Bactericidal Kinetics of Marine-Derived Napyradiomycins against Contemporary Methicillin-Resistant Staphylococcus aureus

 and
and
Abstract
:1. Introduction
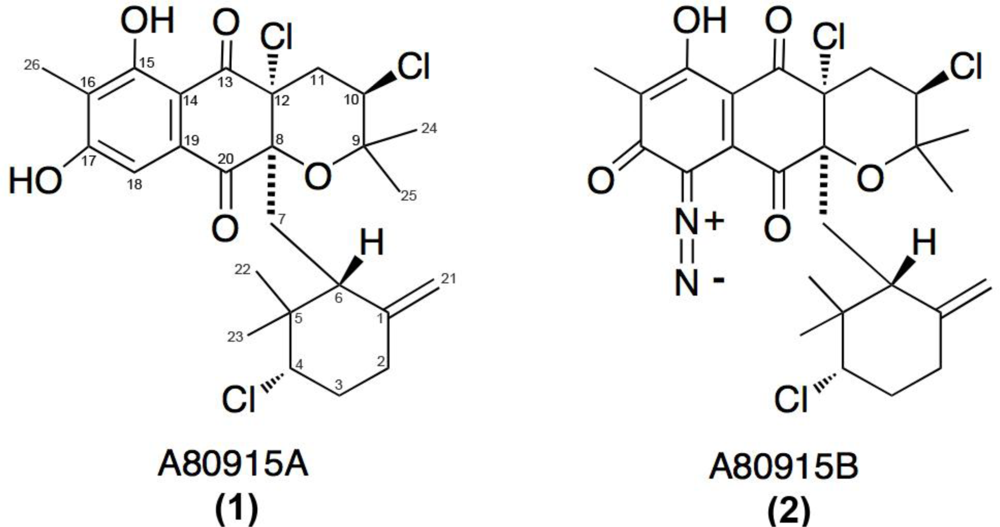
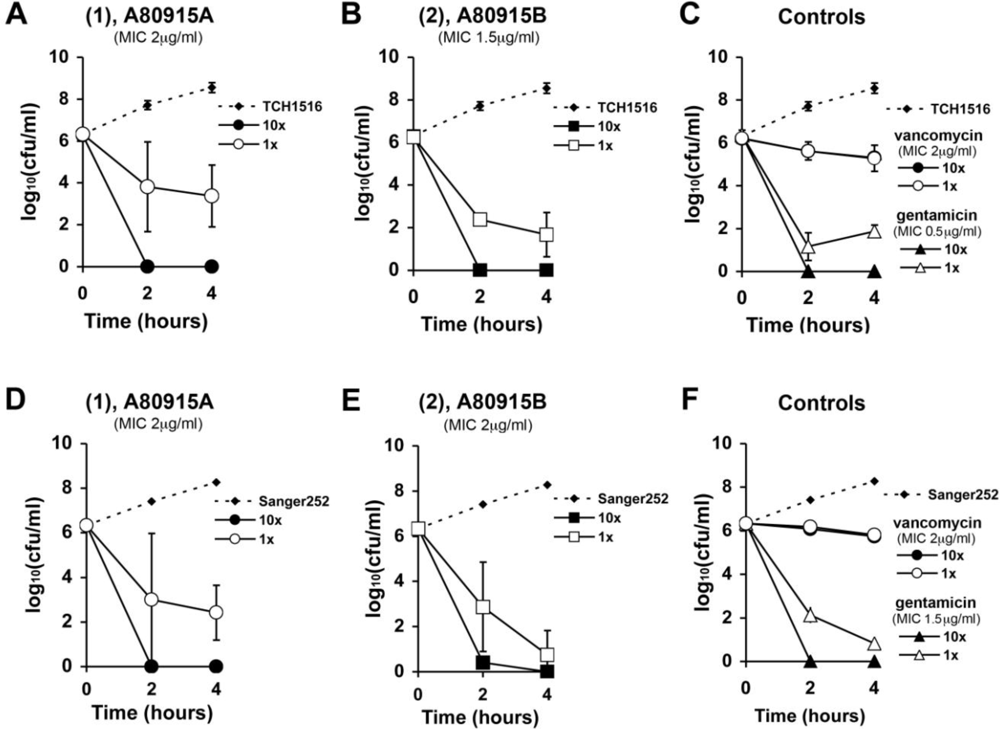
2. Results and Discussion
3. Experimental Section
3.1. Isolation and Cultivation of CNQ-525 and Purification of Napyradiomycin Derivative 1 (A80915A) and Derivative 2 (A80915B)
3.2. Bacterial Strains
3.3. Susceptibility Testing
3.4. Time-Kill Analyses
3.5. Effect of Human Serum on Antibiotic Activity
3.6. Antiproliferative Bioassay: Test for Cytotoxicity
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Como-Sabetti, K; Harriman, KH; Buck, JM; Glennen, A; Boxrud, DJ; Lynfield, R. Community-associated methicillin-resistant Staphylococcus aureus: Trends in case and isolate characteristics from six years of prospective surveillance. Public Health Rep 2009, 124, 427–435. [Google Scholar]
- Boucher, HW; Talbot, GH; Bradley, JS; Edwards, JE; Gilbert, D; Rice, LB; Scheld, M; Spellberg, B; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the infectious diseases society of America. Clin Infect Dis 2009, 48, 1–12. [Google Scholar]
- Spellberg, B; Talbot, GH; Boucher, HW; Bradley, JS; Gilbert, D; Scheld, WM; Edwards, J, Jr; Bartlett, JG. Antimicrobial agents for complicated skin and skin-structure infections: Justification of noninferiority margins in the absence of placebo-controlled trials. Clin Infect Dis 2009, 49, 383–391. [Google Scholar]
- Clardy, J; Fischbach, MA; Walsh, CT. New antibiotics from bacterial natural products. Nat Biotechnol 2006, 24, 1541–1550. [Google Scholar]
- Hopwood, DA. Therapeutic treasures from the deep. Nat Chem Biol 2007, 3, 457–458. [Google Scholar]
- Shiomi, K; Iinuma, H; Hamada, M; Naganawa, H; Manabe, M; Matsuki, C; Takeuchi, T; Umezawa, H. Novel antibiotics napyradiomycins. Production, isolation, physico-chemical properties and biological activity. J Antibiot (Tokyo) 1986, 39, 487–493. [Google Scholar]
- Shiomi, K; Nakamura, H; Iinuma, H; Naganawa, H; Isshiki, K; Takeuchi, T; Umezawa, H; Iitaka, Y. Structures of new antibiotics napyradiomycins. J Antibiot (Tokyo) 1986, 39, 494–501. [Google Scholar]
- Shiomi, K; Iinuma, H; Naganawa, H; Isshiki, K; Takeuchi, T; Umezawa, H. Biosynthesis of napyradiomycins. J Antibiot (Tokyo) 1987, 40, 1740–1745. [Google Scholar]
- Shiomi, K; Nakamura, H; Iinuma, H; Naganawa, H; Takeuchi, T; Umezawa, H; Iitaka, Y. New antibiotic napyradiomycins A2 and B4 and stereochemistry of napyradiomycins. J Antibiot (Tokyo) 1987, 40, 1213–1219. [Google Scholar]
- Fukuda, DS; Mynderse, JS; Baker, PJ; Berry, DM; Boeck, LD; Yao, RC; Mertz, FP; Nakatsukasa, WM; Mabe, J; Ott, J; et al. A80915, a new antibiotic complex produced by Streptomyces aculeolatus. Discovery, taxonomy, fermentation, isolation, characterization, and antibacterial evaluation. J Antibiot (Tokyo) 1990, 43, 623–633. [Google Scholar]
- Fukuda, DS; Mynderse, JS; Yao, RC. Antibiotic A80915 and process for its production. US Patent 4,904,590 1990. [Google Scholar]
- Gomi, S; Ohuchi, S; Sasaki, T; Itoh, J; Sezaki, M. Studies on new antibiotics SF2415. II. The structural elucidation. J Antibiot (Tokyo) 1987, 40, 740–749. [Google Scholar]
- Shomura, T; Gomi, S; Ito, M; Yoshida, J; Tanaka, E; Amano, S; Watabe, H; Ohuchi, S; Itoh, J; Sezaki, M; et al. Studies on new antibiotics SF2415. I. Taxonomy, fermentation, isolation, physico-chemical properties and biological activities. J Antibiot (Tokyo) 1987, 40, 732–739. [Google Scholar]
- Umezawa, K; Masuoka, S; Ohse, T; Naganawa, H; Kondo, S; Ikeda, Y; Kinoshita, N; Hamada, M; Sawa, T; Takeuchi, T. Isolation from Streptomyces of a novel naphthoquinone compound, naphthablin, that inhibits Abl oncogene functions. J Antibiot (Tokyo) 1995, 48, 604–607. [Google Scholar]
- Jensen, PR; Gontang, E; Mafnas, C; Mincer, TJ; Fenical, W. Culturable marine actinomycete diversity from tropical Pacific Ocean sediments. Environ Microbiol 2005, 7, 1039–1048. [Google Scholar]
- Cho, JY; Kwon, HC; Williams, PG; Jensen, PR; Fenical, W. Azamerone, a terpenoid phthalazinone from a marine-derived bacterium related to the genus Streptomyces (Actinomycetales). Org Lett 2006, 8, 2471–2474. [Google Scholar]
- Soria-Mercado, IE; Prieto-Davo, A; Jensen, PR; Fenical, W. Antibiotic terpenoid chloro-dihydroquinones from a new marine actinomycete. J Nat Prod 2005, 68, 904–910. [Google Scholar]
- Soria-Mercado, IE; Jensen, PR; Fenical, W; Kassel, S; Golen, J. 3,4a-Dichloro-10a-(3-chloro-6-hydroxy-2,2,6-trimethylcyclohexylmethyl)-6,8-dihydroxy-2,2,7-trimethyl-3,4,4a,10a-tetrahydro-2H-benzo[g]chromene-5,10-dione. Acta Crystallogr 2004, E60, o1627–o1629. [Google Scholar]
- Gallagher, KA; Fenical, W; Jensen, PR. Hybrid isoprenoid secondary metabolite production in terrestrial and marine actinomycetes. Curr Opin Biotechnol 2010, 21, 794–800. [Google Scholar]
- Sakoulas, G; Eliopoulos, GM; Moellering, RC, Jr; Wennersten, C; Venkataraman, L; Novick, RP; Gold, HS. Accessory gene regulator (agr) locus in geographically diverse Staphylococcus aureus isolates with reduced susceptibility to vancomycin. Antimicrob Agents Chemother 2002, 46, 1492–1502. [Google Scholar]
- Farnaes, LLL. Novel Analogs and a Protein Target for the Napyradiomycins. PhD Thesis, University of California, San Diego, CA, USA. 2009. [Google Scholar]
- Motohashi, K; Sue, M; Furihata, K; Ito, S; Seto, H. Terpenoids produced by actinomycetes: Napyradiomycins from Streptomyces antimycoticus NT17. J Nat Prod 2008, 71, 595–601. [Google Scholar]
- Motohashi, K; Irie, K; Toda, T; Matsuo, Y; Kasai, H; Sue, M; Furihata, K; Seto, H. Studies on terpenoids produced by actinomycetes. 5-dimethylallylindole-3-carboxylic Acid and A80915G-8″-acid produced by marine-derived Streptomyces sp. MS239. J Antibiot (Tokyo) 2008, 61, 75–80. [Google Scholar]
- Winter, JM; Moffitt, MC; Zazopoulos, E; McAlpine, JB; Dorrestein, PC; Moore, BS. Molecular basis for chloronium-mediated meroterpene cyclization: Cloning, sequencing, and heterologous expression of the napyradiomycin biosynthetic gene cluster. J Biol Chem 2007, 282, 16362–16368. [Google Scholar]
- Snyder, SA; Tang, ZY; Gupta, R. Enantioselective total synthesis of (−)-Napyradiomycin A1 via asymmetric chlorination of an isolated olefin. J Am Chem Soc 2009, 131, 5744–5745. [Google Scholar]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 7th ed.; Approved Standard M7-A7; CLSI: Wayne, PA, USA, 2006; Volume 26, 2. [Google Scholar]
- Haste, NM; Perera, VR; Maloney, KN; Tran, DN; Jensen, P; Fenical, W; Nizet, V; Hensler, ME. Activity of the streptogramin antibiotic etamycin against methicillin-resistant Staphylococcus aureus. J Antibiot (Tokyo) 2010, 63, 219–224. [Google Scholar]


{kind=link}
{kind=link}
| Strain | Compound 1 | Compound 2 |
|---|---|---|
| S. aureus ATCC 29213 | 2 | 4 |
| MRSA-ATCC33591 a | 1–2 | 2 |
| NRS70 (N315) a | 1 | 2 |
| Sanger 252 a | 1–2 | 1 |
| NRS100 (COL) a | 1 | 2 |
| MRSA clinical bacteremia isolate c-44 a | 3 | 1.5 |
| MRSA clinical bacteremia isolate c-88 a | 1.5–3 | 1.5 |
| MRSA USA300 (UAMS1182) b | 1.5–3 | 1–3 |
| MRSA USA300 (TCH1516) b | 2 | 1–2 |
| VRSA (Michigan Isolate) | 1–2 | 2–4 |
| VRSA (Pennsylvania Isolate) | 1–2 | 4 |
| GISA (HIP5836) (New Jersey) c | 0.5 | 1 |
| GISA (PC-3) (New York) c | 0.5 | 1 |
| Hetero-GISA A5940 c | 1 | 4 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Haste, N.M.; Farnaes, L.; Perera, V.R.; Fenical, W.; Nizet, V.; Hensler, M.E. Bactericidal Kinetics of Marine-Derived Napyradiomycins against Contemporary Methicillin-Resistant Staphylococcus aureus. Mar. Drugs 2011, 9, 680-689. https://doi.org/10.3390/md9040680
Haste NM, Farnaes L, Perera VR, Fenical W, Nizet V, Hensler ME. Bactericidal Kinetics of Marine-Derived Napyradiomycins against Contemporary Methicillin-Resistant Staphylococcus aureus. Marine Drugs. 2011; 9(4):680-689. https://doi.org/10.3390/md9040680
Chicago/Turabian StyleHaste, Nina M., Lauge Farnaes, Varahenage R. Perera, William Fenical, Victor Nizet, and Mary E. Hensler. 2011. "Bactericidal Kinetics of Marine-Derived Napyradiomycins against Contemporary Methicillin-Resistant Staphylococcus aureus" Marine Drugs 9, no. 4: 680-689. https://doi.org/10.3390/md9040680
APA StyleHaste, N. M., Farnaes, L., Perera, V. R., Fenical, W., Nizet, V., & Hensler, M. E. (2011). Bactericidal Kinetics of Marine-Derived Napyradiomycins against Contemporary Methicillin-Resistant Staphylococcus aureus. Marine Drugs, 9(4), 680-689. https://doi.org/10.3390/md9040680