Vampire Bat Rabies: Ecology, Epidemiology and Control

Abstract
:1. Introduction
2. Vampire Bat Biology and Ecology
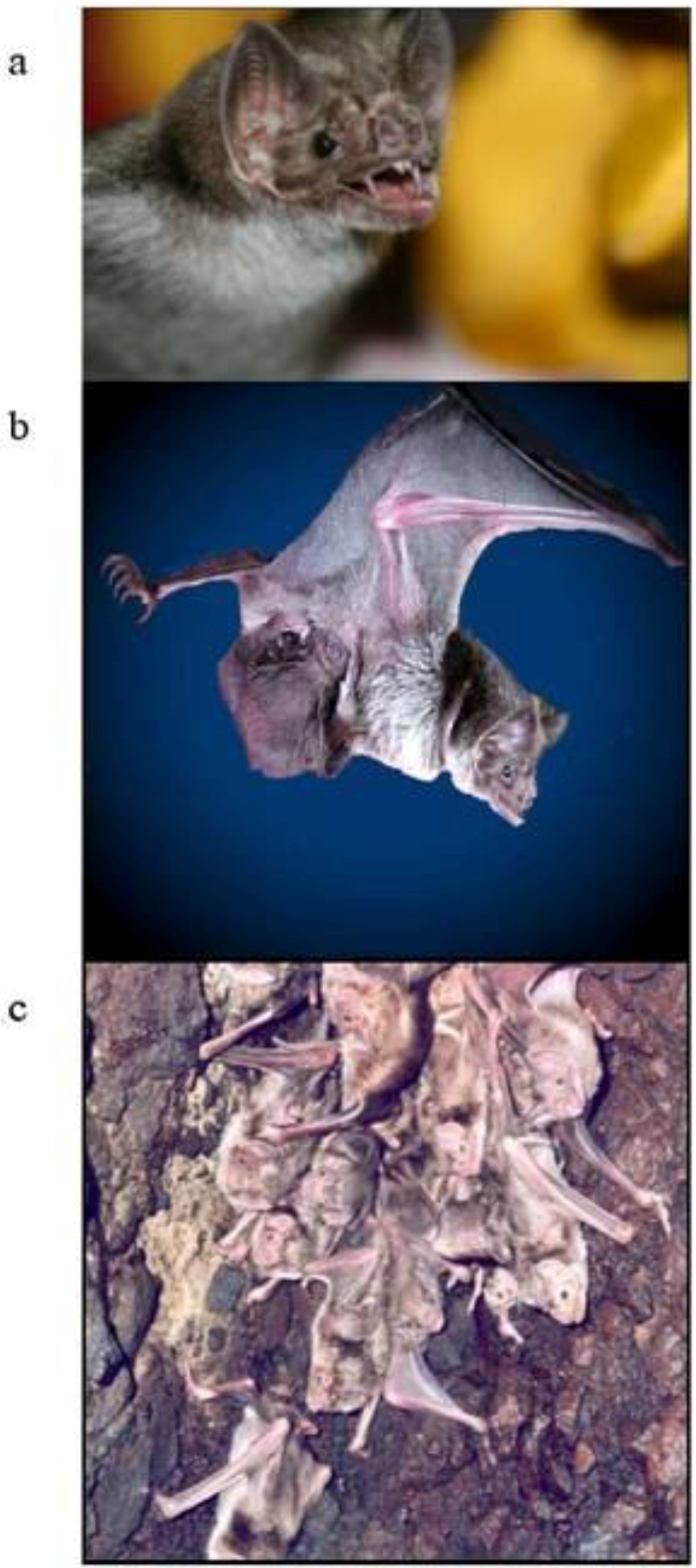

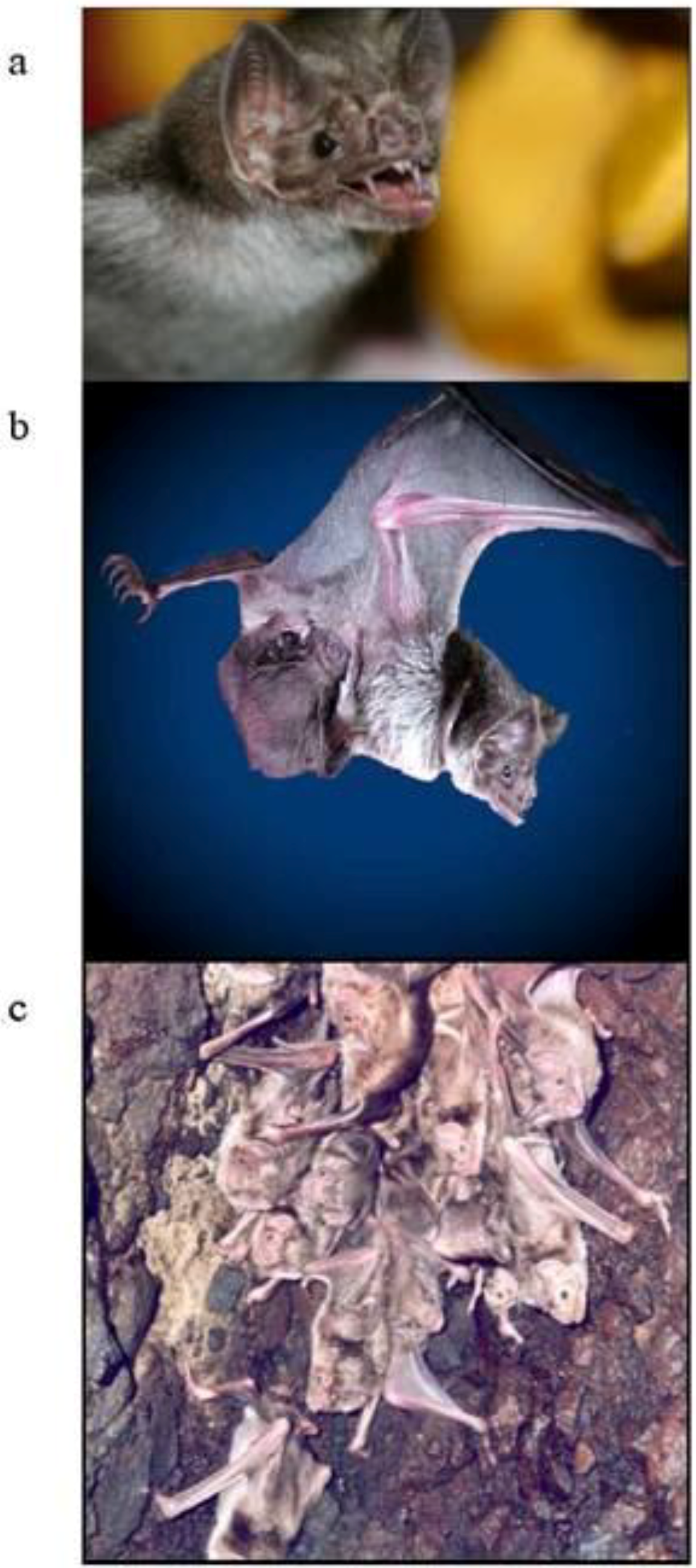{kind=link}
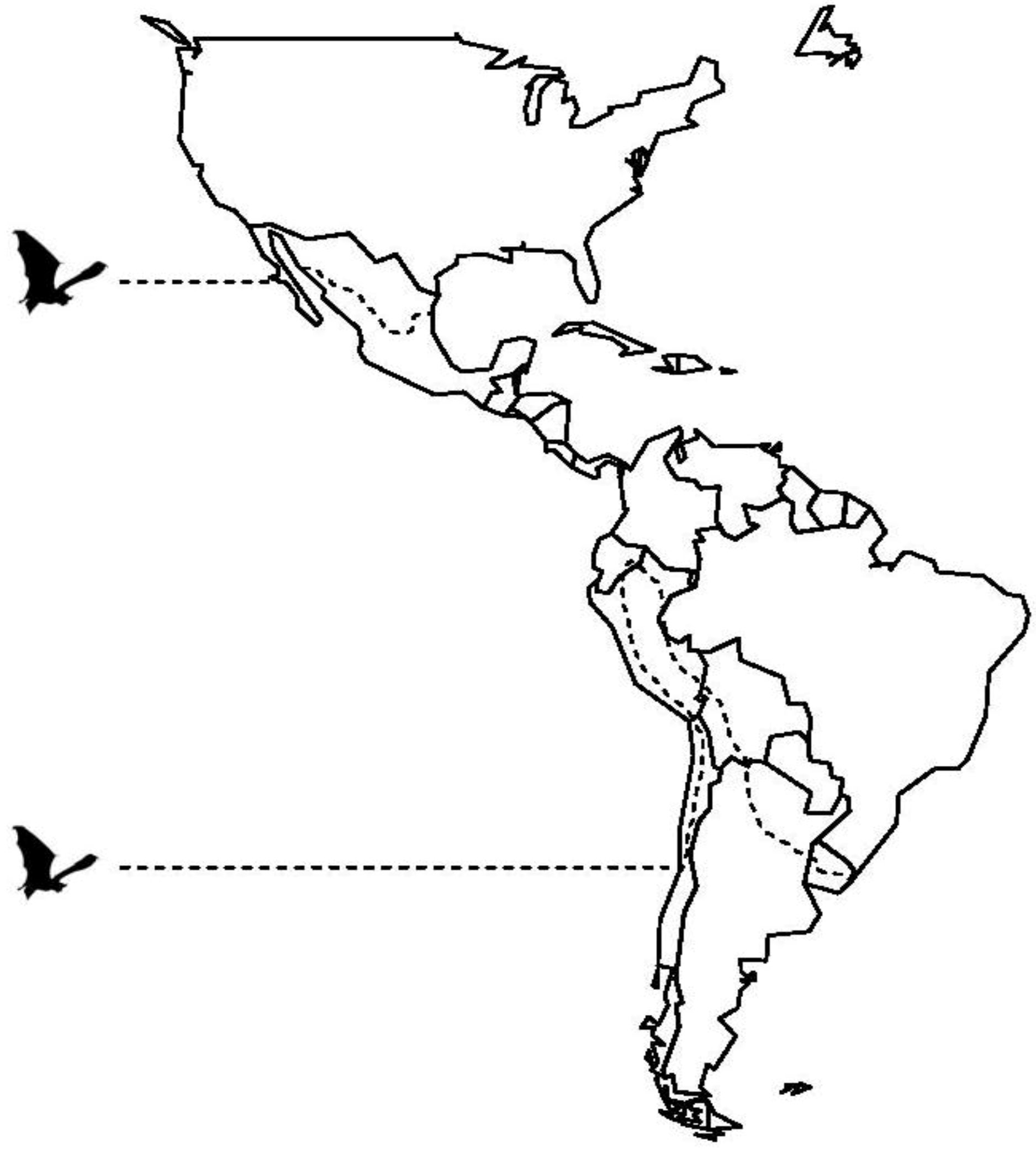{kind=link}
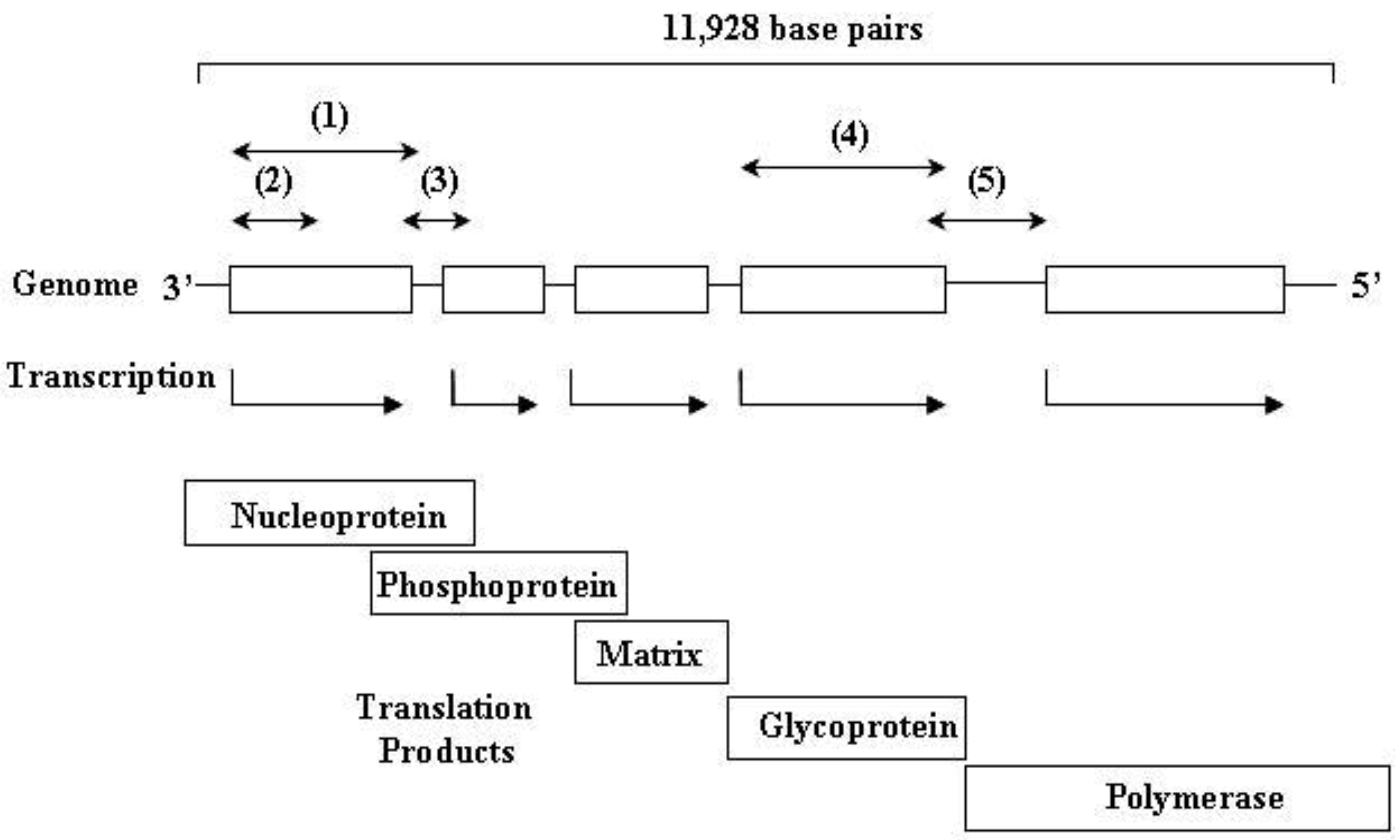{kind=link}
{kind=link}
| Species | Common name | Description |
|---|---|---|
| Desmodus rotundus | Common vampire bat | Weight: 30–40 g |
| Wingspan: 35–40 cm | ||
| Colony size: 20–1000 | ||
| Prey: mammals | ||
| Diaemus youngi | White-winged vampire bat | Weight: 30–45 g |
| Wingspan: 32–35 cm | ||
| Colony size: up to 30 | ||
| Prey: birds | ||
| Diphylla ecaudata | Hairy-legged vampire bat | Weight: 25–40 g |
| Wingspan: 37–45 cm | ||
| Colony size: 20–500 | ||
| Prey: mammals and birds |
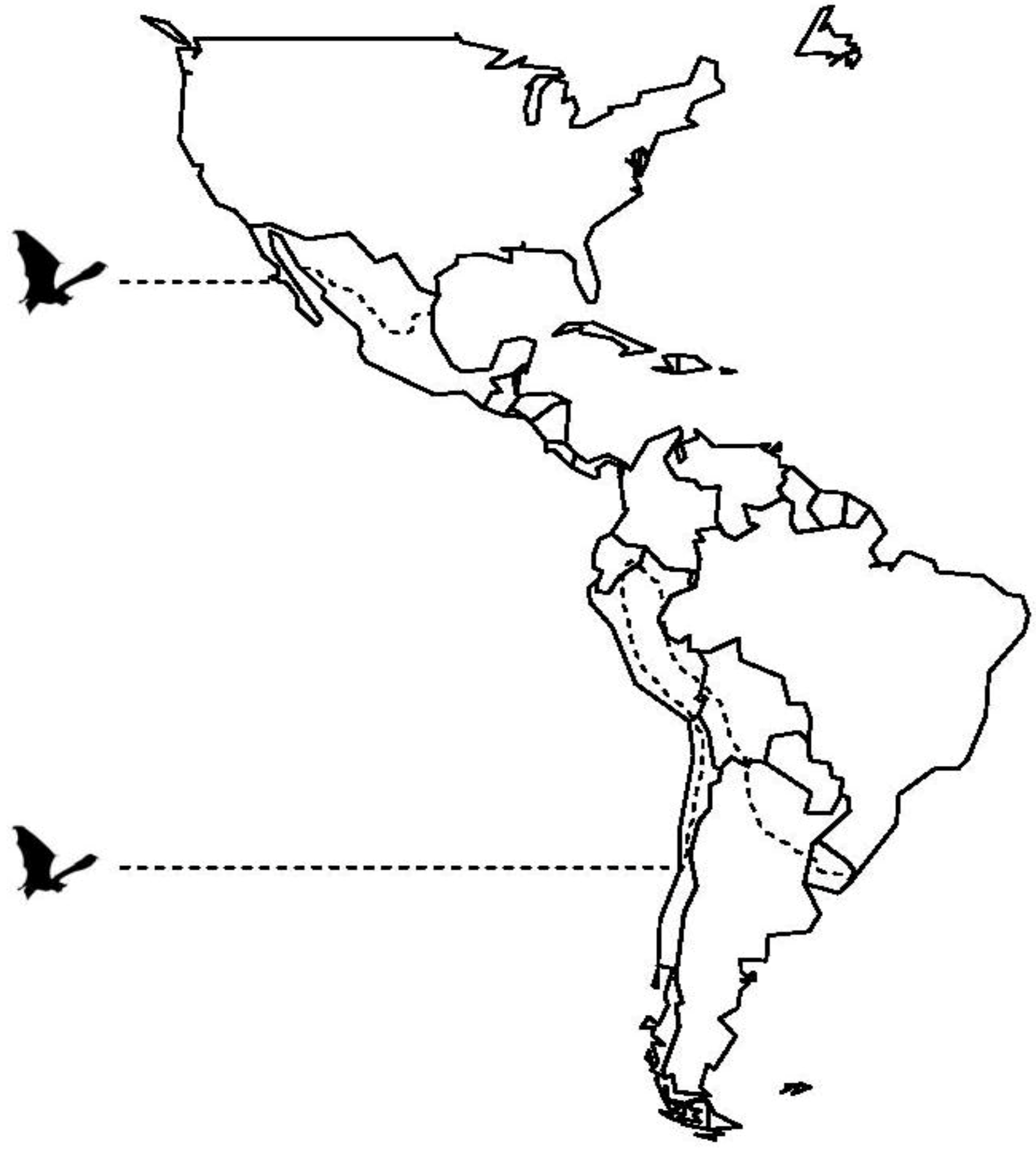
3. Rabies Virus Dynamics in Vampire Bat Populations
4. Virus Typing
4.1. Antigenic Typing
4.2. Phylogenetic Typing
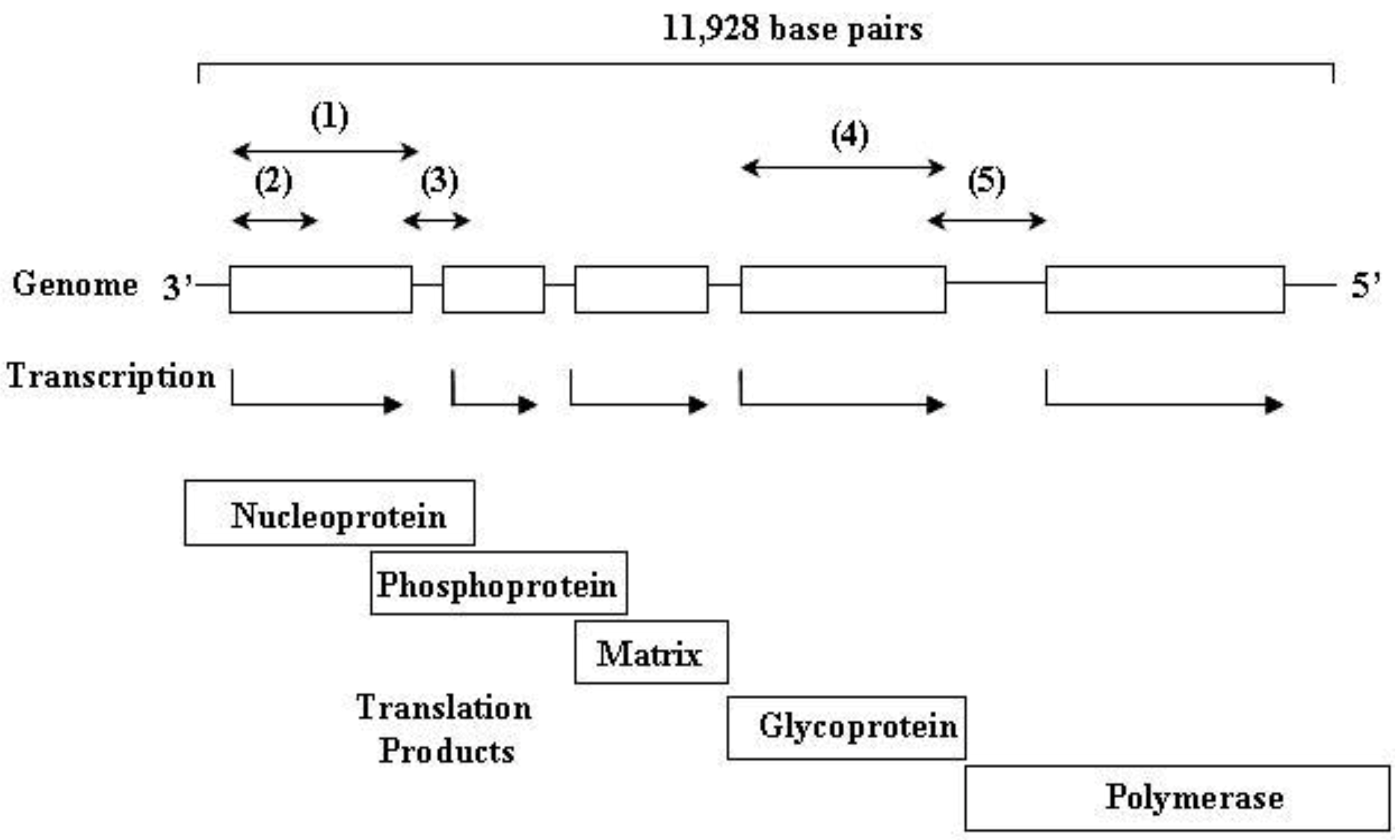
5. Impact on Livestock
| Country | 1982 | 2002 |
|---|---|---|
| Argentina | 92 | 13 |
| Bolivia | 159 | 59 |
| Brazil | 5900 | 1321 |
| Chile | 0 | 0 |
| Colombia | 139 | 47 |
| Ecuador | 45 | 16 |
| El Salvador | 7 | 19 |
| Guatemala | 22 | 11 |
| Honduras | 19 | 0 |
| Mexico | 35 | 154 |
| Nicaragua | 1 | 2 |
| Panama | 8 | 9 |
| Paraguay | 9 | 79 |
| Peru | 32 | 110 |
| Suriname | 0 | 5 |
| Venezuela | 54 | 19 |
| Year | Human | Dogs | Cattle | Bats | Other | Total |
|---|---|---|---|---|---|---|
| 2003 [53] | 1 | 75 | 201 (60.7) | 13 | 41 | 331 |
| 2004 [54] | 3 | 45 | 186 (69.7 | 0 | 33 | 267 |
| 2005 [55] | 8 | 103 | 252 (63.8) | 10 | 22 | 395 |
| 2006 [56] | 9 | 77 | 181 (62.0) | 5 | 20 | 292 |
| 2007 [57] | 0 | 42 | 227 (78.8) | 0 | 19 | 288 |
| 2008 [58] | 3 | 31 | 183 (77.9) | 0 | 18 | 235 |
| 2009 [59] | 4 | 12 | 134 (76.6) | 0 | 25 | 175 |
| 2010 [60] | 4 | 20 | 296 (82.0) | 0 | 41 | 361 |
| 2011 [61] | 3 | 20 | 121 (80.1) | 0 | 7 | 151 |
6. Impact on Public Health
Box 1. Case History: An imported case of human rabies in Louisiana, USA, due to a vampire bat bite.
7. Control of Vampire Bats and Vampire Bat Rabies
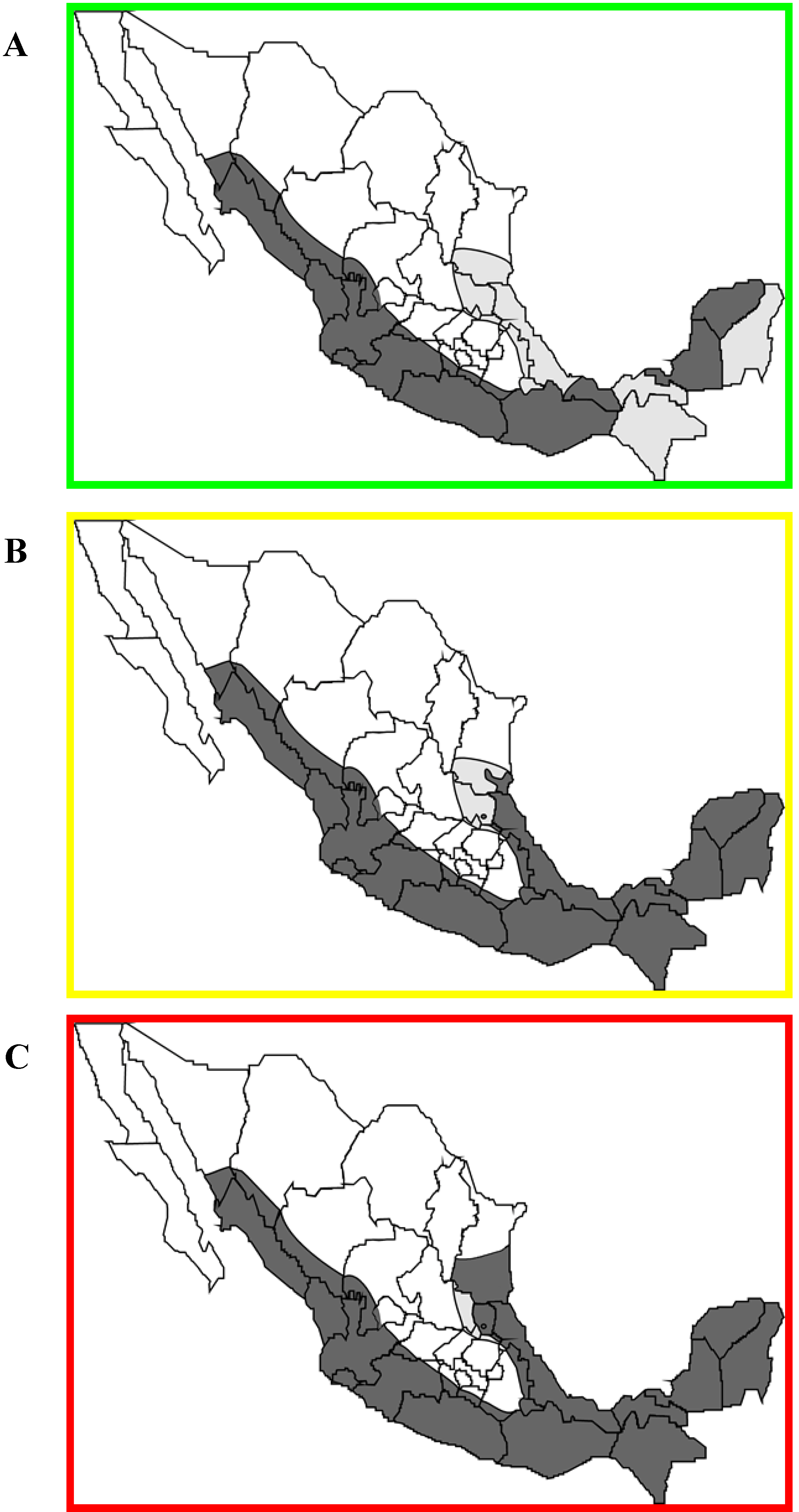
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Daszak, P.; Cunningham, A.A.; Hyatt, A.D. Emerging infectious diseases of wildlife-threats to biodiversity and human health. Science 2000, 287, 443–449. [Google Scholar] [CrossRef]
- Banyard, A.C.; Hayman, D.; Johnson, N.; McElhinney, L.M.; Fooks, A.R. Bats and Lyssaviruses. Adv. Virus Res. 2011, 79, 239–289. [Google Scholar] [CrossRef]
- Schneider, M.C.; Belotto, A.; Adé, M.P.; Hendrickx, S.; Leanes, L.F.; Rodrigues, M.J.; Medina, G.; Correa, E. Current status of human rabies transmitted by dogs in Latin America. Cad. Saúde Pública 2007, 23, 2049–2063. [Google Scholar] [CrossRef]
- McColl, K.A.; Tordo, N.; Aguilar-Setién, A. Bat lyssavirus infections. Rev. Sci. Tech. 2000, 19, 177–196. [Google Scholar]
- Sodre, M.M.; da Gama, A.R.; de Almeida, M.F. Updated list of bat species positive for rabies in Brazil. Rev. Inst. Med. Trop. Sao Paulo 2010, 52, 75–81. [Google Scholar]
- Schneider, M.C.; Romijn, P.C.; Uieda, W.; Tamayo, H.; da Silva, D.F.; Belotto, A.; da Silva, J.B.; Leanes, L.F. Rabies transmitted by vampire bats to humans: An emerging zoonotic disease in Latin America? Rev. Panam. Salad Publica 2009, 25, 260–269. [Google Scholar]
- De, Oviedo; Valdes, F. Sumario de la Natural historia de las Indias, 1526; (Spanish); Fond de Cultura Economica: Mexico City, Mexico, 1950. [Google Scholar]
- Carini, A. Sur une grande épizootie de rage (French). Ann. Instit. Past. 1911, 25, 843–846. [Google Scholar]
- Haupt, H.; Rehaag, B. Durch Fledermäuse verbreitete seuchenhafte Tolwut unter Viehbeständen in Santa Catharina (Süd-Brasilien) (German). Ztschr. F. Infectionskr. 1921, 22, 104–127. [Google Scholar]
- Belotto, A.; Leanes, L.F.; Schneider, M.C.; Tamayo, H.; Correa, F. Overview of rabies in the Americas. Virus Res. 2005, 111, 5–12. [Google Scholar] [CrossRef]
- Hurst, E.W.; Pawan, J.L. A further account of the Trinidad outbreak of acute rabies myelitis. J. Path. Bacteriol. 1932, 35, 301–321. [Google Scholar] [CrossRef]
- Pawan, J.L. The transmission of paralytic rabies in Trinidad by the Vampire bat (Desmodus rotundus murinus Wagner, 1840). Ann. Trop. Med. Parasitol. 1936, 30, 101–130. [Google Scholar]
- Streicker, D.G.; Recuenco, S.; Valderrama, W.; Benavides, J.G.; Vargas, I.; Pacheco, V.; Condori, R.E.; Montgomery, J.; Rupprecht, C.E.; Rohani, P.; et al. Ecological and anthropongenic drivers of rabies exposure in vampire bats: Implications for transmission and control. Proc. Biol. Sci. 2012, 279, 3384–3392. [Google Scholar] [CrossRef]
- Delpietro, H.A.; Marchevsky, N.; Simonetti, E. Relative population densities and predation of the common vampire bat (Desmodus rotundus) in natural and cattle-raising areas in north-east Argentina. Prev. Vet. Med. 1992, 14, 13–20. [Google Scholar] [CrossRef]
- Belwood, J.J.; Morton, P.A. Vampires: The real story. Bats Mag. 1991, 9, 11–16. [Google Scholar]
- Johnson, H.N. Derriengue: Vampire bat rabies in Mexico. Am. J. Hyg. 1948, 47, 189–204. [Google Scholar]
- Real, L.A.; Russell, C.; Waller, L.; Smith, D.L.; Childs, J. Spatial dynamics and molecular ecology of North American rabies. J. Hered. 2005, 96, 253–260. [Google Scholar] [CrossRef]
- Langoni, H.; Souza, L.C.; Zetun, C.B.; Silva, T.C.C.; Hoffmann, J.L.; Silva, R.C. Serological survey for rabies in serum samples from vampire bats (Desmodus rotundus) in Botucatu Region, Sp., Brazil. J. Venom. Anim. Toxins Trop. Dis. 2008, 14, 651–659. [Google Scholar] [CrossRef]
- Davies, J.S.; Nicolay, C.W.; Williams, S.H. A comparative study of incisor procumbency and mandibular morphology in vampire bats. J. Morphol. 2010, 271, 853–862. [Google Scholar]
- Arellano-Sota, C. Biology, ecology, and control of the vampire bat. Rev. Infect. Dis. 1988, 10, S615–S619. [Google Scholar] [CrossRef]
- Constantine, D.G. Bat Rabies: Current Knowledge and Future Research. In Rabies; Nagano, Y., Davenport, M., Eds.; University Park Press: Baltimore, MD, USA, 1971; p. 253. [Google Scholar]
- Delpietro, H.; de Díaz, A.M.; Fuenzalida, E.; Bell, J.F. Determination of the rate of rabies attack in bats. Bol. Oficina. Sanit. Panam. 1972, 73, 222–230. [Google Scholar]
- Lord, D.R.D.; Fuenzalida, E.; Delpietro, H.; Larghi, O.; de Diaz, A.M.; Lázaro, L. Observations on the epizootiology of vampire bat rabies. Bull. Pan. Am. Health. Organ. 1975, 9, 189–195. [Google Scholar]
- Blackwood, J.C.; Streicker, D.G.; Altizer, D.; Rohani, P. Resolving the roles of immunity, pathogenesis, and immigration for rabies persistence in vampire bats. Proc. Natl. Acad. Sci. USA 2013, 110, 20837–20842. [Google Scholar] [CrossRef]
- Lord, R.D. An ecological strategy for controlling bovine rabies through elimination of vampire bats. In Proceedings of the 9th Vertebrate Pest Conference, Fresno, USA, March, 1980; p. 22.
- Mungrue, K.; Mahabir, R. The rabies epidemic in Trinidad of 1923 to 1937: An evaluation with a geographic information system. Wilderness Environ. Med. 2011, 22, 28–36. [Google Scholar] [CrossRef]
- Gilbert, A.T.; Petersen, B.W.; Recuenco, S.; Niezgoda, M.; Gómez, J.; Laguna-Torres, V.A.; Rupprecht, C.E. Evidence of rabies virus exposure among humans in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2012, 87, 206–215. [Google Scholar] [CrossRef]
- Moreno, J.; Baer, G.M. Experimental rabies in the vampire bat. Am. J. Trop. Med. Hyg. 1980, 29, 254–259. [Google Scholar]
- Aguilar-Setién, A.; Loza-Rubio, E.; Salas-Rojas, M.; Brisseau, N.; Cliquet, F.; Pastoret, P.P.; Rojas-Dotor, S.; Tesoro, E.; Kretschmer, R. Salivary excretion of rabies virus in healthy vampire bats. Epidemiol. Infect. 2005, 133, 517–522. [Google Scholar] [CrossRef]
- Almeida, M.F.; Martorelli, L.F.A.; Aires, C.C.; Sallum, P.C.; Dungun, E.L.; Massad, E. Experimental rabies infection in haematophagus bats Desmodus rotundus. Epidemiol. Infect. 2005, 133, 523–527. [Google Scholar] [CrossRef]
- Carniero, A.J.B.; Franke, C.R.; Stöcker, A.; dos Santos, F.; de Sa, J.F.U.; Moraes-Silva, E.; Alves, J.N.; Brunink, S.; Corman, V.M.; Drosten, C.; et al. Rabies virus RNA in naturally infected vampire bats, Northeastern Brazil. Emerg. Infect. Dis. 2010, 12, 20040–20046. [Google Scholar]
- Johnson, N.; Wakeley, P.R.; Brookes, S.M.; Fooks, A.R. European bat lyssavirus type 2 RNA in Myotis daubentonii. Emerg. Infect. Dis. 2006, 12, 1142–1144. [Google Scholar] [CrossRef]
- Marston, D.A.; McElhinney, L.M.; Johnson, N.; Müller, T.; Conzelmann, K.K.; Tordo, N.; Fooks, A.R. Comparative analysis of the full genome sequence of European bat lyssavirus type 1 and type 2 with other lyssaviruses and evidence for a conserved transcription termination and polyadenylation motif in the G-L 3' non-translated region. J. Gen. Virol. 2007, 88, 1302–1314. [Google Scholar] [CrossRef]
- Wiktor, T.J.; Koprowski, H. Monoclonal antibodies against rabies virus produced by somatic cell hybridization: Detection of antigenic variants. Proc. Natl. Acad. Sci. USA 1978, 75, e3928. [Google Scholar] [CrossRef]
- Rupprecht, C.E.; Glickman, L.T.; Spencer, P.A.; Wiktor, T.J. Epidemiology of rabies virus variants. Differentiation using monoclonal antibodies and discriminant analysis. Am. J. Epidemiol. 1987, 126, 298–309. [Google Scholar] [CrossRef]
- Velasco-Villa, A.; Gómez-Sierra, M.; Hernández-Rodríguez, G.; Juárez-Islas, V.; Meléndez-Félix, A.; Vargos-Pino, F.; Velázquez-Monroy, O.; Flisser, A. Antigenic diversity and distribution of rabies virus in Mexico. J. Clin. Microbiol. 2002, 40, 951–958. [Google Scholar] [CrossRef]
- Nadin-Davis, S.A.; Loza-Rubio, E. The molecular epidemiology of rabies associated with chiropteran hosts in Mexico. Virus Res. 2006, 117, 215–226. [Google Scholar] [CrossRef]
- Sacramento, D.; Bourhy, H.; Tordo, N. PCR technique as an alternative method for diagnosis and molecular epidemiology of rabies virus. Mol. Cell. Probes 1989, 6, 229–240. [Google Scholar]
- Bourhy, H.; Kissi, B.; Audry, L.; Smreczak, M.; Sadkowska-Todys, H.; Kulonen, K.; Tordo, N.; Zmudzinski, J.F.; Holmes, E.C. Ecology and evolution of rabies virus in Europe. J. Gen. Virol. 1999, 80, 2545–2557. [Google Scholar]
- Talbi, C.; Holmes, E.C.; de Benedictis, P.; Faye, O.; Nakouné, E.; Gamatié, D.; Diarra, A.; Elmanny, B.O.; Sow, A.; Adjogoua, E.V.; et al. Evolutionary history and dynamics of dog rabies virus in western and central Africa. J. Gen. Virol. 2009, 90, 783–791. [Google Scholar] [CrossRef]
- Johnson, N.; Black, C.; Smith, J.; Un, H.; McElhinney, L.M.; Aylan, O.; Fooks, A.R. Rabies emergence among foxes in Turkey. J. Wildl. Dis. 2003, 39, 262–270. [Google Scholar] [CrossRef]
- Condori-Condori, R.E.; Streicker, D.G.; Cabezas-Sanchez, C.; Velasco-Villa, A. Enzootic and epizootic rabies associated with vampire bats, Peru. Emerg. Infect. Dis. 2013, 19, 1463–1469. [Google Scholar]
- Sabeta, C.T.; Bingham, J.; Nel, L.H. Molecular epidemiology of canid rabies in Zimbabwe and South Africa. Virus Res. 2003, 91, 203–211. [Google Scholar] [CrossRef]
- Johnson, N.; McElhinney, L.M.; Smith, J.; Lowings, P.; Fooks, A.R. Phylogenetic comparison of the genus Lyssavirus using distal coding sequences of the glycoprotein and nucleoprotein. Arch. Virol. 2003, 147, 2111–2123. [Google Scholar]
- Streicker, D.G.; Turmelle, A.S.; Vonhof, M.J.; Kuzmin, I.V.; McCracken, G.F.; Rupprecht, C.E. Host phylogeny contrains cross-species emergence and establishment of rabies virus in bats. Science 2010, 329, 676–679. [Google Scholar] [CrossRef]
- Forment, W.L.; Schmidt, U.; Greenhall, A.M. Movement and population studies of the vampire bat (Desmodus rotundus) in Mexico. J. Mammol. 1971, 52, 227–228. [Google Scholar] [CrossRef]
- Shoji, Y.; Kobayashi, Y.; Sato, G.; Itou, T.; Miura, Y.; Mikami, T.; Cunha, E.M.; Samara, S.I.; Carvalho, A.A.; Nocitti, D.P.; et al. Genetic characterization of rabies viruses isolated from frugivorous bat (Artibeus spp.) in Brazil. J. Med. Vet. Sci. 2004, 66, 1271–1273. [Google Scholar] [CrossRef]
- Kobayashi, Y.; Sato, G.; Shoji, Y.; Sato, T.; Itou, T.; Cunham, E.M.; Samara, S.I.; Carvalho, A.A.; Nocitti, D.P.; Ito, F.H.; et al. Molecular epidemiological analysis of bat rabies viruses in Brazil. J. Med. Vet. Sci. 2005, 67, 647–652. [Google Scholar] [CrossRef]
- Velasco-Villa, A.; Orciari, L.A.; Juarez-Islas, V.; Gómez-Sierra, M.; Castillo, A.; Flisser, A.; Rupprecht, C.E. Molecular diversity of rabies viruses associated with bats in Mexico and other countries of the Americas. J. Clin. Microbiol. 2006, 44, 1697–1710. [Google Scholar] [CrossRef]
- Albas, A.; Campos, A.C.; Araujo, D.B.; Rodrigues, C.S.; Sodré, M.M.; Dungon, E.L.; Favoretto, S.R. Molecular characterization of rabies virus isolated from nonhaematophagous bats in Brazil. Rev. Soc. Bras. Med. Trop. 2011, 44, 678–683. [Google Scholar] [CrossRef]
- Rawlins, S.C. Current trends in screwworm myiasis in the Caribbean region. Vet. Parasitol. 1985, 18, 241–250. [Google Scholar] [CrossRef]
- Sulkin, S.E.; Allen, R. Virus infection in bats. Monogr. Virol. 1974, 8, 1–103. [Google Scholar]
- Krebs, J.W.; Mandel, E.J.; Swerdlow, D.L.; Rupprecht, C.E. Rabies surveillance in the United States during 2003. J. Am. Vet. Med. Assoc. 2004, 225, 1837–1849. [Google Scholar] [CrossRef]
- Krebs, J.W.; Mandel, E.J.; Swerdlow, D.L.; Rupprecht, C.E. Rabies surveillance in the United States during 2004. J. Am. Vet. Med. Assoc. 2005, 227, 1912–1925. [Google Scholar] [CrossRef]
- Blanton, J.D.; Krebs, J.W.; Hanlon, C.A.; Rupprecht, C.E. Rabies surveillance in the United States during 2005. J. Am. Vet. Med. Assoc. 2006, 229, 1897–1911. [Google Scholar] [CrossRef]
- Blanton, J.D.; Hanlon, C.A.; Rupprecht, C.E. Rabies surveillance in the United States during 2006. J. Am. Vet. Med. Assoc. 2007, 231, 540–556. [Google Scholar] [CrossRef]
- Blanton, J.D.; Palmer, D.; Christian, K.A.; Rupprect, C.E. Rabies surveillance in the United States during 2007. J. Am. Vet. Med. Assoc. 2008, 233, 884–897. [Google Scholar] [CrossRef]
- Blanton, J.D.; Robertson, K.; Palmer, D.; Rupprecht, C.E. Rabies surveillance in the United States during 2008. J. Am. Vet. Med. Assoc. 2009, 235, 676–689. [Google Scholar] [CrossRef]
- Blanton, J.D.; Palmer, D.; Rupprecht, C.E. Rabies surveillance in the United States during 2009. J. Am. Vet. Med. Assoc. 2010, 237, 646–657. [Google Scholar] [CrossRef]
- Blanton, J.D.; Palmer, D.; Dyer, J.; Rupprecht, C.E. Rabies surveillance in the United States during 2010. J. Am. Vet. Med. Assoc. 2011, 239, 773–783. [Google Scholar] [CrossRef]
- Blanton, J.D.; Dyer, J.; Brayer, J.; Rupprecht, C.E. Rabies surveillance in the United States during 2011. J. Am. Vet. Med. Assoc. 2012, 241, 712–722. [Google Scholar] [CrossRef]
- SIEPI Sistema de Información Epidemiológica. Available online: http://siepi.panaftosa.org.br (accessed on 11 January 2013).
- Balsamo, G.; Ratard, R.C.; Thoppil, D.R.; Thoppil, M.; Pino, F.V.; Rupprecht, C.E.; Sprecher, A.G.; Petersen, B.W. Human rabies from exposure to a vampire bat in Mexico-Louisiana, 2010. MMWR 2011, 60, 1050–1052. [Google Scholar]
- Aranda, M.; López-de Buen, L. Rabies in skunks from Mexico. J. Wild. Dis. 1999, 35, 574–577. [Google Scholar] [CrossRef]
- Loza-Rubio, E.; Aguilar-Setién, A.; Bahloul, C.; Brochier, B.; Pastoret, P.P.; Tordo, N. Discrimination between epidemiological cycles of rabies in Mexico. Arch. Med. Res. 1999, 30, 144–149. [Google Scholar]
- Delpietro, H.A.; Russo, R.G. Ecological and epidemiologic aspects of the attacks by vampire bats and paralytic rabies in Argentina and analysis of the proposals carried out for their control. Rev. Sci. Tech. 1996, 15, 971–984. [Google Scholar]
- Avila-Flores, R.; Medellín, R.A. Ecological, taxonomic, and physiological correlates of cave use by Mexican bats. J. Mammol. 2004, 85, 675–687. [Google Scholar] [CrossRef]
- Aguilar-Setién, A.; Mexican Social Security Institute, Mexico City, Mexico; Aréchiga-Ceballos, N.; Institute of Epidemiology Diagnostic and Reference, Mexico City, Mexico. Unpublished data. 2013.
- Wilkinson, G.S. Social grooming in the common vampire bat, Desmodus rotundus. Anim. Behav. 1986, 34, 1880–1889. [Google Scholar] [CrossRef]
- Fornes, A.; Lord, R.D.; Kuns, M.L.; Larghi, O.P.; Fuenzalida, E.; Lazara, L. Control of bovine rabies through vampire bat control. J. Wildl. Dis. 1974, 10, 310–316. [Google Scholar] [CrossRef]
- Mayen, F. Haematophagous bats in Brazil, their role in rabies transmission, impact on public health, livestock industry and alternatives to an indiscriminate reduction of bat population. J. Vet. Med. B 2003, 50, 469–472. [Google Scholar] [CrossRef]
- Serrano, H.; Pérez Rivero, J.J.; Aguilar Setién, A.; de Paz, O.; Villa-Godoy, A. Vampire bat reproductive control by naturally occurring Phytoestrogen. Reprod. Fertil. Devel. 2007, 19, 470–472. [Google Scholar] [CrossRef]
- Crespo, R.F.; Fernández, S.; de Anda López, D.; Valarde, F.I.; Anaya, R.M. Intramuscular inoculation of cattle with warfarin: A new technique for the control of vampire bats. Bull. Pan. Am. Health Organ. 1979, 87, 283–299. [Google Scholar]
- Aguilar-Sétien, A.; Brochier, B.; Tordo, N.; de Paz, O.; Desmettre, P.; Péharpé, D.; Pastoret, P.P. Experimental rabies infection and oral vaccination in vampire bats (Desmodus rotundus). Vaccine 1998, 16, 1122–1126. [Google Scholar]
- Aguilar-Setién, A.; Campos, Y.L.; Cruz, E.T.; Kretschmer, R.; Brochier, B.; Pastoret, P.P. Vaccination of vampire bats using recombinant vaccinia-rabies virus. J. Wildl. Dis. 2002, 38, 539–544. [Google Scholar] [CrossRef]
- Gut, H.J. A pleistocene vampire bat from Florida. J. Mammol. 1959, 40, 534–538. [Google Scholar] [CrossRef]
- Mistry, S.; Moreno-Valdez, A. Climate change and bats: Vampire bats offer clues to the future. Bats 2008, 26, 8–11. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Johnson, N.; Aréchiga-Ceballos, N.; Aguilar-Setien, A. Vampire Bat Rabies: Ecology, Epidemiology and Control. Viruses 2014, 6, 1911-1928. https://doi.org/10.3390/v6051911
Johnson N, Aréchiga-Ceballos N, Aguilar-Setien A. Vampire Bat Rabies: Ecology, Epidemiology and Control. Viruses. 2014; 6(5):1911-1928. https://doi.org/10.3390/v6051911
Chicago/Turabian StyleJohnson, Nicholas, Nidia Aréchiga-Ceballos, and Alvaro Aguilar-Setien. 2014. "Vampire Bat Rabies: Ecology, Epidemiology and Control" Viruses 6, no. 5: 1911-1928. https://doi.org/10.3390/v6051911
APA StyleJohnson, N., Aréchiga-Ceballos, N., & Aguilar-Setien, A. (2014). Vampire Bat Rabies: Ecology, Epidemiology and Control. Viruses, 6(5), 1911-1928. https://doi.org/10.3390/v6051911