The Role of Bacterial Chaperones in the Circulative Transmission of Plant Viruses by Insect Vectors


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction — Circulative Transmission of Plant Viruses
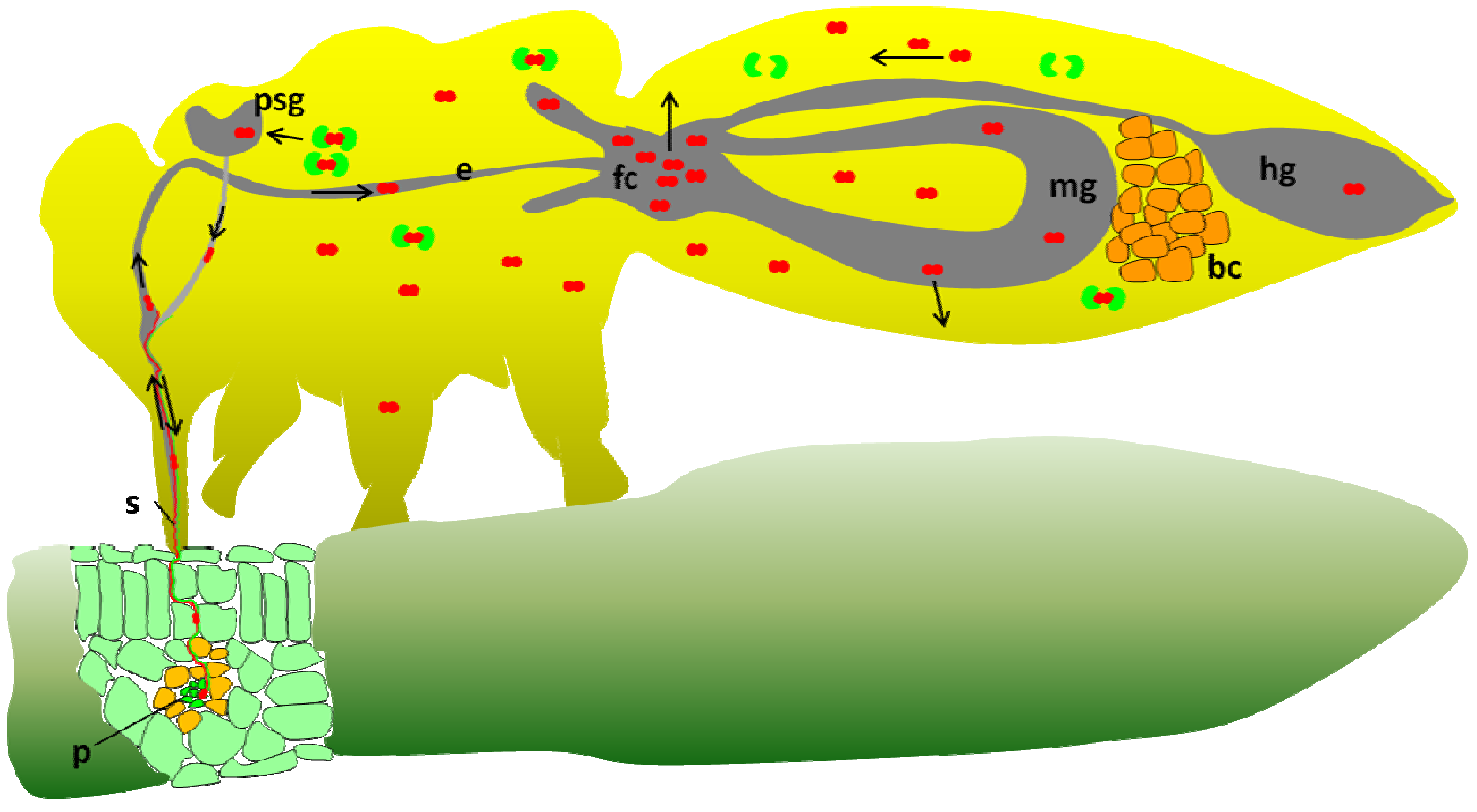
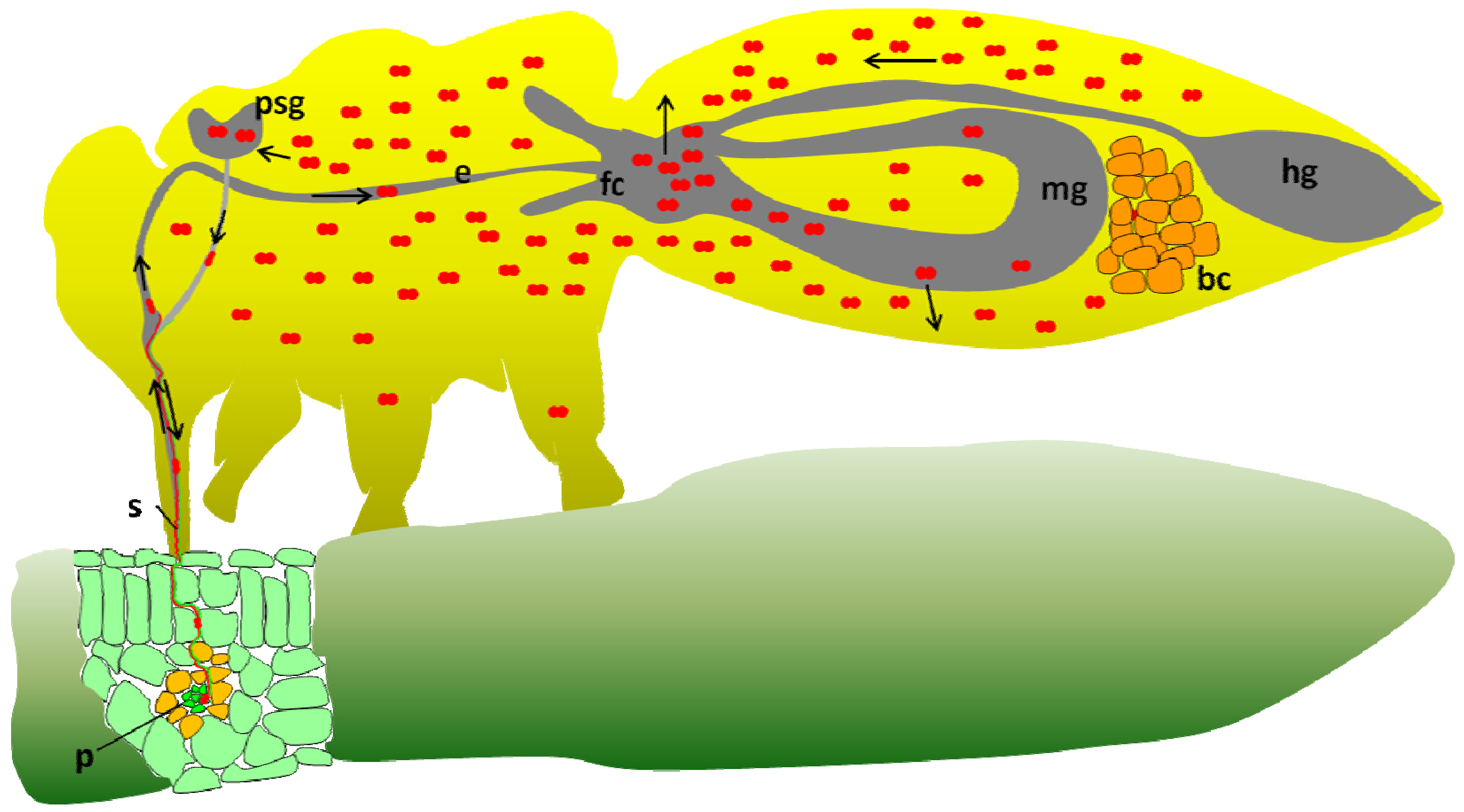
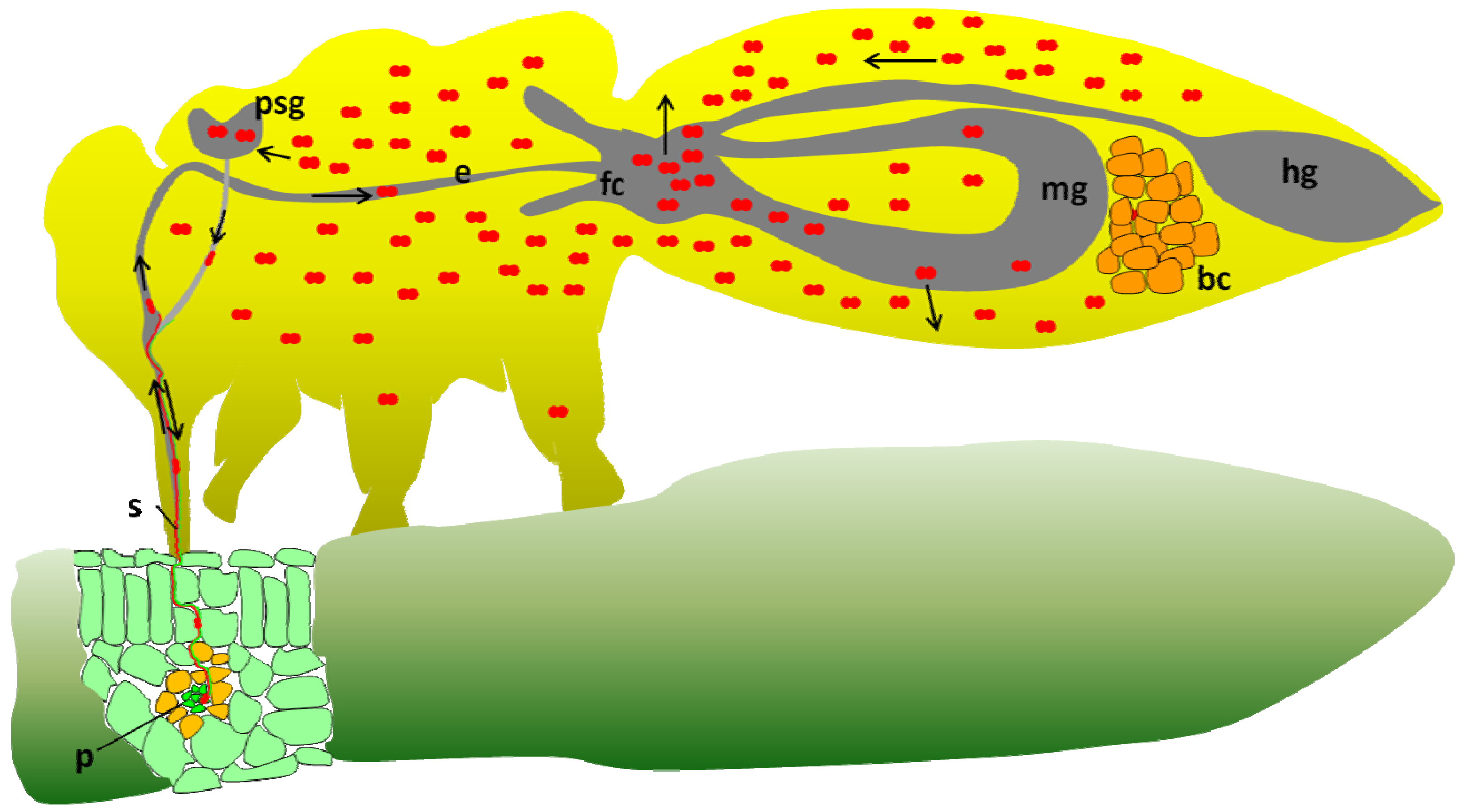
2. Potato Leafroll Virus (PLRV) Transmission by the Peach Potato Aphid Myzus persicae and the Coat Protein Role
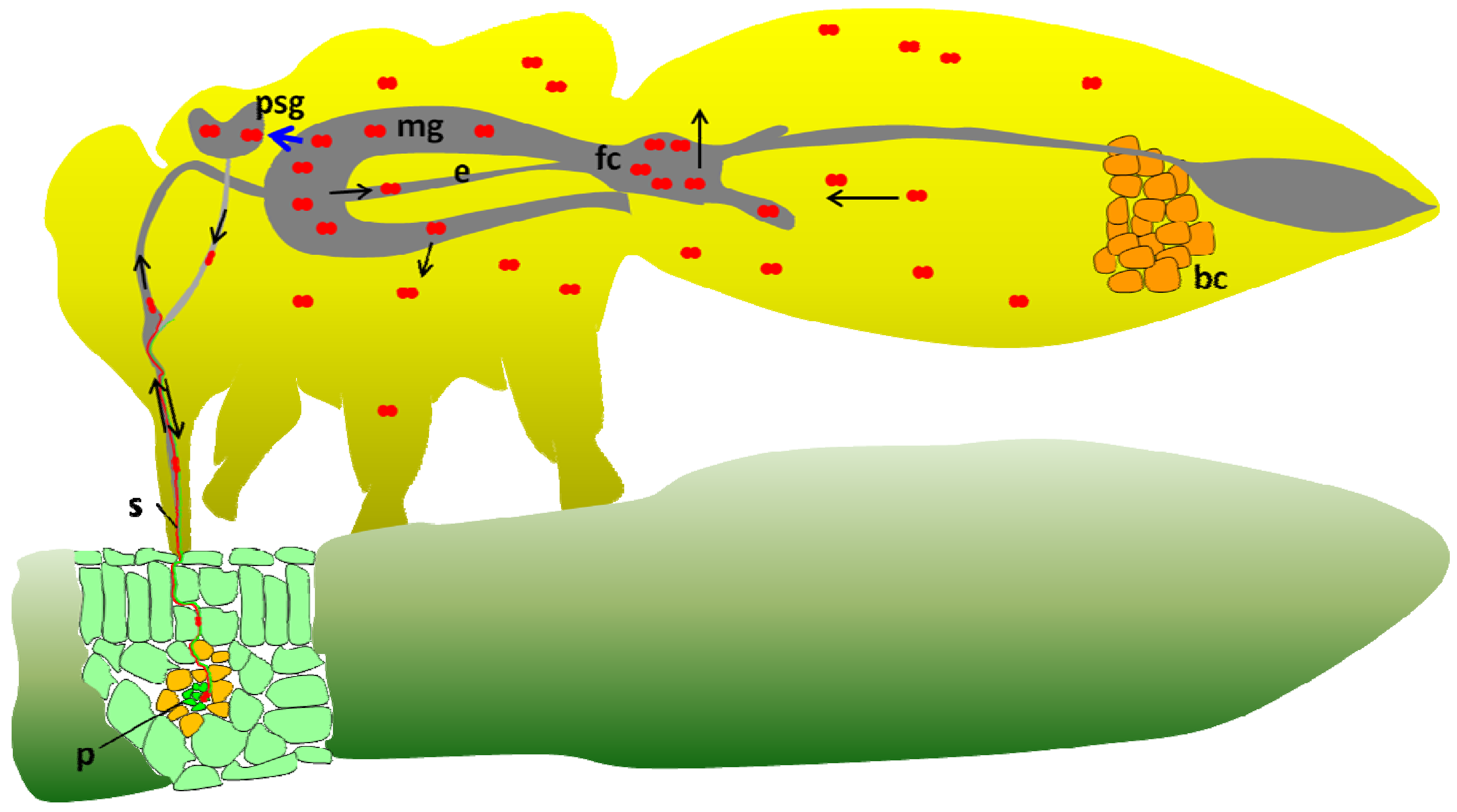
3. Tomato Yellow Leaf Curl Virus (TYLCV) Transmission by the Sweetpotato Whitefly Bemisia tabaci
4. Proteins Influencing the Circulative Transmission of Plant Viruses
5. Bacterial GroEL Protein, Structure and Function, Including Symbiotic Bacteria of Insects
6. Implication of GroEL Proteins in the Circulative Transmission of Aphid Transmitted Luteoviruses
7. Implication of the GroEL Protein in the Circulative Transmission of TYLCV


8. Conclusions
Acknowledgments
Conflict of interests
References and Notes
- Gergerich, R.C.; Scott, H.A. Determinants in the specificity of virus transmission by leaf-feeding beetles. In Advances in Disease Vector Research; Springer: New York, NY, USA, 1991; pp. 1–13. [Google Scholar]
- Ammar, E. Propagative transmission of plant and animal viruses by insects: Factors affecting vector specificity and competence. Adv. Dis. Vector Res 1994, 10, 289–331. [Google Scholar] [CrossRef]
- Nuttall, P.A.; Jones, L.D.; Davies, C.R. The role of arthropod vectors in arbovirus evolution. In Advances in Disease Vector Research; Springer: New York, NY, USA, 1991; pp. 15–45. [Google Scholar]
- Weaver, S.C. Vector biology in viral pathogenesis. In Viral Pathogenesis; Lippincott-Raven: New York, NY, USA, 1997; pp. 329–352. [Google Scholar]
- Brown, J.K.; Czosnek, H. Whitefly transmission of plant viruses. Adv. Bot. Res. 2002, 36, 65–100. [Google Scholar] [CrossRef]
- Ghanim, M.; Morin, S.; Czosnek, H. Rate of Tomato yellow leaf curl virus translocation in the circulative transmission pathway of its vector, the whitefly Bemisia tabaci. Phytopathology 2001, 91, 188–196. [Google Scholar] [CrossRef]
- Caciagli, P.; Bosco, D. Quantitation over time of tomato yellow leaf curl geminivirus DNA in its whitefly vector. Phytopathology 1997, 87, 610–613. [Google Scholar] [CrossRef]
- Cohen, S.; Duffus, J.; Ursen, R.; Liu, H.; Flock, R. Purification, serology, and vector relationships of squash leaf curl virus, a whitefh-transmitted geminivirus. Phytopathology 1983, 73, 1669–1673. [Google Scholar] [CrossRef]
- Muniyappa, V.; Venkatesh, H.; Ramappa, H.; Kulkarni, R.; Zeidan, M.; Tarba, C.-Y.; Ghanim, M.; Czosnek, H. Tomato leaf curl virus from Bangalore (ToLCV-Ban4): Sequence comparison with Indian ToLCV isolates, detection in plants and insects, and vector relationships. Arch. Virol. 2000, 145, 1583–1598. [Google Scholar] [CrossRef]
- Ghanim, M.; Morin, S.; Zeidan, M.; Czosnek, H. Evidence for transovarial transmission of tomato yellow leaf curl virus by its vector, the whitefly Bemisia tabaci. Virology 1998, 240, 295–303. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; Ammar, E.D.; Whitfield, A.E.; Redinbaugh, M.G. Insect vector interactions with persistently transmitted viruses. Annu. Rev. Phytopathol. 2008, 46, 327–359. [Google Scholar] [CrossRef]
- Power, A.G. Insect transmission of plant viruses: A constraint on virus variability. Curr. Opin. Plant Biol. 2000, 3, 336–340. [Google Scholar] [CrossRef]
- Stanley, J. Subviral DNAs associated with geminivirus disease complexes. Vet. Microbiol. 2004, 98, 121–129. [Google Scholar] [CrossRef]
- Gray, S.; Gildow, F.E. Luteovirus-aphid interactions. Annu. Rev. Phytopathol. 2003, 41, 539–566. [Google Scholar] [CrossRef]
- Gray, S.M.; Banerjee, N. Mechanisms of arthropod transmission of plant and animal viruses. Microbiol. Mol. Biol. Rev. 1999, 63, 128–148. [Google Scholar]
- Czosnek, H.; Ghanim, M.; Ghanim, M. The circulative pathway of begomoviruses in the whitefly vector Bemisia tabaci—Insights from studies with Tomato yellow leaf curl virus. Ann. Appl. Biol. 2002, 140, 215–231. [Google Scholar] [CrossRef]
- Hunter, W.; Hiebert, E.; Webb, S.; Tsai, J.; Polston, J. Location of geminiviruses in the whitefly Bemisia tabaci (Homoptera: Aleyrodidae). Plant Dis. 1998, 82, 1147–1151. [Google Scholar] [CrossRef]
- Skaljac, M.; Ghanim, M. Tomato yellow leaf curl disease and plant-virus vector interactions. Isr. J. Plant Sci. 2010, 58, 103–111. [Google Scholar] [CrossRef]
- Ghanim, M.; Medina, V. Localization of Tomato yellow leaf curl virus in its whitefly vector Bemisia tabaci. In Tomato Yellow Leaf Curl Virus Disease; Springer: Dordrecht, The Netherlands, 2007; pp. 171–183. [Google Scholar]
- Garret, A.; Kerlan, C.; Thomas, D. The intestine is a site of passage for potato leafroll virus from the gut lumen into the haemocoel in the aphid vector, Myzus persicae Sulz. Arch. Virol. 1993, 131, 377–392. [Google Scholar] [CrossRef]
- Hogenhout, S.A.; van Der Wilk, F.; Verbeek, M.; Goldbach, R.W.; van den Heuvel, J.F.J.M. Identifying the determinants in the equatorial domain of buchnera GroEL implicated in binding potato leafroll virus. J. Virol. 2000, 74, 4541–4548. [Google Scholar] [CrossRef]
- Ponsen, M.B. The Site of Potato Leafroll Virus Multiplication in Its Vector, Myzus Persicae: An Anatomical Study; H. Veenman: Wageningen, The Netherlands, 1972. [Google Scholar]
- Wang, H.; Nuttall, P. Excretion of host immunoglobulin in tick saliva and detection of IgG-binding proteins in tick haemolymph and salivary glands. Parasitology 1994, 109, 525–530. [Google Scholar] [CrossRef]
- Smith, H.G.; Barker, H. The Luteoviridae; CABI: Wallingford, UK, 1999. [Google Scholar]
- Rouze-Jouan, J.; Terradot, L.; Pasquer, F.; Tanguy, S.; Ducray-Bourdin, D.G. The passage of Potato leafroll virus through Myzus persicae gut membrane regulates transmission efficiency. J. Gen. Virol. 2001, 82, 17–23. [Google Scholar]
- Kennedy, J.S.; DAY, M.F.; Eastop, V.F. A Conspectus of Aphids as Vectors of Plant Viruses; Cambridge University Press: Cambridge, UK, 1962. [Google Scholar]
- Weidemann, H. On the multiplication of potato leafroll virus in the aphid Myzus persicae (Sulz.). Z. Angew. Entomol. 1982, 94, 321–330. [Google Scholar] [CrossRef]
- Tamada, T.; Harrison, B. Quantitative studies on the uptake and retention of potato leafroll virus by aphids in laboratory and field conditions. Ann. Appl. Biol. 1981, 98, 261–276. [Google Scholar] [CrossRef]
- Gildow, F.; Rochow, W. Importance of capsid integrity for interference between two isolates of barley yellow dwarf virus in an aphid. Phytopathology 1980, 70, 1013–1015. [Google Scholar] [CrossRef]
- Rochow, W. Barley yellow dwarf virus: phenotypic mixing and vector specificity. Science 1970, 167, 875–878. [Google Scholar]
- Chay, C.; Gunasinge, U.; Dinesh-Kumar, S.; Miller, W.; GRAY, S.M. Aphid transmission and systemic plant infection determinants of barley yellow dwarf luteovirus-PAV are contained in the coat protein readthrough domain and 17-kDa protein, respectively. Virology 1996, 219, 57–65. [Google Scholar] [CrossRef]
- Gildow, F.; Reavy, B.; Mayo, M.; Duncan, G.; Woodford, J.; Lamb, J.; Hay, R. Aphid acquisition and cellular transport of Potato leafroll virus-like particles lacking P5 readthrough protein. Phytopathology 2000, 90, 1153–1161. [Google Scholar] [CrossRef]
- Reinbold, C.; Gildow, F.; Herrbach, E.; Ziegler-Graff, V.; Goncalves, M.; van Den Heuvel, J.; Brault, V. Studies on the role of the minor capsid protein in transport of Beet western yellows virus through Myzus persicae. J. Gen. Virol. 2001, 82, 1995–2007. [Google Scholar]
- Jolly, C.; Mayo, M. Changes in the amino acid sequence of the coat protein readthrough domain of potato leafroll luteovirus affect the formation of an epitope and aphid transmission. Virology 1994, 201, 182–185. [Google Scholar] [CrossRef]
- Brault, V.; Mutterer, J.; Scheidecker, D.; Simonis, M.; Herrbach, E.; Richards, K.; Ziegler-Graff, V. Effects of point mutations in the readthrough domain of the beet western yellows virus minor capsid protein on virus accumulation in planta and on transmission by aphids. J. Virol. 2000, 74, 1140–1148. [Google Scholar] [CrossRef]
- Lee, L.; Kaplan, I.B.; Ripoll, D.R.; Liang, D.; Palukaitis, P.; Gray, S.M. A surface loop of the potato leafroll virus coat protein is involved in virion assembly, systemic movement, and aphid transmission. J. Virol. 2005, 79, 1207–1214. [Google Scholar] [CrossRef]
- Gildow, F. Coated-vesicle transport of luteoviruses through salivary glands of myzus persicae. Phytopathology 1982, 72, 1289–1296. [Google Scholar]
- Gildow, F.; Damsteegt, V.; Stone, A.; Smith, O.; Gray, S. Virus:Vector Cell Interactions Regulating Transmission Specificity of Soybean Dwarf Luteoviruses. J. Phytopathol. 2000, 148, 333–342. [Google Scholar]
- Rubinstein, G.; Czosnek, H. Long-term association of tomato yellow leaf curl virus with its whitefly vector Bemisia tabaci: Effect on the insect transmission capacity, longevity and fecundity. J. Gen. Virol. 1997, 78, 2683–2689. [Google Scholar]
- Czosnek, H.; Ghanim, M. Back to basics: Are begomoviruses whitefly pathogens? J. Integr. Agric. 2012, 11, 225–234. [Google Scholar] [CrossRef]
- Ghanim, M.; Czosnek, H. Tomato yellow leaf curl geminivirus (TYLCV-Is) is transmitted among whiteflies (Bemisia tabaci) in a sex-related manner. J. Virol. 2000, 74, 4738–4745. [Google Scholar] [CrossRef]
- Sinisterra, X.H.; McKenzie, C.; Hunter, W.B.; Powell, C.A.; Shatters, R.G. Differential transcriptional activity of plant-pathogenic begomoviruses in their whitefly vector (Bemisia tabaci, Gennadius: Hemiptera Aleyrodidae). J. Gen. Virol. 2005, 86, 1525–1532. [Google Scholar]
- Cohen, S.; Nitzany, F. Transmission and host range of the tomato yellow leaf curl virus. Phytopathology 1966, 56, 1127–1131. [Google Scholar]
- Cicero, J.M.; Brown, J.K. Anatomy of accessory salivary glands of the whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) and correlations to Begomovirus transmission. Ann. Entomol. Soc. Am. 2011, 104, 280–286. [Google Scholar]
- Cicero, J.M.; Brown, J.K. Functional anatomy of whitefly organs associated with squash leaf curl virus (Geminiviridae: Begomovirus) transmission by the B Biotype of Bemisia tabaci (Hemiptera: Aleyrodidae). Ann. Entomol. Soc. Am. 2011, 104, 261–279. [Google Scholar]
- De Barro, P.J.; Liu, S.-S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Brown, J.K. The Bemisia tabaci complex: Genetic and phenotypic variability drives begomovirus spread and virus diversification. APSnet Feature Story January 2007. [Google Scholar] [CrossRef]
- Brown, J.K. The Bemisia tabaci complex: Genetic and phenotypic variation and relevance to TYLCV-vector interactions. In Tomato Yellow Leaf Curl Virus Disease; Springer: Dordrecht, The Netherlands, 2007; pp. 25–56. [Google Scholar]
- Brown, J.K. Phylogenetic biology of the Bemisia tabaci sibling species group. In Bemisia: Bionomics and Management of a Global Pest; Springer: Dordrecht, The Netherlands, 2010; pp. 31–67. [Google Scholar]
- Brown, J.; Frohlich, D.; Rosell, R. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol. 1995, 40, 511–534. [Google Scholar] [CrossRef]
- Frohlich, D.; Torres-Jerez, I.; Bedford, I.; Markham, P.; Brown, J. A phylogeographical analysis of the Bemisia tabaci species complex based on mitochondrial DNA markers. Mol. Ecol. 1999, 8, 1683–1691. [Google Scholar] [CrossRef]
- Dinsdale, A.; Cook, L.; Riginos, C.; Buckley, Y.; Barro, P.D. Refined global analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha: Aleyrodoidea: Aleyrodidae) mitochondrial cytochrome oxidase 1 to identify species level genetic boundaries. Ann. Entomol. Soc. Am. 2010, 103, 196–208. [Google Scholar] [CrossRef]
- Costa, H.; Brown, J. Variation in biological characteristics and esterase patterns among populations of Bemisia tabaci, and the association of one population with silverleaf symptom induction. Entomol. Exp. Appl. 1991, 61, 211–219. [Google Scholar] [CrossRef]
- Horowitz, A.R.; Kontsedalov, S.; Khasdan, V.; Ishaaya, I. Biotypes B and Q of Bemisia tabaci and their relevance to neonicotinoid and pyriproxyfen resistance. Arch. Insect Biochem. Physiol. 2005, 58, 216–225. [Google Scholar] [CrossRef]
- Hanley-Bowdoin, L.; Settlage, S.B.; Orozco, B.M.; Nagar, S.; Robertson, D. Geminiviruses: Models for plant DNA replication, transcription, and cell cycle regulation. Crit. Rev. Plant Sci. 1999, 18, 71–106. [Google Scholar] [CrossRef]
- Cicero, J.; Hiebert, E.; Webb, S. The alimentary canal of Bemisia tabaci and Trialeurodes abutilonea (Homoptera, Sternorrhynchi): Histology, ultrastructure and correlations to function. Zoomorphology 1995, 115, 31–39. [Google Scholar] [CrossRef]
- Lucio-Zavaleta, E.; Smith, D.; Gray, S. Variation in transmission efficiency among Barley yellow dwarf virus-RMV isolates and clones of the normally inefficient aphid vector, Rhopalosiphum padi. Phytopathology 2001, 91, 792–796. [Google Scholar] [CrossRef]
- Morin, S.; Ghanim, M.; Sobol, I.; Czosnek, H. The GroEL protein of the whitefly Bemisia tabaci interacts with the coat protein of transmissible and nontransmissible begomoviruses in the yeast two-hybrid system. Virology 2000, 276, 404–416. [Google Scholar] [CrossRef]
- Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Kontsedalov, S.; Skaljac, M.; Brumin, M.; Sobol, I.; Czosnek, H.; Vavre, F.; Fleury, F.; et al. The transmission efficiency of tomato yellow leaf curl virus by the whitefly bemisia tabaci is correlated with the presence of a specific symbiotic bacterium species. J. Virol. 2010, 84, 9310–9317. [Google Scholar] [CrossRef]
- Gotz, M.; Popovski, S.; Kollenberg, M.; Gorovitz, R.; Brown, J.K.; Cicero, J.M.; Czosnek, H.; Winter, S.; Ghanim, M. Implication of Bemisia tabaci heat shock protein 70 in begomovirus—Whitefly interactions. J. Virol. 2012, 86, 13241–13252. [Google Scholar]
- Franz, A.W.; van der Wilk, F.; Verbeek, M.; Dullemans, A.M.; van den Heuvel, J.F. Faba Bean Necrotic Yellows Virus (Genus Nanovirus) requires a helper factor for its aphid transmission. Virology 1999, 262, 210–219. [Google Scholar] [CrossRef]
- Gildow, F. Evidence for receptor-mediated endocytosis regulating luteovirus acquisition by aphids. Phytopathology 1993, 83, 270–277. [Google Scholar] [CrossRef]
- Gildow, F.; Gray, S. The aphid salivary gland basal lamina as a selective barrier associated with vector-specific transmission of barley yellow dwarf luteoviruses. Phytopathology 1993, 83, 1293–1302. [Google Scholar] [CrossRef]
- Peiffer, M.; Gildow, F.; Gray, S. Two distinct mechanisms regulate luteovirus transmission efficiency and specificity at the aphid salivary gland. J. Gen. Virol. 1997, 78, 495–503. [Google Scholar]
- Gildow, F.; Rochow, W. Transmission interference between two isolates of barley yellow dwarf virus in Macrosiphum avenae. Phytopathology 1980, 70, 122–126. [Google Scholar] [CrossRef]
- Seddas, P.; Boissinot, S.; Strub, J.-M.; van Dorsselaer, A.; van Regenmortel, M.H.; Pattus, F. Rack-1, GAPDH3, and actin: Proteins of Myzus persicae potentially involved in the transcytosis of beet western yellows virus particles in the aphid. Virology 2004, 325, 399–412. [Google Scholar] [CrossRef]
- Li, C.; Cox-Foster, D.; Gray, S.M.; Gildow, F. Vector specificity of Barley Yellow Dwarf Virus (BYDV) transmission: Identification of potential cellular receptors binding BYDV-MAV in the aphid, Sitobion avenae. Virology 2001, 286, 125–133. [Google Scholar] [CrossRef]
- Azzam, O.; Frazer, J.; de La Rosa, D.; Beaver, J.; Ahlquist, P.; Maxwell, D. Whitefly transmission and efficient ssDNA accumulation of bean golden mosaic geminivirus require functional coat protein. Virology 1994, 204, 289–296. [Google Scholar] [CrossRef]
- Briddon, R.; Pinner, M.; Stanley, J.; Markham, P. Geminivirus coat protein replacement alters insect specificity. Virology 1990, 177, 85–94. [Google Scholar] [CrossRef]
- Höfer, P.; Bedford, I.D.; Markham, P.G.; Jeske, H.; Frischmuth, T. Coat protein gene replacement results in whitefly transmission of an insect nontransmissible geminivirus isolate. Virology 1997, 236, 288–295. [Google Scholar] [CrossRef]
- Noris, E.; Vaira, A.; Caciagli, P.; Masenga, V.; Gronenborn, B.; Accotto, G. Amino acids in the capsid protein of tomato yellow leaf curl virus that are crucial for systemic infection, particle formation, and insect transmission. J. Virol. 1998, 72, 10050–10057. [Google Scholar]
- Soto, M.J.; Chen, L.-F.; Seo, Y.-S.; Gilbertson, R.L. Identification of regions of the Beet mild curly top virus (family Geminiviridae) capsid protein involved in systemic infection, virion formation and leafhopper transmission. Virology 2005, 341, 257–270. [Google Scholar] [CrossRef]
- Nuttall, P.A.; Jones, L.D.; Labuda, M.; Kaufman, W.R. Adaptations of arboviruses to ticks. J. Med. Entomol. 1994, 31, 1–9. [Google Scholar]
- Perez de Leon, A.A.; Ribeiro, J.M.; Tabachnick, W.J.; Valenzuela, J.G. Identification of a salivary vasodilator in the primary North American vector of bluetongue viruses, Culicoides variipennis. Am. J. Trop. Med. Hyg. 1997, 57, 375–381. [Google Scholar]
- Kubes, M.; Fuchsberger, N.; Labuda, M.; Zuffova, E.; Nuttall, P. Salivary gland extracts of partially fed Dermacentor reticulatus ticks decrease natural killer cell activity in vitro. Immunology 1994, 82, 113. [Google Scholar]
- Valenzuela, J.G.; Chuffe, O.M.; Ribeiro, J.C. Apyrase and anti-platelet activities from the salivary glands of the bed bug Cimex lectularius. Insect Biochem. Mol. Biol. 1996, 26, 557–562. [Google Scholar] [CrossRef]
- Mowry, T.M. Within-plant accumulation of potato leafroll virus by aggregated green peach aphid feeding. Phytopathology 1995, 85, 859–863. [Google Scholar] [CrossRef]
- Bedford, I.; Briddon, R.; Brown, J.; Rosell, R.; Markham, P. Geminivirus transmission and biological characterisation of Bemisia tabaci (Gennadius) biotypes from different geographic regions. Ann. Appl. Biol. 1994, 125, 311–325. [Google Scholar] [CrossRef]
- Rosell, R.; Brown, J. Mechanisms of geminivirus acquisition and transmission by the whitefly Bemisia tabaci (Genn.). Phytopathology 1994, 84, 1131–1132. [Google Scholar]
- Wu, Z.; Hu, J.; Polston, J.; Ullman, D.; Hiebert, E. Complete nucleotide sequence of a nonvector-transmissible strain of Abutilon mosaic geminivirus in Hawaii. Phytopathology 1996, 86, 608–613. [Google Scholar] [CrossRef]
- Brown, J.K.; Hartitz, M.D.; Rosell, R.C.; Bisaro, D.M. Subgroup III geminivirus coat protein plays a direct role in whitefly-mediated transmission. Phytopathology 1996, 86, S43. [Google Scholar]
- Hwang, D.-J.; Tumer, N.; Wilson, T. Chaperone protein GrpE and the GroEL/GroES complex promote the correct folding of tobacco mosaic virus coat protein for ribonucleocapsid assembly in vivo. Arch. Virol. 1998, 143, 2203–2214. [Google Scholar] [CrossRef]
- Thirumalai, D.; Lorimer, G.H. Chaperonin-mediated protein folding. Annu. Rev. Biophys. Biomol. Struct. 2001, 30, 245–269. [Google Scholar] [CrossRef]
- Gething, M.-J.; Sambrook, J. Protein folding in the cell. Nature 1992, 335, 33–45. [Google Scholar]
- De Beus, M.D.; Doyle, S.M.; Teschke, C.M. GroEL binds a late folding intermediate of phage P22 coat protein. Cell Stress Chaperones 2000, 5, 163. [Google Scholar] [CrossRef]
- Xu, Z.; Horwich, A.L.; Sigler, P.B. The crystal structure of the asymmetric GroEL–GroES–(ADP) 7 chaperonin complex. Nature 1997, 388, 741–750. [Google Scholar] [CrossRef]
- Fisher, M. GroE chaperonin-assisted folding and assembly of dodecameric glutamine synthetase. Biochemistry 1998, 63, 382. [Google Scholar]
- Akad, F.; Dotan, N.; Czosnek, H. Trapping of Tomato yellow leaf curl virus (TYLCV) and other plant viruses with a GroEL homologue from the whitefly Bemisia tabaci. Arch. Virol. 2004, 149, 1481–1497. [Google Scholar]
- Van den Heuvel, J.; Verbeek, M.; van der Wilk, F. Endosymbiotic bacteria associated with circulative transmission of potato leafroll virus by Myzus persicae. J. Gen. Virol. 1994, 75, 2559–2565. [Google Scholar] [CrossRef]
- Ishikawa, H. Host-symbiont interactions in the protein synthesis in the pea aphid, Acyrthosiphon pisum. Insect Biochem. 1982, 12, 613–622. [Google Scholar]
- Hogenhout, S.A.; van Der Wilk, F.; Verbeek, M.; Goldbach, R.W.; van den Heuvel, J.F.J.M. Potato leafroll virus binds to the equatorial domain of the aphid endosymbiotic GroEL homolog. J. Virol. 1998, 72, 358–365. [Google Scholar]
- Van den Heuvel, J.; Bruyere, A.; Hogenhout, S.; Ziegler-Graff, V.; Brault, V.; Verbeek, M.; van Der Wilk, F.; Richards, K. The N-terminal region of the luteovirus readthrough domain determines virus binding to Buchnera GroEL and is essential for virus persistence in the aphid. J. Virol. 1997, 71, 7258–7265. [Google Scholar]
- Chiel, E.; Gottlieb, Y.; Zchori-Fein, E.; Mozes-Daube, N.; Katzir, N.; Inbar, M.; Ghanim, M. Biotype-dependent secondary symbiont communities in sympatric populations of Bemisia tabaci. Bull. Entomol. Res. 2007, 97, 407–413. [Google Scholar] [CrossRef]
- Škaljac, M.; Žanić, K.; Hrnčić, S.; Radonjić, S.; Perović, T.; Ghanim, M. Diversity and localization of bacterial symbionts in three whitefly species (Hemiptera: Aleyrodidae) from the east coast of the Adriatic Sea. Bull. Entomol. Res. 2013, 103, 48–59. [Google Scholar] [CrossRef]
- Skaljac, M.; Zanic, K.; Ban, S.G.; Kontsedalov, S.; Ghanim, M. Co-infection and localization of secondary symbionts in two whitefly species. BMC Microbiol. 2010, 10, 142. [Google Scholar] [CrossRef]
- Bing, X.-L.; Yang, J.; Zchori-Fein, E.; Wang, X.-W.; Liu, S.-S. Characterization of a newly discovered symbiont of the Whitefly Bemisia tabaci (Hemiptera: Aleyrodidae). Appl. Environ. Microbiol. 2013, 79, 569–575. [Google Scholar] [CrossRef]
- Morin, S.; Ghanim, M.; Zeidan, M.; Czosnek, H.; Verbeek, M.; van den Heuvel, J.F.J.M. A GroEL homologue from endosymbiotic bacteria of the Whitefly Bemisia tabaci is implicated in the circulative transmission of Tomato Yellow Leaf Curl Virus. Virology 1999, 256, 75–84. [Google Scholar] [CrossRef]
- Jiu, M.; Zhou, X.-P.; Tong, L.; Xu, J.; Yang, X.; Wan, F.-H.; Liu, S.-S. Vector-virus mutualism accelerates population increase of an invasive whitefly. PLoS One 2007, 2, e182. [Google Scholar] [CrossRef]
- Kontsedalov, S.; Abu-Moch, F.; Lebedev, G.; Czosnek, H.; Horowitz, A.R.; Ghanim, M. Bemisia tabaci biotype dynamics and resistance to insecticides in Israel during the years 2008–2010. J. Intg. Agr. 2012, 11, 312–320. [Google Scholar]
- Ghanim, M.; Rosell, R.C.; Campbell, L.R.; Czosnek, H.; Brown, J.K.; Ullman, D.E. Digestive, salivary, and reproductive organs of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) B type. J. Morphol. 2001, 248, 22–40. [Google Scholar] [CrossRef]
- Luan, J.-B.; Li, J.-M.; Varela, N.L.; Wang, Y.-L.; Li, F.-F.; Bao, Y.-Y.; Zhang, C.-X.; Liu, S.-S.; Wang, X.-W. Global analysis of the transcriptional response of whitefly to tomato yellow leaf curl China virus reveals the relationship of coevolved adaptations. J. Virol. 2011, 85, 3330–3340. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kliot, A.; Ghanim, M. The Role of Bacterial Chaperones in the Circulative Transmission of Plant Viruses by Insect Vectors. Viruses 2013, 5, 1516-1535. https://doi.org/10.3390/v5061516
Kliot A, Ghanim M. The Role of Bacterial Chaperones in the Circulative Transmission of Plant Viruses by Insect Vectors. Viruses. 2013; 5(6):1516-1535. https://doi.org/10.3390/v5061516
Chicago/Turabian StyleKliot, Adi, and Murad Ghanim. 2013. "The Role of Bacterial Chaperones in the Circulative Transmission of Plant Viruses by Insect Vectors" Viruses 5, no. 6: 1516-1535. https://doi.org/10.3390/v5061516