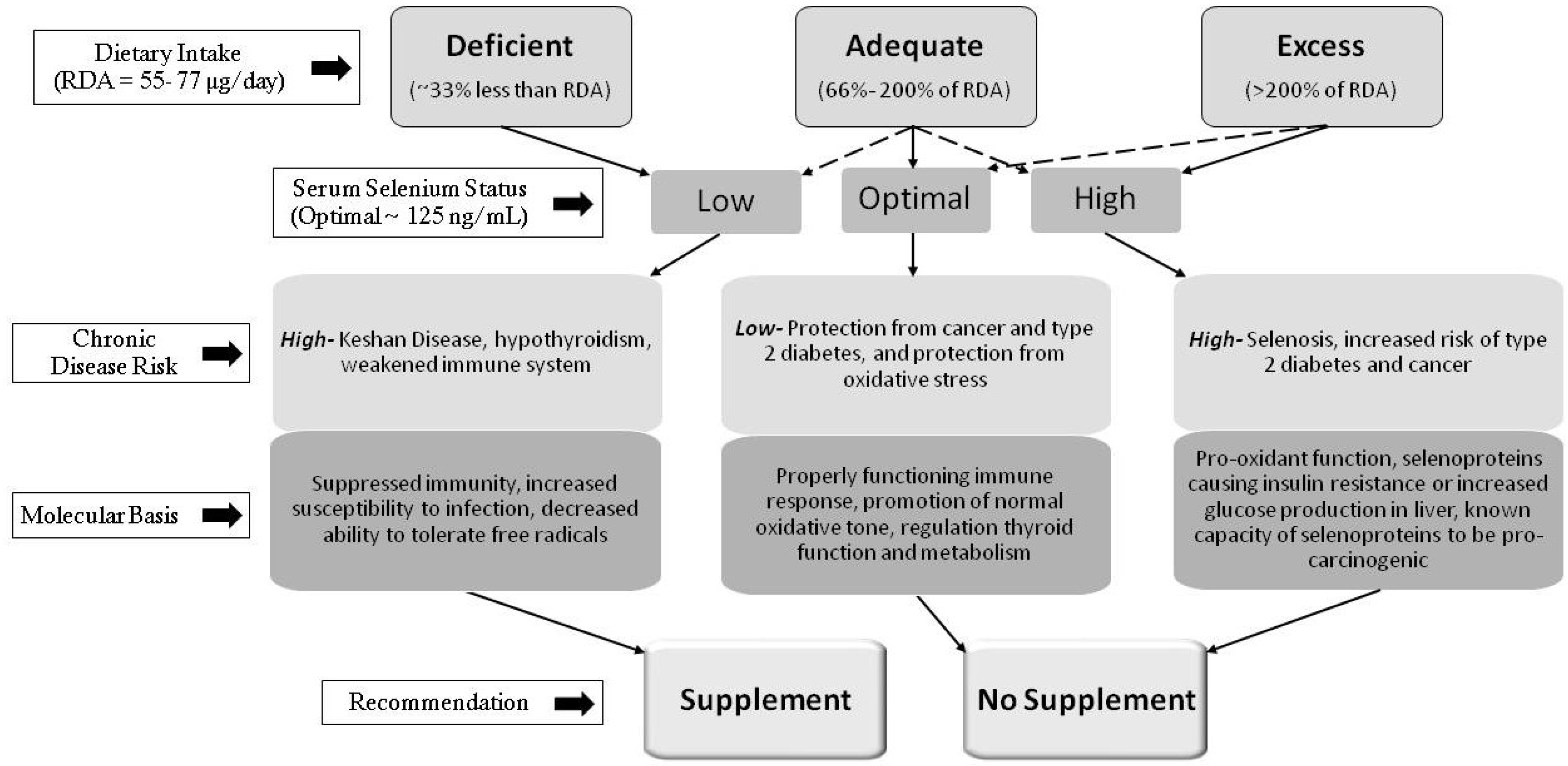
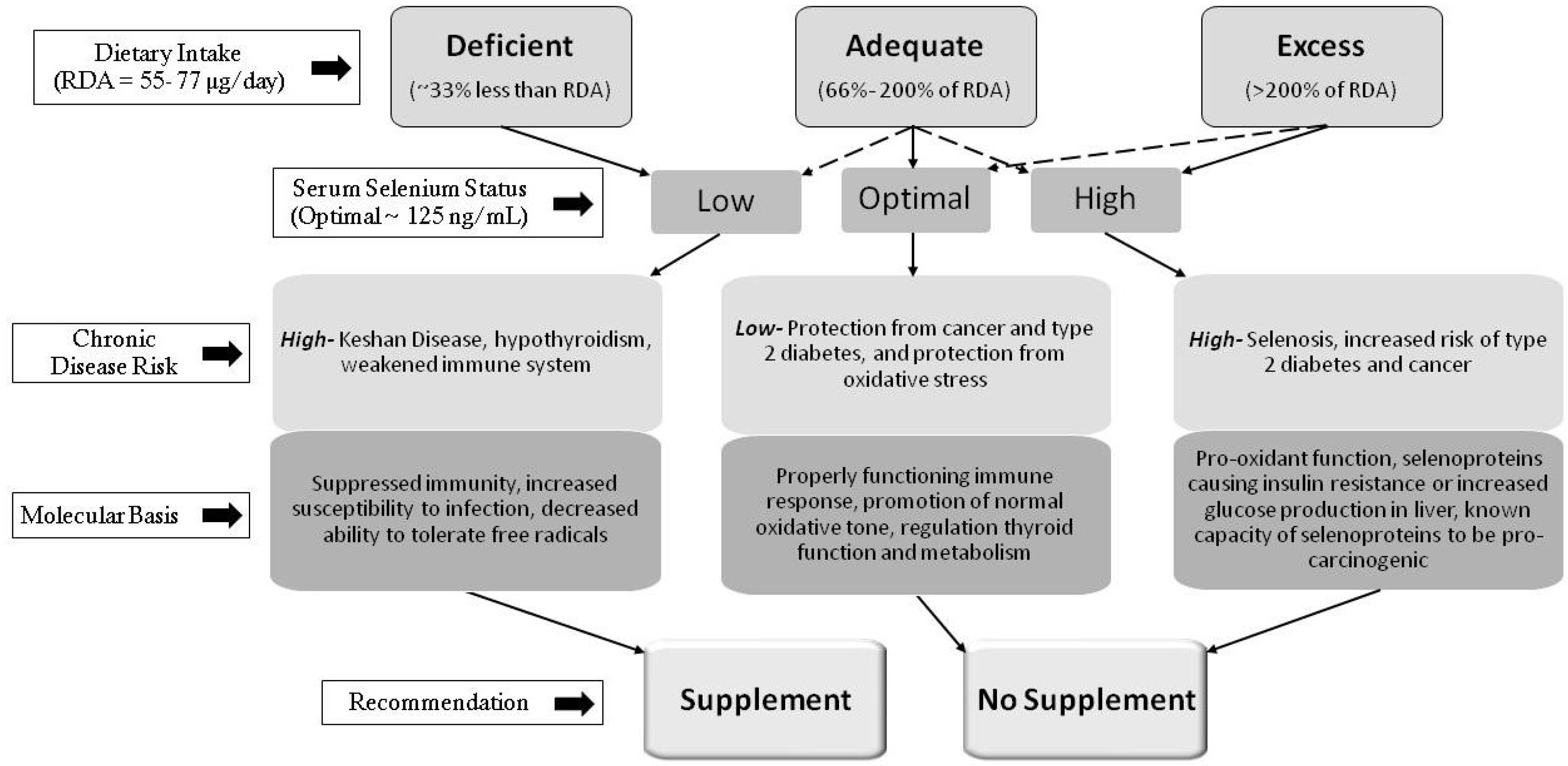
:
{kind=link}
{kind=link}
1. Introduction
2. Nutrition and Cancer
2.1. Selenium Supranutrition and Cancer Prevention
2.2. Type 2 Diabetes and Increased Cancer Risk
3. Type 2 Diabetes and Selenium
3.1. Linking Type 2 Diabetes and Selenium
3.2. Proposed Mechanisms
4. Conclusions and Perspectives
Conflict of Interest
References
- Burk, R.F.; Hill, K.E.; Motley, A.K. Plasma selenium in specific and non-specific forms. Biofactors 2001, 14, 107–114. [Google Scholar] [CrossRef]
- Xiong, Y.M.; Mo, X.Y.; Zou, X.Z.; Song, R.X.; Sun, W.Y.; Lu, W.; Chen, Q.; Yu, Y.X.; Zang, W.J. Association study between polymorphisms in selenoprotein genes and susceptibility to kashin-beck disease. Osteoarthritis Cartilage 2010, 18, 817–824. [Google Scholar] [CrossRef]
- Sun, W.; Wang, X.; Zou, X.; Song, R.; Du, X.; Hu, J.; Xiong, Y. Selenoprotein p gene r25191g/a polymorphism and quantification of selenoprotein P mRNA level in patients with Kashin-Beck disease. Br. J. Nutr. 2010, 104, 1283–1287. [Google Scholar] [CrossRef]
- Monsen, E.R. Dietary reference intakes for the antioxidant nutrients: Vitamin C, vitamin E, selenium, and carotenoids. J. Am. Diet. Assoc. 2000, 100, 637–640. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigo, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef]
- Ursini, F.; Bindoli, A. The role of selenium peroxidases in the protection against oxidative damage of membranes. Chem. Phys. Lipids 1987, 44, 255–276. [Google Scholar] [CrossRef]
- McKenzie, R.C.; Rafferty, T.S.; Beckett, G.J. Selenium: An essential element for immune function. Immunol. Today 1998, 19, 342–345. [Google Scholar] [CrossRef]
- Arthur, J.R.; Nicol, F.; Beckett, G.J. The role of selenium in thyroid hormone metabolism and effects of selenium deficiency on thyroid hormone and iodine metabolism. Biol. Trace Elem. Res. 1992, 34, 321–325. [Google Scholar] [CrossRef]
- Reeves, M.A.; Hoffmann, P.R. The human selenoproteome: Recent insights into functions and regulation. Cell. Mol. Life Sci. 2009, 66, 2457–2478. [Google Scholar] [CrossRef]
- Rayman, M.P. Food-chain selenium and human health: Emphasis on intake. Br. J. Nutr. 2008, 100, 254–268. [Google Scholar]
- Rayman, M.P. Selenium and human health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC)National Center for Health Statistics (NCHS)National Health and Nutrition Examination Survey Data; U.S. Department of Health and Human Services, Centers for Disease Control and Prevention: Hyattsville, MD, USA, 2012. Available online: http://www.ars.usda.gov/SP2UserFiles/Place/12355000/pdf/0910/Table_37_SUP_GEN_09.pdf (accessed on 28 February 2013).
- Laclaustra, M.; Navas-Acien, A.; Stranges, S.; Ordovas, J.M.; Guallar, E. Serum selenium concentrations and diabetes in U.S. Adults: National health and nutrition examination survey (NHANES) 2003–2004. Environ. Health Perspect. 2009, 117, 1409–1413. [Google Scholar]
- Stranges, S.; Sieri, S.; Vinceti, M.; Grioni, S.; Guallar, E.; Laclaustra, M.; Muti, P.; Berrino, F.; Krogh, V. A prospective study of dietary selenium intake and risk of type 2 diabetes. BMC Public Health 2010, 10, 564. [Google Scholar] [CrossRef]
- Penney, K.L.; Schumacher, F.R.; Li, H.; Kraft, P.; Morris, J.S.; Kurth, T.; Mucci, L.A.; Hunter, D.J.; Kantoff, P.W.; Stampfer, M.J.; et al. A large prospective study of sep15 genetic variation, interaction with plasma selenium levels, and prostate cancer risk and survival. Cancer Prev. Res. 2010, 3, 604–610. [Google Scholar] [CrossRef]
- Curran, J.E.; Jowett, J.B.; Elliott, K.S.; Gao, Y.; Gluschenko, K.; Wang, J.; Abel Azim, D.M.; Cai, G.; Mahaney, M.C.; Comuzzie, A.G.; et al. Genetic variation in selenoprotein S influences inflammatory response. Nat. Genet. 2005, 37, 1234–1241. [Google Scholar] [CrossRef]
- Peters, U.; Chatterjee, N.; Hayes, R.B.; Schoen, R.E.; Wang, Y.; Chanock, S.J.; Foster, C.B. Variation in the selenoenzyme genes and risk of advanced distal colorectal adenoma. Cancer Epidemiol. Biomarkers Prev. 2008, 17, 1144–1154. [Google Scholar] [CrossRef]
- Duffield-Lillico, A.J.; Reid, M.E.; Turnbull, B.W.; Combs, G.F., Jr.; Slate, E.H.; Fischbach, L.A.; Marshall, J.R.; Clark, L.C. Baseline characteristics and the effect of selenium supplementation on cancer incidence in a randomized clinical trial: A summary report of the nutritional prevention of cancer trial. Cancer Epidemiol. Biomarkers Prev. 2002, 11, 630–639. [Google Scholar]
- Swanson, C.A.; Longnecker, M.P.; Veillon, C.; Howe, M.; Levander, O.A.; Taylor, P.R.; McAdam, P.A.; Brown, C.C.; Stampfer, M.J.; Willett, W.C. Selenium intake, age, gender, and smoking in relation to indices of selenium status of adults residing in a seleniferous area. Am. J. Clin. Nutr. 1990, 52, 858–862. [Google Scholar]
- Johansson, U.; Johnsson, F.; Joelsson, B.; Berglund, M.; Akesson, B. Selenium status in patients with liver cirrhosis and alcoholism. Br. J. Nutr. 1986, 55, 227–233. [Google Scholar] [CrossRef]
- De Jong, N.; Gibson, R.S.; Thomson, C.D.; Ferguson, E.L.; McKenzie, J.E.; Green, T.J.; Horwath, C.C. Selenium and zinc status are suboptimal in a sample of older New Zealand women in a community-based study. J. Nutr. 2001, 131, 2677–2684. [Google Scholar]
- Allard, J.P.; Aghdassi, E.; Chau, J.; Salit, I.; Walmsley, S. Oxidative stress and plasma antioxidant micronutrients in humans with HIV infection. Am. J. Clin. Nutr. 1998, 67, 143–147. [Google Scholar]
- Dworkin, B.M.; Rosenthal, W.S.; Wormser, G.P.; Weiss, L.; Nunez, M.; Joline, C.; Herp, A. Abnormalities of blood selenium and glutathione peroxidase activity in patients with acquired immunodeficiency syndrome and aids-related complex. Biol. Trace Elem. Res. 1988, 15, 167–177. [Google Scholar] [CrossRef]
- Ashton, K.; Hooper, L.; Harvey, L.J.; Hurst, R.; Casgrain, A.; Fairweather-Tait, S.J. Methods of assessment of selenium status in humans: A systematic review. Am. J. Clin. Nutr. 2009, 89, 2025S–2039S. [Google Scholar] [CrossRef]
- Hurst, R.; Armah, C.N.; Dainty, J.R.; Hart, D.J.; Teucher, B.; Goldson, A.J.; Broadley, M.R.; Motley, A.K.; Fairweather-Tait, S.J. Establishing optimal selenium status: Results of a randomized, double-blind, placebo-controlled trial. Am. J. Clin. Nutr. 2010, 91, 923–931. [Google Scholar] [CrossRef]
- Xia, Y.; Hill, K.E.; Byrne, D.W.; Xu, J.; Burk, R.F. Effectiveness of selenium supplements in a low-selenium area of china. Am. J. Clin. Nutr. 2005, 81, 829–834. [Google Scholar]
- Zeng, H.; Combs, G.F., Jr. Selenium as an anticancer nutrient: Roles in cell proliferation and tumor cell invasion. J. Nutr. Biochem. 2008, 19, 1–7. [Google Scholar] [CrossRef]
- Clark, L.C.; Combs, G.F., Jr.; Turnbull, B.W.; Slate, E.H.; Chalker, D.K.; Chow, J.; Davis, L.S.; Glover, R.A.; Graham, G.F.; Gross, E.G.; et al. Effects of selenium supplementation for cancer prevention in patients with carcinoma of the skin. A randomized controlled trial. Nutritional prevention of cancer study group. JAMA 1996, 276, 1957–1963. [Google Scholar] [CrossRef]
- Rocourt, C.R.; Wu, M.; Chen, B.P.; Cheng, W.H. The catalytic subunit of DNA-dependent protein kinase is downstream of atm and feeds forward oxidative stress in the selenium-induced senescence response. J. Nutr. Biochem. 2012. [Google Scholar] [CrossRef]
- Wu, M.; Wu, R.T.; Wang, T.T.; Cheng, W.H. Role for p53 in selenium-induced senescence. J. Agric. Food Chem. 2011, 59, 11882–11887. [Google Scholar]
- Wu, M.; Kang, M.M.; Schoene, N.W.; Cheng, W.H. Selenium compounds activate early barriers of tumorigenesis. J. Biol. Chem. 2010, 285, 12055–12062. [Google Scholar] [CrossRef]
- Zu, K.; Bihani, T.; Lin, A.; Park, Y.M.; Mori, K.; Ip, C. Enhanced selenium effect on growth arrest by BiP/GRP78 knockdown in p53-null human prostate cancer cells. Oncogene 2006, 25, 546–554. [Google Scholar]
- Xu, J.X.; Wang, J.; Wang, T. Effects of vitamin E and selenium on the metabolism of free radicals in broilers. Ying Yong Sheng Tai Xue Bao 2007, 18, 1789–1793. [Google Scholar]
- Soto-Reyes, E.; Del Razo, L.M.; Valverde, M.; Rojas, E. Role of the alkali labile sites, reactive oxygen species and antioxidants in DNA damage induced by methylated trivalent metabolites of inorganic arsenic. Biometals 2005, 18, 493–506. [Google Scholar] [CrossRef]
- Lanfear, J.; Fleming, J.; Wu, L.; Webster, G.; Harrison, P.R. The selenium metabolite selenodiglutathione induces p53 and apoptosis: Relevance to the chemopreventive effects of selenium? Carcinogenesis 1994, 15, 1387–1392. [Google Scholar] [CrossRef]
- Ip, C.; Thompson, H.J.; Zhu, Z.; Ganther, H.E. In vitro and in vivo studies of methylseleninic acid: Evidence that a monomethylated selenium metabolite is critical for cancer chemoprevention. Cancer Res. 2000, 60, 2882–2886. [Google Scholar]
- Last, K.; Maharaj, L.; Perry, J.; Strauss, S.; Fitzgibbon, J.; Lister, T.A.; Joel, S. The activity of methylated and non-methylated selenium species in lymphoma cell lines and primary tumours. Ann. Oncol. 2006, 17, 773–779. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Speckmann, B.; Sies, H. Toward understanding success and failures in the use of selenium for cancer prevention. Antioxid. Redox Signal. 2013. [Google Scholar] [CrossRef]
- Trumbo, P.R. The level of evidence for permitting a qualified health claim: FDA’s review of the evidence for selenium and cancer and vitamin E and heart disease. J. Nutr. 2005, 135, 354–356. [Google Scholar]
- Lippman, S.M.; Klein, E.A.; Goodman, P.J.; Lucia, M.S.; Thompson, I.M.; Ford, L.G.; Parnes, H.L.; Minasian, L.M.; Gaziano, J.M.; Hartline, J.A.; et al. Effect of selenium and vitamin E on risk of prostate cancer and other cancers: The selenium and vitamin E cancer prevention trial (SELECT). JAMA 2009, 301, 39–51. [Google Scholar] [CrossRef]
- Wu, J.; Salisbury, C.; Graham, R.; Lyons, G.; Fenech, M. Increased consumption of wheat biofortified with selenium does not modify biomarkers of cancer risk, oxidative stress, or immune function in healthy australian males. Environ. Mol. Mutagen. 2009, 50, 489–501. [Google Scholar] [CrossRef]
- Hatfield, D.L.; Yoo, M.H.; Carlson, B.A.; Gladyshev, V.N. Selenoproteins that function in cancer prevention and promotion. Biochim. Biophys. Acta 2009, 1790, 1541–1545. [Google Scholar] [CrossRef]
- Yoo, M.H.; Xu, X.M.; Carlson, B.A.; Patterson, A.D.; Gladyshev, V.N.; Hatfield, D.L. Targeting thioredoxin reductase 1 reduction in cancer cells inhibits self-sufficient growth and DNA replication. PLoS One 2007, 2, e1112. [Google Scholar] [CrossRef]
- Boosalis, M.G. The role of selenium in chronic disease. Nutr. Clin. Pract. 2008, 23, 152–160. [Google Scholar] [CrossRef]
- Bleys, J.; Navas-Acien, A.; Guallar, E. Selenium and diabetes: More bad news for supplements. Ann. Intern. Med. 2007, 147, 271–272. [Google Scholar] [CrossRef]
- Mueller, A.S.; Mueller, K.; Wolf, N.M.; Pallauf, J. Selenium and diabetes: An enigma? Free Radic. Res. 2009, 43, 1029–1059. [Google Scholar] [CrossRef]
- Marcason, W. What is the latest research on the connection between selenium and diabetes? J. Am. Diet. Assoc. 2008, 108, 188. [Google Scholar] [CrossRef]
- Willett, W.C.; Polk, B.F.; Morris, J.S.; Stampfer, M.J.; Pressel, S.; Rosner, B.; Taylor, J.O.; Schneider, K.; Hames, C.G. Prediagnostic serum selenium and risk of cancer. Lancet 1983, 2, 130–134. [Google Scholar]
- Fairweather-Tait, S.J.; Bao, Y.; Broadley, M.R.; Collings, R.; Ford, D.; Hesketh, J.E.; Hurst, R. Selenium in human health and disease. Antioxid. Redox Signal. 2011, 14, 1337–1383. [Google Scholar] [CrossRef]
- Klein, E.A.; Thompson, I.M., Jr.; Tangen, C.M.; Crowley, J.J.; Lucia, M.S.; Goodman, P.J.; Minasian, L.M.; Ford, L.G.; Parnes, H.L.; Gaziano, J.M.; et al. Vitamin E and the risk of prostate cancer: The selenium and vitamin E cancer prevention trial (SELECT). JAMA 2011, 306, 1549–1556. [Google Scholar] [CrossRef]
- Li, G.X.; Lee, H.J.; Wang, Z.; Hu, H.; Liao, J.D.; Watts, J.C.; Combs, G.F., Jr.; Lu, J. Superior in vivo inhibitory efficacy of methylseleninic acid against human prostate cancer over selenomethionine or selenite. Carcinogenesis 2008, 29, 1005–1012. [Google Scholar] [CrossRef]
- Wang, C.; Wang, H.; Luo, J.; Hu, Y.; Wei, L.; Duan, M.; He, H. Selenium deficiency impairs host innate immune response and induces susceptibility to listeria monocytogenes infection. BMC Immunol. 2009, 10, 55. [Google Scholar] [CrossRef]
- Hu, F.B. Globalization of diabetes: The role of diet, lifestyle, and genes. Diabetes Care 2011, 34, 1249–1257. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- Vigneri, P.; Frasca, F.; Sciacca, L.; Pandini, G.; Vigneri, R. Diabetes and cancer. Endocr. Relat. Cancer 2009, 16, 1103–1123. [Google Scholar] [CrossRef]
- Stumvoll, M.; Goldstein, B.J.; van Haeften, T.W. Type 2 diabetes: Principles of pathogenesis and therapy. Lancet 2005, 365, 1333–1346. [Google Scholar] [CrossRef]
- Kaaks, R. Nutrition, hormones, and breast cancer: Is insulin the missing link? Cancer Causes Control 1996, 7, 605–625. [Google Scholar] [CrossRef]
- Lindblad, P.; Chow, W.H.; Chan, J.; Bergstrom, A.; Wolk, A.; Gridley, G.; McLaughlin, J.K.; Nyren, O.; Adami, H.O. The role of diabetes mellitus in the aetiology of renal cell cancer. Diabetologia 1999, 42, 107–112. [Google Scholar] [CrossRef]
- Larsson, S.C.; Orsini, N.; Brismar, K.; Wolk, A. Diabetes mellitus and risk of bladder cancer: A meta-analysis. Diabetologia 2006, 49, 2819–2823. [Google Scholar] [CrossRef]
- Limburg, P.J.; Vierkant, R.A.; Fredericksen, Z.S.; Leibson, C.L.; Rizza, R.A.; Gupta, A.K.; Ahlquist, D.A.; Melton, L.J., III; Sellers, T.A.; Cerhan, J.R. Clinically confirmed type 2 diabetes mellitus and colorectal cancer risk: A population-based, retrospective cohort study. Am. J. Gastroenterol. 2006, 101, 1872–1879. [Google Scholar] [CrossRef]
- Larsson, S.C.; Orsini, N.; Wolk, A. Diabetes mellitus and risk of colorectal cancer: A meta-analysis. J. Natl. Cancer Inst. 2005, 97, 1679–1687. [Google Scholar] [CrossRef]
- Stadler, J.; Yeung, K.S.; Furrer, R.; Marcon, N.; Himal, H.S.; Bruce, W.R. Proliferative activity of rectal mucosa and soluble fecal bile acids in patients with normal colons and in patients with colonic polyps or cancer. Cancer Lett. 1988, 38, 315–320. [Google Scholar] [CrossRef]
- Will, J.C.; Galuska, D.A.; Vinicor, F.; Calle, E.E. Colorectal cancer: Another complication of diabetes mellitus? Am. J. Epidemiol. 1998, 147, 816–825. [Google Scholar] [CrossRef]
- Mitri, J.; Castillo, J.; Pittas, A.G. Diabetes and risk of non-Hodgkin’s lymphoma: A meta-analysis of observational studies. Diabetes Care 2008, 31, 2391–2397. [Google Scholar] [CrossRef]
- Kasper, J.S.; Giovannucci, E. A meta-analysis of diabetes mellitus and the risk of prostate cancer. Cancer Epidemiol. Biomarkers. Prev. 2006, 15, 2056–2062. [Google Scholar] [CrossRef]
- Barrett-Connor, E. Lower endogenous androgen levels and dyslipidemia in men with non-insulin-dependent diabetes mellitus. Ann. Intern. Med. 1992, 117, 807–811. [Google Scholar] [CrossRef]
- Betancourt-Albrecht, M.; Cunningham, G.R. Hypogonadism and diabetes. Int. J. Impot. Res. 2003, 15, S14–S20. [Google Scholar] [CrossRef]
- Shamberger, R.J.; Rukovena, E.; Longfield, A.K.; Tytko, S.A.; Deodhar, S.; Willis, C.E. Antioxidants and cancer. I. Selenium in the blood of normals and cancer patients. J. Natl. Cancer Inst. 1973, 50, 863–870. [Google Scholar]
- Bleys, J.; Navas-Acien, A.; Guallar, E. Serum selenium and diabetes in U.S. Adults. Diabetes Care 2007, 30, 829–834. [Google Scholar] [CrossRef]
- Czernichow, S.; Couthouis, A.; Bertrais, S.; Vergnaud, A.C.; Dauchet, L.; Galan, P.; Hercberg, S. Antioxidant supplementation does not affect fasting plasma glucose in the supplementation with antioxidant vitamins and minerals (SU.VI.MAX) study in France: Association with dietary intake and plasma concentrations. Am. J. Clin. Nutr. 2006, 84, 395–399. [Google Scholar]
- Stranges, S.; Marshall, J.R.; Natarajan, R.; Donahue, R.P.; Trevisan, M.; Combs, G.F.; Cappuccio, F.P.; Ceriello, A.; Reid, M.E. Effects of long-term selenium supplementation on the incidence of type 2 diabetes: A randomized trial. Ann. Intern. Med. 2007, 147, 217–223. [Google Scholar] [CrossRef]
- Kornhauser, C.; Garcia-Ramirez, J.R.; Wrobel, K.; Perez-Luque, E.L.; Garay-Sevilla, M.E.; Wrobel, K. Serum selenium and glutathione peroxidase concentrations in type 2 diabetes mellitus patients. Prim. Care Diabetes 2008, 2, 81–85. [Google Scholar] [CrossRef]
- Ruiz, C.; Alegria, A.; Barbera, R.; Farre, R.; Lagarda, J. Selenium, zinc and copper in plasma of patients with type 1 diabetes mellitus in different metabolic control states. J. Trace Elem. Med. Biol. 1998, 12, 91–95. [Google Scholar] [CrossRef]
- Navarro-Alarcon, M.; de la Serrana Lopez, G.H.; Perez-Valero, V.; Lopez-Martinez, C. Serum and urine selenium concentrations as indicators of body status in patients with diabetes mellitus. Sci. Total Environ. 1999, 228, 79–85. [Google Scholar] [CrossRef]
- Quilliot, D.; Dousset, B.; Guerci, B.; Dubois, F.; Drouin, P.; Ziegler, O. Evidence that diabetes mellitus favors impaired metabolism of zinc, copper, and selenium in chronic pancreatitis. Pancreas 2001, 22, 299–306. [Google Scholar] [CrossRef]
- Rajpathak, S.; Rimm, E.; Morris, J.S.; Hu, F. Toenail selenium and cardiovascular disease in men with diabetes. J. Am. Coll. Nutr. 2005, 24, 250–256. [Google Scholar]
- Park, K.; Rimm, E.B.; Siscovick, D.S.; Spiegelman, D.; Manson, J.E.; Morris, J.S.; Hu, F.B.; Mozaffarian, D. Toenail selenium and incidence of type 2 diabetes in U.S. Men and women. Diabetes Care 2012, 35, 1544–1551. [Google Scholar] [CrossRef]
- Al-Saleh, E.; Nandakumaran, M.; Al-Shammari, M.; Al-Harouny, A. Maternal-fetal status of copper, iron, molybdenum, selenium and zinc in patients with gestational diabetes. J. Matern. Fetal Neonatal Med. 2004, 16, 15–21. [Google Scholar] [CrossRef]
- Kilinc, M.; Guven, M.A.; Ezer, M.; Ertas, I.E.; Coskun, A. Evaluation of serum selenium levels in turkish women with gestational diabetes mellitus, glucose intolerants, and normal controls. Biol. Trace Elem. Res. 2008, 123, 35–40. [Google Scholar] [CrossRef]
- Rayman, M.P.; Blundell-Pound, G.; Pastor-Barriuso, R.; Guallar, E.; Steinbrenner, H.; Stranges, S. A randomized trial of selenium supplementation and risk of type-2 diabetes, as assessed by plasma adiponectin. PLoS One 2012, 7, e45269. [Google Scholar]
- Labunskyy, V.M.; Lee, B.C.; Handy, D.E.; Loscalzo, J.; Hatfield, D.L.; Gladyshev, V.N. Both maximal expression of selenoproteins and selenoprotein deficiency can promote development of type 2 diabetes-like phenotype in mice. Antioxid. Redox Signal. 2011, 14, 2327–2336. [Google Scholar] [CrossRef]
- Lebedeva, E.A. Relationship between serum selenium level and lipid spectrum in patients with diabetes mellitus type I. Probl. Endokrinol. (Mosk.) 1991, 37, 23–25. [Google Scholar]
- Steinbrenner, H.; Speckmann, B.; Pinto, A.; Sies, H. High selenium intake and increased diabetes risk: Experimental evidence for interplay between selenium and carbohydrate metabolism. J. Clin. Biochem. Nutr. 2011, 48, 40–45. [Google Scholar] [CrossRef]
- Yang, S.J.; Hwang, S.Y.; Choi, H.Y.; Yoo, H.J.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Baik, S.H.; Choi, D.S.; Choi, K.M. Serum selenoprotein P levels in patients with type 2 diabetes and prediabetes: Implications for insulin resistance, inflammation, and atherosclerosis. J. Clin. Endocrinol. Metab. 2011, 96, E1325–E1329. [Google Scholar] [CrossRef]
- Gao, Y.; Feng, H.C.; Walder, K.; Bolton, K.; Sunderland, T.; Bishara, N.; Quick, M.; Kantham, L.; Collier, G.R. Regulation of the selenoprotein SelS by glucose deprivation and endoplasmic reticulum stress - SelS is a novel glucose-regulated protein. FEBS Lett. 2004, 563, 185–190. [Google Scholar] [CrossRef]
- Misu, H.; Takamura, T.; Takayama, H.; Hayashi, H.; Matsuzawa-Nagata, N.; Kurita, S.; Ishikura, K.; Ando, H.; Takeshita, Y.; Ota, T.; et al. A liver-derived secretory protein, selenoprotein P, causes insulin resistance. Cell Metab. 2010, 12, 483–495. [Google Scholar] [CrossRef]
- Douillet, C.; Tabib, A.; Bost, M.; Accominotti, M.; Borson-Chazot, F.; Ciavatti, M. A selenium supplement associated or not with vitamin E delays early renal lesions in experimental diabetes in rats. Proc. Soc. Exp. Biol. Med. 1996, 211, 323–331. [Google Scholar]
- Ghaffari, T.; Nouri, M.; Saei, A.A.; Rashidi, M.R. Aldehyde and xanthine oxidase activities in tissues of streptozotocin-induced diabetic rats: Effects of vitamin E and selenium supplementation. Biol. Trace Elem. Res. 2012, 147, 217–225. [Google Scholar] [CrossRef]
- Hwang, D.; Seo, S.; Kim, Y.; Kim, C.; Shim, S.; Jee, S.; Lee, S.; Jang, M.; Kim, M.; Yim, S.; et al. Selenium acts as an insulin-like molecule for the down-regulation of diabetic symptoms via endoplasmic reticulum stress and insulin signalling proteins in diabetes-induced non-obese diabetic mice. J. Biosci. 2007, 32, 723–735. [Google Scholar] [CrossRef]
- Iizuka, Y.; Ueda, Y.; Yagi, Y.; Sakurai, E. Significant improvement of insulin resistance of gk rats by treatment with sodium selenate. Biol. Trace Elem. Res. 2010, 138, 265–271. [Google Scholar] [CrossRef]
- Kljai, K.; Runje, R. Selenium and glycogen levels in diabetic patients. Biol. Trace Elem. Res. 2001, 83, 223–229. [Google Scholar] [CrossRef]
- Stapleton, S.R. Selenium: An insulin-mimetic. Cell. Mol. Life Sci. 2000, 57, 1874–1879. [Google Scholar] [CrossRef]
- Maritim, A.C.; Sanders, R.A.; Watkins, J.B., III. Diabetes, oxidative stress, and antioxidants: A review. J. Biochem. Mol. Toxicol. 2003, 17, 24–38. [Google Scholar]
- Baynes, J.W. Role of oxidative stress in development of complications in diabetes. Diabetes 1991, 40, 405–412. [Google Scholar]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Oxidative stress and stress-activated signaling pathways: A unifying hypothesis of type 2 diabetes. Endocr. Rev. 2002, 23, 599–622. [Google Scholar] [CrossRef]
- Monnier, L.; Mas, E.; Ginet, C.; Michel, F.; Villon, L.; Cristol, J.P.; Colette, C. Activation of oxidative stress by acute glucose fluctuations compared with sustained chronic hyperglycemia in patients with type 2 diabetes. JAMA 2006, 295, 1681–1687. [Google Scholar] [CrossRef]
- Steinbrenner, H.; Sies, H. Protection against reactive oxygen species by selenoproteins. Biochem. Biophys. Acta 2009, 1790, 1478–1485. [Google Scholar] [CrossRef]
- Naziroglu, M.; Cay, M. Protective role of intraperitoneally administered vitamin E and selenium on the antioxidative defense mechanisms in rats with diabetes induced by streptozotocin. Biol. Trace Elem. Res. 2001, 79, 149–159. [Google Scholar] [CrossRef]
- Guney, M.; Erdemoglu, E.; Mungan, T. Selenium-vitamin E combination and melatonin modulates diabetes-induced blood oxidative damage and fetal outcomes in pregnant rats. Biol. Trace Elem. Res. 2011, 143, 1091–1102. [Google Scholar] [CrossRef]
- Agbor, G.A.; Vinson, J.A.; Patel, S.; Patel, K.; Scarpati, J.; Shiner, D.; Wardrop, F.; Tompkins, T.A. Effect of selenium- and glutathione-enriched yeast supplementation on a combined atherosclerosis and diabetes hamster model. J. Agric. Food Chem. 2007, 55, 8731–8736. [Google Scholar] [CrossRef]
- Can, B.; Ulusu, N.N.; Kilinc, K.; Leyla Acan, N.; Saran, Y.; Turan, B. Selenium treatment protects diabetes-induced biochemical and ultrastructural alterations in liver tissue. Biol. Trace Elem. Res. 2005, 105, 135–150. [Google Scholar] [CrossRef]
- Gebre-Medhin, M.; Ewald, U.; Tuvemo, T. Serum selenium is related to low-density lipoproteins in healthy children but not in children with diabetes. Ups. J. Med. Sci. 1988, 93, 57–62. [Google Scholar]
- Yadav, S.; Day, J.P.; Mohan, V.; Snehalatha, C.; Braganza, J.M. Selenium and diabetes in the tropics. Pancreas 1991, 6, 528–533. [Google Scholar] [CrossRef]
- Douillet, C.; Bost, M.; Accominotti, M.; Borson-Chazot, F.; Ciavatti, M. Effect of selenium and vitamin E supplements on tissue lipids, peroxides, and fatty acid distribution in experimental diabetes. Lipids 1998, 33, 393–399. [Google Scholar] [CrossRef]
- Kim, S.S.; Koo, J.H.; Kwon, I.S.; Oh, Y.S.; Lee, S.J.; Kim, E.J.; Kim, W.K.; Lee, J.; Cho, J.Y. Exercise training and selenium or a combined treatment ameliorates aberrant expression of glucose and lactate metabolic proteins in skeletal muscle in a rodent model of diabetes. Nutr. Res. Pract. 2011, 5, 205–213. [Google Scholar] [CrossRef]
- Ayaz, M.; Can, B.; Ozdemir, S.; Turan, B. Protective effect of selenium treatment on diabetes-induced myocardial structural alterations. Biol. Trace Elem. Res. 2002, 89, 215–226. [Google Scholar] [CrossRef]
- Sheng, X.Q.; Huang, K.X.; Xu, H.B. New experimental observation on the relationship of selenium and diabetes mellitus. Biol. Trace Elem. Res. 2004, 99, 241–253. [Google Scholar] [CrossRef]
- Skripchenko, N.D.; Sharafetdinov, K.; Plotnikova, O.A.; Meshcheriakova, V.A.; Mal’tsev, G. Effect of selenium enriched diet on lipid peroxidation in patients with diabetes mellitus type 2. Vopr. Pitan. 2003, 72, 14–17. [Google Scholar]
- McClung, J.P.; Roneker, C.A.; Mu, W.; Lisk, D.J.; Langlais, P.; Liu, F.; Lei, X.G. Development of insulin resistance and obesity in mice overexpressing cellular glutathione peroxidase. Proc. Natl. Acad. Sci. USA 2004, 101, 8852–8857. [Google Scholar]
- Loh, K.; Deng, H.; Fukushima, A.; Cai, X.; Boivin, B.; Galic, S.; Bruce, C.; Shields, B.J.; Skiba, B.; Ooms, L.M.; et al. Reactive oxygen species enhance insulin sensitivity. Cell Metab. 2009, 10, 260–272. [Google Scholar] [CrossRef]
- Bindoli, A.; Rigobello, M.P. Principles in redox signaling: From chemistry to functional significance. Antioxid. Redox Signal. 2013, 18, 1557–1593. [Google Scholar] [CrossRef]
- Samoylenko, A.; Hossain, J.A.; Mennerich, D.; Kellokumpu, S.; Hiltunen, K.; Kietzmann, T. Nutritional countermeasures targeting reactive oxygen species in cancer: From mechanisms to biomarkers and clinical evidence. Antioxid. Redox Signal. 2013. [Google Scholar] [CrossRef]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar]
- Rhee, S.G. Redox signaling: Hydrogen peroxide as intracellular messenger. Exp. Mol. Med. 1999, 31, 53–59. [Google Scholar] [CrossRef]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef]
- Stone, J.R.; Yang, S. Hydrogen peroxide: A signaling messenger. Antioxid. Redox Signal. 2006, 8, 243–270. [Google Scholar] [CrossRef]
- Veal, E.A.; Day, A.M.; Morgan, B.A. Hydrogen peroxide sensing and signaling. Mol. Cell 2007, 26, 1–14. [Google Scholar] [CrossRef]
- Roman, M.; Lapolla, A.; Jitaru, P.; Sechi, A.; Cosma, C.; Cozzi, G.; Cescon, P.; Barbante, C. Plasma selenoproteins concentrations in type 2 diabetes mellitus—A pilot study. Transl. Res. 2010, 156, 242–250. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rocourt, C.R.B.; Cheng, W.-H. Selenium Supranutrition: Are the Potential Benefits of Chemoprevention Outweighed by the Promotion of Diabetes and Insulin Resistance? Nutrients 2013, 5, 1349-1365. https://doi.org/10.3390/nu5041349
Rocourt CRB, Cheng W-H. Selenium Supranutrition: Are the Potential Benefits of Chemoprevention Outweighed by the Promotion of Diabetes and Insulin Resistance? Nutrients. 2013; 5(4):1349-1365. https://doi.org/10.3390/nu5041349
Chicago/Turabian StyleRocourt, Caroline R. B., and Wen-Hsing Cheng. 2013. "Selenium Supranutrition: Are the Potential Benefits of Chemoprevention Outweighed by the Promotion of Diabetes and Insulin Resistance?" Nutrients 5, no. 4: 1349-1365. https://doi.org/10.3390/nu5041349
APA StyleRocourt, C. R. B., & Cheng, W.-H. (2013). Selenium Supranutrition: Are the Potential Benefits of Chemoprevention Outweighed by the Promotion of Diabetes and Insulin Resistance? Nutrients, 5(4), 1349-1365. https://doi.org/10.3390/nu5041349