Polymer Networks Produced by Marine Diatoms in the Northern Adriatic Sea

Abstract
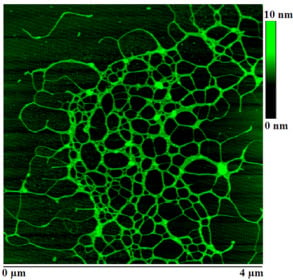
:
1. Introduction
2. Results and Discussion
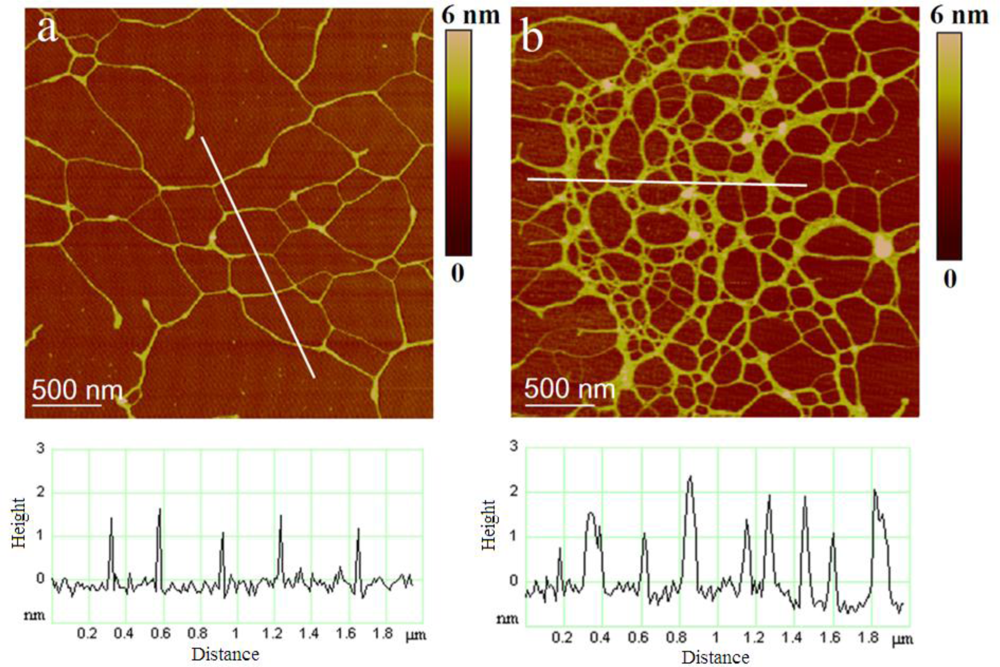
2.1. Extracellular Polymers Released by Cylindrotheca Closterium
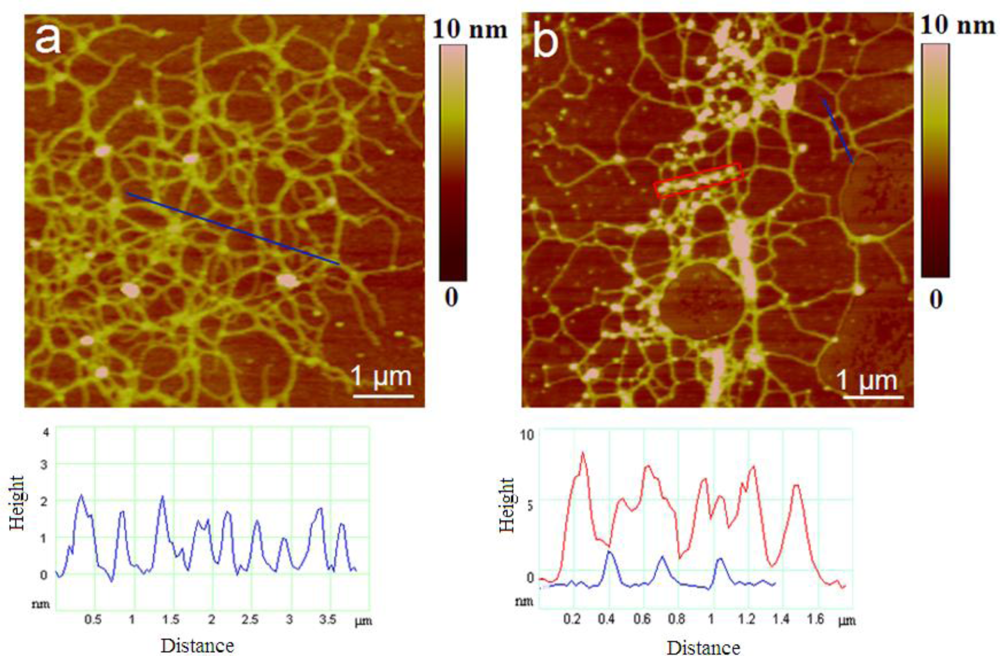
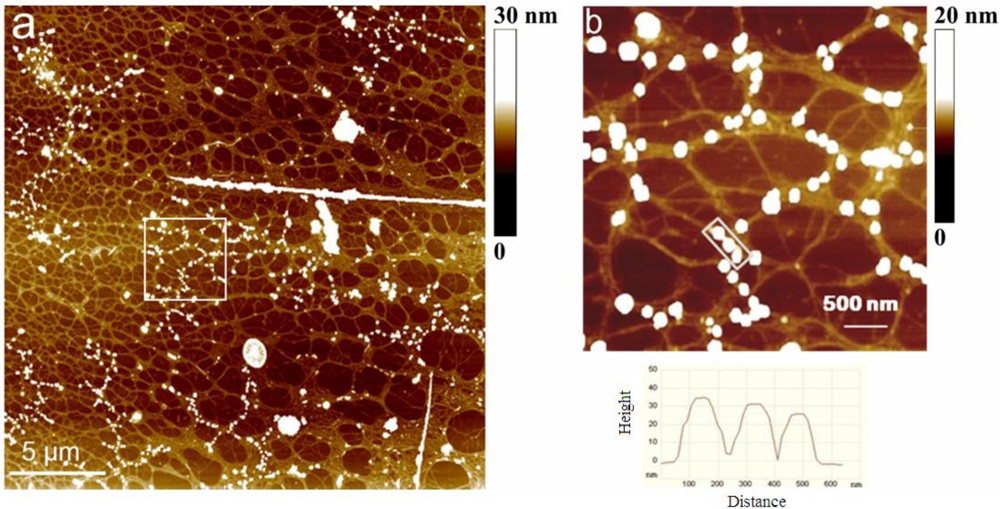
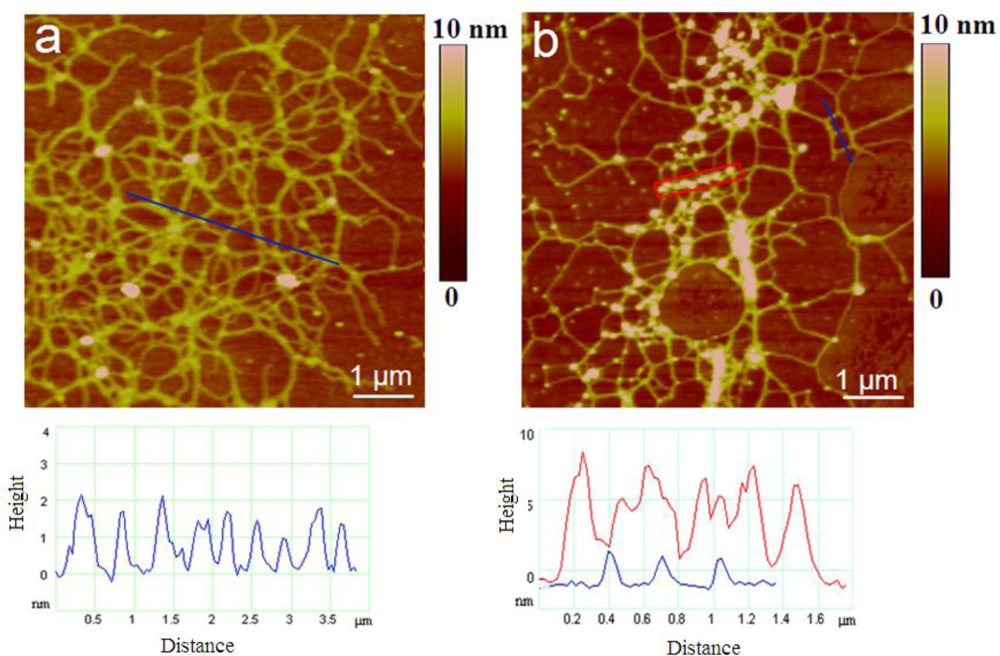
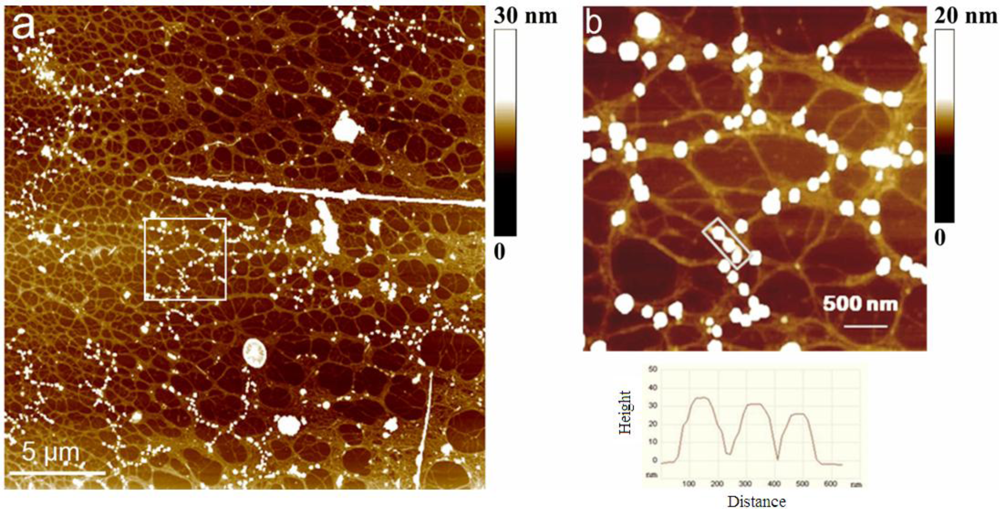
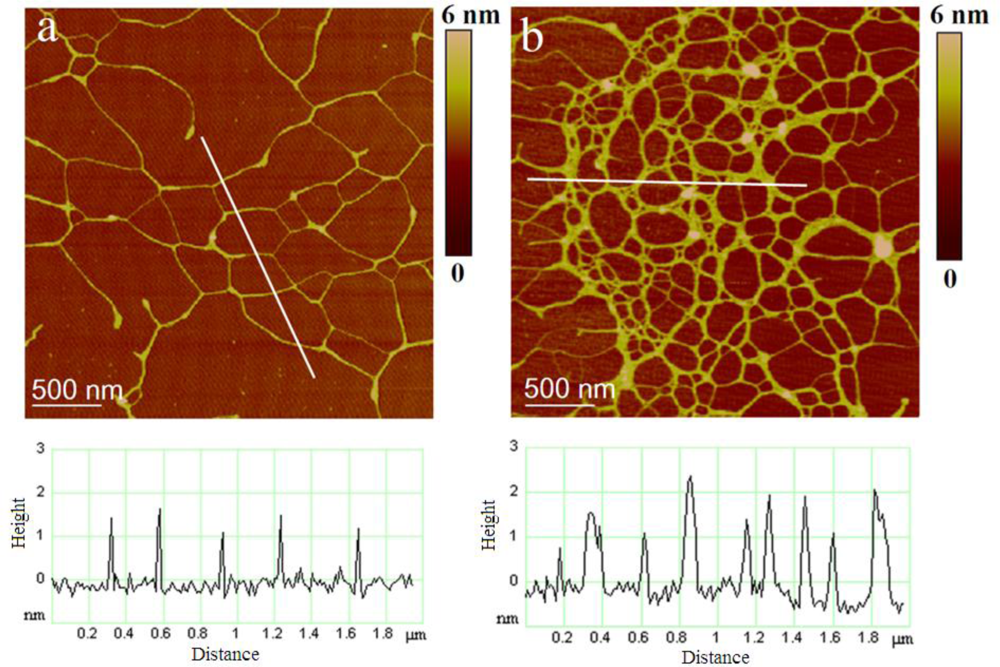
2.2. Polymer Networks of Marine Gel
2.3. Macromolecular Properties of the Isolated Polysaccharide Fraction of Marine Gel
2.4. Polysaccharide Fraction Isolated from Cell Culture
3. Experimental Section
3.1. Cell Culture
3.2. Macroscopic Gel Phase
3.3. AFM Imaging
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- CIESM, Production and Fate of Dissolved Organic Matter in the Mediterranean Sea; CIESM Workshop Monographs 28; CIESM: Cambados, Spain, 2006; pp. 5–10.
- Marine mucilages. In Science of the Total Environment; Vollenweider, R; Rinaldi, A (Eds.) Elsevier: Amsterdam, The Netherlands, 1995; Volume 165, pp. 1–235.
- Marine coastal eutrophication. In Science of the Total Environment Supplement; Vollenweider, RA; Marchetti, R; Viviani, R (Eds.) Elsevier: Amsterdam, The Netherlands, 1992; pp. 1–131.
- Pugnetti, A; Armeni, M; Camatti, E; Crevatin, E; Dell’Anno, A; Del Negro, P; Milandri, A; Socal, G; Fonda Umani, S; Danovaro, R. Imbalance between phytoplankton production and bacterial carbon demand in relation to mucilage formation in the Northern Adriatic Sea. Sci Total Environ 2005, 353, 162–177. [Google Scholar]
- The environmental impact (reduction in eutrophication) that would result from banning sodium tripolyphosphate (STPP) in household detergents. In Opinion of the Scientific Committee on Toxicity, Ecotoxicity and the Environment (CSTEE); European Commission Health & Consumer Protection Direcotorate-General: Brussels, Belgium, 2003.
- Fonda Umani, S; Milani, L; Borme, D; De Olazabal, A; Parlato, S; Precali, R; Kraus, R; Lučić, D; Njire, J; Totti, C; Romagnoli, T; Pompei, M; Cangini, M. Inter-annual variations of planktonic food webs in the northern Adriatic Sea. Sci Total Environ 2005, 353, 218–231. [Google Scholar]
- Precali, R; Giani, M; Marini, M; Grilli, F; Pečar, O; Paschini, E. Mucilaginous aggregates in the northern Adriatic in the period 1999–2002: Typology and distribution. Sci Total Environ 2005, 353, 10–23. [Google Scholar]
- Cozzi, S; Ivančić, I; Catalano, G; Đakovac, T; Degobbis, D. Dynamics of the oceanographic properties during mucilage appearance in the Northern Adriatic Sea: Analysis of the 1997 event in comparison to earlier events. J Mar Syst 2004, 50, 223–241. [Google Scholar]
- Penna, N; Kovač, N; Ricci, F; Penna, A; Capellacci, S; Faganeli, J. The role of dissolved carbohydrates in the northern Adriatic macroaggregate formation. Acta Chim Slov 2009, 56, 305–314. [Google Scholar]
- Najdek, M; Blažina, M; Đakovac, T; Kraus, R. The role of the diatom Cylindrotheca closterium in a mucilage event in the northern Adriatic Sea: Coupling with high salinity water intrusions. J Plankton Res 2005, 27, 851–862. [Google Scholar]
- Kovač, N; Mozetič, P; Trichet, J; Defarge, C. Phytoplankton composition and organic matter organization of mucous aggregates by means of light and cryo-scanning electron microscopy. Mar Biol 2005, 147, 261–271. [Google Scholar] [Green Version]
- Alcoverro, T; Conte, E; Mazzella, L. Production of mucilage by the adriatic epipelic diatom Cylindrotheca closterium (Bacillariophycea) under nutrient limitation. J Phycol 2000, 36, 1087–1095. [Google Scholar]
- Najdek, M; Degobbis, D; Mioković, D; Ivančić, I. Fatty acid and phytoplankton compositions of different types of mucilaginous aggregates in the northern Adriatic. J Plankton Res 2002, 24, 429–441. [Google Scholar]
- Del Negro, P; Crevatin, E; Larato, C; Ferrari, C; Totti, C; Pompei, M; Giani, M; Berto, D; Fonda Umani, S. Mucilage microcosms. Sci Total Environ 2005, 353, 258–269. [Google Scholar]
- Underwood, GJC; Peterson, DM. The importance of extracellular carbohydrate producton by marine epipelic diatoms. In Advances in Botanical Research; Callow, JA, Ed.; Elsevier; Academic Press: New York, NY, USA, 2003; pp. 184–240. [Google Scholar]
- Magaletti, E; Urbani, R; Sist, P; Ferrari, CR; Cicero, AM. Abundance and chemical characterization of extracellular carbohydrates released by the marine diatom Cylindrotheca fusiformis under N- and P-limitation. Eur J Phycol 2004, 39, 133–142. [Google Scholar]
- Urbani, R; Magaletti, E; Sist, P; Cicero, AM. Extracellular carbohydrates released by the marine diatoms Cylindrotheca closterium,Thalassiosira pseudonana and Skeletonema costatum: Effect of P-depletion and growth status. Sci Total Environ 2005, 353, 300–306. [Google Scholar]
- Bhaskar, PV; Bhosle, NB. Microbial extracellular polymeric substances in marine biogeochemical processes. Curr Sci 2005, 88, 45–53. [Google Scholar]
- Hoagland, KD; Rosowski, JR; Gretz, MR; Roemer, SC. Diatom extracellular polymeric substances: Function, fine structure, chemistry and physiology. J Phycol 1993, 29, 537–566. [Google Scholar]
- Wingender, J; Neu, TR; Flemming, H-C. What are bacterial extracellular polymeric substances? In Microbial Extracellular Polymeric Substances—Characterization, Structure and Function; Wingender, J, Neu, TR, Flemming, H-C, Eds.; Springer-Verlag: Heidelberg, Germany, 1999; pp. 1–20. [Google Scholar]
- Mišić Radić, T; Svetličić, V; Žutić, V; Boulgaropoulos, B. Seawater at the nanoscale: Marine gel imaged by atomic force microscopy. J Mol Recognit 2011, 24, 397–405. [Google Scholar]
- Pletikapić, G; Mišić Radić, T; Hozić Zimmermann, A; Svetličić, V; Pfannkuchen, M; Marić, D; Godrijan, J; Žutić, V. AFM imaging of extracellular polymer release by marine diatom Cylindrotheca closterium (Ehrenberg) Reiman & J.C. Lewin. J Mol Recognit 2011, 24, 436–445. [Google Scholar]
- Azam, F; Fonda-Umani, S; Funari, E. Significance of bacteria in the mucilage phenomenon in the northen Adriatic Sea. Ann Ist Super Sanità 1999, 35, 411–419. [Google Scholar]
- Faganeli, J; Mohar, B; Kofol, R; Pavlica, V; Marinšek, T; Rozman, A; Kovač, N; Šurca Vuk, A. Nature and lability of Northern Adriatic macroaggregates. Mar Drugs 2010, 8, 2480–2492. [Google Scholar]
- Morris, VJ; Kirby, AR; Gunning, AP. Atomic Force Microscopy for Biologists; Imperial College Press: London, UK, 1999; p. 332. [Google Scholar]
- Magonov, SN. Atomic force microscopy in analysis of polymers. In Encyclopedia of Analytical Chemistry; Meyers, RA, Ed.; John Wiley & Sons Ltd: Chichester, UK, 2000; pp. 7432–7491. [Google Scholar]
- Radotić, K; Mićić, M; Jeremić, M. Scanning probe microscopy of plant cell wall and its constituents. In Force Microscopy: Applications in Biology and Medicine; Jena, BP, Hoerber, JKH, Eds.; John Wiley & Sons Inc: New York, NY, USA, 2006; pp. 95–112. [Google Scholar]
- Schaeffer, TE. High-speed atomic force microscopy of biomolecules in motion. In Force Microscopy: Applications in Biology and Medicine; Jena, BP, Hoerber, JKH, Eds.; John Wiley & Sons: New York, NY, USA, 2006; pp. 221–248. [Google Scholar]
- Kirby, AR; Gunning, AP; Morris, VJ. Imaging polysaccharides by atomic force microscopy. Biopolymers 1996, 38, 355–366. [Google Scholar]
- Abu-Lail, NI; Camesano, TA. Polysaccharide properties probed with atomic force microscopy. J Microsc 2003, 212, 217–238. [Google Scholar]
- Abu-Lail, NI; Camesano, TA. Atomic force microscope and single-molecule force microscopy studies of biopolymers. In Dekker Encyclopedia of Nanoscience and Nanotechnology; Schwarz, J, Contescu, C, Putyera, K, Eds.; Taylor & Francis: London, UK, 2004; pp. 119–131. [Google Scholar]
- McIntire, TM; Brant, DA. Imaging of individual biopolymers and supramolecular assemblies using noncontact atomic force microscopy. Biopolymers 1997, 42, 133–146. [Google Scholar]
- Ikeda, S; Morris, VJ; Nishinari, K. Microstructure of aggregated and nonaggregated kappa-carrageenan helices visualized by atomic force microscopy. Biomacromolecules 2001, 2, 1331–1337. [Google Scholar]
- Keresztes, Z; Rigó, T; Telegdi, J; Kálmán, E. Investigation of biopolymer networks by means of AFM. Appl Phys A 2001, 72, S113–S116. [Google Scholar]
- Funami, T; Hiroe, M; Noda, S; Asai, I; Ikeda, S; Nishimari, K. Influence of molecular structure imaged with atomic force microscopy on the rheological behavior of carrageenan aqueous systems in the presence or absence of cations. Food Hydrocolloid 2007, 21, 617–629. [Google Scholar]
- Noda, S; Funami, T; Nakauma, M; Asai, I; Takahashi, R; Al-Assaf, S; Ikeda, S; Nishinari, K; Phillips, GO. Molecular structures of gellan gum imaged with atomic force microscopy in relation to the rheological behavior in aqueous systems. 1. Gellan gum with various acyl contents in the presence and absence of potassium. Food Hydrocolloid 2008, 22, 1148–1159. [Google Scholar]
- Balnois, E; Wilkinson, KJ. Sample preparation techniques for the observation of environmental biopolymers by atomic force microscopy. Colloid Surf A 2002, 207, 229–242. [Google Scholar]
- Shchipunov, YA; Kojima, A; ImaebI, T. Polysaccharides as a template for silicate generated by sol-gel processes. J Colloid Interface Sci 2005, 285, 574–580. [Google Scholar]
- Klaine, SJ; Alvarez, PJJ; Batley, GE; Fernandes, TF; Handy, RD; Lyon, DY; Mahendra, S; McLaughlin, MJ; Lead, JR. Nanomaterials in the environment: Behavior, fate, bioavailability, and effects. Environ Toxicol Chem 2008, 27, 1825–1851. [Google Scholar]
- Sumper, M; Kröger, N. Silica formation in diatoms: The function of long-chain polyamines and silaffins. J Mater Chem 2004, 14, 2059–2065. [Google Scholar]
- Hildebrand, M; Holton, G; Joy, DC; Doktycz, MJ; Allison, DP. Diverse and conserved nano- and mesoscale-structures of diatom silica revealed by atomic force microscopy. J Microsc 2009, 235, 172–187. [Google Scholar]
- Pletikapić, G; Berquand, A; Mišić Radić, T; Svetličić, V. Quantitative nanomechanical mapping of marine diatom in seawater using peak force tapping AFM. J Phycol 2011, in press. [Google Scholar]
- Smidsrød, O; Haug, A. Estimation of the relative stiffness of the molecular chain in polyelectrolytes from measurements of viscosity at different ionic strengths. Biopolymers 1971, 10, 1213–1227. [Google Scholar]
- Trzeinski, S; Varum, K; Staszewska, D; Smidsrød, O; Bohdanecký, M. Comparative studies on molecular chain parameters of chitosans and poly(diallyldimethylammonium chloride): The stiffness B-parameter and the temperature coefficient of intrinsic viscosity. Carbohydr Polym 2002, 48, 171–178. [Google Scholar]
- Huglin, M. Light Scattering from Polymer Solutions; Academic Press: London, UK, 1972; p. 885. [Google Scholar]
- Urbani, R; Brant, D. Kelco microbial polysaccharides S-130 (welan) and S-657 Display similar dilute aqueous solution behavior. Carbohydr Polym 1989, 11, 169–191. [Google Scholar]
- Urbani, R; Sist, P. Studio di caratterizzazione chimica delle componenti polisaccaridiche e del loro ruolo nel meccanismo di formazione degli aggregati gelatinosi. MAT Final Report. In ICRAM-Ministero dell’Ambiente e della Tutela del Territorio; ICRAM: Rome, Italy, 2003; pp. 135–170. [Google Scholar]
- Urbani, R; Cesaro, A. Chain conformations of polysaccharides in different solvents. In Handbook of Solvents; Wypych, G, Ed.; ChemTec Publishing: Toronto, Canada, 2001; pp. 706–735. [Google Scholar]
- Sartoni, G; Urbani, R; Sist, P; Berto, D; Nuccio, C; Giani, M. Benthic mucilaginous aggregates in the Mediterranean Sea: Origin, chemical composition and polysaccharide characterization. Mar Chem 2008, 111, 184–198. [Google Scholar]
- Verdugo, P; Alldredge, AL; Azam, F; Kirchman, DL; Passow, U; Santschi, PH. The oceanic gel phase: A bridge in the DOM-POM continuum. Mar Chem 2004, 92, 67–85. [Google Scholar]
- Verdugo, P; Orellana, MV; Chin, WC; Petersen, TW; Van den Eng, G; Benner, R; Hedges, JI. Marine biopolymer self-assembly: Implications for carbon cycling in the ocean. Faraday Discuss 2008, 139, 393–398. [Google Scholar]
- Ding, Y-X; Hung, C-C; Santaschi, PH; Verdugo, P; Chin, W-C. Spontaneouns assembly of exopolymers from phytoplankton. Terr Atmos Ocean Sci 2009, 20, 741–747. [Google Scholar]
- Guillard, RRL. Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Smith, WL, Chanley, MH, Eds.; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Stachowitsch, M; Fanuko, N; Richter, M. Mucous aggregates in the Adriatic Sea: An overview of stages and accourence. PSZNI: Mar Ecol 1990, 11, 327–350. [Google Scholar]
- Balnois, E; Papastavrou, G; Wilkinson, KJ. Force microscopy and force measurements of environmental colloids. In IUPAC Series on Analytical and Physical Chemistry of Environmental Systems Environmental Colloids and Particles—Behaviour, Separation and Characterization; Wilkinson, KJ, Lead, JR, Eds.; John Wiley & Sons: Chichester, UK, 2006; pp. 405–467. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharide fibrils | Number of fibrils analyzed | Fibril height/nm | |
|---|---|---|---|
| Mean value | Range | ||
| Attached to the diatom cell | 120 | 0.85 ± 0.32 | 0.4–1.8 |
| Marine gel network: early stage | 189 | 0.92 ± 0 .40 | 0.4–2.0 |
| Marine gel network: mature gel | 178 | 3.58 ± 0.76 | 1.6–5.0 |
| Sample | NaCl M | Mw kDa | A2×106 cm3 mol g−2 | RG nm |
|---|---|---|---|---|
| PS1 | 0.10 | 264 | −0.41 | 77 |
| 0.30 | 145 | −0.81 | 83 | |
| 0.50 | 218 | −1.3 | 99 | |
| 0.70 | 234 | −4.8 | 94 | |
| PS2 | 0.30 | 222 | −0.43 | 155 |
| 0.70 | 217 | −0.56 | 131 |
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Svetličić, V.; Žutić, V.; Radić, T.M.; Pletikapić, G.; Zimmermann, A.H.; Urbani, R. Polymer Networks Produced by Marine Diatoms in the Northern Adriatic Sea. Mar. Drugs 2011, 9, 666-679. https://doi.org/10.3390/md9040666
Svetličić V, Žutić V, Radić TM, Pletikapić G, Zimmermann AH, Urbani R. Polymer Networks Produced by Marine Diatoms in the Northern Adriatic Sea. Marine Drugs. 2011; 9(4):666-679. https://doi.org/10.3390/md9040666
Chicago/Turabian StyleSvetličić, Vesna, Vera Žutić, Tea Mišić Radić, Galja Pletikapić, Amela Hozić Zimmermann, and Ranieri Urbani. 2011. "Polymer Networks Produced by Marine Diatoms in the Northern Adriatic Sea" Marine Drugs 9, no. 4: 666-679. https://doi.org/10.3390/md9040666
APA StyleSvetličić, V., Žutić, V., Radić, T. M., Pletikapić, G., Zimmermann, A. H., & Urbani, R. (2011). Polymer Networks Produced by Marine Diatoms in the Northern Adriatic Sea. Marine Drugs, 9(4), 666-679. https://doi.org/10.3390/md9040666