Digestive Enzymes of the Crustaceans Munida and Their Application in Cheese Manufacturing: A Review

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. The Interest for Marine Enzymes in Biotechnological Processes
2. The Crustaceans Munida as a Source of Digestive Enzymes for Cheese Technology
3. The Enzymes of Whole Munida Crustaceans
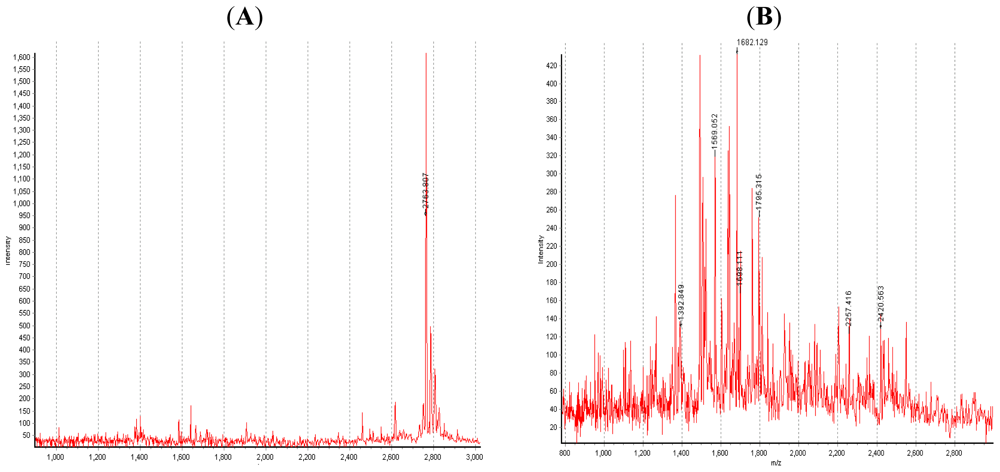
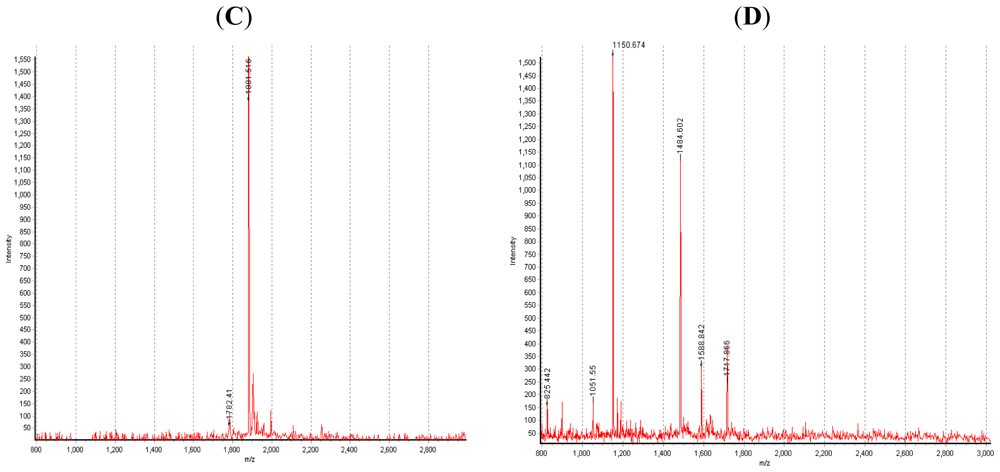
4. The Digestive Enzymes from the Hepatopancreas of Crustaceans Munida
5. Detection of Munida Proteolytic Activities by Casein Gel Zymography
6. Cheesemaking Trials Using the Enzymes Extracted from the Hepatopancreas of Munida Crustaceans
6.1. Mini Cheddar-Type Cheeses
6.2. Ewe Mini-Cheeses
7. Conclusions
References
- Zhang, C; Kim, SK. Research and application of marine microbial enzymes: Status and prospects. Mar. Drugs 2010, 8, 1920–1934. [Google Scholar]
- Rasmussen, RS; Morrissey, MT. Marine biotechnology for production of food ingredients. Adv. Food Nutr. Res 2007, 52, 237–292. [Google Scholar]
- Debashish, G; Malay, S; Barindra, S; Joydeep, M. Marine enzymes. Adv. Biochem. Eng. Biotechnol 2005, 96, 189–218. [Google Scholar]
- Haefner, B. Drugs from the deep: Marine natural products as drug candidates. Drug Discov. Today 2003, 8, 536–544. [Google Scholar]
- Bernan, VS; Greenstein, M; Maiese, WM. Marine micro-organisms as a source of new natural products. Adv. Appl. Microbiol 1997, 43, 57–89. [Google Scholar]
- Diaz-Lopez, M; Garcia-Carreno, FL. Applications of fish and shellfish enzymes in food and feed products. In Seafood Enzymes; Haard, NF, Simpson, BK, Eds.; M Dekker: New York, NY, USA, 2000; pp. 571–618. [Google Scholar]
- Shahidi, F; Janak Kamil, YVA. Enzymes from fish and aquatic invertebrates and their application in the food industry. Trends Food Sci. Technol 2001, 12, 435–464. [Google Scholar]
- Haard, NF; Simpson, BK. Proteases from aquatic organisms and their uses in the seafood industry. In Fisheries Processing: Biotechnological Applications; Martin, AM, Ed.; Chapman and Hall: London, UK, 1994; pp. 132–154. [Google Scholar]
- Venugopal, V; Shahidi, F. Value-added products from underutilized fish species. Crit. Rev. Food Sci. Nutr 1995, 35, 431–453. [Google Scholar]
- Simpson, BK. Digestive proteinases from marine animals. In Seafood Enzymes; Haard, NF, Simpson, BK, Eds.; M Dekker: New York, NY, USA, 2000; pp. 191–214. [Google Scholar]
- Haard, NF; Simpson, BK; Sikorski, ZE. Biotechnological applications of seafood proteins and others nitrogenous compounds. In Seafood Proteins; Sikorski, ZE, Pan, BS, Shahidi, F, Eds.; Chapman and Hall: New York, NY, USA, 1994; pp. 194–202. [Google Scholar]
- Reece, P. Recovery of proteases from fish waste. Process Biochem 1988, 6, 62–66. [Google Scholar]
- Gudmundsdóttir, A; Pálsdóttir, HM. Atlantic cod trypsins: From basic research to practical applications. Mar. Biotechnol. (NY) 2005, 7, 77–88. [Google Scholar]
- Shamsuzzaman, K; Haard, NF. Evaluation of harp seal gastric protase as a rennet substitute for cheddar cheese. J. Food Sci 1983, 48, 179–182. [Google Scholar]
- Shamsuzzaman, K; Haard, NF. Purification and characterization of a chymosin-like protease from gastric mucosa of harp seal (Paophilus groenlandicus). Can. J. Biochem. Cell Biol 1984, 62, 699–708. [Google Scholar]
- Shamsuzzaman, K; Haard, NF. Milk clotting and cheese making properties of a chymosin-like enzyme from harp seal mucosa. J. Food Biochem 1985, 9, 173–192. [Google Scholar]
- Tavares, JFP; Baptista, JAB; Marcone, MF. Milk coagulating enzymes of tuna fish waste as a rennet substitute. Int. J. Food Sci. Technol 1997, 48, 169–176. [Google Scholar]
- Garcìa-Carreño, FL; Hernandez-Cortes, MP; Haard, NF. Enzymes whit peptides and proteinase activity from the digestive system of a freshwater and a marine decapod. J. Agric. Food Chem 1994, 42, 1456–1461. [Google Scholar]
- Falciai, L; Minervini, R. Galatheideae. In Guida dei Crostacei Decapodi d’Europa; Muzio, F, Ed.; Franco Muzzio Editore: Rome, Italy, 1992; pp. 168–174. [Google Scholar]
- Celis-Guerrero, LE; Garcìa-Carreño, FL; Navarrete del Toro, MA. Characterization of proteases in the digestive system of spiny lobster (Panulirus interruptus). Mar. Biotechnol 2004, 6, 262–269. [Google Scholar]
- Muhlia-Almazán, A; Sánchez-Paz, A; García-Carreño, FL. Invertebrate trypsins: A review. J. Comp. Physiol. B 2008, 178, 655–672. [Google Scholar]
- Wu, Z; Jiang, G; Xiang, P; Yang, D; Wang, N. Purification and characterization of trypsin-like enzymes from North Pacific krill (Euphausia pacifica). Biotechnol. Lett 2008, 30, 67–72. [Google Scholar]
- Dall, W; Moriarty, DJW. Functional aspects of nutrition and digestion. In The Biology of Crustaceans, Internal Anatomy and Physiological Regulation; Mantel, LH, Ed.; Academic Press: New York, NY, USA, 1983; pp. 215–261. [Google Scholar]
- Galgani, F; Nagayama, F. Digestive proteinases in the Japanese spiny lobster Panulirus japonicus. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 1987, 87, 889–893. [Google Scholar]
- Fang, L; Lee, B. Ontogenic changes in digestive enzymes in Penaeus monodon. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 1992, 103, 1033–1037. [Google Scholar]
- Garcìa-Carreño, FL. The digestive proteases of langostilla (Pleuroncodes planipes, Decapoda): Their partial characterization and the effect of feed on their composition. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 1992, 103, 575–578. [Google Scholar]
- Glass, H; Stark, J. Protein digestion in the European lobster Homarus gammarus (L). Comp. Biochem. Physiol. B: Biochem. Mol. Biol 1994, 108, 225–235. [Google Scholar]
- Jones, DA; Kumlu, M; Le Vay, L; Fletcher, DJ. The digestive physiology of herbivorous, omnivorous and carnivorous crustacean larvae: a review. Aquaculture 1997, 155, 285–295. [Google Scholar]
- Navarrete del Toro, MA; García-Carreño, FL; Díaz, LM; Celis-Guerrero, L; Saborowski, R. Aspartic proteinases in the digestive tract of marine decapod crustaceans. J. Exp. Zool. A Comp. Exp. Biol 2006, 305A, 645–654. [Google Scholar]
- D’Ambrosio, A; Rossano, R; Ungaro, N; Riccio, P. Proteolytic and milk clotting activities in extracts obtained from the crustaceans Munida. J. Mol. Catal. B Enzym 2003, 865, 1–6. [Google Scholar]
- Rossano, R; Larocca, M; Lamaina, A; Viggiani, S; Riccio, P. The hepatopancreas enzymes of the crustaceans Munida and their potential application in cheese biotechnology. LWT-Food Sci. Technol 2011, 44, 173–180. [Google Scholar]
- Hill, RD; Lahav, E; Givol, D. A rennin-sensitive bond in αs1-B-casein. J. Dairy Res 1974, 41, 147–153. [Google Scholar]
- McSweeney, PLH; Olson, NF; Fox, PF; Healy, A; Højrup, P. Proteolytic specificity of chymosin on bovine αs1-b-casein. J. Dairy Res 1993, 60, 401–412. [Google Scholar]
- Visser, S; Slangen, KJ. On the specificity of chymosin (rennin) in its action on β-casein. Neth. Milk Dairy J 1997, 31, 16–30. [Google Scholar]
- Visser, S; Slangen, KJ; Hup, G; Stadhouders, J. Bitter flavour in cheese III. Comparative gel-chromatographic analysis of hydrophobic peptide fractions from twelve Gouda-type cheeses and identification of bitter peptides isolated from a cheese made with Streptococcus cremoris strain HP. Neth. Milk Dairy J 1983, 37, 181–192. [Google Scholar]
- Kelly, M; Fox, PF; McSweeney, PLH. Influence of salting-moisture on proteolysis in Cheddar-type cheese. Milchwissenschaft 1996, 51, 498–501. [Google Scholar]
- Singh, TK; Young, ND; Drake, M; Cadwallader, KR. Production and sensory characterization of a bitter peptide from b-Casein. J. Agric. Food Chem 2005, 53, 1185–1189. [Google Scholar]
- Soeryapranata, E; Powers, JR; Weller, K; Hill, H; Siems, W. Differentiation of intracellular peptidases of starter and adjunct cultures using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. LWT-Food Sci. Technol 2004, 37, 17–22. [Google Scholar]
- Parra, L; De Palencia, F; Casal, V; Requena, T; Pelaez, C. Hydrolysis of b-casein (193–209) fragment of whole cells and fractions of Lactobacillus casei and Lactococcus lactis. J. Food Sci 1999, 64, 899–902. [Google Scholar]
- Garcìa-Carreño, F; Haard, N. Characterization of proteinase classes in langostilla (Pleuroncodes planipes) and crayfish (Pacifastacus astacus) extracts. J. Food Biochem 1993, 17, 97–113. [Google Scholar]
- Lemos, D; Ezquerra, JM; Garcìa-Carreño, F. Protein digestion in penaeid shrimp: Digestive proteinases, proteinase inhibitors and feed digestibility. Aquaculture 2000, 186, 89–105. [Google Scholar]
- Sainz, JC; García-Carreño, FL; Hernández-Cortés, P. Penaeus vannamei isotrypsins: Purification and characterization. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 2004, 138, 155–162. [Google Scholar]
- Osnes, KK; Mohr, V. On the purification and characterization of three serine-type peptide hydrolases from Antarctic krill, Euphausia superba. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 1985, 82, 607–619. [Google Scholar]
- Fernández Gimenez, AV; García-Carreño, FL; Navarrete del Toro, MA; Fenucci, JL. Digestive proteinases of Artemesia longinaris (Decapoda, Penaeidae) and relationship with molting. Comp. Biochem. Physiol. B: Biochem. Mol. Biol 2002, 132, 593–598. [Google Scholar]
- Shakeel-Ur-Rehman; McSweeney, PLH; Fox, PF. Protocol for the manufacture of miniature cheeses. Lait 1998, 78, 607–620. [Google Scholar]
- Rossano, R; Piraino, P; D’Ambrosio, A; O’Connell, OF; Ungaro, N; McSweeney, PLH; Riccio, P. Proteolysis in miniature Cheddar cheeses manufactured using extracts from the crustacean Munida as coagulant. J. Biotechnol 2005, 120, 220–227. [Google Scholar]
- Sousa, MJ; Ardo, Y; McSweeney, PLH. Advances in the study of proteolysis during cheese ripening. Int. Dairy J 2001, 11, 327–345. [Google Scholar]
- Broome, MC; Xu, X; Mayes, JJ. Proteolysis in Cheddar cheese made with alternative coagulants. Aust. J. Dairy Technol 2006, 61, 85–87. [Google Scholar]
- Rogelj, IB; Perko, B; Francky, A; Penca, V; Pungerčar, J. Recombinant lamb chymosin as an alternative coagulating enzyme in cheese production. J. Dairy Sci 2001, 84, 1020–1026. [Google Scholar]
- Koaze, Y; Goi, H; Ezawa, K; Yamada, Y; Hara, T. Fungal proteolytic enzymes. I. Isolation of two kinds of acid-proteases excreted by Aspergillus niger varmacrosporus. Agric. Biol. Chem 1964, 28, 216–223. [Google Scholar]
- Sternberg, MZ. Crystalline milk-clotting protease from Mucor miehei and some of its properties. J. Dairy Sci 1971, 54, 159–167. [Google Scholar]
- Ostoslavskaya, VI; Revina, LP; Kotlova, EK; Surova, IA; Levin, ED; Timokhina, EA; Stepanov, VM. The primary structure of aspergillopepsin A, aspartic proteinase from Aspergillus awamori. IV. Amino acid sequence of the enzyme. Bioorg. Khim 1986, 12, 1030–1047. [Google Scholar]
- Pitts, JE. Crystallization by centrifugation. Nature 1992, 355, 117–120. [Google Scholar]
- Mackay, VL; Welch, SK; Insley, MY; Manney, TR; Holly, J; Saari, GC; Parker, ML. The Saccharomyces cerevisiae BAR1 gene encodes an exported protein with homology to pepsin. Proc. Natl. Acad. Sci. USA 1988, 8, 55–59. [Google Scholar]
- Togni, G; Sanglard, D; Falchetto, R; Monod, M. Isolation and nucleotide sequence of the extracellular acid protease gene (ACP) from the yeast Candida tropicalis. FEBS Lett 1991, 286, 181–185. [Google Scholar]
- Macedo, IQ; Faro, CJ; Pires, EV. Specificity and kinetics of the milk-clotting enzyme from Cardoon (Cynara cardunculus L.) toward bovine k-casein. J. Agric. Food Chem 1993, 41, 1537–1540. [Google Scholar]
- Fernández-Salguero, J; Sanjuán, E. Influence of vegetable and animal rennet on proteolysis during ripening in ewe’s milk cheese. Food Chem 1999, 64, 177–183. [Google Scholar]
- Sousa, MJ; Malcata, FX. Advances in the role of a plant coagulant (Cynara cardunculus) in vitro and during ripening of cheeses from several milk species. Lait 2002, 82, 151–170. [Google Scholar]
- Fernàndez-Salguero, J; Tejada, L; Gòmez, R. Use of powdered vegetable coagulant in the manufacture of ewe’s milk cheeses. J. Sci. Food Agric 2002, 82, 464–468. [Google Scholar]
- Vioque, M; Gómez, R; Sánchez, E; Mata, C; Tejada, L; Fernández-Salguero, J. Chemical and microbiological characteristics of ewe’s milk cheese manufactured with extracts from flowers of Cynara cardunculus and Cynara humilis. J. Agric. Food Chem 2000, 48, 451–456. [Google Scholar]
- Lopes, A; Teixerira, G; Liberato, MC; Pais, MS; Clemente, A. New vegetal sources of milk clotting enzyme. J. Mol. Catal. B Enzym 1998, 5, 63–68. [Google Scholar]
- Yousif, BH; McMahon, DJ; Shammet, KM. Milk-clotting enzyme from Solanum dobium plant. Int. Dairy J 1996, 6, 637–644. [Google Scholar]
- Tavaria, F; Sousa, MJ; Domingos, A; Malcata, FX; Brodelius, P; Clemente, A; Pais, MS. Degradation of caseins from milk of different species by extracts of Centaurea calcitrapa. J. Agric. Food Chem 1997, 45, 3760–3765. [Google Scholar]
- Pintado, AI; Macedo, AC; Teixeira, G; Pais, MS; Clemente, A; Malcata, FX. Caseinolytic activity of fruit extract from Opuntia ficus-indica on bovine, caprine, and ovine sodium caseinates. Biotechnol. Prog 2001, 17, 643–646. [Google Scholar]
- Reis, PM; Lourenco, PL; Domingos, A; Clemente, AF; Pais, MS; Malcata, FX. Applicability of extracts from Centaurea calcitrapa in ripening of bovine cheese. Int. Dairy J 2000, 10, 775–780. [Google Scholar]
- Li, J; Peng, Y; Wang, X; Chi, Z. Optimum production and characterization of an acid protease from marine yeast Metschnikowia reukaufii W6b. J. Ocean Univ. China 2010, 9, 359–364. [Google Scholar]



© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Rossano, R.; Larocca, M.; Riccio, P. Digestive Enzymes of the Crustaceans Munida and Their Application in Cheese Manufacturing: A Review. Mar. Drugs 2011, 9, 1220-1231. https://doi.org/10.3390/md9071220
Rossano R, Larocca M, Riccio P. Digestive Enzymes of the Crustaceans Munida and Their Application in Cheese Manufacturing: A Review. Marine Drugs. 2011; 9(7):1220-1231. https://doi.org/10.3390/md9071220
Chicago/Turabian StyleRossano, Rocco, Marilena Larocca, and Paolo Riccio. 2011. "Digestive Enzymes of the Crustaceans Munida and Their Application in Cheese Manufacturing: A Review" Marine Drugs 9, no. 7: 1220-1231. https://doi.org/10.3390/md9071220
APA StyleRossano, R., Larocca, M., & Riccio, P. (2011). Digestive Enzymes of the Crustaceans Munida and Their Application in Cheese Manufacturing: A Review. Marine Drugs, 9(7), 1220-1231. https://doi.org/10.3390/md9071220