Macrophages and Cell-Cell Spread of HIV-1

Virus-Cell Interaction Section, HIV Drug Resistance Program, National Cancer Institute, Frederick, MD 21702, USA
*
Author to whom correspondence should be addressed.
Viruses 2010, 2(8), 1603-1620; https://doi.org/10.3390/v2081603
Submission received: 14 June 2010
/
Revised: 22 July 2010
/
Accepted: 27 July 2010
/
Published: 5 August 2010
(This article belongs to the Special Issue Transmission of Retroviruses across Virological Synapses)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Macrophages have been postulated to play an important role in the pathogenesis of HIV-1 infection. Their ability to cross the blood-brain barrier and their resistance to virus-induced cytopathic effects allows them to serve as reservoirs for long-term infection. Thus, exploring the mechanisms of virus transmission from macrophages to target cells such as other macrophages or T lymphocytes is central to our understanding of HIV-1 pathogenesis and progression to AIDS, and is vital to the development of vaccines and novel antiretroviral therapies. This review provides an overview of the current understanding of cell-cell transmission in macrophages.
1. Introduction
Infection by HIV-1, a member of the genus Lentiviridae, results in an acute period characterized by high viral loads and a transient drop in CD4+ T cell counts. Following a vigorous adaptive immune response, viral loads decline and CD4+ T-cell counts recover. Over time, however, CD4 counts decline progressively, eventually leading to immunodeficiency and death [1]. HIV-1 enters target cells in most cases by using CD4 (a major attachment receptor), and chemokine coreceptors (CCR5 or CXCR4) [2]. CD4+ T cells represent a major target cell population in vivo. However, other cell populations, notably macrophages, are also productively infected by HIV-1. Unlike T cells, macrophages survive for long periods of time post-infection [3,4] and their ability to cross the blood-brain barrier suggests an important role in disseminating virus to the central nervous system [5].
3. HIV-1 replication
As mentioned above, HIV-1 targets immune cells – primarily T cells, macrophages, and dendritic cells – that express CD4 and chemokine coreceptors CCR5 or CXCR4. The levels of coreceptor expression differ between T-cell lines, primary CD4+ T cells, and macrophages. Most T-cell lines express high levels of CXCR4 but are CCR5-negative. In contrast, CCR5 is expressed abundantly on macrophages. Primary CD4+ T-cells generally express both CXCR4 and CCR5. Thus, CXCR4 (X4)-tropic strains of HIV-1 can infect T-cell lines, primary CD4+ T cells, but only to a limited extent macrophages; CCR5 (R5)-tropic strains infect monocyte-derived macrophages (MDMs) and primary CD4+ T cells; and dual-tropic (R5/X4) strains can infect all of these cell types. During the course of HIV-1 infection in vivo, a shift in coreceptor usage from R5 to R5/X4 and in some cases X4 often occurs [13]. This shift towards increased CXCR4 usage is typically accompanied by a decline in CD4+ cell numbers. It has been suggested that X4-tropic strains are more readily neutralized than R5-tropic isolates; the former thus emerge near the end of the disease course when the immune system has been largely incapacitated [14].
Because virus isolates present early in infection are typically R5-tropic, it has long been assumed that macrophages serve as an early target cell during person-to-person transmission. However, a recent study that used single genome sequencing to isolate and characterize transmitted (“founder”) viruses observed that these HIV-1 strains replicated poorly in macrophages despite being R5-tropic [15]. This finding, which extends earlier reports that CD4+ memory T-cells are the primary target for primate lentiviral infection early post-transmission [16], raises questions about the importance of macrophages in primary infection.
The HIV-1 replication cycle begins with the binding of the viral envelope (Env) glycoprotein complex – composed of the surface Env glycoprotein gp120 and the transmembrane Env glycoprotein gp41 – to CD4 and coreceptor on the target cell. These interactions induce conformational changes in gp120 and gp41 that lead to the exposure of the fusion peptide at the N-terminus of gp41. Fusion peptide insertion into the target membrane triggers the fusion of viral and host lipid bilayers and subsequent release of the viral core into the target cell cytoplasm [17]. After gaining access to the cytosol, the viral RNA genome is reverse transcribed to double-stranded DNA, which is translocated to the nucleus and integrated into the host cell genome. In vitro studies have shown that macrophages are capable of maintaining unintegrated viral DNAs for several weeks post-infection [18].
Once the HIV-1 genome has integrated into the host cell chromosome, it serves as the template for the synthesis of a number of viral RNAs, which are exported from the nucleus to the cytoplasm for translation and packaging. The Env glycoprotein precursor, gp160, is synthesized in the endoplasmic reticulum (ER) and transported to the plasma membrane via the secretory pathway. In contrast, the Gag and GagPol polyprotein precursors are synthesized on free ribosomes in the cytosol and are targeted to the plasma membrane by a poorly understood mechanism. Assembly generally takes place at the plasma membrane [19,20], and nascent particles bud off from the cell surface. So-called “late” domains near the C-terminus of Gag recruit components of the ESCRT (for endosomal sorting complex required for transport) machinery to promote the pinching-off of virus particles from the plasma membrane. This ESCRT machinery normally functions in membrane budding and fission reactions in late endosomes and during the abscission step of cytokinesis [21,22,23,24]. During or shortly after virus release, the viral protease (PR) cleaves the Gag and GagPol precursors, leading to virion maturation - a structural and morphological rearrangement of the virion that is essential for infectivity. With the completion of maturation, the mature virions can undergo a new round of infection.
4. HIV-1 Gag trafficking in macrophages
Although it is well established that the assembly and release of HIV-1 virions take place at the plasma membrane in T cells, the site of assembly in macrophages has been the subject of controversy [25]. Early electron microscopy (EM) data demonstrated that virus assembly in monocyte-derived macrophages in culture takes place in an internal, vesicle-like compartment [26]. This compartment was later shown to bear tetraspanin markers, such as CD63 and CD81, that are characteristic components of late endosomes or multivesicular bodies (MVBs) [27,18,29]. The prevailing hypothesis at that time was that in infected macrophages HIV-1 assembles in MVBs and is released from the cell upon fusion of the virus-containing MVBs with the plasma membrane – a pathway used to export exosomes from antigen-presenting cells [27,18,29]. More recently, Deneka et al. and Welsch et al. observed that this putative internal virus-containing compartment in macrophages is connected to the exterior of the cell by a thin channel, and is thus continuous with – and part of – the plasma membrane rather than being a late endosome or MVB [30,31]. Bennett and colleagues used a 3D imaging technique known as ion abrasion scanning EM to visualize the internal virus-containing compartment in MDMs [32]. Interestingly, many of the narrow channels connecting the internal virus-containing compartment with the cell surface contained numerous virions lined up “single file” [32] (Figure 1). An additional viewpoint was offered by Benaroch and colleagues, who observed HIV-1 assembly in MDMs in an intracellularly closed compartment that failed to acidify [33]. In their study, they observed that only 20% of the virus-containing compartments could be stained by a membrane-impermeable dye. Collectively, these studies suggest that the apparently intracellular compartment in which HIV-1 assembles in MDMs is distinct from conventional endosomes/MVBs. Consistent with this hypothesis, Gousset and colleagues showed that virions formed by a Gag mutant that assembles in MVBs failed to relocate to sites of cell-cell contact, in contrast to WT virions, which efficiently translocated to the synapse [34].
Numerous studies have observed colocalization of Gag and tetraspanin markers within infected MDMs and at macrophage-macrophage and macrophage-T cell contacts; however, what role these molecules play in HIV-1 replication remains an interesting and unresolved topic of research. Ruiz-Mateos et al. showed that knocking down CD63 expression in MDMs with siRNA affected neither particle production nor infectivity [35]. However, overexpression of tetraspanins or treatment of virus-producing cells with anti-tetraspanin antibodies was shown to disrupt particle production, reduce the infectivity of HIV-1 virions [36,37,38], and suppress virus-induced syncytium formation [39]. Furthermore, depletion of CD81 was reported to enhance virus transmission to target cells [37]. Opposing data on the role of endogenous CD63 were presented by Chen and colleagues who reported that CD63 depletion inhibited HIV-1 replication in MDMs [40]. Overall, these data suggest that tetraspanins play regulatory role(s) in particle infectivity and cell-cell virus transmission.
The Gag polyprotein precursor, Pr55Gag – often referred to simply as “Gag” – is the viral protein predominantly responsible for driving HIV-1 assembly [19,20]. No other viral proteins are required for the generation of immature VLPs. Pr55Gag is composed of four major domains arranged from N- to C-terminus: matrix (MA), capsid (CA), nucleocapsid (NC), and p6. Concomitant with virus budding and release, these domains are liberated from Pr55Gag by PR-mediated cleavage. Each domain of Gag carries out distinct functions during the assembly process. MA bears several determinants – a covalently attached myristic acid and a highly basic patch of amino acid residues – that are critical for directing the membrane association of Gag. The basic patch also regulates the localization of virus assembly, as mutations in this region of MA induce a retargeting of virus assembly to MVBs [27]. CA promotes Gag multimerization during assembly and also forms the outer shell of the viral core during virion maturation. NC, by virtue of its RNA binding capability, also directs Gag-Gag multimerization and is responsible for packaging the viral RNA genome into virions. The p6 domain contains binding sites for the ESCRT machinery that, as mentioned above, is responsible for particle budding and release. Specifically, p6 binds the ESCRT-I component Tsg101 and the ESCRT-associated factor Alix [21,22,24,41].
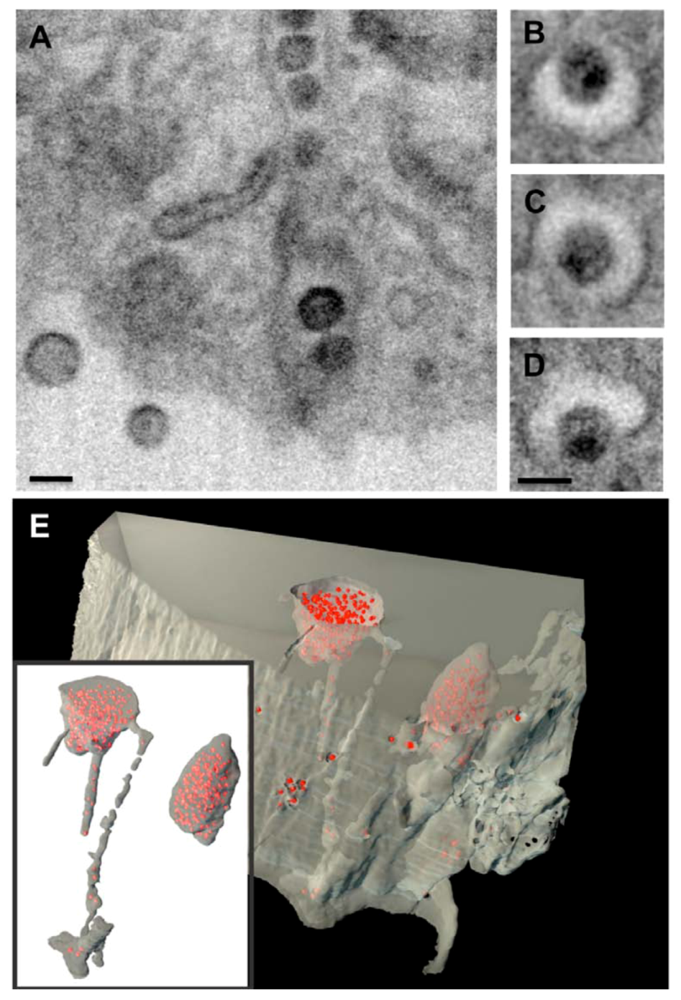
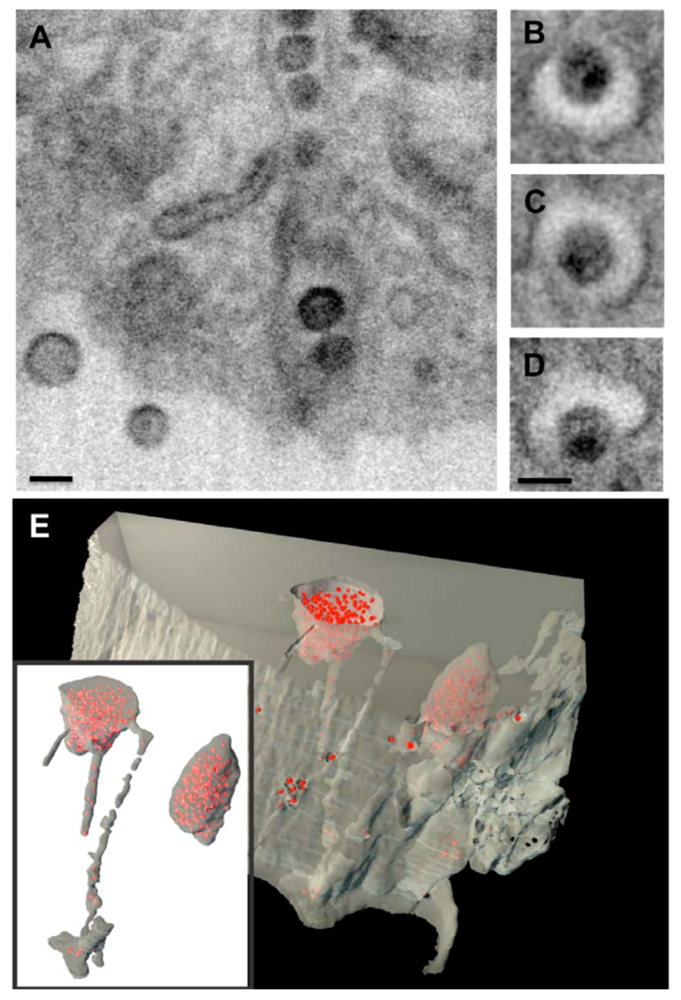
Figure 1.
Assembly of HIV-1 in an internal compartment in primary monocyte-derived macrophages (MDMs).Ion-abrasion scanning EM shows the internal virus-containing compartments in MDMs and the virion channels connecting these compartments with the cell surface. (A) Transverse and (B-D) axial sections are shown, with scale bars = 100nm. (E) Reconstruction of a 3D image from IA-SEM, showing internal HIV-1 particles (red) and virion channels. Reprinted from Bennett et al. (2009) [32].
Figure 1.
Assembly of HIV-1 in an internal compartment in primary monocyte-derived macrophages (MDMs).Ion-abrasion scanning EM shows the internal virus-containing compartments in MDMs and the virion channels connecting these compartments with the cell surface. (A) Transverse and (B-D) axial sections are shown, with scale bars = 100nm. (E) Reconstruction of a 3D image from IA-SEM, showing internal HIV-1 particles (red) and virion channels. Reprinted from Bennett et al. (2009) [32].
As is the case with other steps in the HIV-1 replication cycle, Gag trafficking to the site of assembly requires the participation of host cell factors. In cells in which HIV-1 assembles at and buds from the plasma membrane, Gag is recruited to cholesterol/sphingolipid-enriched lipid raft domains at the plasma membrane [42]. This is reflected by raft association of Pr55Gag[43,44,45] and also by the raft-like composition of the viral lipid bilayer [46,47,48]. Lipid rafts likely serve as a platform for Gag assembly and for incorporation of the Env glycoprotein complex into virions [49,50,51]. The lipid phosphoinositide, phosphatidylinositol-4,5-bisphosphate [PI(4,5)P2], also promotes the targeting of Gag to the inner leaflet of the plasma membrane. Depletion of PI(4,5)P2 redirects HIV-1 assembly to MVBs [52], and HIV-1 MA binds directly to PI(4,5)P2 [53,54,55]. HIV-1 MA shows specificity for PI(4,5)P2 and its interaction with this lipid triggers increased exposure of the covalently attached myristic acid moiety [54], thus stabilizing Gag interaction with the membrane.
As mentioned above, mutation of basic residues in MA or PI(4,5)P2 depletion causes HIV-1 assembly to be redirected to MVBs. It remains to be determined in these cases what signals in Gag are responsible for MVB targeting. One might reasonably assume that late domain motifs in p6, which are known to interact with ESCRT machinery, would play a major role. However, deletion of p6 does not affect the MVB localization of MA basic domain mutants [27]. While it has been suggested that HIV-1 Gag relocalizes ESCRT machinery away from the MVB to the plasma membrane [56], Welsch et al. showed that HIV-1 infection did not significantly affect the localization pattern of ESCRT and ESCRT-associated proteins [57]. This study suggests that ESCRT components are sufficiently abundant both at the plasma membrane and in MVBs to promote HIV-1 budding.
In addition to lipid rafts and PI(4,5)P2 , a number of cellular proteins have been implicated in Gag trafficking. These include the clathrin adaptor complexes AP-1, AP-2, AP-3, [58,59,60,61,62], the Golgi-localized γ-ear containing, Arf-binding (GGA) proteins, the ADP ribosylation factors (Arfs) [63], SOCS1 [64], the kinesin KIF4 [65,66], the ubiquitin E3-ligase POSH [67] – which binds the ESCRT-associated protein Alix – and Annexin 2 [68]. The mechanisms by which these diverse factors influence Gag targeting remain to be elucidated. Proteomics analysis has revealed the presence of a large number of host proteins in HIV-1 particles produced from infected macrophages [69]; in most cases, the roles of these proteins in virus replication remains to be determined.
5. Virus spread at points of cell-cell contact: the immunological and virological synapse.
It is now well established that HIV-1 and other retroviruses transfer from infected to uninfected cells at cell-cell contact sites that resemble immunological synapses (ISs) [73,74]. These have been termed virological synapses (VSs) [75,76,77] (Figure 2). ISs form between antigen-presenting cells (APCs) – such as macrophages and dendritic cells – and either CD4+ or CD8+ T cells. At the IS, antigenic peptide-loaded major histocompatibility complex (pMHC) on the APC and its cognate T-cell receptor (TCR)/CD3 complex on the T cell are recruited to the cell contact site. The pMHC-TCR interaction activates the T cell to induce its subsequent differentiation to become an effector cell. Additional ligand/receptor interactions stabilize the IS for prolonged periods of time to enable completion of the signaling event. The mediators of these interactions include intercellular adhesion molecule-1 (ICAM-1) on the APC, which binds to lymphocyte function-associated adhesion molecule (LFA-1) on the T cell; and CD28 on the T cell, which binds to CD80/CD86 on the APC. The recruitment of these molecules and the stability of the IS involves the actin/microtubule cytoskeletal network and lipid rafts [78,79,80]. Additional details on the IS and VS are reviewed in two recent papers [81,82].
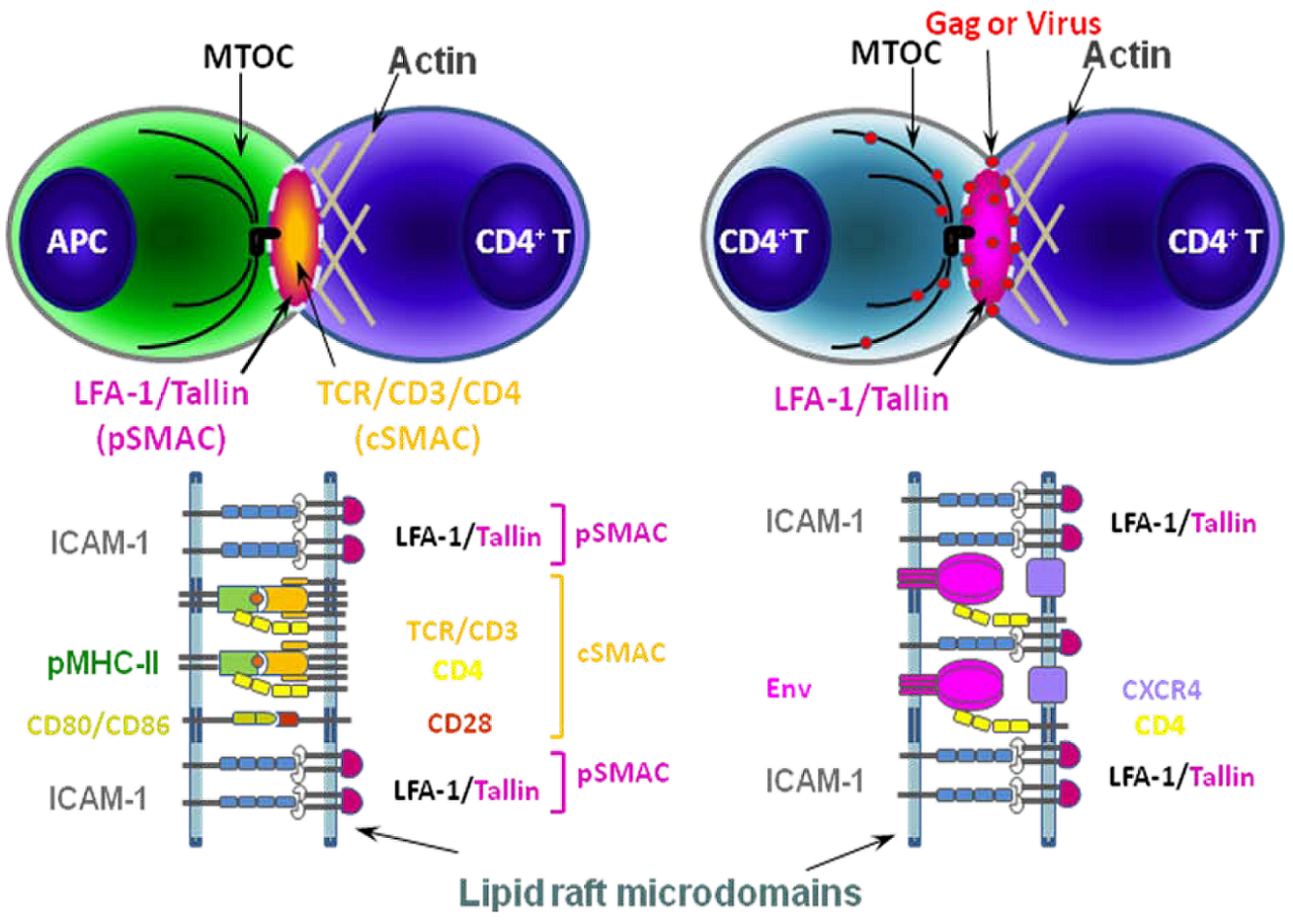
Figure 2.
Schematic models of immunological synapse (IS) between APC and CD4+ T cell (left) and virological synapse (VS) between CD4+ T cells (right).The formation of an IS between APCs and CD4+ T cells begins with the recognition of antigenic peptide loaded on MHC-II (pMHC-II) on the APC by its cognate TCR/CD3/CD4 receptors on the CD4+ T cell. The adhesion molecules (ICAM-1 and LFA-1) and co-stimulatory molecules (CD80/86 and CD28) are also recruited to lipid raft microdomains (dotted line) at the IS to form the supramolecular activation complex (SMAC) [83]. These additional ligand-receptor interactions stabilize the complex. HIV-1-infected cells form a structurally similar contact, the VS, with uninfected cells. pSMAC, peripheral SMAC; cSMAC, central SMAC. Modified with permission from Piguet and Sattentau, 2004 [77].
Figure 2.
Schematic models of immunological synapse (IS) between APC and CD4+ T cell (left) and virological synapse (VS) between CD4+ T cells (right).The formation of an IS between APCs and CD4+ T cells begins with the recognition of antigenic peptide loaded on MHC-II (pMHC-II) on the APC by its cognate TCR/CD3/CD4 receptors on the CD4+ T cell. The adhesion molecules (ICAM-1 and LFA-1) and co-stimulatory molecules (CD80/86 and CD28) are also recruited to lipid raft microdomains (dotted line) at the IS to form the supramolecular activation complex (SMAC) [83]. These additional ligand-receptor interactions stabilize the complex. HIV-1-infected cells form a structurally similar contact, the VS, with uninfected cells. pSMAC, peripheral SMAC; cSMAC, central SMAC. Modified with permission from Piguet and Sattentau, 2004 [77].
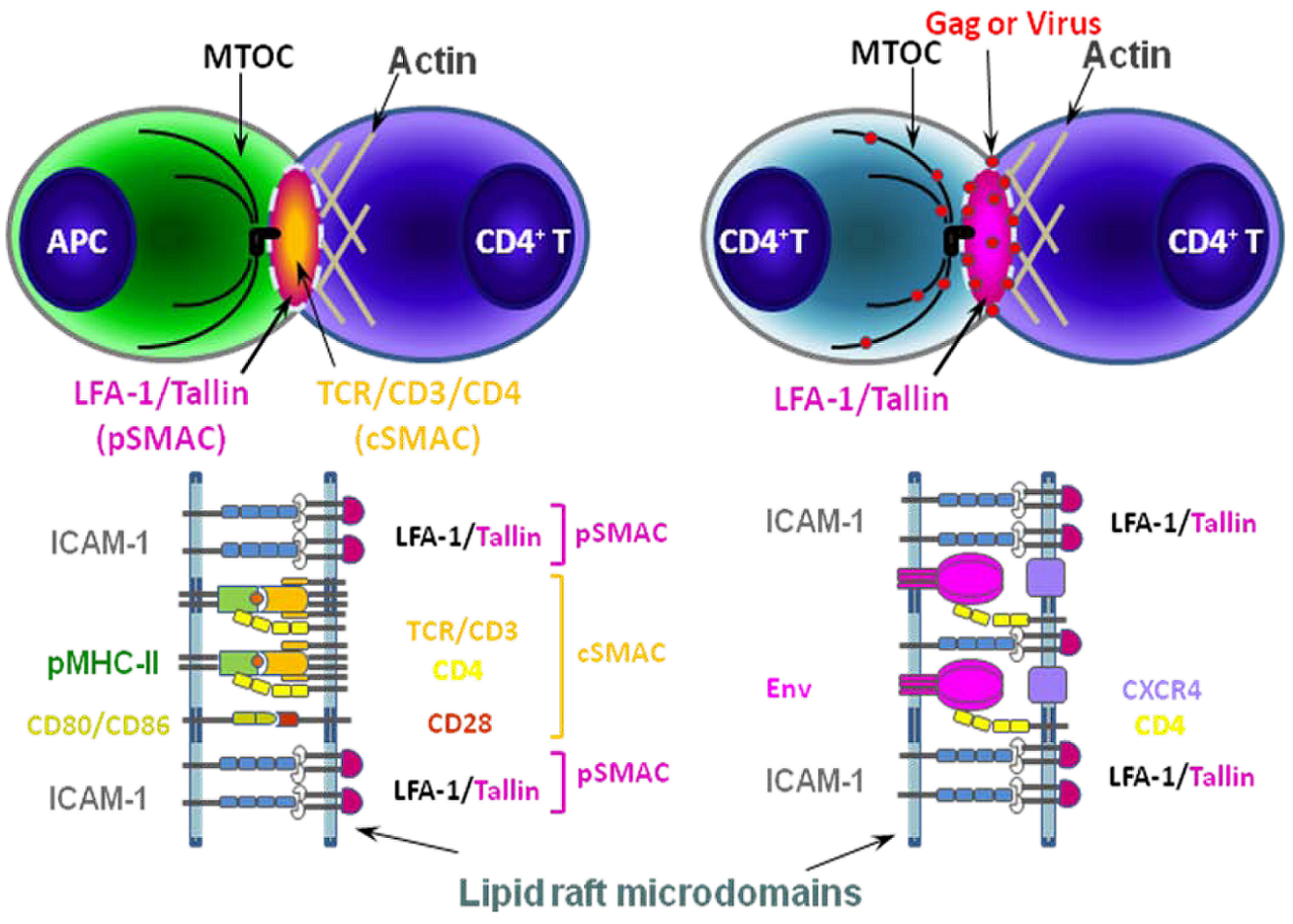
The VS is formed between cells to transfer infectious viruses either newly assembled in the donor cell (cis-infection) or captured by the donor cell from the extracellular environment (trans-infection). The first HIV-1 cell-associated transfer across a synapse was demonstrated in the context of trans-infection from DCs to T lymphocytes [76], where viruses and their receptors were recruited to the contact sites. During the differentiation process, DCs appear to participate in different modes of transfer due to their differential susceptibility to HIV-1 infection [84]; immature DCs are relatively efficiently infected and can therefore transfer viruses by the cis-infection route. In contrast, direct infection of mature DCs is less efficient; these cells thus transfer virus primarily in trans [85]. Although cis- and trans-infection modes are different in terms of the origins of the virus being transferred, they share common features: 1) virus accumulates in tetraspanin-enriched compartments [62]; 2) HIV-1-harboring compartments are accessible to the cell surface [62,86]; and 3) the virus in the internal compartment is relocated to the VS upon contact with target cell [62]. Although the mechanism of virus recruitment to the VS in the cis-infection mode remains poorly studied, in trans-infection the recruitment of the following molecules has been observed: tetraspanins (CD81 and CD9) [85], DC-SIGN (dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin), ICAM-1, and LFA-1 [87,88]. How the cytoskeletal network functions in DC-mediated trans-infection needs to be elucidated. However, recent studies indicate the involvement of the cytoskeletal network at multiple steps: the formation of the invaginated plasma-membrane-derived compartment is actin-dependent, but microtubule-independent [86]; actin-dependent macropinocytosis plays a role in the putative uptake of HIV-1 [89]; and actin and microtubules are involved in the formation of the synapse itself [89].
VS formation between T cells was first recognized in human T-cell leukemia virus type 1 (HTLV-1) infections [75]. Similar VS structures were also found in HIV-1-infected T cells [90], where viral molecules (Gag and Env), receptors, adhesion molecules, and actin were recruited to the VS. Cholesterol/GM1-enriched lipid rafts were shown to be required for VS formation and for the recruitment of Gag and Env [91]. Different cytoskeletal networks are involved on each side of the VS: F-actin on the target cells and both actin and tubulin on the infected cells [92]. Virus transfer and uptake seem to be carried out by an endocytic mechanism [93,94], probably in a clathrin- and dynamin-dependent manner [93]. Transfer can take place simultaneously from one infected cell to several uninfected cells at structures referred to as polysynapses [95]. A recent study observed that the VS is loosely structured and viral transfer across the VS can be inhibited by neutralizing antibodies [96]. To some extent, these observations challenge the view that the VS is a privileged microenvironment that is sequestered from the host humoral immune response.
Additional cell-associated viral transfer modes have been observed. These include the use of filopodial bridges, cytonemes, and tunneling nanotubes (TNT). These structures are actin-based, thread-like extensions of the plasma membrane, which can reach relatively distant target cells and require less contact area compared to synapses. Filopodial bridges have been shown to be utilized for the transfer of the retrovirus murine leukemia virus (MLV) in fibroblasts [97]. The MLV particles were observed to “surf” along the bridges, the formation of which required Env-receptor interactions. HIV-1 transfer was also found to move across filopodial bridges towards CD4/CXCR4-expressing cells [97].
TNTs, which allow the exchange of relatively large cytoplasmic components - including vesicles - between connected cells, are a well-established pathway for direct cell-to-cell communication [65]. HIV-1 has been reported to use TNTs for transfer between T cells and between macrophages [98,99]. Macrophages form TNTs of heterogeneous length, and increase in number during active viral replication [98]. Infected macrophages were observed to simultaneously form different types of TNTs in addition to filopodia [95].
6. Transfer of HIV-1 across the macrophage VS
Compared to the T-cell and DC VS, relatively little is known about the formation of, and transfer of virus across, the VS in macrophages. Initial studies, however, suggest that much of what has been learned about the VS in T cells and DCs will apply to macrophages. Two recent studies showed that macrophages could form VSs to transfer HIV-1 to uninfected macrophages and T cells, with features similar to those seen in T cells and DCs [34,100] (Figure 3). Gousset and colleagues used the biarsenical probe FlAsH in living cells to study the localization and movement of HIV-1 Gag bearing a tetra-Cys tag – the binding site for FlAsH. The tetra-Cys tag, which was introduced near the C-terminus of the MA domain in Gag, was shown to not interfere with HIV-1 assembly and release or particle infectivity. In the infected MDMs, virus particles accumulated both at the plasma membrane and in internal compartments containing the tetraspanin markers CD81 and CD82 [34]. There appeared to be little movement of Gag between these two compartments during the time course of the live-cell imaging (~90 min). However, upon coculture with T cells and subsequent VS formation, the internal virus moved rapidly (within 5-10 min) to the VS. In this system, Env was found not to be required for either VS formation or movement of Gag to the synapse [34], though in another study Env interactions with receptor and coreceptor were shown to be required for transfer of Gag across the synapse [100]. Interestingly, as mentioned above, a MA mutant that targets to MVBs also demonstrated localization to an internal tetraspanin-enriched compartment but did not translocate to the synapse [34]. This observation suggests that the WT and the mutant Gag assemble in different compartments in MDM; the most likely explanation for these findings is that the WT Gag assembles in the internal surface-connected compartment discussed above [30,31,32] rather than in true MVBs.
Groot and colleagues [100] cocultivated infected MDMs with autologous CD4+ T cells for up to six hours. Within one hour of coculture, Gag, Env, and CD4 could be observed accumulating at VSs. The VS formation appeared to be transient, with 5-10% of T cell becoming Gag-positive by six hours of coculture. The authors used a transwell system to show that whereas nearly 10% of T cells were virus-positive after 10 hours, when cell-cell contact was prevented only 0.5% of T cells became infected [100].
It remains unclear by what mechanism HIV-1 particles that assemble and accumulate in the internal virus-containing compartment traffic to the VS. This could occur by particle movement along the virion channel [32] to the cell surface or could result from the evagination of the internal compartment and wholesale “dumping” of particles at the VS. The live-cell imaging data of Gousset and colleagues [34] favor the latter hypothesis. Also remaining to be defined are the molecular signals that drive particle movement and/or evagination reactions. The recent studies discussed above [34,100] reported that T-cells often extend pseudopod or uropod-like extensions that contact the infected MDM at the VS; the signals that induce the formation of these T-cell extensions remain to be defined.
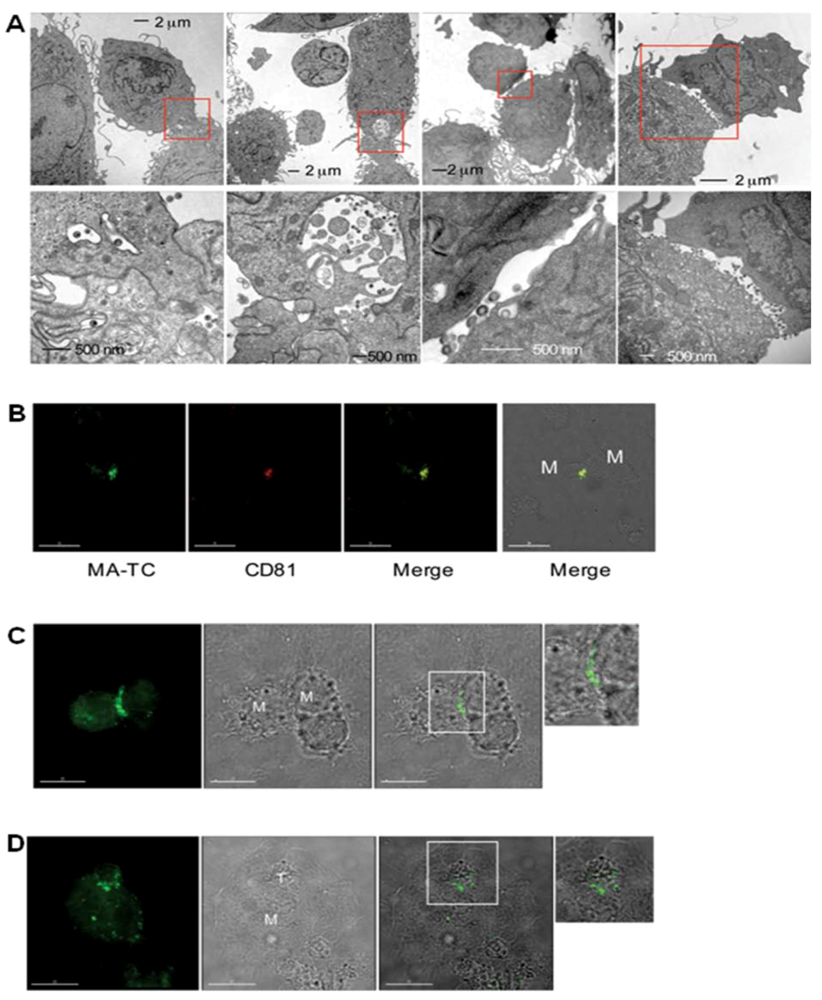
Figure 3.
Recruitment and accumulation of Gag at the MDM/MDM and MDM/T-cell synapse.Cells were infected with a tetra-Cys-tagged HIV-1 clone (A, B) or an Env(-) derivative (C, D). EM images show the accumulation of virions in intracellular compartments near the plasma membrane and the contact sites between infected MDM and uninfected MDM (A). Gag and the tetraspanin CD81 are recruited to the MDM/MDM contact site (B). Gag in infected MDMs is relocated to the contact sites with both uninfected MDMs (C) and Jurkat T cells (D) in the absence of Env expression. Scale bars, 30 μm (B) and 15 μm (C) and (D). From Gousset et al. 2008 [34].
Figure 3.
Recruitment and accumulation of Gag at the MDM/MDM and MDM/T-cell synapse.Cells were infected with a tetra-Cys-tagged HIV-1 clone (A, B) or an Env(-) derivative (C, D). EM images show the accumulation of virions in intracellular compartments near the plasma membrane and the contact sites between infected MDM and uninfected MDM (A). Gag and the tetraspanin CD81 are recruited to the MDM/MDM contact site (B). Gag in infected MDMs is relocated to the contact sites with both uninfected MDMs (C) and Jurkat T cells (D) in the absence of Env expression. Scale bars, 30 μm (B) and 15 μm (C) and (D). From Gousset et al. 2008 [34].

7. Perspectives
Macrophages have long been thought to play an important role in lentiviral biology, and it has been clear for decades that the site at which HIV-1 assembles in MDMs differs from the primarily plasma-membrane-associated assembly observed in T cells. However, many issues relating to HIV-1 infection of this cell type remain to be fully understood. For example, what precisely is the nature of the compartment in which virus assembles in MDMs? How are particles that assemble in the putative internal compartment exported to the exterior of the cell? What is the trigger for movement of internally assembled virions to the VS? What role do tetraspanins and other markers of the IS play in MDM synapse formation and movement of virus across the synapse? Also unclear is to what extent do observations made in MDM tissue culture systems apply to the in vivo biology of HIV-1. In any event, a more complete characterization of HIV-1 replication in this fascinating cell type will likely contribute to our understanding of HIV-1 biology and to the development of anti-HIV-1 vaccines and antiretroviral therapies.
Acknowledgments
We thank V. Piguet, Q. Sattentau, and S. Subramaniam for permission to reproduce figures and members of Freed laboratory for helpful discussions and critical reading of the manuscript. Research in Freed laboratory is supported by the Intramural Research Program of the NIH, National Cancer Institute, Center for Cancer Research and by the Intramural AIDS Targeted Antiviral Program.
References
- Kuritzkes, D.R.; Walker, B.D. HIV pathogenesis, clinical manifestations and treatment. In Fields Virology, 5th; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007. [Google Scholar]
- Berger, E.A.; Murphy, P.M.; Farber, J.M. Chemokine receptors as HIV-1 coreceptors: roles in viral entry, tropism, and disease. Annu. Rev. Immunol. 1999, 17, 657–700. [Google Scholar] [CrossRef] [PubMed]
- Gartner, S.; Markovits, P.; Markovitz, D.M.; Kaplan, M.H.; Gallo, R.C.; Popovic, M. The role of mononuclear phagocytes in HTLV-III/LAV infection. Science 1986, 233, 215–219. [Google Scholar] [PubMed]
- Sharova, N.; Swingler, C.; Sharkey, M.; Stevenson, M. Macrophages archive HIV-1 virions for dissemination in trans. EMBO J. 2005, 24, 2481–2489. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Scarano, F.; Martin-Garcia, J. The neuropathogenesis of AIDS. Nat. Rev. Immunol. 2005, 5, 69–81. [Google Scholar] [CrossRef] [PubMed]
- Zink, M.C.; Laast, V.A.; Helke, K.L.; Brice, A.K.; Barber, S.A.; Clements, J.E.; Mankowski, J.L. From mice to macaques--animal models of HIV nervous system disease. Curr. HIV Res. 2006, 4, 293–305. [Google Scholar] [CrossRef]
- Desrosiers, R.C. Nonhuman Lentiviruses. In Field's Virology, 5th; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams and Wilkins: Philadelphia, PA, USA, 2007. [Google Scholar]
- Bendinelli, M.; Pistello, M.; Lombardi, S.; Poli, A.; Garzelli, C.; Matteucci, D.; Ceccherini-Nelli, L.; Malvaldi, G.; Tozzini, F. Feline immunodeficiency virus: an interesting model for AIDS studies and an important cat pathogen. Clin. Microbiol. Rev. 1995, 8, 87–112. [Google Scholar] [PubMed]
- Hirsch, V.M.; Fuerst, T.R.; Sutter, G.; Carroll, M.W.; Yang, L.C.; Goldstein, S.; Piatak, M.; Elkins, W.R.; Alvord, W.G.; Montefiori, D.C.; Moss, B.; Lifson, J.D. Patterns of viral replication correlate with outcome in simian immunodeficiency virus (SIV)-infected macaques: effect of prior immunization with a trivalent SIV vaccine in modified vaccinia virus Ankara . J. Virol. 1996, 70, 3741–3752. [Google Scholar] [PubMed]
- Matsumura, S.; Ishida, T.; Washizu, T.; Tomoda, I. Histopathology and viral antigen distribution in lymph nodes of cats naturally infected with feline immunodeficiency virus. J. Vet. Med. Sci. 1994, 56, 523–528. [Google Scholar] [PubMed]
- O'Neil, S.P.; Suwyn, C.; Anderson, D.C.; Niedziela, G.; Bradley, J.; Novembre, F.J.; Herndon, J.G.; McClure, H.M. Correlation of acute humoral response with brain virus burden and survival time in pig-tailed macaques infected with the neurovirulent simian immunodeficiency virus SIVsmmFGb. Am. J. Pathol. 2004, 164, 1157–1172. [Google Scholar] [PubMed]
- Igarashi, T.; Brown, C.R.; Endo, Y.; Buckler-White, A.; Plishka, R.; Bischofberger, N.; Hirsch, V.; Martin, M.A. Macrophage are the principal reservoir and sustain high virus loads in rhesus macaques after the depletion of CD4+ T cells by a highly pathogenic simian immunodeficiency virus/HIV type 1 chimera (SHIV): Implications for HIV-1 infections of humans. Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Schuitemaker, H.; Koot, M.; Kootstra, N.A.; Dercksen, M.W.; de Goede, R.E.; van Steenwijk, R.P.; Lange, J.M.; Schattenkerk, J.K.; Miedema, F.; Tersmette, M. Biological phenotype of human immunodeficiency virus type 1 clones at different stages of infection: progression of disease is associated with a shift from monocytotropic to T-cell-tropic virus population. J. Virol. 1992, 66, 1354–1360. [Google Scholar] [PubMed]
- Bunnik, E.M.; Quakkelaar, E.D.; van Nuenen, A.C.; Boeser-Nunnink, B.; Schuitemaker, H. Increased neutralization sensitivity of recently emerged CXCR4-using human immunodeficiency virus type 1 strains compared to coexisting CCR5-using variants from the same patient. J. Virol. 2007, 81, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Gonzalez, J.F.; Salazar, M.G.; Keele, B.F.; Learn, G.H.; Giorgi, E.E.; Li, H.; Decker, J.M.; Wang, S.; Baalwa, J.; Kraus, M.H.; Parrish, N.F.; Shaw, K.S.; Guffey, M.B.; Bar, K.J.; Davis, K.L.; Ochsenbauer-Jambor, C.; Kappes, J.C.; Saag, M.S.; Cohen, M.S.; Mulenga, J.; Derdeyn, C.A.; Allen, S.; Hunter, E.; Markowitz, M.; Hraber, P.; Perelson, A.S.; Bhattacharya, T.; Haynes, B.F.; Korber, B.T.; Hahn, B.H.; Shaw, G.M. Genetic identity, biological phenotype, and evolutionary pathways of transmitted/founder viruses in acute and early HIV-1 infection. J. Exp. Med. 2009, 206, 1273–1289. [Google Scholar] [CrossRef] [PubMed]
- Haase, A.T. Perils at mucosal front lines for HIV and SIV and their hosts. Nat. Rev. Immunol. 2005, 5, 783–792. [Google Scholar] [PubMed]
- Freed, E.O.; Martin, M.A. HIVs and their replication. In Field's Virology, 5th; Knipe, D.M., Howley, P.M., Griffin, D.E., Lamb, R.A., Martin, M.A., Roizman, B., Straus, S.E., Eds.; Lippincott Williams and Wilkins: Philadelphia, 2007. [Google Scholar]
- Kelly, J.; Beddall, M.H.; Yu, D.; Iyer, S.R.; Marsh, J.W.; Wu, Y. Human macrophages support persistent transcription from unintegrated HIV-1 DNA. Virology 2008, 372, 300–312. [Google Scholar] [CrossRef] [PubMed]
- Adamson, C.S.; Freed, E.O. Human immunodeficiency virus type 1 assembly, release, and maturation. Adv. Pharmacol. 2007, 55, 347–387. [Google Scholar] [PubMed]
- Bieniasz, P.D. The cell biology of HIV-1 virion genesis. Cell Host Microbe 2009, 5, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Bieniasz, P.D. Late budding domains and host proteins in enveloped virus release. Virology 2006, 344, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Demirov, D.G.; Freed, E.O. Retrovirus budding. Virus Res. 2004, 106, 87–102. [Google Scholar] [CrossRef] [PubMed]
- Hurley, J.H. ESCRT complexes and the biogenesis of multivesicular bodies. Curr. Opin. Cell Biol. 2008, 20, 4–11. [Google Scholar] [CrossRef]
- Morita, E.; Sundquist, W.I. Retrovirus budding. Annu. Rev. Cell Dev. Biol. 2004, 20, 395–425. [Google Scholar] [CrossRef] [PubMed]
- Joshi, A.; Freed, E.O. HIV-1 Gag trafficking. Future HIV Therapy 2007, 1, 427–438. [Google Scholar] [CrossRef]
- Orenstein, J.M.; Meltzer, M.S.; Phipps, T.; Gendelman, H.E. Cytoplasmic assembly and accumulation of human immunodeficiency virus types 1 and 2 in recombinant human colony-stimulating factor-1-treated human monocytes: an ultrastructural study. J. Virol. 1988, 62, 2578–2586. [Google Scholar] [PubMed]
- Ono, A.; Freed, E.O. Cell-type-dependent targeting of human immunodeficiency virus type 1 assembly to the plasma membrane and the multivesicular body. J. Virol. 2004, 78, 1552–1563. [Google Scholar] [CrossRef] [PubMed]
- Pelchen-Matthews, A.; Kramer, B.; Marsh, M. Infectious HIV-1 assembles in late endosomes in primary macrophages. J. Cell Biol. 2003, 162, 443–455. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Moore, M.; Innes, D.; Leijendekker, R.; Leigh-Brown, A.; Benaroch, P.; Geuze, H. Human macrophages accumulate HIV-1 particles in MHC II compartments. Traffic 2002, 3, 718–729. [Google Scholar] [CrossRef] [PubMed]
- Deneka, M.; Pelchen-Matthews, A.; Byland, R.; Ruiz-Mateos, E.; Marsh, M. In macrophages, HIV-1 assembles into an intracellular plasma membrane domain containing the tetraspanins CD81, CD9, and CD53. J. Cell Biol. 2007, 177, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Keppler, O.T.; Habermann, A.; Allespach, I.; Krijnse-Locker, J.; Krausslich, H.G. HIV-1 buds predominantly at the plasma membrane of primary human macrophages . PLoS Pathog. 2007, 3, e36. [Google Scholar] [CrossRef] [PubMed]
- Bennett, A.E.; Narayan, K.; Shi, D.; Hartnell, L.M.; Gousset, K.; He, H.; Lowekamp, B.C.; Yoo, T.S.; Bliss, D.; Freed, E.O.; Subramaniam, S. Ion-abrasion scanning electron microscopy reveals surface-connected tubular conduits in HIV-infected macrophages . PLoS Pathog. 2009, 5, e1000591. [Google Scholar] [CrossRef] [PubMed]
- Jouve, M.; Sol-Foulon, N.; Watson, S.; Schwartz, O.; Benaroch, P. HIV-1 buds and accumulates in "nonacidic" endosomes of macrophages. Cell Host Microbe 2007, 2, 85–95. [Google Scholar] [CrossRef]
- Gousset, K.; Ablan, S.D.; Coren, L.V.; Ono, A.; Soheilian, F.; Nagashima, K.; Ott, D.E.; Freed, E.O. Real-time visualization of HIV-1 GAG trafficking in infected macrophages . PLoS Pathog. 2008, 4, e1000015. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Mateos, E.; Pelchen-Matthews, A.; Deneka, M.; Marsh, M. CD63 is not required for production of infectious human immunodeficiency virus type 1 in human macrophages. J. Virol. 2008, 82, 4751–4761. [Google Scholar] [CrossRef] [PubMed]
- Grigorov, B.; Attuil-Audenis, V.; Perugi, F.; Nedelec, M.; Watson, S.; Pique, C.; Darlix, J.L.; Conjeaud, H.; Muriaux, D. A role for CD81 on the late steps of HIV-1 replication in a chronically infected T cell line. Retrovirology 2009, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Krementsov, D.N.; Weng, J.; Lambele, M.; Roy, N.H.; Thali, M. Tetraspanins regulate cell-to-cell transmission of HIV-1. Retrovirology 2009, 6, 64. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Aoki, J.; Misawa, N.; Daikoku, E.; Sano, K.; Tanaka, Y.; Koyanagi, Y. Modulation of human immunodeficiency virus type 1 infectivity through incorporation of tetraspanin proteins. J. Virol. 2008, 82, 1021–1033. [Google Scholar] [CrossRef] [PubMed]
- Weng, J.; Krementsov, D.N.; Khurana, S.; Roy, N.H.; Thali, M. Formation of syncytia is repressed by tetraspanins in human immunodeficiency virus type 1-producing cells. J. Virol. 2009, 83, 7467–7474. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Dziuba, N.; Friedrich, B.; von Lindern, J.; Murray, J.L.; Rojo, D.R.; Hodge, T.W.; O'Brien, W.A.; Ferguson, M.R. A critical role for CD63 in HIV replication and infection of macrophages and cell lines. Virology 2008, 379, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Fujii, K.; Hurley, J.H.; Freed, E.O. Beyond Tsg101: the role of Alix in 'ESCRTing' HIV-1. Nat. Rev. Microbiol. 2007, 5, 912–916. [Google Scholar] [CrossRef] [PubMed]
- Waheed, A.A.; Freed, E.O. The role of lipids in retrovirus replication. Viriuses 2010, 2, 1146–1180. [Google Scholar] [CrossRef]
- Lindwasser, O.W.; Resh, M.D. Multimerization of human immunodeficiency virus type 1 Gag promotes its localization to barges, raft-like membrane microdomains. J. Virol. 2001, 75, 7913–7924. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.H.; Hildreth, J.E. Evidence for budding of human immunodeficiency virus type 1 selectively from glycolipid-enriched membrane lipid rafts. J. Virol. 2000, 74, 3264–3272. [Google Scholar] [CrossRef] [PubMed]
- Ono, A.; Freed, E.O. Plasma membrane rafts play a critical role in HIV-1 assembly and release. Proc. Natl. Acad. Sci. U. S. A. 2001, 98, 13925–13930. [Google Scholar] [CrossRef] [PubMed]
- Aloia, R.C.; Tian, H.; Jensen, F.C. Lipid composition and fluidity of the human immunodeficiency virus envelope and host cell plasma membranes. Proc. Natl. Acad. Sci. U. S. A. 1993, 90, 5181–5185. [Google Scholar] [CrossRef] [PubMed]
- Brugger, B.; Glass, B.; Haberkant, P.; Leibrecht, I.; Wieland, F.T.; Krausslich, H.G. The HIV lipidome: a raft with an unusual composition. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 2641–2646. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.; Uchil, P.D.; Jin, J.; Shui, G.; Ott, D.E.; Mothes, W.; Wenk, M.R. Retroviruses human immunodeficiency virus and murine leukemia virus are enriched in phosphoinositides. J. Virol. 2008, 82, 11228–11238. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, J.; Repik, A.; Clapham, P.R. Gag regulates association of human immunodeficiency virus type 1 envelope with detergent-resistant membranes. J. Virol. 2006, 80, 5292–5300. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.; Kim, J.O.; Ganesh, L.; Kabat, J.; Schwartz, O.; Nabel, G.J. HIV-1 assembly: viral glycoproteins segregate quantally to lipid rafts that associate individually with HIV-1 capsids and virions. Cell Host Microbe 2008, 3, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Wyma, D.J.; Kotov, A.; Aiken, C. Evidence for a stable interaction of gp41 with Pr55(Gag) in immature human immunodeficiency virus type 1 particles. J. Virol. 2000, 74, 9381–9387. [Google Scholar] [CrossRef] [PubMed]
- Ono, A.; Ablan, S.D.; Lockett, S.J.; Nagashima, K.; Freed, E.O. Phosphatidylinositol (4,5) bisphosphate regulates HIV-1 Gag targeting to the plasma membrane. Proc. Natl. Acad. Sci. U. S. A. 2004, 101, 14889–14894. [Google Scholar] [CrossRef] [PubMed]
- Chukkapalli, V.; Hogue, I.B.; Boyko, V.; Hu, W.S.; Ono, A. Interaction between the human immunodeficiency virus type 1 Gag matrix domain and phosphatidylinositol-(4,5)-bisphosphate is essential for efficient gag membrane binding. J. Virol. 2008, 82, 2405–2417. [Google Scholar] [CrossRef] [PubMed]
- Saad, J.S.; Miller, J.; Tai, J.; Kim, A.; Ghanam, R.H.; Summers, M.F. Structural basis for targeting HIV-1 Gag proteins to the plasma membrane for virus assembly. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 11364–11369. [Google Scholar] [CrossRef] [PubMed]
- Shkriabai, N.; Datta, S.A.; Zhao, Z.; Hess, S.; Rein, A.; Kvaratskhelia, M. Interactions of HIV-1 Gag with assembly cofactors. Biochemistry 2006, 45, 4077–4083. [Google Scholar] [CrossRef] [PubMed]
- Eastman, S.W.; Martin-Serrano, J.; Chung, W.; Zang, T.; Bieniasz, P.D. Identification of human VPS37C, a component of endosomal sorting complex required for transport-I important for viral budding. J. Biol. Chem. 2005, 280, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Welsch, S.; Habermann, A.; Jager, S.; Muller, B.; Krijnse-Locker, J.; Krausslich, H.G. Ultrastructural analysis of ESCRT proteins suggests a role for endosome-associated tubular-vesicular membranes in ESCRT function. Traffic 2006, 7, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, C.; Burton, A.; Ruiz-Mateos, E.; Marsh, M.; Saiardi, A. Inositol pyrophosphate mediated pyrophosphorylation of AP3B1 regulates HIV-1 Gag release. Proc. Natl. Acad. Sci. U. S. A. 2009, 106, 21161–21166. [Google Scholar] [CrossRef] [PubMed]
- Batonick, M.; Favre, M.; Boge, M.; Spearman, P.; Honing, S.; Thali, M. Interaction of HIV-1 Gag with the clathrin-associated adaptor AP-2. Virology 2005, 342, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Camus, G.; Segura-Morales, C.; Molle, D.; Lopez-Verges, S.; Begon-Pescia, C.; Cazevieille, C.; Schu, P.; Bertrand, E.; Berlioz-Torrent, C.; Basyuk, E. The clathrin adaptor complex AP-1 binds HIV-1 and MLV Gag and facilitates their budding. Mol. Biol. Cell 2007, 18, 3193–3203. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Li, H.; Derdowski, A.; Ding, L.; Burnett, A.; Chen, X.; Peters, T.R.; Dermody, T.S.; Woodruff, E.; Wang, J.J.; Spearman, P. AP-3 directs the intracellular trafficking of HIV-1 Gag and plays a key role in particle assembly. Cell 2005, 120, 663–674. [Google Scholar] [CrossRef] [PubMed]
- Garcia, E.; Nikolic, D.S.; Piguet, V. HIV-1 replication in dendritic cells occurs through a tetraspanin-containing compartment enriched in AP-3. Traffic 2008, 9, 200–214. [Google Scholar] [PubMed]
- Joshi, A.; Garg, H.; Nagashima, K.; Bonifacino, J.S.; Freed, E.O. GGA and Arf proteins modulate retrovirus assembly and release. Mol. Cell 2008, 30, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Ryo, A.; Tsurutani, N.; Ohba, K.; Kimura, R.; Komano, J.; Nishi, M.; Soeda, H.; Hattori, S.; Perrem, K.; Yamamoto, M.; Chiba, J.; Mimaya, J.; Yoshimura, K.; Matsushita, S.; Honda, M.; Yoshimura, A.; Sawasaki, T.; Aoki, I.; Morikawa, Y.; Yamamoto, N. SOCS1 is an inducible host factor during HIV-1 infection and regulates the intracellular trafficking and stability of HIV-1 Gag. Proc. Natl. Acad. Sci. U. S. A. 2008, 105, 294–299. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.; Tang, Y.; Okada, Y.; Torrey, T.A.; Chattopadhyay, S.K.; Pfleiderer, M.; Falkner, F.G.; Dorner, F.; Choi, W.; Hirokawa, N.; Morse 3rd., H.C. Binding of murine leukemia virus Gag polyproteins to KIF4, a microtubule-based motor protein . J. Virol. 1998, 72, 6898–6901. [Google Scholar] [PubMed]
- Martinez, N.W.; Xue, X.; Berro, R.G.; Kreitzer, G.; Resh, M.D. Kinesin KIF4 regulates intracellular trafficking and stability of the human immunodeficiency virus type 1 Gag polyprotein. J. Virol. 2008, 82, 9937–9950. [Google Scholar] [CrossRef] [PubMed]
- Alroy, I.; Tuvia, S.; Greener, T.; Gordon, D.; Barr, H.M.; Taglicht, D.; Mandil-Levin, R.; Ben-Avraham, D.; Konforty, D.; Nir, A.; Levius, O.; Bicoviski, V.; Dori, M.; Cohen, S.; Yaar, L.; Erez, O.; Propheta-Meiran, O.; Koskas, M.; Caspi-Bachar, E.; Alchanati, I.; Sela-Brown, A.; Moskowitz, H.; Tessmer, U.; Schubert, U.; Reiss, Y. The trans-Golgi network-associated human ubiquitin-protein ligase POSH is essential for HIV type 1 production. Proc. Natl. Acad. Sci. U. S. A. 2005, 102, 1478–1483. [Google Scholar] [CrossRef] [PubMed]
- Ryzhova, E.V.; Vos, R.M.; Albright, A.V.; Harrist, A.V.; Harvey, T.; Gonzalez-Scarano, F. Annexin 2: a novel human immunodeficiency virus type 1 Gag binding protein involved in replication in monocyte-derived macrophages. J. Virol. 2006, 80, 2694–2704. [Google Scholar] [CrossRef] [PubMed]
- Chertova, E.; Chertov, O.; Coren, L.V.; Roser, J.D.; Trubey, C.M.; Bess Jr., J.W.; Sowder, R.C.; Barsov, E.; Hood, B.L.; Fisher, R.J.; Nagashima, K.; Conrads, T.P.; Veenstra, T.D.; Lifson, J.D.; Ott, D.E. Proteomic and biochemical analysis of purified human immunodeficiency virus type 1 produced from infected monocyte-derived macrophages . J. Virol. 2006, 80, 9039–9052. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.M.; Hocking, H.; Li, P.; Burrell, C.J. Rapid and efficient cell-to-cell transmission of human immunodeficiency virus infection from monocyte-derived macrophages to peripheral blood lymphocytes. Virology 1999, 265, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, D.S.; Willey, R.L.; Sato, H.; Chang, L.J.; Blumenthal, R.; Martin, M.A. Quantitation of human immunodeficiency virus type 1 infection kinetics. J. Virol. 1993, 67, 2182–2190. [Google Scholar] [PubMed]
- Phillips, D.M.; Tan, X.; Perotti, M.E.; Zacharopoulos, V.R. Mechanism of monocyte-macrophage-mediated transmission of HIV . AIDS Res. Hum. Retroviruses 1998, 14, S67–S70. [Google Scholar] [PubMed]
- Dustin, M.L.; Colman, D.R. Neural and immunological synaptic relations. Science 2002, 298, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Grakoui, A.; Bromley, S.K.; Sumen, C.; Davis, M.M.; Shaw, A.S.; Allen, P.M.; Dustin, M.L. The immunological synapse: a molecular machine controlling T cell activation. Science 1999, 285, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Igakura, T.; Stinchcombe, J.C.; Goon, P.K.; Taylor, G.P.; Weber, J.N.; Griffiths, G.M.; Tanaka, Y.; Osame, M.; Bangham, C.R. Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science 2003, 299, 1713–1716. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Wu, L.; Bohks, S.M.; KewalRamani, V.N.; Unutmaz, D.; Hope, T.J. Recruitment of HIV and its receptors to dendritic cell-T cell junctions. Science 2003, 300, 1295–1297. [Google Scholar] [CrossRef] [PubMed]
- Piguet, V.; Sattentau, Q. Dangerous liaisons at the virological synapse. J. Clin. Invest. 2004, 114, 605–610. [Google Scholar] [PubMed]
- Chhabra, E.S.; Higgs, H.N. The many faces of actin: matching assembly factors with cellular structures. Nat. Cell Biol. 2007, 9, 1110–1121. [Google Scholar] [CrossRef] [PubMed]
- Harder, T.; Rentero, C.; Zech, T.; Gaus, K. Plasma membrane segregation during T cell activation: probing the order of domains. Curr. Opin. Immunol. 2007, 19, 470–475. [Google Scholar] [CrossRef] [PubMed]
- Vogt, A.B.; Spindeldreher, S.; Kropshofer, H. Clustering of MHC-peptide complexes prior to their engagement in the immunological synapse: lipid raft and tetraspan microdomains. Immunol. Rev. 2002, 189, 136–151. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C. T cell polarization at the virological synapse. Viruses 2010, 2, 1261–1278. [Google Scholar] [CrossRef]
- Vasiliver-Shamis, G.; Dustin, M.L.; Hioe, C.E. HIV-1 virological synapse is not simply a copycat of the immunological synapse. Viruses 2010, 2, 1239–1260. [Google Scholar] [CrossRef]
- Monks, C.R.; Freiberg, B.A.; Kupfer, H.; Sciaky, N.; Kupfer, A. Three-dimensional segregation of supramolecular activation clusters in T cells. Nature 1998, 395, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Granelli-Piperno, A.; Delgado, E.; Finkel, V.; Paxton, W.; Steinman, R.M. Immature dendritic cells selectively replicate macrophagetropic (M-tropic) human immunodeficiency virus type 1, while mature cells efficiently transmit both M- and T-tropic virus to T cells. J. Virol. 1998, 72, 2733–2737. [Google Scholar] [PubMed]
- Garcia, E.; Pion, M.; Pelchen-Matthews, A.; Collinson, L.; Arrighi, J.F.; Blot, G.; Leuba, F.; Escola, J.M.; Demaurex, N.; Marsh, M.; Piguet, V. HIV-1 trafficking to the dendritic cell-T-cell infectious synapse uses a pathway of tetraspanin sorting to the immunological synapse. Traffic 2005, 6, 488–501. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.J.; Reuter, M.A.; McDonald, D. HIV traffics through a specialized, surface-accessible intracellular compartment during trans-infection of T cells by mature dendritic cells . PLoS Pathog. 2008, 4, e1000134. [Google Scholar] [CrossRef] [PubMed]
- Arrighi, J.F.; Pion, M.; Garcia, E.; Escola, J.M.; van Kooyk, Y.; Geijtenbeek, T.B.; Piguet, V. DC-SIGN-mediated infectious synapse formation enhances X4 HIV-1 transmission from dendritic cells to T cells. J. Exp. Med. 2004, 200, 1279–1288. [Google Scholar] [CrossRef] [PubMed]
- Groot, F.; Kuijpers, T.W.; Berkhout, B.; de Jong, E.C. Dendritic cell-mediated HIV-1 transmission to T cells of LAD-1 patients is impaired due to the defect in LFA-1. Retrovirology 2006, 3, 75. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.H.; Wells, C.; Wu, L. Macropinocytosis and cytoskeleton contribute to dendritic cell-mediated HIV-1 transmission to CD4+ T cells. Virology 2008, 381, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.; Kashefi, K.; Hollinshead, M.; Sattentau, Q.J. HIV-1 cell to cell transfer across an Env-induced, actin-dependent synapse. J. Exp. Med. 2004, 199, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.; Sattentau, Q.J. Human immunodeficiency virus type 1 virological synapse formation in T cells requires lipid raft integrity. J. Virol. 2005, 79, 12088–12094. [Google Scholar] [CrossRef] [PubMed]
- Jolly, C.; Mitar, I.; Sattentau, Q.J. Requirement for an intact T-cell actin and tubulin cytoskeleton for efficient assembly and spread of human immunodeficiency virus type 1. J. Virol. 2007, 81, 5547–5560. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.; Grigorov, B.; Senserrich, J.; Clotet, B.; Darlix, J.L.; Muriaux, D.; Este, J.A. A clathrin-dynamin-dependent endocytic pathway for the uptake of HIV-1 by direct T cell-T cell transmission. Antiviral. Res. 2008, 80, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Hubner, W.; McNerney, G.P.; Chen, P.; Dale, B.M.; Gordon, R.E.; Chuang, F.Y.; Li, X.D.; Asmuth, D.M.; Huser, T.; Chen, B.K. Quantitative 3D video microscopy of HIV transfer across T cell virological synapses. Science 2009, 323, 1743–1747. [Google Scholar] [CrossRef] [PubMed]
- Rudnicka, D.; Feldmann, J.; Porrot, F.; Wietgrefe, S.; Guadagnini, S.; Prevost, M.C.; Estaquier, J.; Haase, A.T.; Sol-Foulon, N.; Schwartz, O. Simultaneous cell-to-cell transmission of human immunodeficiency virus to multiple targets through polysynapses. J. Virol. 2009, 83, 6234–6246. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.; Welsch, S.; Jolly, C.; Briggs, J.A.; Vaux, D.; Sattentau, Q.J. Virological synapse-mediated spread of human immunodeficiency virus type 1 between T cells is sensitive to entry inhibition. J. Virol. 2010, 84, 3516–3527. [Google Scholar] [CrossRef] [PubMed]
- Sherer, N.M.; Lehmann, M.J.; Jimenez-Soto, L.F.; Horensavitz, C.; Pypaert, M.; Mothes, W. Retroviruses can establish filopodial bridges for efficient cell-to-cell transmission. Nat. Cell Biol. 2007, 9, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Eugenin, E.A.; Gaskill, P.J.; Berman, J.W. Tunneling nanotubes (TNT) are induced by HIV-infection of macrophages: a potential mechanism for intercellular HIV trafficking. Cell. Immunol. 2009, 254, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Sowinski, S.; Jolly, C.; Berninghausen, O.; Purbhoo, M.A.; Chauveau, A.; Kohler, K.; Oddos, S.; Eissmann, P.; Brodsky, F.M.; Hopkins, C.; Onfelt, B.; Sattentau, Q.; Davis, D.M. Membrane nanotubes physically connect T cells over long distances presenting a novel route for HIV-1 transmission. Nat. Cell Biol. 2008, 10, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Groot, F.; Welsch, S.; Sattentau, Q.J. Efficient HIV-1 transmission from macrophages to T cells across transient virological synapses. Blood 2008, 111, 4660–4663. [Google Scholar] [CrossRef] [PubMed]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
MDPI and ACS Style
Waki, K.; Freed, E.O. Macrophages and Cell-Cell Spread of HIV-1. Viruses 2010, 2, 1603-1620. https://doi.org/10.3390/v2081603
AMA Style
Waki K, Freed EO. Macrophages and Cell-Cell Spread of HIV-1. Viruses. 2010; 2(8):1603-1620. https://doi.org/10.3390/v2081603
Chicago/Turabian StyleWaki, Kayoko, and Eric O. Freed. 2010. "Macrophages and Cell-Cell Spread of HIV-1" Viruses 2, no. 8: 1603-1620. https://doi.org/10.3390/v2081603