Bunyaviruses and the Type I Interferon System

Abstract
:1. Introduction
2. Diseases caused by the Bunyaviridae

{kind=link}
| Genus/ virus | Disease | Vector | Distribution |
|---|---|---|---|
| Orthobunyavirus | |||
| La Crosse (LACV) | Human: encephalitis | Mosquito | N America |
| Ngari (NRIV) | Human: hemorrhagic fever | Mosquito | Africa |
| Oropouche (OROV) | Human: fever | Midge | S. America |
| Tahyna | Human: fever | Mosquito | Europe |
| Phlebovirus | |||
| Punta Toro (PTV) | Human: fever | Sandfly | M America |
| Rift Valley fever (RVFV) | Human: encephalitis, hemorrhagic fever, retinitis, fatality 1℅.Domestic ruminants: necrotic hepatitis, hemorrhage, abortion | Mosquito | Africa |
| Sicilian sandfly fever (SFSV) | Human: fever | Sandfly | Europe, Africa |
| Toscana (TOSV) | Human: fever | Sandfly | Europe |
| Hantavirus | |||
| Hantaan (HTNV) | Human: severe hemorrhagic fever with renal syndrome (HFRS), fatality 5-15℅ | Field mouse | Eastern Europe, Asia |
| Puumala (PUUV) | Human: mild HFRS, fatality 0.1℅ | Bank vole | Western Europe |
| Seoul (SEOV) | Human: moderate HFRS, fatality 1℅ | Rat | Worldwide |
| Sin Nombre (SNV) | Human: hantavirus cardiopulmonary syndrome, fatality 40℅ | Deer mouse | N America |
| Nairovirus | |||
| Crimean-Congo hemorrhagic fever (CCHFV) | Human: hemorrhagic fever, fatality 20-80℅ | Tick, culicoid fly | Eastern Europe, Africa, Asia |
| Nairobi sheep disease | Sheep, goat: fever, hemorrhagic gastroenteritis, abortion | Tick, culicoid fly, mosquito | Africa, Asia |
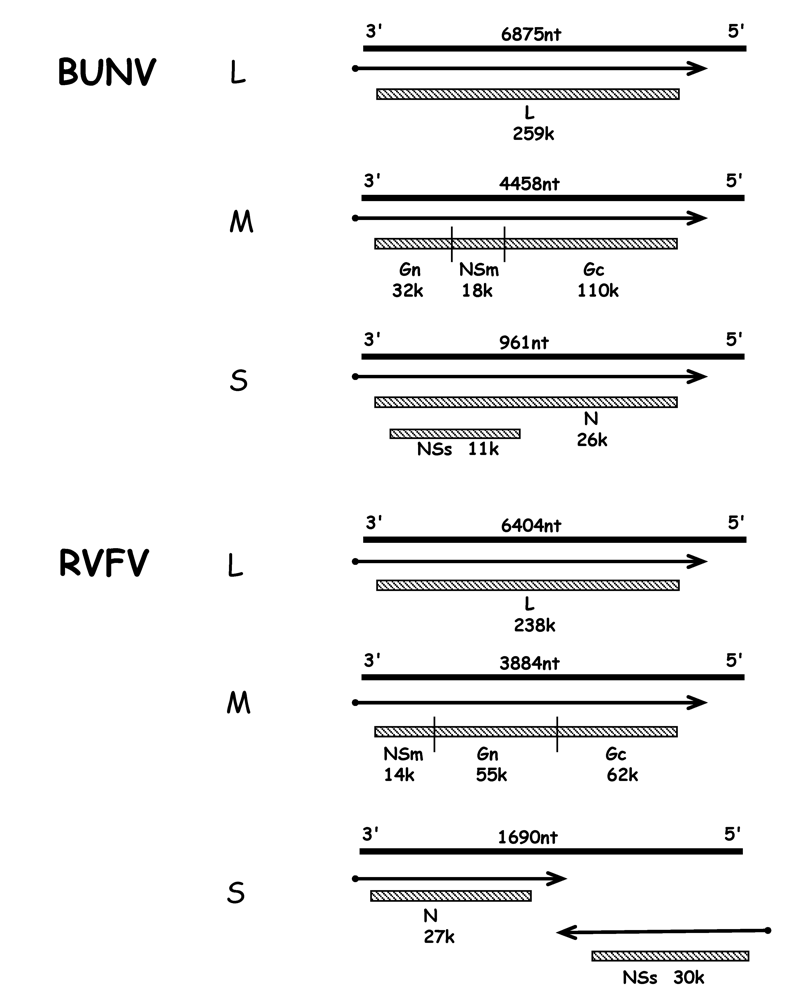
3. Virus particles and infection cycle
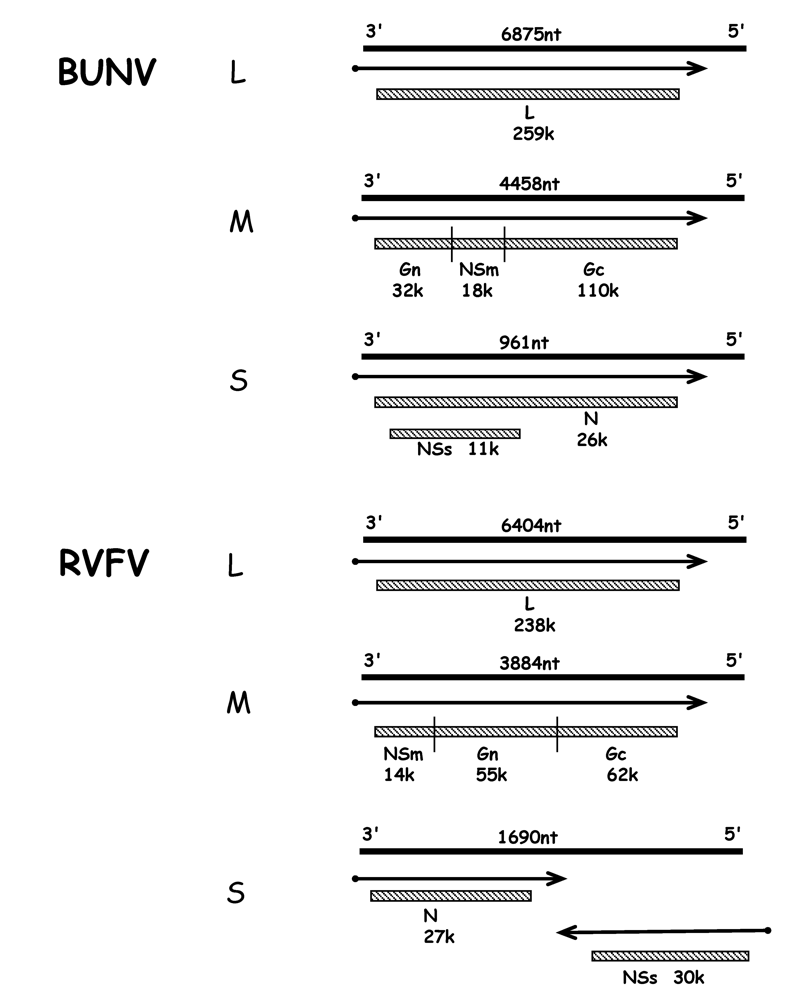
4. Innate immune responses – the type I interferon system
5. IFN responses to bunyaviruses
6. IFN induction by bunyaviruses
7. Viral countermeasures
8. Conclusions
Acknowledgments
References
- Elliott, R.M. The Bunyaviridae. 1996; Plenum Press: New York, NY, USA. [Google Scholar]
- Elliott, R.M. Emerging viruses: the Bunyaviridae. Mol Med 1997, 3, 572–577. [Google Scholar] [PubMed]
- Pinheiro, F.P.; Travassos da Rosa, A.P.; Travassos da Rosa, J.F.; Ishak, R.; Freitas, R.B.; Gomes, M.L.; LeDuc, J.W.; Oliva, O.F. Oropouche virus I. A review of clinical, epidemiological, and ecological findings. Am J Trop Med Hyg 1981, 30, 149–160. [Google Scholar] [PubMed]
- Azevedo, R.S.; Nunes, M.R.; Chiang, J.O.; Bensabath, G.; Vasconcelos, H.B.; Pinto, A.Y.; Martins, L.C.; Monteiro, H.A.; Rodrigues, S.G.; Vasconcelos, P.F. Reemergence of Oropouche fever, northern Brazil. Emerg Infect Dis 2007, 13, 912–915. [Google Scholar] [PubMed]
- Dionisio, D.; Esperti, F.; Vivarelli, A.; Valassina, M. Epidemiological, clinical and laboratory aspects of sandfly fever. Curr Opin Infect Dis 2003, 16, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Charrel, R.N.; Gallian, P.; Navarro-Mari, J.M.; Nicoletti, L.; Papa, A.; Sanchez-Seco, M.P.; Tenorio, A.; de Lamballerie, X. Emergence of Toscana virus in Europe. Emerg Infect Dis 2005, 11, 1657–1663. [Google Scholar] [PubMed]
- Valassina, M.; Cusi, M.G.; Valensin, P.E. A Mediterranean arbovirus: the Toscana virus. J Neurovirol 2003, 9, 577–583. [Google Scholar] [PubMed]
- Nichol, S.T.; Beaty, B.J.; Elliott, R.M.; Goldbach, R.; Plyusnin, A.; Schmaljohn, C.S.; Tesh, R.B. Genus Phlebovirus. Fauguet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier Academic Press: San Diego, CA, USA, 2005. [Google Scholar]
- McJunkin, J.E.; de los Reyes, E.C.; Irazuzta, J.E.; Caceres, M.J.; Khan, R.R.; Minnich, L.L.; Fu, K.D.; Lovett, G.D.; Tsai, T.; Thompson, A. La Crosse encephalitis in children. N Engl J Med 2001, 344, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Calisher, C.H. Medically important arboviruses of the United States and Canada. Clin Microbiol Rev 1994, 7, 89–116. [Google Scholar] [PubMed]
- Haddow, A.D.; Odoi, A. The incidence risk, clustering, and clinical presentation of La Crosse virus infections in the eastern United States, 2003-2007. PLoS One 2009, 4, e6145. [Google Scholar] [CrossRef] [PubMed]
- Utz, J.T.; Apperson, C.S.; MacCormack, J.N.; Salyers, M.; Dietz, E.J.; McPherson, J.T. Economic and social impacts of La Crosse encephalitis in western North Carolina. Am J Trop Med Hyg 2003, 69, 509–518. [Google Scholar] [PubMed]
- Daubney, R.; Hudson, J.R.; Gamham, P.C. Enzootic hepatitis of Rift Valley fever: an undescribed virus disease of sheep, cattle and man from East Africa. J. Pathol. Bacteriol. 1931, 34, 545–549. [Google Scholar] [CrossRef]
- Balkhy, H.H.; Memish, Z.A. Rift Valley fever: an uninvited zoonosis in the Arabian peninsula. Int J Antimicrob Agents 2003, 21, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Bird, B.H.; Ksiazek, T.G.; Nichol, S.T.; Maclachlan, N.J. Rift Valley fever virus. J Am Vet Med Assoc 2009, 234, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Flick, R.; Bouloy, M. Rift Valley fever virus. Curr Mol Med 2005, 5, 827–834. [Google Scholar] [CrossRef]
- Borio, L.; Inglesby, T.; Peters, C.J.; Schmaljohn, A.L.; Hughes, J.M.; Jahrling, P.B.; Ksiazek, T.; Johnson, K.M.; Meyerhoff, A.; O'Toole, T.; Ascher, M.S.; Bartlett, J.; Breman, J.G.; Eitzen Jr., E.M.; Hamburg, M.; Hauer, J.; Henderson, D.A.; Johnson, R.T.; Kwik, G.; Layton, M.; Lillibridge, S.; Nabel, G.J.; Osterholm, M.T.; Perl, T.M.; Russell, P.; Tonat, K. Hemorrhagic fever viruses as biological weapons: medical and public health management. Jama 2002, 287, 2391–2405. [Google Scholar] [CrossRef] [PubMed]
- Bowen, M.D.; Trappier, S.G.; Sanchez, A.J.; Meyer, R.F.; Goldsmith, C.S.; Zaki, S.R.; Dunster, L.M.; Peters, C.J.; Ksiazek, T.G.; Nichol, S.T. A reassortant bunyavirus isolated from acute hemorrhagic fever cases in Kenya and Somalia. Virology 2001, 291, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Briese, T.; Bird, B.; Kapoor, V.; Nichol, S.T.; Lipkin, W.I. Batai and Ngari viruses: M segment reassortment and association with severe febrile disease outbreaks in East Africa. J Virol 2006, 80, 5627–5630. [Google Scholar] [CrossRef] [PubMed]
- Gerrard, S.R.; Li, L.; Barrett, A.D.; Nichol, S.T. Ngari virus is a Bunyamwera virus reassortant that can be associated with large outbreaks of hemorrhagic fever in Africa. J Virol 2004, 78, 8922–8926. [Google Scholar] [CrossRef] [PubMed]
- Yanase, T.; Kato, T.; Yamakawa, M.; Takayoshi, K.; Nakamura, K.; Kokuba, T.; Tsuda, T. Genetic characterization of Batai virus indicates a genomic reassortment between orthobunyaviruses in nature. Arch Virol 2006, 151, 2253–2260. [Google Scholar] [CrossRef] [PubMed]
- Ergonul, O. Crimean-Congo haemorrhagic fever. Lancet Infect Dis 2006, 6, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Mirazimi, A. Interferon and cytokine responses to Crimean Congo hemorrhagic fever virus; an emerging and neglected viral zonoosis. Cytokine Growth Factor Rev 2008, 19, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Nichol, S.T.; Spiropoulou, C.F.; Morzunov, S.; Rollin, P.E.; Ksiazek, T.G.; Feldmann, H.; Sanchez, A.; Childs, J.; Zaki, S.; Peters, C.J. Genetic identification of a hantavirus associated with an outbreak of acute respiratory illness. Science 1993, 262, 914–917. [Google Scholar] [PubMed]
- Freiberg, A.N.; Sherman, M.B.; Morais, M.C.; Holbrook, M.R.; Watowich, S.J. Three-dimensional organization of Rift Valley fever virus revealed by cryoelectron tomography. J Virol 2008, 82, 10341–10348. [Google Scholar] [CrossRef] [PubMed]
- Huiskonen, J.T.; Overby, A.K.; Weber, F.; Grunewald, K. Electron cryo-microscopy and single-particle averaging of Rift Valley fever virus: evidence for GN-GC glycoprotein heterodimers. J Virol 2009, 83, 3762–3769. [Google Scholar] [CrossRef] [PubMed]
- Overby, A.K.; Pettersson, R.F.; Grunewald, K.; Huiskonen, J.T. Insights into bunyavirus architecture from electron cryotomography of Uukuniemi virus. Proc Natl Acad Sci U S A 2008, 105, 2375–2379. [Google Scholar] [CrossRef] [PubMed]
- Sherman, M.B.; Freiberg, A.N.; Holbrook, M.R.; Watowich, S.J. Single-particle cryo-electron microscopy of Rift Valley fever virus. Virology 2009, 387, 11–15. [Google Scholar] [CrossRef] [PubMed]
- Albarino, C.G.; Bird, B.H.; Nichol, S.T. A shared transcription termination signal on negative and ambisense RNA genome segments of Rift Valley fever, sandfly fever Sicilian, and Toscana viruses. J Virol 2007, 81, 5246–5256. [Google Scholar] [CrossRef] [PubMed]
- Barr, J.N.; Rodgers, J.W.; Wertz, G.W. Identification of the Bunyamwera bunyavirus transcription termination signal. J Gen Virol 2006, 87, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Won, S.; Peters, C.J.; Makino, S. Characterization of Rift Valley fever virus transcriptional terminations. J Virol 2007, 81, 8421–8438. [Google Scholar] [CrossRef] [PubMed]
- Pichlmair, A.; Reis e Sousa, C. Innate recognition of viruses. Immunity 2007, 27, 370–383. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: an interplay between induction, signalling, antiviral responses and virus countermeasures. J Gen Virol 2008, 89, 1–47. [Google Scholar] [CrossRef]
- Yoneyama, M.; Fujita, T. RNA recognition and signal transduction by RIG-I-like receptors. Immunol Rev 2009, 227, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Gilfoy, F.D.; Mason, P.W. West Nile virus-induced interferon production is mediated by the double-stranded RNA-dependent protein kinase PKR. J Virol 2007, 81, 11148–11158. [Google Scholar] [CrossRef] [PubMed]
- McAllister, C.S.; Samuel, C.E. The RNA-activated protein kinase enhances the induction of interferon-beta and apoptosis mediated by cytoplasmic RNA sensors. J Biol Chem 2009, 284, 1644–1651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Samuel, C.E. Induction of protein kinase PKR-dependent activation of interferon regulatory factor 3 by vaccinia virus occurs through adapter IPS-1 signaling. J Biol Chem 2008, 283, 34580–34587. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takeuchi, O.; Mikamo-Satoh, E.; Hirai, R.; Kawai, T.; Matsushita, K.; Hiiragi, A.; Dermody, T.S.; Fujita, T.; Akira, S. Length-dependent recognition of double-stranded ribonucleic acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene 5. J Exp Med 2008, 205, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Hornung, V.; Ellegast, J.; Kim, S.; Brzozka, K.; Jung, A.; Kato, H.; Poeck, H.; Akira, S.; Conzelmann, K.K.; Schlee, M.; Endres, S.; Hartmann, G. 5'-Triphosphate RNA Is the Ligand for RIG-I. Science 2006, 314. [Google Scholar] [CrossRef]
- Pichlmair, A.; Schulz, O.; Tan, C.P.; Naslund, T.I.; Liljestrom, P.; Weber, F.; Reis, E.S.C. RIG-I-Mediated Antiviral Responses to Single-Stranded RNA Bearing 5' Phosphates. Science 2006, 314, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Plumet, S.; Herschke, F.; Bourhis, J.M.; Valentin, H.; Longhi, S.; Gerlier, D. Cytosolic 5'-Triphosphate Ended Viral Leader Transcript of Measles Virus as Activator of the RIG I-Mediated Interferon Response. PLoS ONE 2007, 2, e279. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Wagner, V.; Rasmussen, S.B.; Hartmann, R.; Paludan, S.R. Double-stranded RNA is produced by positive-strand RNA viruses and DNA viruses but not in detectable amounts by negative-strand RNA viruses. J Virol 2006, 80, 5059–5064. [Google Scholar] [CrossRef] [PubMed]
- Kato, H.; Takeuchi, O.; Sato, S.; Yoneyama, M.; Yamamoto, M.; Matsui, K.; Uematsu, S.; Jung, A.; Kawai, T.; Ishii, K.J.; Yamaguchi, O.; Otsu, K.; Tsujimura, T.; Koh, C.S.; Reis e Sousa, C.; Matsuura, Y.; Fujita, T.; Akira, S. Differential roles of MDA5 and RIG-I helicases in the recognition of RNA viruses. Nature 2006, 441, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Habjan, M.; Andersson, I.; Klingstrom, J.; Schumann, M.; Martin, A.; Zimmermann, P.; Wagner, V.; Pichlmair, A.; Schneider, U.; Muhlberger, E.; Mirazimi, A.; Weber, F. Processing of genome 5' termini as a strategy of negative-strand RNA viruses to avoid RIG-I-dependent interferon induction. PLoS One 2008, 3, e2032. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, J. Triggering the innate antiviral response through IRF-3 activation. J Biol Chem 2007, 282, 15325–15329. [Google Scholar] [CrossRef] [PubMed]
- Paun, A.; Pitha, P.M. The IRF family, revisited. Biochimie 2007, 89, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Escalante, C.R.; Nistal-Villan, E.; Shen, L.; Garcia-Sastre, A.; Aggarwal, A.K. Structure of IRF-3 bound to the PRDIII-I regulatory element of the human interferon-beta enhancer. Mol Cell 2007, 26, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin Microbiol Rev 2009, 22, 240–273. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, S.B.; Reinert, L.S.; Paludan, S.R. Innate recognition of intracellular pathogens: detection and activation of the first line of defense. Apmis 2009, 117, 323–337. [Google Scholar] [CrossRef] [PubMed]
- de Weerd, N.A.; Samarajiwa, S.A.; Hertzog, P.J. Type I interferon receptors: biochemistry and biological functions. J Biol Chem 2007, 282, 20053–20057. [Google Scholar] [CrossRef] [PubMed]
- Schindler, C.; Levy, D.E.; Decker, T. JAK-STAT signaling: from interferons to cytokines. J Biol Chem 2007, 282, 20059–20063. [Google Scholar] [CrossRef] [PubMed]
- Der, S.D.; Zhou, A.; Williams, B.R.; Silverman, R.H. Identification of genes differentially regulated by interferon alpha, beta, or gamma using oligonucleotide arrays. Proc Natl Acad Sci U S A 1998, 95, 15623–15628. [Google Scholar] [CrossRef] [PubMed]
- Haller, O.; Kochs, G.; Weber, F. Interferon, Mx, and viral countermeasures. Cytokine Growth Factor Rev 2007, 18, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Kochs, G.; Haller, O. GTP-bound human MxA protein interacts with the nucleocapsids of Thogoto virus (Orthomyxoviridae). J Biol Chem 1999, 274, 4370–4376. [Google Scholar] [CrossRef] [PubMed]
- Kochs, G.; Haller, O. Interferon-induced human MxA GTPase blocks nuclear import of Thogoto virus nucleocapsids. Proc Natl Acad Sci U S A 1999, 96, 2082–2086. [Google Scholar] [CrossRef] [PubMed]
- Kochs, G.; Janzen, C.H.; Hohenberg, H.; Haller, O. Antivirally active MxA protein sequesters La Crosse virus nucleocapsid protein into perinuclear complexes. Proc Natl Acad Sci U S A 2002, 99, 3153–3158. [Google Scholar] [CrossRef] [PubMed]
- Nallagatla, S.R.; Hwang, J.; Toroney, R.; Zheng, X.; Cameron, C.E.; Bevilacqua, P.C. 5'-triphosphate-dependent activation of PKR by RNAs with short stem-loops. Science 2007, 318, 1455–1458. [Google Scholar] [CrossRef] [PubMed]
- Dauber, B.; Martinez-Sobrido, L.; Schneider, J.; Hai, R.; Waibler, Z.; Kalinke, U.; Garcia-Sastre, A.; Wolff, T. Influenza B virus ribonucleoprotein is a potent activator of the antiviral kinase PKR. PLoS Pathog 2009, 5, e1000473. [Google Scholar] [CrossRef] [PubMed]
- Garcia, M.A.; Meurs, E.F.; Esteban, M. The dsRNA protein kinase PKR: virus and cell control. Biochimie 2007, 89, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Hovanessian, A.G.; Justesen, J. The human 2'-5'oligoadenylate synthetase family: unique interferon-inducible enzymes catalyzing 2'-5' instead of 3'-5' phosphodiester bond formation. Biochimie 2007, 89, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Bisbal, C.; Silverman, R.H. Diverse functions of RNase L and implications in pathology. Biochimie 2007, 89, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Samuel, C.E. Antiviral actions of interferons. Clin Microbiol Rev 2001, 14, 778–809. [Google Scholar] [CrossRef] [PubMed]
- Sadler, A.J.; Williams, B.R. Interferon-inducible antiviral effectors. Nat Rev Immunol 2008, 8, 559–568. [Google Scholar] [CrossRef]
- Haller, O.; Kochs, G.; Weber, F. The interferon response circuit: induction and suppression by pathogenic viruses. Virology 2006, 344, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Scarano, F.; Bupp, K.; Nathanson, N. Pathogenesis of diseases caused by viruses of the Bunyavirus genus. The Bunyaviridae 1996. [Google Scholar]
- Kim, G.R.; McKee Jr., K.T. Pathogenesis of Hantaan virus infection in suckling mice: clinical, virologic, and serologic observations. Am J Trop Med Hyg 1985, 34, 388–395. [Google Scholar] [PubMed]
- Morahan, P.S.; Grossberg, S.E. Age-related cellular resistance of the chicken embryo to viral infections I. Interferon and natural resistance to myxoviruses and vesicular stomatitis virus. J Infect Dis 1970, 121, 615–623. [Google Scholar] [PubMed]
- Pfeifer, K.; Ushijima, H.; Lorenz, B.; Muller, W.E.; Schroder, H.C. Evidence for age-dependent impairment of antiviral 2',5'-oligoadenylate synthetase/ribonuclease L-system in tissues of rat. Mech Ageing Dev 1993, 67, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Andersson, I.; Lundkvist, A.; Haller, O.; Mirazimi, A. Type I interferon inhibits Crimean-Congo hemorrhagic fever virus in human target cells. J Med Virol 2006, 78, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Habjan, M.; Pichlmair, A.; Elliott, R.M.; Overby, A.K.; Glatter, T.; Gstaiger, M.; Superti-Furga, G.; Unger, H.; Weber, F. NSs protein of rift valley fever virus induces the specific degradation of the double-stranded RNA-dependent protein kinase. J Virol 2009, 83, 4365–4375. [Google Scholar] [CrossRef] [PubMed]
- Livonesi, M.C.; de Sousa, R.L.; Badra, S.J.; Figueiredo, L.T. In vitro and in vivo studies of the Interferon-alpha action on distinct Orthobunyavirus. Antiviral Res 2007, 75, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Luby, J.P. Sensitivities of neurotropic arboviruses to human interferon. J Infect Dis 1975, 132, 361–367. [Google Scholar] [PubMed]
- Morrill, J.C.; Jennings, G.B.; Cosgriff, T.M.; Gibbs, P.H.; Peters, C.J. Prevention of Rift Valley fever in rhesus monkeys with interferon-alpha. Rev Infect Dis 1989, 11, S815–825. [Google Scholar] [PubMed]
- Tamura, M.; Asada, H.; Kondo, K.; Takahashi, M.; Yamanishi, K. Effects of human and murine interferons against hemorrhagic fever with renal syndrome (HFRS) virus (Hantaan virus). Antiviral Res 1987, 8, 171–178. [Google Scholar] [CrossRef]
- Temonen, M.; Lankinen, H.; Vapalahti, O.; Ronni, T.; Julkunen, I.; Vaheri, A. Effect of interferon-alpha and cell differentiation on Puumala virus infection in human monocyte/macrophages. Virology 1995, 206, 8–15. [Google Scholar] [CrossRef]
- Blakqori, G.; Delhaye, S.; Habjan, M.; Blair, C.D.; Sanchez-Vargas, I.; Olson, K.E.; Attarzadeh-Yazdi, G.; Fragkoudis, R.; Kohl, A.; Kalinke, U.; Weiss, S.; Michiels, T.; Staeheli, P.; Weber, F. La Crosse bunyavirus nonstructural protein NSs serves to suppress the type I interferon system of mammalian hosts. J Virol 2007, 81, 4991–4999. [Google Scholar] [CrossRef] [PubMed]
- Bouloy, M.; Janzen, C.; Vialat, P.; Khun, H.; Pavlovic, J.; Huerre, M.; Haller, O. Genetic evidence for an interferon-antagonistic function of rift valley fever virus nonstructural protein NSs. J Virol 2001, 75, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.; Fazakerley, J.K.; Bridgen, A. Pathogenesis of Dugbe virus infection in wild-type and interferon-deficient mice. J Gen Virol 2006, 87, 2005–2009. [Google Scholar] [CrossRef] [PubMed]
- Hefti, H.P.; Frese, M.; Landis, H.; Di Paolo, C.; Aguzzi, A.; Haller, O.; Pavlovic, J. Human MxA protein protects mice lacking a functional alpha/beta interferon system against La crosse virus and other lethal viral infections. J Virol 1999, 73, 6984–6991. [Google Scholar] [PubMed]
- Weber, F.; Dunn, E.F.; Bridgen, A.; Elliott, R.M. The Bunyamwera virus nonstructural protein NSs inhibits viral RNA synthesis in a minireplicon system. Virology 2001, 281, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Wichmann, D.; Grone, H.J.; Frese, M.; Pavlovic, J.; Anheier, B.; Haller, O.; Klenk, H.D.; Feldmann, H. Hantaan virus infection causes an acute neurological disease that is fatal in adult laboratory mice. J Virol 2002, 76, 8890–8899. [Google Scholar] [CrossRef] [PubMed]
- Andersson, I.; Bladh, L.; Mousavi-Jazi, M.; Magnusson, K.E.; Lundkvist, A.; Haller, O.; Mirazimi, A. Human MxA protein inhibits the replication of Crimean-Congo hemorrhagic fever virus. J Virol 2004, 78, 4323–4329. [Google Scholar] [CrossRef] [PubMed]
- Frese, M.; Kochs, G.; Feldmann, H.; Hertkorn, C.; Haller, O. Inhibition of bunyaviruses, phleboviruses, and hantaviruses by human MxA protein. J Virol 1996, 70, 915–923. [Google Scholar] [PubMed]
- Habjan, M.; Penski, N.; Wagner, V.; Spiegel, M.; Overby, A.K.; Kochs, G.; Huiskonen, J.T.; Weber, F. Efficient production of Rift Valley fever virus-like particles: The antiviral protein MxA can inhibit primary transcription of bunyaviruses. Virology 2009, 385, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Kanerva, M.; Melen, K.; Vaheri, A.; Julkunen, I. Inhibition of puumala and tula hantaviruses in Vero cells by MxA protein. Virology 1996, 224, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Sandrock, M.; Frese, M.; Haller, O.; Kochs, G. Interferon-induced rat Mx proteins confer resistance to Rift Valley fever virus and other arthropod-borne viruses. J Interferon Cytokine Res 2001, 21, 663–668. [Google Scholar] [PubMed]
- Miura, T.A.; Carlson, J.O.; Beaty, B.J.; Bowen, R.A.; Olson, K.E. Expression of human MxA protein in mosquito cells interferes with LaCrosse virus replication. J Virol 2001, 75, 3001–3003. [Google Scholar] [CrossRef] [PubMed]
- Reichelt, M.; Stertz, S.; Krijnse-Locker, J.; Haller, O.; Kochs, G. Missorting of LaCrosse virus nucleocapsid protein by the interferon-induced MxA GTPase involves smooth ER membranes. Traffic 2004, 5, 772–784. [Google Scholar] [CrossRef] [PubMed]
- Oelschlegel, R.; Kruger, D.H.; Rang, A. MxA-independent inhibition of Hantaan virus replication induced by type I and type II interferon in vitro. Virus Res 2007, 127, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Bridgen, A.; Fazakerley, J.K.; Streitenfeld, H.; Randall, R.E.; Elliott, R.M. Bunyamwera bunyavirus nonstructural protein NSs counteracts the induction of alpha/beta interferon. J. Virol. 2002, 76, 7949–7955. [Google Scholar] [CrossRef] [PubMed]
- Streitenfeld, H.; Boyd, A.; Fazakerley, J.K.; Bridgen, A.; Elliott, R.M.; Weber, F. Activation of PKR by Bunyamwera virus is independent of the viral interferon antagonist NSs. J. Virol. 2003, 77, 5507–5511. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, T.; Narayanan, K.; Won, S.; Kamitani, W.; Peters, C.J.; Makino, S. Rift Valley fever virus NSs protein promotes post-transcriptional downregulation of protein kinase PKR and inhibits eIF2alpha phosphorylation. PLoS Pathog 2009, 5, e1000287. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.F.; Wickham, M.E.; Coombes, B.K.; Finlay, B.B. Crossing the line: selection and evolution of virulence traits. PLoS Pathog 2006, 2, e42. [Google Scholar] [CrossRef] [PubMed]
- Morrill, J.C.; Jennings, G.B.; Johnson, A.J.; Cosgriff, T.M.; Gibbs, P.H.; Peters, C.J. Pathogenesis of Rift Valley fever in rhesus monkeys: role of interferon response. Arch Virol 1990, 110, 195–212. [Google Scholar] [CrossRef] [PubMed]
- Anderson Jr., G.W.; Slayter, M.V.; Hall, W.; Peters, C.J. Pathogenesis of a phleboviral infection (Punta Toro virus) in golden Syrian hamsters. Arch Virolrch V 1990, 114, 203–212. [Google Scholar]
- Perrone, L.A.; Narayanan, K.; Worthy, M.; Peters, C.J. The S segment of Punta Toro virus (Bunyaviridae, Phlebovirus) is a major determinant of lethality in the Syrian hamster and codes for a type I interferon antagonist. J Virol 2007, 81, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Alff, P.J.; Gavrilovskaya, I.N.; Gorbunova, E.; Endriss, K.; Chong, Y.; Geimonen, E.; Sen, N.; Reich, N.C.; Mackow, E.R. The pathogenic NY-1 hantavirus G1 cytoplasmic tail inhibits RIG-I- and TBK-1-directed interferon responses. J Virol 2006, 80, 9676–9686. [Google Scholar] [CrossRef] [PubMed]
- Geimonen, E.; Neff, S.; Raymond, T.; Kocer, S.S.; Gavrilovskaya, I.N.; Mackow, E.R. Pathogenic and nonpathogenic hantaviruses differentially regulate endothelial cell responses. Proc Natl Acad Sci U S A 2002, 99, 13837–13842. [Google Scholar] [CrossRef] [PubMed]
- Handke, W.; Oelschlegel, R.; Franke, R.; Kruger, D.H.; Rang, A. Hantaan virus triggers TLR3-dependent innate immune responses. J Immunol 2009, 182, 2849–2858. [Google Scholar] [CrossRef] [PubMed]
- Kraus, A.A.; Raftery, M.J.; Giese, T.; Ulrich, R.; Zawatzky, R.; Hippenstiel, S.; Suttorp, N.; Kruger, D.H.; Schonrich, G. Differential antiviral response of endothelial cells after infection with pathogenic and nonpathogenic hantaviruses. J Virol 2004, 78, 6143–6150. [Google Scholar] [CrossRef] [PubMed]
- Spiropoulou, C.F.; Albarino, C.G.; Ksiazek, T.G.; Rollin, P.E. Andes and Prospect Hill Hantaviruses Differ in Early Induction of Interferon although Both Can Downregulate Interferon Signaling. J Virol 2007, 81, 2769–2776. [Google Scholar] [CrossRef] [PubMed]
- Stoltz, M.; Ahlm, C.; Lundkvist, A.; Klingstrom, J. Lambda Interferon (IFN-{lambda}) in Serum Is Decreased in Hantavirus-Infected Patients, and In Vitro-Established Infection Is Insensitive to Treatment with All IFNs and Inhibits IFN-{gamma}-Induced Nitric Oxide Production. J Virol 2007, 81, 8685–8691. [Google Scholar] [CrossRef] [PubMed]
- Gowen, B.B.; Hoopes, J.D.; Wong, M.H.; Jung, K.H.; Isakson, K.C.; Alexopoulou, L.; Flavell, R.A.; Sidwell, R.W. TLR3 Deletion Limits Mortality and Disease Severity due to Phlebovirus Infection. J Immunol 2006, 177, 6301–6307. [Google Scholar] [PubMed]
- Garcin, D.; Lezzi, M.; Dobbs, M.; Elliott, R.M.; Schmaljohn, C.; Kang, C.Y.; Kolakofsky, D. The 5' ends of Hantaan virus (Bunyaviridae) RNAs suggest a prime-and-realign mechanism for the initiation of RNA synthesis. J Virol 1995, 69, 5754–5762. [Google Scholar] [PubMed]
- Prescott, J.; Ye, C.; Sen, G.; Hjelle, B. Induction of innate immune response genes by Sin Nombre hantavirus does not require viral replication. J Virol 2005, 79, 15007–15015. [Google Scholar] [CrossRef] [PubMed]
- Prescott, J.B.; Hall, P.R.; Bondu-Hawkins, V.S.; Ye, C.; Hjelle, B. Early innate immune responses to Sin Nombre hantavirus occur independently of IFN regulatory factor 3, characterized pattern recognition receptors, and viral entry. J Immunol 2007, 179, 1796–1802. [Google Scholar] [PubMed]
- Khaiboullina, S.F.; Rizvanov, A.A.; Otteson, E.; Miyazato, A.; Maciejewski, J.; St Jeor, S. Regulation of cellular gene expression in endothelial cells by sin nombre and prospect hill viruses. Viral Immunol 2004, 17, 234–251. [Google Scholar] [PubMed]
- Khaiboullina, S.F.; Rizvanov, A.A.; Deyde, V.M.; St Jeor, S.C. Andes virus stimulates interferon-inducible MxA protein expression in endothelial cells. J Med Virol 2005, 75, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Andersson, I.; Karlberg, H.; Mousavi-Jazi, M.; Martinez-Sobrido, L.; Weber, F.; Mirazimi, A. Crimean-Congo hemorrhagic fever virus delays activation of the innate immune response. J Med Virol 2008, 80, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.E.; Noyce, R.S.; Mossman, K.L. Innate cellular response to virus particle entry requires IRF3 but not virus replication. J Virol 2004, 78, 1706–1717. [Google Scholar] [CrossRef] [PubMed]
- Paladino, P.; Cummings, D.T.; Noyce, R.S.; Mossman, K.L. The IFN-independent response to virus particle entry provides a first line of antiviral defense that is independent of TLRs and retinoic acid-inducible gene I. J Immunol 2006, 177, 8008–8016. [Google Scholar] [PubMed]
- Shabman, R.S.; Morrison, T.E.; Moore, C.; White, L.; Suthar, M.S.; Hueston, L.; Rulli, N.; Lidbury, B.; Ting, J.P.; Mahalingam, S.; Heise, M.T. Differential induction of type I interferon responses in myeloid dendritic cells by mosquito and mammalian-cell-derived alphaviruses. J Virol 2007, 81, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Bridgen, A.; Weber, F.; Fazakerley, J.K.; Elliott, R.M. Bunyamwera bunyavirus nonstructural protein NSs is a nonessential gene product that contributes to viral pathogenesis. Proc Natl Acad Sci U S A 2001, 98, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Le May, N.; Dubaele, S.; De Santis, L.P.; Billecocq, A.; Bouloy, M.; Egly, J.M. TFIIH transcription factor, a target for the Rift Valley hemorrhagic fever virus. Cell 2004, 116, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Le May, N.; Mansuroglu, Z.; Leger, P.; Josse, T.; Blot, G.; Billecocq, A.; Flick, R.; Jacob, Y.; Bonnefoy, E.; Bouloy, M. A SAP30 complex inhibits IFN-beta expression in Rift Valley fever virus infected cells. PLoS Pathog 2008, 4, e13. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Blakqori, G.; Wagner, V.; Banholzer, M.; Kessler, N.; Elliott, R.M.; Haller, O.; Weber, F. Inhibition of RNA polymerase II phosphorylation by a viral interferon antagonist. J Biol Chem 2004, 279, 31471–31477. [Google Scholar] [CrossRef] [PubMed]
- Leonard, V.H.; Kohl, A.; Hart, T.J.; Elliott, R.M. Interaction of Bunyamwera Orthobunyavirus NSs protein with mediator protein MED8: a mechanism for inhibiting the interferon response. J Virol 2006, 80, 9667–9675. [Google Scholar] [CrossRef] [PubMed]
- Blakqori, G.; Weber, F. Efficient cDNA-based rescue of La Crosse bunyaviruses expressing or lacking the nonstructural protein NSs. J Virol 2005, 79, 10420–10428. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, M.; McLees, A.; Elliott, R.M. Viruses in the Anopheles A, Anopheles B, and Tete serogroups in the Orthobunyavirus genus (family Bunyaviridae) do not encode an NSs protein. J Virol 2009, 83, 7612–7618. [Google Scholar] [CrossRef] [PubMed]
- Frias-Staheli, N.; Giannakopoulos, N.V.; Kikkert, M.; Taylor, S.L.; Bridgen, A.; Paragas, J.; Richt, J.A.; Rowland, R.R.; Schmaljohn, C.S.; Lenschow, D.J.; Snijder, E.J.; Garcia-Sastre, A.; Virgin, H.W.t. Ovarian tumor domain-containing viral proteases evade ubiquitin- and ISG15-dependent innate immune responses. Cell Host Microbe 2007, 2, 404–416. [Google Scholar] [CrossRef]
- Plyusnin, A. Genetics of hantaviruses: implications to taxonomy. Arch Virol 2002, 147, 665–682. [Google Scholar] [CrossRef] [PubMed]
- Jaaskelainen, K.M.; Kaukinen, P.; Minskaya, E.S.; Plyusnina, A.; Vapalahti, O.; Elliott, R.M.; Weber, F.; Vaheri, A.; Plyusnin, A. Tula and Puumala hantavirus NSs ORFs are functional and the products inhibit activation of the interferon-beta promoter. J Med Virol 2007, 79, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Jaaskelainen, K.M.; Plyusnina, A.; Lundkvist, A.; Vaheri, A.; Plyusnin, A. Tula hantavirus isolate with the full-length ORF for nonstructural protein NSs survives for more consequent passages in interferon-competent cells than the isolate having truncated NSs ORF. Virol J 2008, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Alff, P.J.; Sen, N.; Gorbunova, E.; Gavrilovskaya, I.N.; Mackow, E.R. The NY-1 hantavirus Gn cytoplasmic tail coprecipitates TRAF3 and inhibits cellular interferon responses by disrupting TBK1-TRAF3 complex formation. J Virol 2008, 82, 9115–9122. [Google Scholar] [CrossRef] [PubMed]
- Sen, N.; Sen, A.; Mackow, E.R. Degrons at the C terminus of the pathogenic but not the nonpathogenic hantavirus G1 tail direct proteasomal degradation. J Virol 2007, 81, 4323–4330. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.L.; Frias-Staheli, N.; Garcia-Sastre, A.; Schmaljohn, C.S. Hantaan virus nucleocapsid protein binds to importin alpha proteins and inhibits tumor necrosis factor alpha-induced activation of nuclear factor kappa B. J Virol 2009, 83, 1271–1279. [Google Scholar] [CrossRef] [PubMed]
- Young, D.F.; Andrejeva, L.; Livingstone, A.; Goodbourn, S.; Lamb, R.A.; Collins, P.L.; Elliott, R.M.; Randall, R.E. Virus replication in engineered human cells that do not respond to interferons. J Virol 2003, 77, 2174–2181. [Google Scholar] [CrossRef] [PubMed]
- Ferko, B.; Stasakova, J.; Romanova, J.; Kittel, C.; Sereinig, S.; Katinger, H.; Egorov, A. Immunogenicity and protection efficacy of replication-deficient influenza A viruses with altered NS1 genes. J Virol 2004, 78, 13037–13045. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Sesma, A.; Marukian, S.; Ebersole, B.J.; Kaminski, D.; Park, M.S.; Yuen, T.; Sealfon, S.C.; Garcia-Sastre, A.; Moran, T.M. Influenza virus evades innate and adaptive immunity via the NS1 protein. J Virol 2006, 80, 6295–6304. [Google Scholar] [CrossRef] [PubMed]
- Richt, J.A.; Lekcharoensuk, P.; Lager, K.M.; Vincent, A.L.; Loiacono, C.M.; Janke, B.H.; Wu, W.H.; Yoon, K.J.; Webby, R.J.; Solorzano, A.; Garcia-Sastre, A. Vaccination of pigs against swine influenza viruses by using an NS1-truncated modified live-virus vaccine. J Virol 2006, 80, 11009–11018. [Google Scholar] [CrossRef] [PubMed]
- Talon, J.; Salvatore, M.; O'Neill, R.E.; Nakaya, Y.; Zheng, H.; Muster, T.; Garcia-Sastre, A.; Palese, P. Influenza A and B viruses expressing altered NS1 proteins: A vaccine approach. Proc Natl Acad Sci U S A 2000, 97, 4309–4314. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Haller, O. Viral suppression of the interferon system. Biochimie 2007, 89, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Bird, B.H.; Albarino, C.G.; Hartman, A.L.; Erickson, B.R.; Ksiazek, T.G.; Nichol, S.T. Rift valley fever virus lacking the NSs and NSm genes is highly attenuated, confers protective immunity from virulent virus challenge, and allows for differential identification of infected and vaccinated animals. J Virol 2008, 82, 2681–2691. [Google Scholar] [CrossRef] [PubMed]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
Share and Cite
Elliott, R.M.; Weber, F. Bunyaviruses and the Type I Interferon System. Viruses 2009, 1, 1003-1021. https://doi.org/10.3390/v1031003
Elliott RM, Weber F. Bunyaviruses and the Type I Interferon System. Viruses. 2009; 1(3):1003-1021. https://doi.org/10.3390/v1031003
Chicago/Turabian StyleElliott, Richard M., and Friedemann Weber. 2009. "Bunyaviruses and the Type I Interferon System" Viruses 1, no. 3: 1003-1021. https://doi.org/10.3390/v1031003