
Marine Natural Product Inhibitors of Neutrophil-Associated Inflammation


Abstract
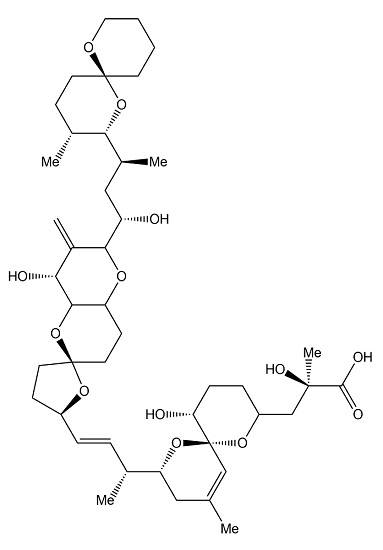
:
1. Introduction
2. Marine Natural Product Inhibitors of Formyl Peptide Receptor-1 (FPR-1) and Its Downstream Signaling Cascades
2.1. IA-LBI07-1
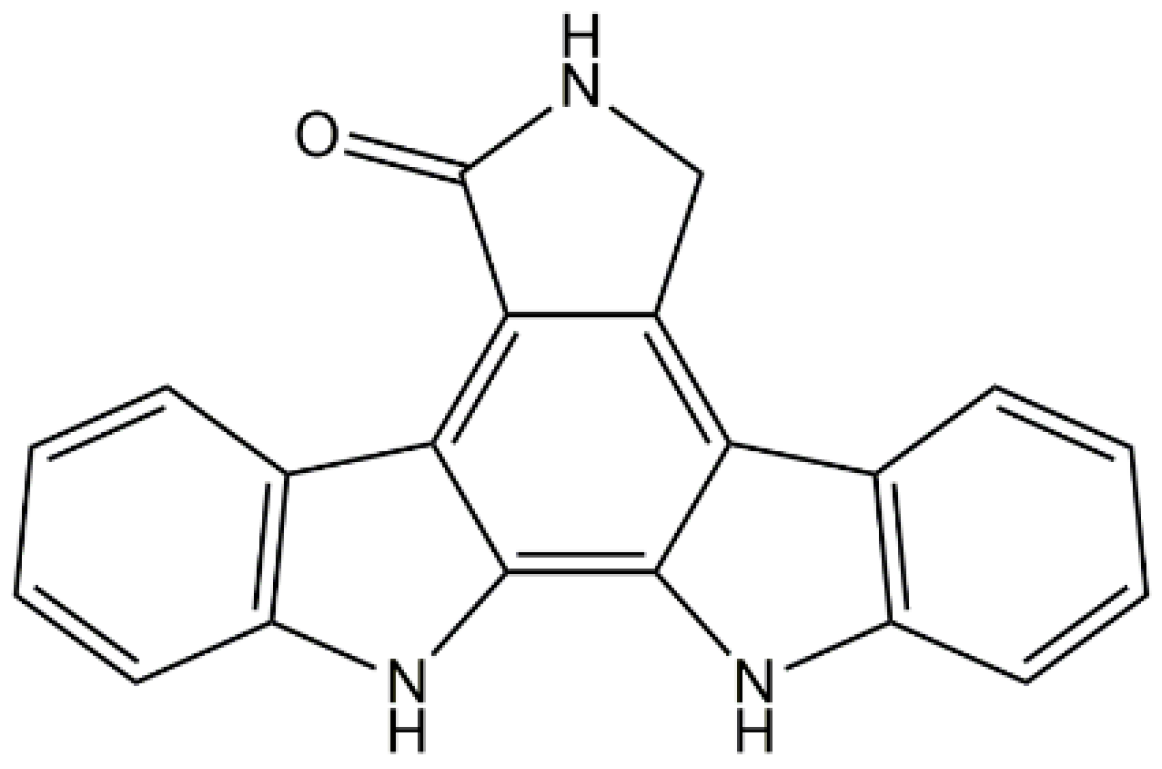
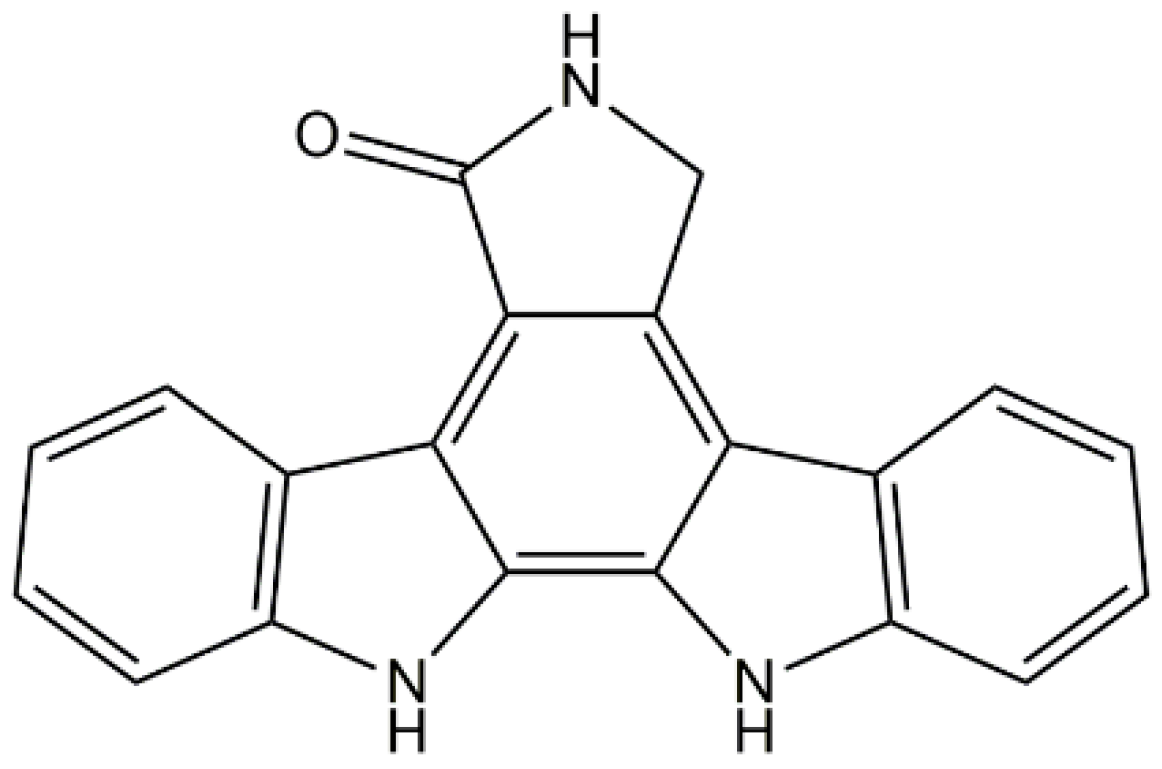
2.2. Staurosporine Aglycone
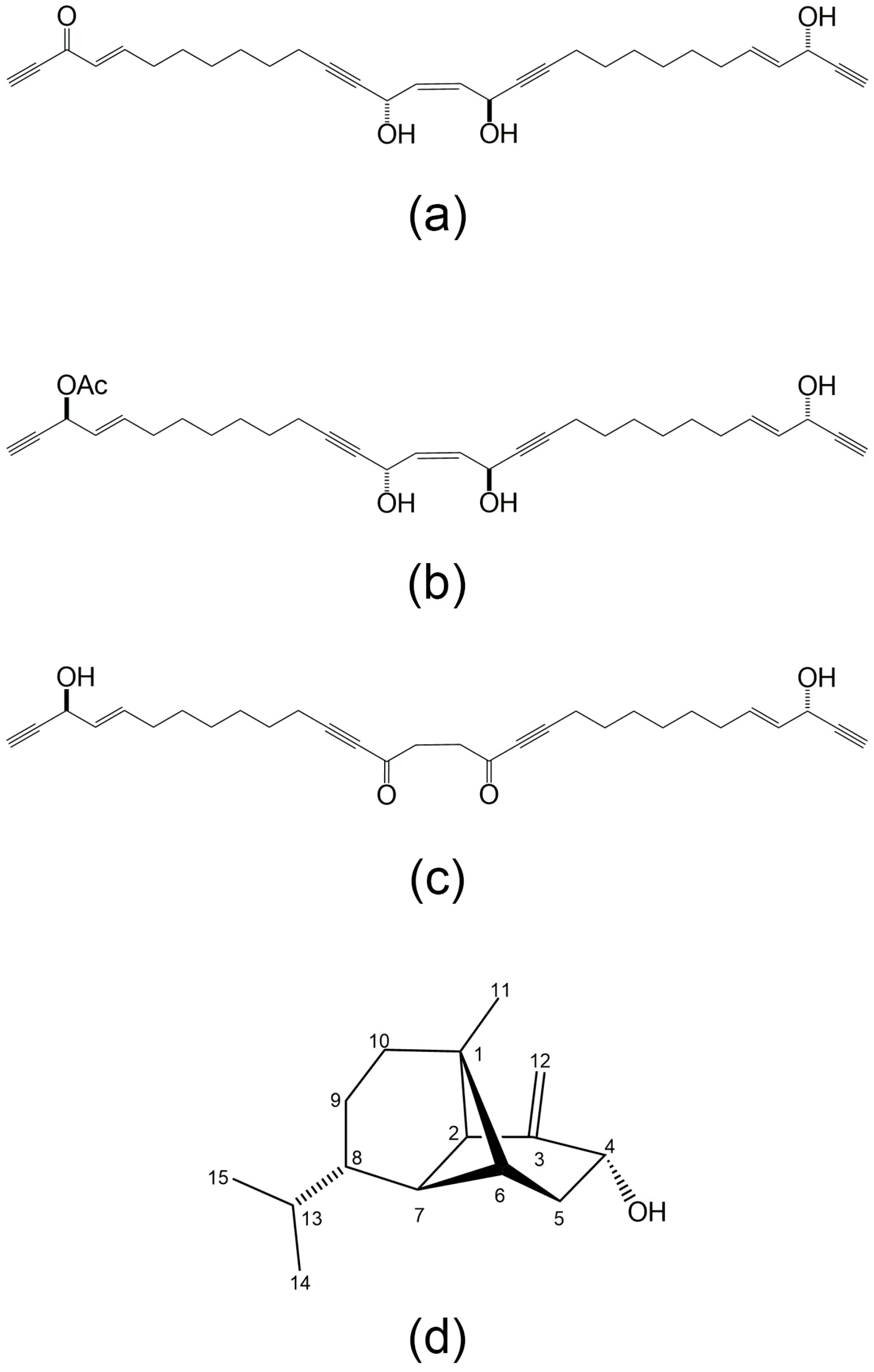
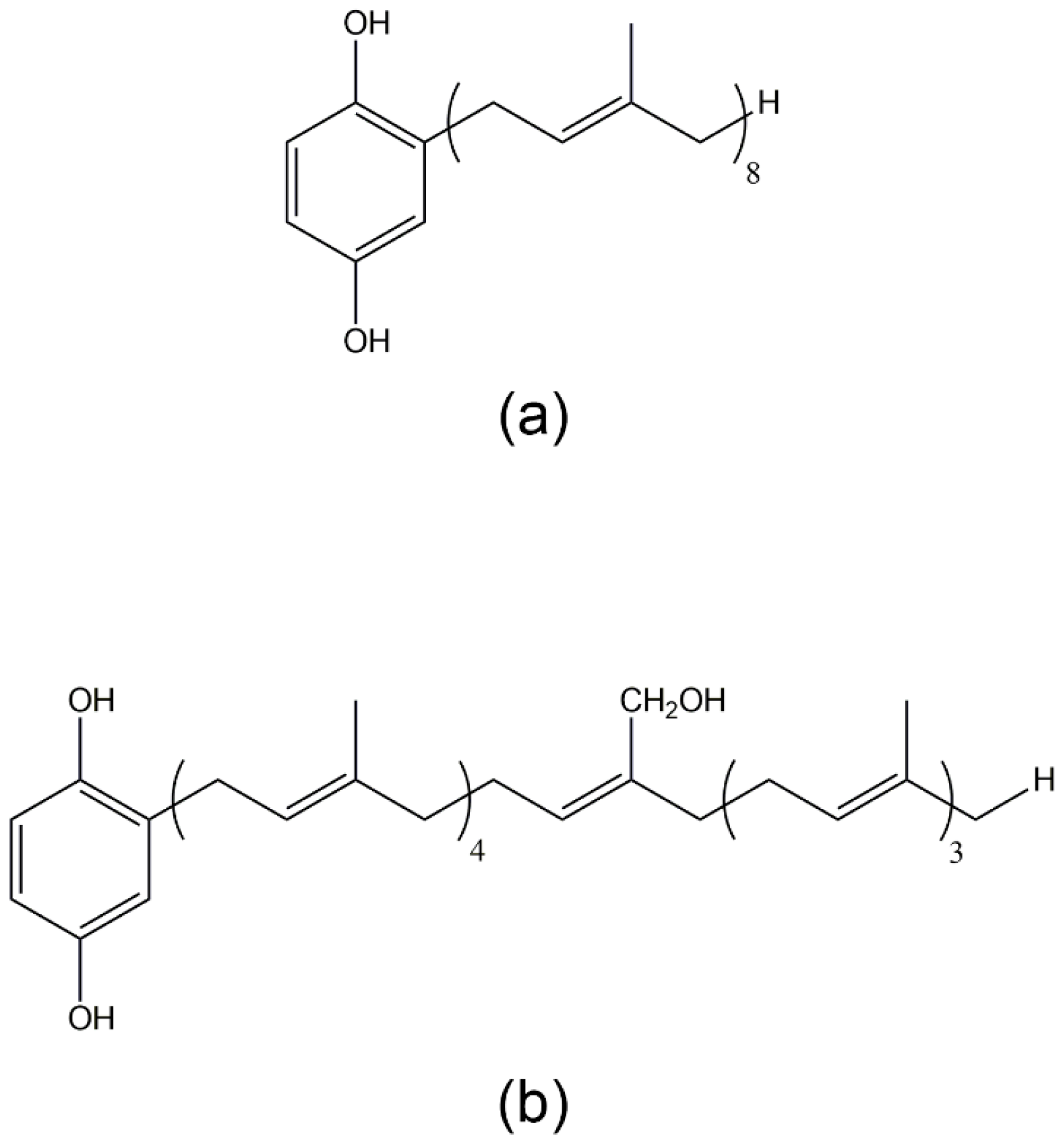
2.3. Secondary Metabolites of Marine Pseudomonas sp. (N11)
3. Marine Natural Product Inhibitors of Phospholipase A2 (PLA2) or Arachidonic Acid (AA) Metabolism
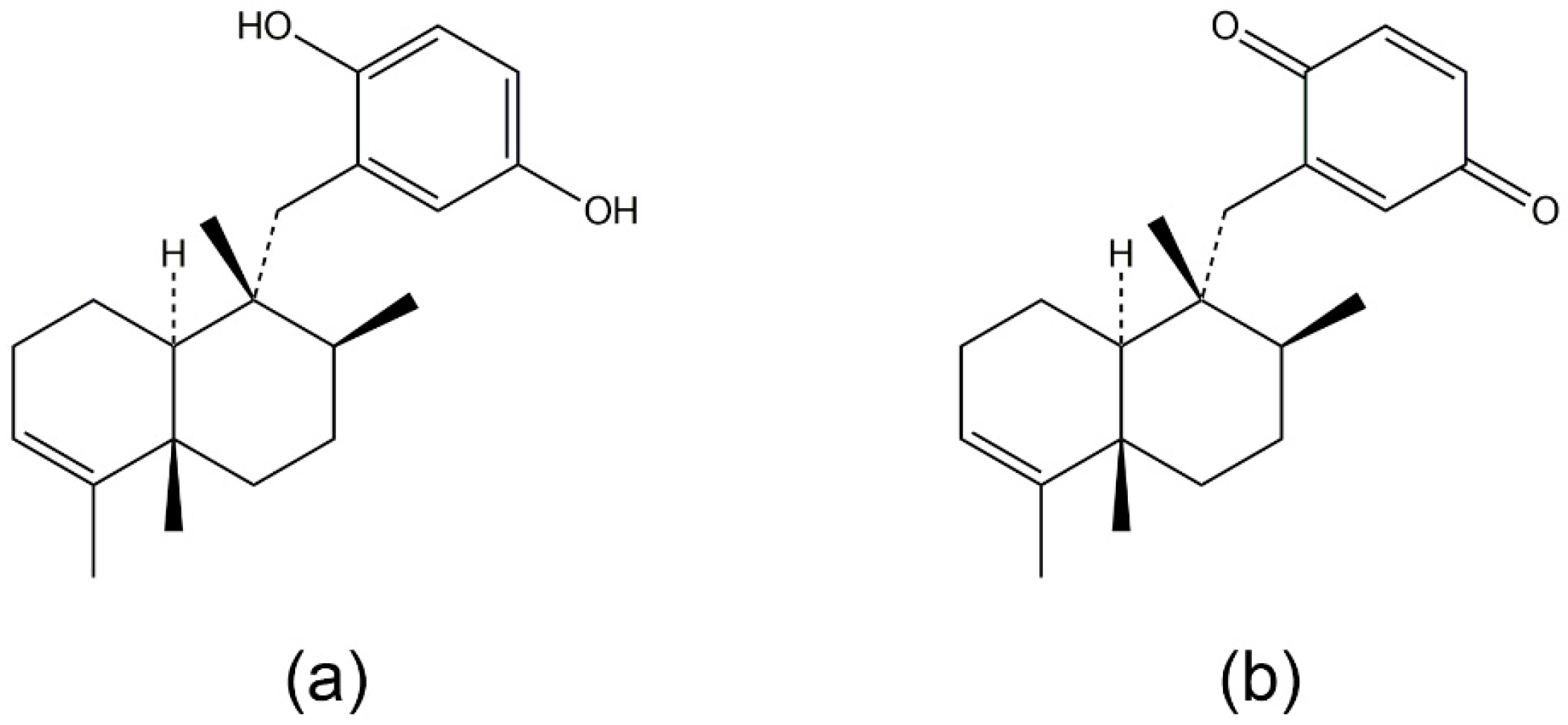
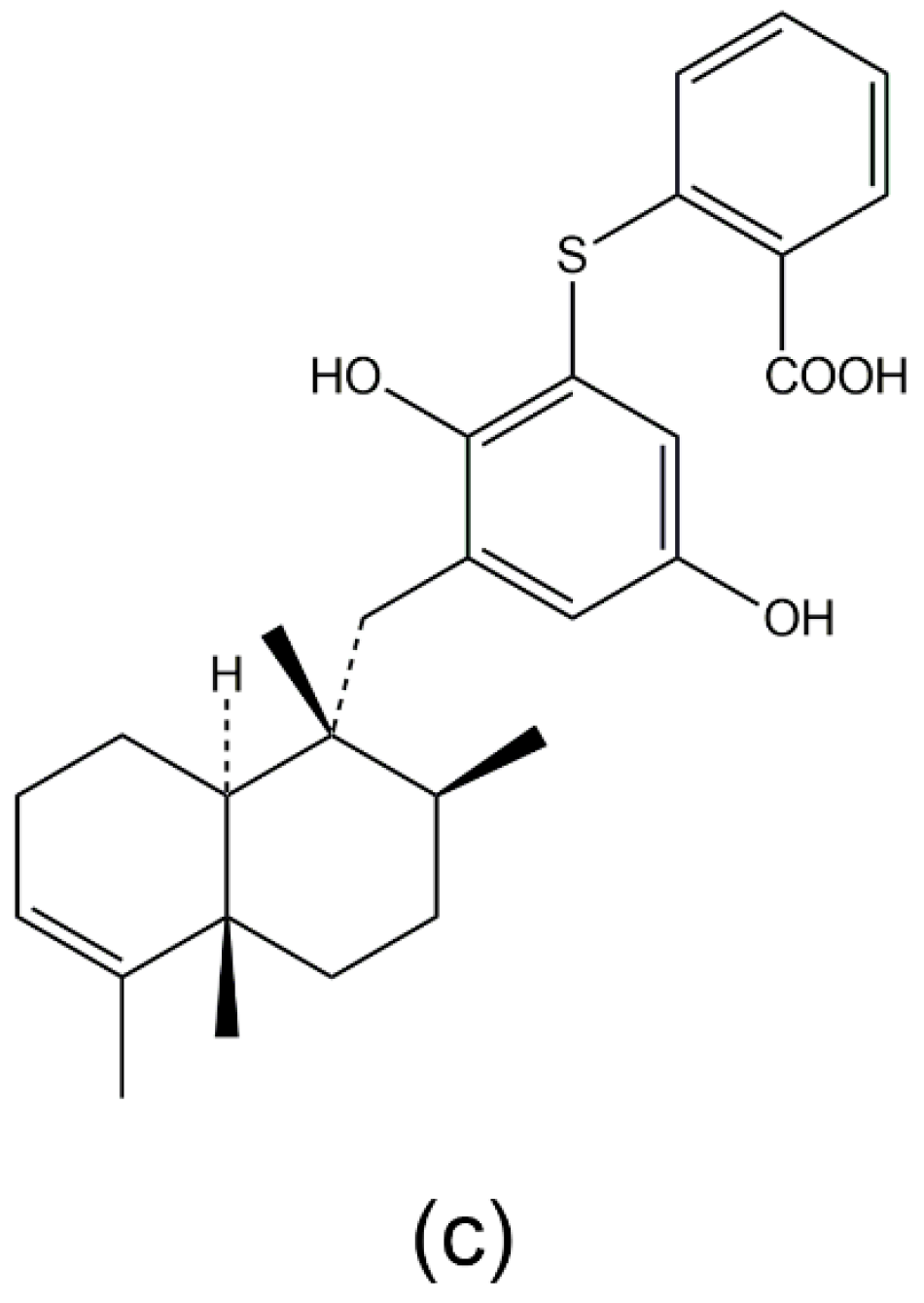
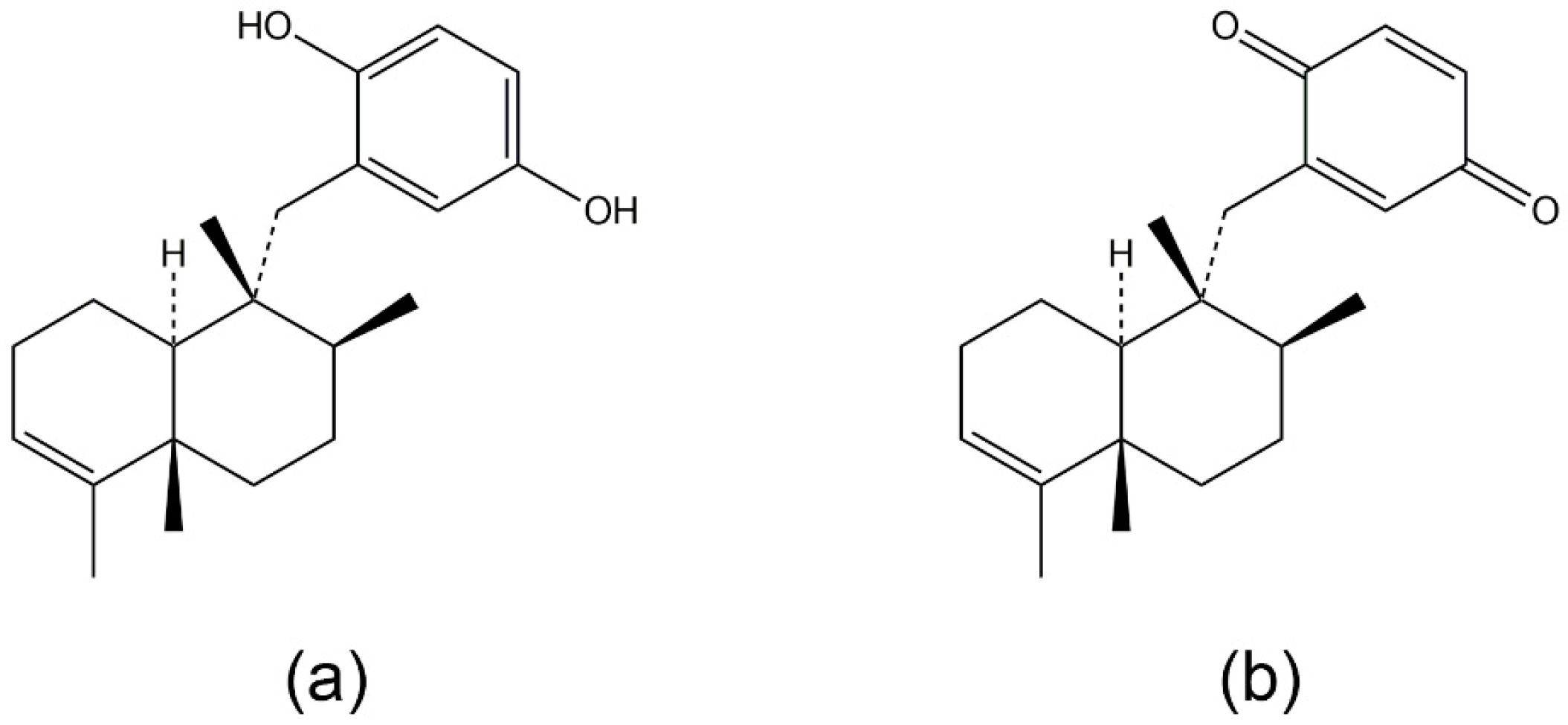
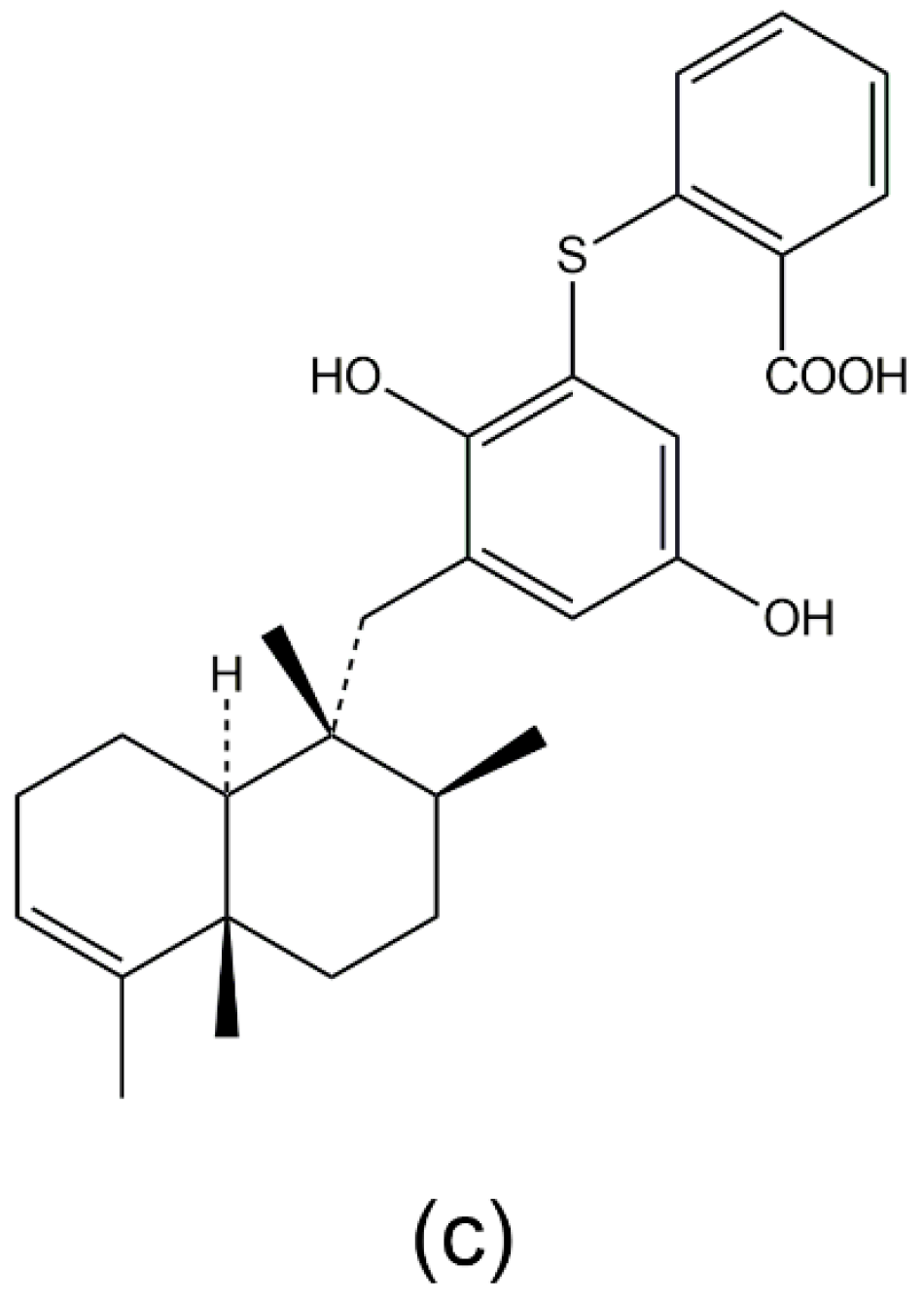
3.1. Avarol, Avarone, and Their Derivatives
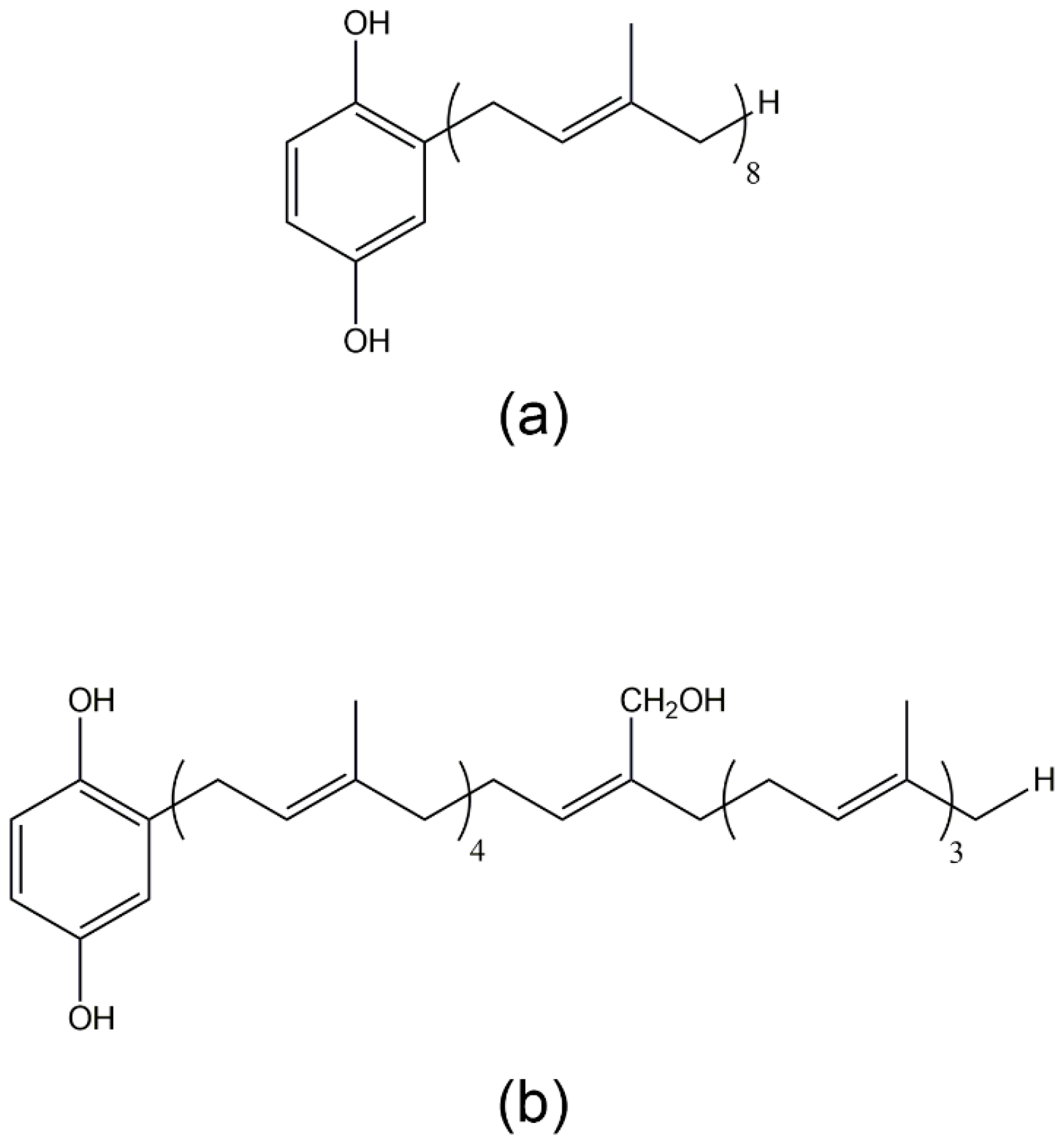
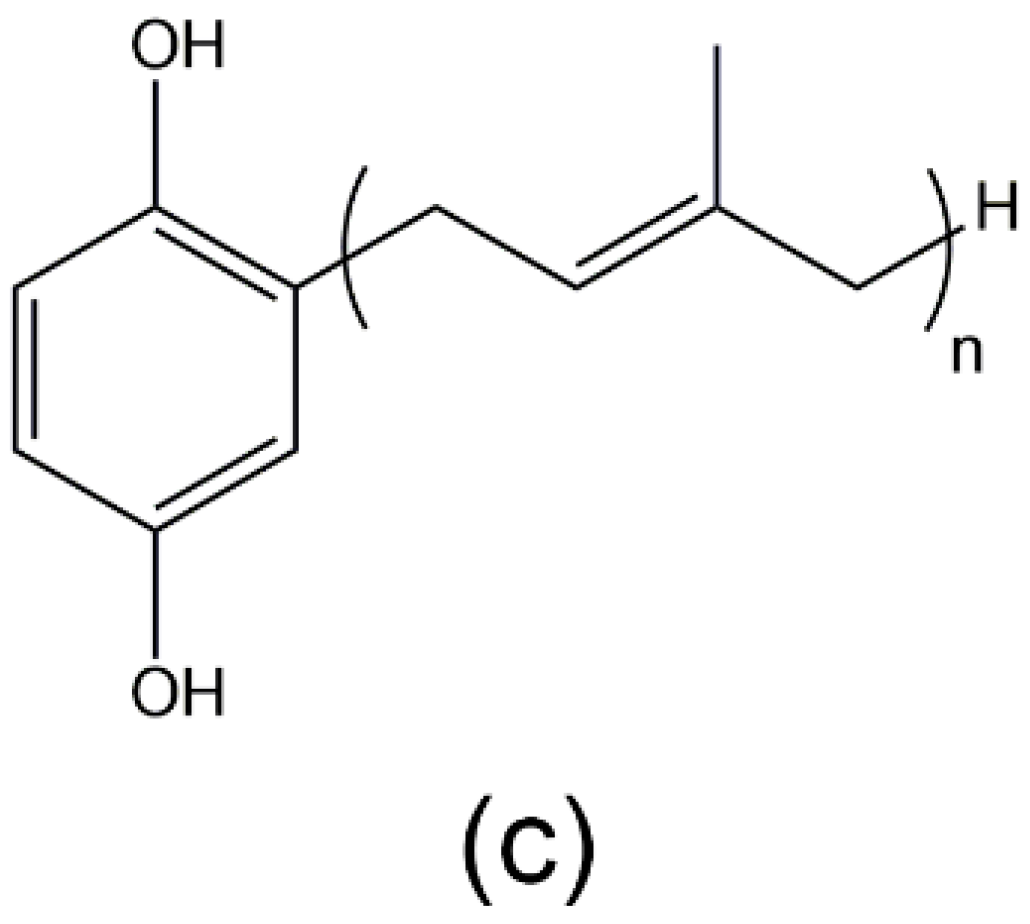
3.2. 2-Octaprenyl-1,4-hydroquinone, 2-[24-Hydroxy]-octaprenyl-1,4-hydroquinone, 2-Prenyl-1,4-hydroquinone, 2-Diprenyl-1,4-hydroquinone, 2-Triprenyl-1,4-hydroquinone, and 2-Tetraprenyl-1,4-hydroquinone
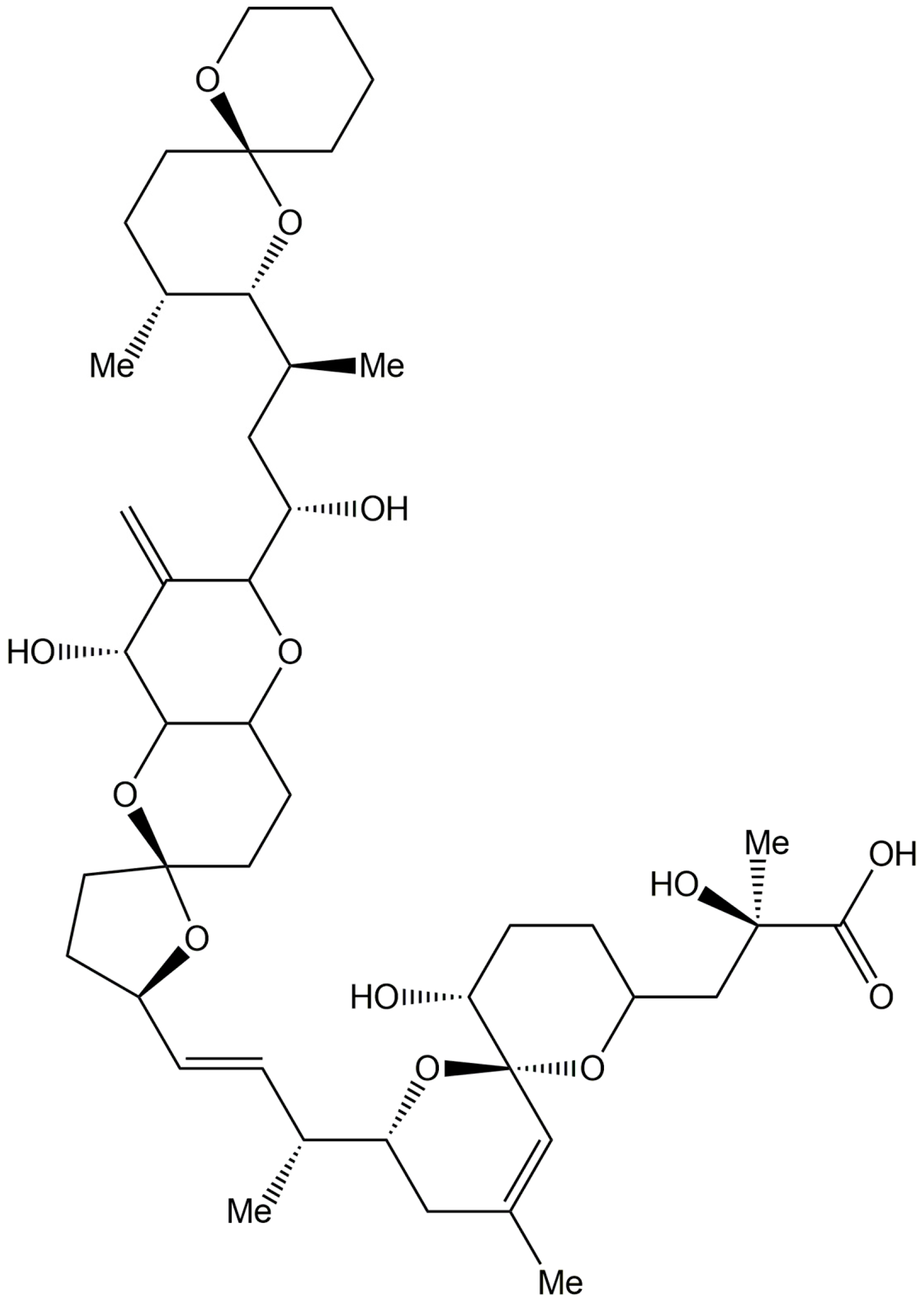
3.3. Lyprinol
3.4. Ircinin
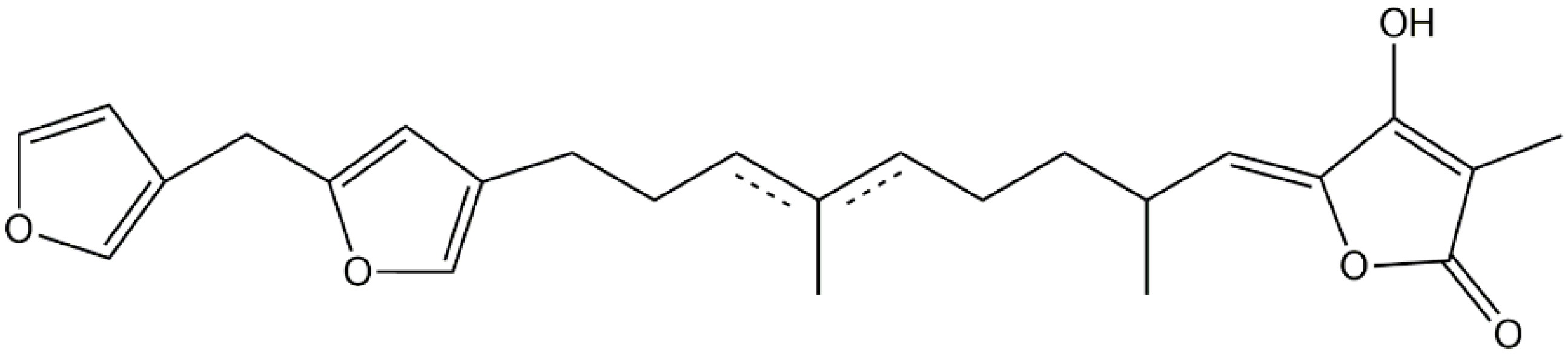
3.5. Dihydroxyicosanoids
4. Marine Natural Products that Regulate Phosphorylation of Intracellular Mediators
5. Marine Natural Product Inhibitors of Neutrophils Migration and Adhesion
5.1. Polysaccharides
5.2. Lectins
5.3. Miscellaneous
6. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ROS | reactive oxygen species |
| FPR | formyl peptide receptor |
| ALX | lipoxin A4 receptor |
| PI3K | phosphatidylinositol-3-kinase |
| MAPK | mitogen-activated kinase |
| PLC | phospholipase C |
| PIP2 | phosphatidylinositol 4,5-bisphosphate |
| IP3 | inositol 1,4,5-trisphosphate |
| PKC | protein kinase C |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| fMLF | formyl-methionyl-leucyl phenylalanine |
| JNK | c-Jun N-terminal kinase |
| ERK | extracellular signal-regulated kinase |
| PMA | phorbol myristate acetate |
| OKA | okadaic acid |
| PP | protein phosphatase |
| PLA2 | phospholipase A2 |
| AA | arachidonic acid |
| 5-LO | 5-lipoxygenase |
| COX | cyclooxygenase |
| 5-HPTETE | 5′-hydroperoxyeicosatetraenoic acid |
| 5-HETE | 5-hydroxyicosatetraenoic acid |
| PGH2 | prostaglandin H |
| LTl | leukotriene |
| TX | thromboxane |
| PLS | polysaccharide |
| Nitric oxide | NO |
| fCS | fucosylated chondroitin sulfate |
| MSU | monosodium urate |
| iNOS | nitric oxide synthase |
| NETs | neutrophil extracellular traps |
References
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Stapels, D.A.; Geisbrecht, B.V.; Rooijakkers, S.H. Neutrophil serine proteases in antibacterial defense. Curr. Opin. Microbiol. 2015, 23, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Hu, P.; Donskov, F.; Wang, G.; Liu, Q.; Du, J. Tumor-associated neutrophils as a new prognostic factor in cancer: A systematic review and meta-analysis. PLoS ONE 2014, 9, e98259. [Google Scholar] [CrossRef] [PubMed]
- Galdiero, M.R.; Bonavita, E.; Barajon, I.; Garlanda, C.; Mantovani, A.; Jaillon, S. Tumor associated macrophages and neutrophils in cancer. Immunobiology 2013, 218, 1402–1410. [Google Scholar] [CrossRef] [PubMed]
- Ciepiela, O.; Ostafin, M.; Demkow, U. Neutrophils in asthma—A review. Respir. Physiol. Neurobiol. 2015, 209, 13–16. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.; Kubes, P. Neutrophils and intravascular immunity in the liver during infection and sterile inflammation. Toxicol. Pathol. 2012, 40, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G.; Moutsopoulos, N.M.; Hajishengallis, E.; Chavakis, T. Immune and regulatory functions of neutrophils in inflammatory bone loss. Semin. Immunol. 2016, 28, 146–158. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M.; Brown, M.A. Innate immunity in the central nervous system. J. Clin. Investig. 2012, 122, 1164–1171. [Google Scholar] [CrossRef] [PubMed]
- Schepetkin, I.A.; Khlebnikov, A.I.; Kirpotina, L.N.; Quinn, M.T. Antagonism of human formyl peptide receptor 1 with natural compounds and their synthetic derivatives. Int. Immunopharmacol. 2015, 37, 43–58. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.K.; Seo, C.H.; Park, Y. Marine peptides and their anti-infective activities. Mar. Drugs 2015, 13, 618–654. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.; Rodriguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine pharmacology in 2009–2011: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [PubMed]
- Rabiet, M.J.; Huet, E.; Boulay, F. The N-formyl peptide receptors and the anaphylatoxin C5a receptors: An overview. Biochimie 2007, 89, 1089–1106. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.D.; Boulay, F.; Wang, J.M.; Dahlgren, C.; Gerard, C.; Parmentier, M.; Serhan, C.N.; Murphy, P.M. International Union of Basic and Clinical Pharmacology. LXXIII. Nomenclature for the formyl peptide receptor (FPR) family. Pharmacol. Rev. 2009, 61, 119–161. [Google Scholar] [CrossRef] [PubMed]
- Dufton, N.; Perretti, M. Therapeutic anti-inflammatory potential of formyl-peptide receptor agonists. Pharmacol. Ther. 2010, 127, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Lammers, K.M.; Chieppa, M.; Liu, L.; Liu, S.; Omatsu, T.; Janka-Junttila, M.; Casolaro, V.; Reinecker, H.C.; Parent, C.A.; Fasano, A. Gliadin Induces Neutrophil Migration via Engagement of the Formyl Peptide Receptor, FPR1. PLoS ONE 2015, 10, e0138338. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Raoof, M.; Chen, Y.; Sumi, Y.; Sursal, T.; Junger, W.; Brohi, K.; Itagaki, K.; Hauser, C.J. Circulating mitochondrial DAMPs cause inflammatory responses to injury. Nature 2010, 464, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Schiffmann, E.; Corcoran, B.A.; Wahl, S.M. N-formylmethionyl peptides as chemoattractants for leucocytes. Proc. Natl. Acad. Sci. USA 1975, 72, 1059–1062. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.C.; Sung, P.J.; Duh, C.Y.; Chen, B.W.; Sheu, J.H.; Yang, N.S. Anti-inflammatory activities of natural products isolated from soft corals of Taiwan between 2008 and 2012. Mar. Drugs 2013, 11, 4083–4126. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, P.C. Synthetic peptide chemotactic factors for neutrophils: The range of active peptides, their efficacy and inhibitory activity, and susceptibility of the cellular response to enzymes and bacterial toxins. Immunology 1979, 36, 579–588. [Google Scholar] [PubMed]
- Sung, P.J.; Chen, B.Y.; Lin, M.R.; Hwang, T.L.; Wang, W.H.; Sheu, J.H.; Wu, Y.C. Excavatoids E and F: Discovery of two new briaranes from the cultured octocoral Briareum excavatum. Mar. Drugs 2009, 7, 472–482. [Google Scholar] [CrossRef] [PubMed]
- Su, J.H.; Chen, B.Y.; Hwang, T.L.; Chen, Y.H.; Huang, I.C.; Lin, M.R.; Chen, J.J.; Fang, L.S.; Wang, W.H.; Li, J.J.; et al. Excavatoids L–N, new 12-hydroxybriaranes from the cultured octocoral Briareum excavatum (Briareidae). Chem. Pharm. Bull. 2010, 58, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.J.; Li, G.Y.; Su, Y.D.; Lin, M.R.; Chang, Y.C.; Kung, T.H.; Lin, C.S.; Chen, Y.H.; Su, J.H.; Lu, M.C.; et al. Excavatoids O and P, new 12-hydroxybriaranes from the octocoral Briareum excavatum. Mar. Drugs 2010, 8, 2639–2646. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Y.; Hwang, T.L.; Lin, M.R.; Chen, Y.H.; Chang, Y.C.; Fang, L.S.; Wang, W.H.; Wu, Y.C.; Sung, P.J. Carijoside A, a bioactive sterol glycoside from an octocoral Carijoa sp. (Clavulariidae). Mar. Drugs 2010, 8, 2014–2020. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.W.; Uvarani, C.; Huang, C.Y.; Hwang, T.L.; Dai, C.F.; Sheu, J.H. New anti-inflammatory tocopherol-derived metabolites from the Taiwanese soft coral Cladiella hirsuta. Bioorg. Med. Chem. Lett. 2015, 25, 92–95. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Z.; Chen, B.W.; Huang, C.Y.; Hwang, T.L.; Dai, C.F.; Sheu, J.H. Eunicellin-based diterpenoids, hirsutalins N-R, from the formosan soft coral Cladiella hirsuta. Mar. Drugs 2014, 12, 2446–2457. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Z.; Chen, B.W.; Huang, C.Y.; Hwang, T.L.; Uvarani, C.; Dai, C.F.; Sung, P.J.; Su, J.H.; Sheu, J.H. Eunicellin-Based Diterpenoids, Hirsutalins S–V, from the Formosan Soft Coral Cladiella hirsuta. Mar. Drugs 2015, 13, 2757–2769. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Tai, C.J.; Hwang, T.L.; Sheu, J.H. Krempfielins J–M, new eunicellin-based diterpenoids from the soft coral Cladiella krempfi. Mar. Drugs 2013, 11, 2741–2750. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.N.; Tai, C.J.; Hwang, T.L.; Sheu, J.H. Krempfielins N–P, New anti-inflammatory eunicellins from a Taiwanese soft coral Cladiella krempfi. Mar. Drugs 2014, 12, 1148–1156. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.J.; Chokkalingam, U.; Cheng, Y.; Shih, S.P.; Lu, M.C.; Su, J.H.; Hwang, T.L.; Sheu, J.H. Krempfielins Q and R, two new eunicellin-based diterpenoids from the soft coral Cladiella krempfi. Int. J. Mol. Sci. 2014, 15, 21865–21874. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Hwang, T.L.; Su, Y.D.; Chang, Y.C.; Chen, Y.H.; Hong, P.H.; Hu, L.C.; Yen, W.H.; Hsu, H.Y.; Huang, S.J.; et al. New 6-hydroxyeunicellins from a soft coral Cladiella sp. Chem. Pharm. Bull. 2012, 60, 160–163. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Tai, C.Y.; Hwang, T.L.; Weng, C.F.; Li, J.J.; Fang, L.S.; Wang, W.H.; Wu, Y.C.; Sung, P.J. Cladielloides A and B: New eunicellin-type diterpenoids from an Indonesian octocoral Cladiella sp. Mar. Drugs 2010, 8, 2936–2945. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Tai, C.Y.; Kuo, Y.H.; Kao, C.Y.; Li, J.J.; Hwang, T.L.; Fang, L.S.; Wang, W.H.; Sheu, J.H.; Sung, P.J. Cladieunicellins A–E, new eunicellins from an Indonesian soft coral Cladiella sp. Chem. Pharm. Bull. 2011, 59, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Tai, C.Y.; Hwang, T.L.; Sung, P.J. Cladieunicellin H, a new hemiketal eunicellin-based diterpenoid from the octocoral Cladiella sp. Nat. Prod. Commun. 2012, 7, 481–484. [Google Scholar] [PubMed]
- Chen, Y.H.; Tai, C.Y.; Su, Y.D.; Chang, Y.C.; Lu, M.C.; Weng, C.F.; Su, J.H.; Hwang, T.L.; Wu, Y.C.; Sung, P.J. Discovery of new eunicellins from an Indonesian octocoral Cladiella sp. Mar. Drugs 2011, 9, 934–943. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.M.; Hong, P.H.; Su, J.H.; Hwang, T.L.; Lu, M.C.; Fang, L.S.; Wu, Y.C.; Li, J.J.; Chen, J.J.; Wang, W.H.; et al. Bioactive compounds from a gorgonian coral Echinomuricea sp. (Plexauridae). Mar. Drugs 2012, 10, 1169–1179. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.H.; Chung, H.M.; Hwang, T.L.; Lu, M.C.; Wen, Z.H.; Kuo, Y.H.; Wang, W.H.; Sung, P.J. Echinoclerodane A: A new bioactive clerodane-type diterpenoid from a gorgonian coral Echinomuricea sp. Molecules 2012, 17, 9443–9450. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.M.; Hu, L.C.; Yen, W.H.; Su, J.H.; Lu, M.C.; Hwang, T.L.; Wang, W.H.; Sung, P.J. Echinohalimane A, a bioactive halimane-type diterpenoid from a Formosan gorgonian Echinomuricea sp. (Plexauridae). Mar. Drugs 2012, 10, 2246–2253. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.J.; Lin, M.R.; Chen, J.J.; Lin, S.F.; Wu, Y.C.; Hwang, T.L.; Fang, L.S. Hydroperoxysterols from the tunicate Eudistoma sp. Chem. Pharm. Bull. 2007, 55, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Solem, M.L.; Jiang, Z.D.; Gerwick, W.H. Three new and bioactive icosanoids from the temperate red marine alga Farlowia mollis. Lipids 1989, 24, 256–260. [Google Scholar] [CrossRef] [PubMed]
- Fang, H.Y.; Chokkalingam, U.; Chiou, S.F.; Hwang, T.L.; Chen, S.L.; Wang, W.L.; Sheu, J.H. Bioactive chemical constituents from the brown alga Homoeostrichus formosana. Int. J. Mol. Sci. 2015, 16, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Liaw, C.C.; Shen, Y.C.; Lin, Y.S.; Hwang, T.L.; Kuo, Y.H.; Khalil, A.T. Frajunolides E-K, briarane diterpenes from Junceella fragilis. J. Nat. Prod. 2008, 71, 1551–1556. [Google Scholar] [CrossRef] [PubMed]
- Sheu, J.H.; Chen, Y.P.; Hwang, T.L.; Chiang, M.Y.; Fang, L.S.; Sung, P.J. Junceellolides J-L, 11,20-epoxybriaranes from the gorgonian coral Junceella fragilis. J. Nat. Prod. 2006, 69, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.J.; Pai, C.H.; Hwang, T.L.; Fan, T.Y.; Su, J.H.; Chen, J.J.; Fang, L.S.; Wang, W.H.; Sheu, J.H. Junceols D-H, new polyoxygenated briaranes from sea whip gorgonian coral Junceella juncea (Ellisellidae). Chem. Pharm. Bull. 2008, 56, 1276–1281. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.C.; Chen, B.W.; Huang, C.Y.; Dai, C.F.; Hwang, T.L.; Sheu, J.H. Eunicellin-based diterpenoids from the Formosan soft coral Klyxum molle with inhibitory activity on superoxide generation and elastase release by neutrophils. J. Nat. Prod. 2013, 76, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.Y.; Su, J.H.; Lu, M.C.; Hwang, T.L.; Wang, W.H.; Chen, J.J.; Sheu, J.H.; Kuo, Y.H.; Weng, C.F.; Fang, L.S.; et al. Lobocrassins A-E: New cembrane-type diterpenoids from the soft coral Lobophytum crassum. Mar. Drugs 2011, 9, 1319–1331. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.Y.; Su, J.H.; Hwang, T.L.; Sheu, J.H.; Wen, Z.H.; Wu, Y.C.; Sung, P.J. Menelloides C and D, new sesquiterpenoids from the Gorgonian coral Menella sp. Mar. Drugs 2011, 9, 1534–1542. [Google Scholar] [CrossRef] [PubMed]
- Kao, S.Y.; Chang, Y.C.; Su, J.H.; Lu, M.C.; Chen, Y.H.; Sheu, J.H.; Wen, Z.H.; Wang, W.H.; Kuo, Y.H.; Hwang, T.L.; et al. (−)-Hydroxylindestrenolide, a new sesquiterpenoid from a gorgonian coral Menella sp. (Plexauridae). Chem. Pharm. Bull. 2011, 59, 1048–1050. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Kuo, J.; Su, J.H.; Hwang, T.L.; Chen, Y.H.; Lee, C.H.; Weng, C.F.; Sung, P.J. Pseudoalteromone B: A novel 15C compound from a marine bacterium Pseudoalteromonas sp. CGH2XX. Mar. Drugs 2012, 10, 1566–1571. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.M.; Wang, W.H.; Hwang, T.L.; Wu, Y.C.; Sung, P.J. Natural clovanes from the gorgonian coral Rumphella antipathies. Nat. Prod. Commun. 2013, 8, 1037–1040. [Google Scholar] [PubMed]
- Chung, H.M.; Wang, W.H.; Hwang, T.L.; Fang, L.S.; Wen, Z.H.; Chen, J.J.; Wu, Y.C.; Sung, P.J. Rumphellaoic acid A, a novel sesquiterpenoid from the formosan gorgonian coral Rumphella antipathies. Mar. Drugs 2014, 12, 5856–5863. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.M.; Wang, W.H.; Hwang, T.L.; Li, J.J.; Fang, L.S.; Wu, Y.C.; Sung, P.J. Rumphellaones B and C, new 4,5-seco-caryophyllane sesquiterpenoids from Rumphellan antipathies. Molecules 2014, 19, 12320–12327. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.M.; Wang, W.H.; Hwang, T.L.; Chen, J.J.; Fang, L.S.; Wen, Z.H.; Wang, Y.B.; Wu, Y.C.; Sung, P.J. Rumphellols A and B, new caryophyllene sesquiterpenoids from a Formosan gorgonian coral, Rumphella antipathies. Int. J. Mol. Sci. 2014, 15, 15679–15688. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Tseng, Y.J.; Chen, B.W.; Hwang, T.L.; Chen, H.Y.; Dai, C.F.; Sheu, J.H. Tortuosenes A and B, new diterpenoid metabolites from the Formosan soft coral Sarcophyton tortuosum. Org. Lett. 2014, 16, 1314–1317. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.H.; Dai, C.F.; Hwang, T.L.; Chen, C.Y.; Li, J.J.; Chen, J.J.; Wu, Y.C.; Sheu, J.H.; Wang, W.H.; Sung, P.J. Discovery of novel diterpenoids from Sinularia arborea. Mar. Drugs 2014, 12, 385–393. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.C.; Su, J.H.; Chiang, M.Y.; Lu, M.C.; Hwang, T.L.; Chen, Y.H.; Hu, W.P.; Lin, N.C.; Wang, W.H.; Fang, L.S.; et al. Flexibilins A–C, new cembrane-type diterpenoids from the Formosan soft coral, Sinularia flexibilis. Mar. Drugs 2013, 11, 1999–2012. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Lin, C.F.; Chang, W.Y.; Kuo, J.; Huang, Y.T.; Chung, P.J.; Hwang, T.L. Bioactive secondary metabolites of a marine Bacillus sp. inhibit superoxide generation and elastase release in human neutrophils by blocking formyl peptide receptor 1. Molecules 2013, 18, 6455–6468. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Iwai, Y.; Hirano, A.; Nakagawa, A.; Awaya, J.; Tsuchya, H.; Takahashi, Y.; Masuma, R. A new alkaloid AM-2282 OF Streptomyces origin. Taxonomy, fermentation, isolation and preliminary characterization. J. Antibiot. 1977, 30, 275–2782. [Google Scholar] [CrossRef] [PubMed]
- Ruegg, U.T.; Burgess, G.M. Staurosporine, K-252 and UCN-01: Potent but nonspecific inhibitors of protein kinases. Trends Pharmacol. Sci. 1989, 10, 218–220. [Google Scholar] [CrossRef]
- Horton, P.A.; Longley, R.E.; McConnell, O.J.; Ballas, L.M. Staurosporine aglycone (K252-c) and arcyriaflavin A from the marine ascidian, Eudistoma sp. Experientia 1994, 50, 843–845. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.C.; Sung, P.J.; Lin, C.F.; Kuo, J.; Chen, C.Y.; Hwang, T.L. Anti-inflammatory effects of secondary metabolites of marine Pseudomonas sp. in human neutrophils are through inhibiting P38 MAPK, JNK, and calcium pathways. PLoS ONE 2014, 9, e114761. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, J.P.; Lin, Y.; Lambeau, G.; Ghomashchi, F.; Lazdunski, M.; Gelb, M.H. Localization of structural elements of bee venom phospholipase A2 involved in N-type receptor binding and neurotoxicity. J. Biol. Chem. 1997, 272, 7173–7181. [Google Scholar] [CrossRef] [PubMed]
- Murakami, M.; Taketomi, Y.; Miki, Y.; Sato, H.; Hirabayashi, T.; Yamamoto, K. Recent progress in phospholipase A(2) research: From cells to animals to humans. Prog. Lipid Res. 2011, 50, 152–192. [Google Scholar] [CrossRef] [PubMed]
- Kramer, R.M.; Roberts, E.F.; Manetta, J.; Putnam, J.E. The Ca2(+)-sensitive cytosolic phospholipase A2 is a 100-kDa protein in human monoblast U937 cells. J. Biol. Chem. 1991, 266, 5268–5272. [Google Scholar] [PubMed]
- Quach, N.D.; Arnold, R.D.; Cummings, B.S. Secretory phospholipase A2 enzymes as pharmacological targets for treatment of disease. Biochem. Pharmacol. 2014, 90, 338–348. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.L.; Wartmann, M.; Lin, A.Y.; Knopf, J.L.; Seth, A.; Davis, R.J. cPLA2 is phosphorylated and activated by MAP kinase. Cell 1993, 72, 269–278. [Google Scholar] [CrossRef]
- Wijkander, J.; Sundler, R. Macrophage arachidonate-mobilizing phospholipase A2: Role of Ca2+ for membrane binding but not for catalytic activity. Biochem. Biophys. Res. Commun. 1992, 184, 118–124. [Google Scholar] [CrossRef]
- Levy, R. The role of cytosolic phospholipase A2-alfa in regulation of phagocytic functions. Biochim. Biophys. Acta 2006, 1761, 1323–1334. [Google Scholar] [CrossRef] [PubMed]
- Nalefski, E.A.; Sultzman, L.A.; Martin, D.M.; Kriz, R.W.; Towler, P.S.; Knopf, J.L.; Clark, J.D. Delineation of two functionally distinct domains of cytosolic phospholipase A2, a regulatory Ca(2+)-dependent lipid-binding domain and a Ca(2+)-independent catalytic domain. J. Biol. Chem. 1994, 269, 18239–18249. [Google Scholar] [PubMed]
- van Rossum, D.B.; Patterson, R.L. PKC and PLA2: Probing the complexities of the calcium network. Cell Calcium 2009, 45, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Folmer, F.; Jaspars, M.; Schumacher, M.; Dicato, M.; Diederich, M. Marine natural products targeting phospholipases A2. Biochem. Pharmacol. 2010, 80, 1793–1800. [Google Scholar] [CrossRef] [PubMed]
- Potts, B.C.; Faulkner, D.J.; Jacobs, R.S. Phospholipase A2 inhibitors from marine organisms. J. Nat. Prod. 1992, 55, 1701–1717. [Google Scholar] [CrossRef] [PubMed]
- Ferrandiz, M.L.; Sanz, M.J.; Bustos, G.; Paya, M.; Alcaraz, M.J.; De Rosa, S. Avarol and avarone, two new anti-inflammatory agents of marine origin. Eur. J. Pharmacol. 1994, 253, 75–82. [Google Scholar] [CrossRef]
- Muller, W.E.; Maidhof, A.; Zahn, R.K.; Schroder, H.C.; Gasic, M.J.; Heidemann, D.; Bernd, A.; Kurelec, B.; Eich, E.; Seibert, G. Potent antileukemic activity of the novel cytostatic agent avarone and its analogues in vitro and in vivo. Cancer Res. 1985, 45, 4822–4826. [Google Scholar] [PubMed]
- Amigo, M.; Paya, M.; De Rosa, S.; Terencio, M.C. Antipsoriatic effects of avarol-3′-thiosalicylate are mediated by inhibition of TNF-alpha generation and NF-kappaB activation in mouse skin. Br. J. Pharmacol. 2007, 152, 353–365. [Google Scholar] [CrossRef] [PubMed]
- Amigo, M.; Terencio, M.C.; Mitova, M.; Iodice, C.; Paya, M.; De Rosa, S. Potential antipsoriatic avarol derivatives as antioxidants and inhibitors of PGE(2) generation and proliferation in the HaCaT cell line. J. Nat. Prod. 2004, 67, 1459–1463. [Google Scholar] [CrossRef] [PubMed]
- Terencio, M.C.; Ferrandiz, M.L.; Posadas, I.; Roig, E.; de Rosa, S.; De Giulio, A.; Paya, M.; Alcaraz, M.J. Suppression of leukotriene B4 and tumour necrosis factor alpha release in acute inflammatory responses by novel prenylated hydroquinone derivatives. Naunyn-Schmiedeberg′s Arch. Pharmacol. 1998, 357, 565–572. [Google Scholar] [CrossRef]
- Gil, B.; Sanz, M.J.; Terencio, M.C.; De Giulio, A.; De Rosa, S.; Alcaraz, M.J.; Paya, M. Effects of marine 2-polyprenyl-1,4-hydroquinones on phospholipase A2 activity and some inflammatory responses. Eur. J. Pharmacol. 1995, 285, 281–288. [Google Scholar] [CrossRef]
- Halpern, G.M. Anti-inflammatory effects of a stabilized lipid extract of Perna canaliculus (Lyprinol). Allerg. Immunol. 2000, 32, 272–278. [Google Scholar]
- Cholbi, R.; Ferrandiz, M.L.; Terencio, M.C.; De Rosa, S.; Alcaraz, M.J.; Paya, M. Inhibition of phospholipase A2 activities and some inflammatory responses by the marine product ircinin. Naunyn-Schmiedeberg's Arch. Pharmacol. 1996, 354, 677–683. [Google Scholar] [CrossRef]
- Berkow, R.L.; Dodson, R.W. Regulation of neutrophil respiratory burst by protein phosphatases. Life Sci. 1993, 52, 1727–1732. [Google Scholar] [CrossRef]
- Garcia, R.C.; Whitaker, M.; Heyworth, P.G.; Segal, A.W. Okadaic acid produces changes in phosphorylation and translocation of proteins and in intracellular calcium in human neutrophils. Relationship with the activation of the NADPH oxidase by different stimuli. Biochem. J. 1992, 286 Pt 3, 687–692. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.J.; Takai, A.; Leto, T.L.; Grinstein, S. Modulation of neutrophil activation by okadaic acid, a protein phosphatase inhibitor. Am. J. Physiol. 1992, 262 Pt 1, C39–C49. [Google Scholar] [PubMed]
- Mayer, A.M.; Choudhry, M.A.; Sayeed, M.M.; Spitzer, J.A. The marine toxin okadaic acid reduces O2-generation and tyrosine phosphorylation in LPS-primed rat neutrophils. Life Sci. 1997, 61, PL199–PL204. [Google Scholar] [CrossRef]
- Shinozaki, T.; Watanabe, H.; Arita, S.; Chigira, M. Amino acid phosphatase activity of alkaline phosphatase. A possible role of protein phosphatase. Eur. J. Biochem. FEBS 1995, 227, 367–371. [Google Scholar] [CrossRef]
- Steinbeck, M.J.; Hegg, G.G.; Karnovsky, M.J. Arachidonate activation of the neutrophil NADPH-oxidase. Synergistic effects of protein phosphatase inhibitors compared with protein kinase activators. J. Biol. Chem. 1991, 266, 16336–16342. [Google Scholar] [PubMed]
- De Brito, T.V.; Prudencio Rda, S.; Sales, A.B.; Vieira, F., Jr.; Candeira, S.J.; Franco, A.X.; Aragao, K.S.; Ribeiro Rde, A.; Ponte de Souza, M.H.; Chaves Lde, S.; et al. Anti-inflammatory effect of a sulphated polysaccharide fraction extracted from the red algae Hypnea musciformis via the suppression of neutrophil migration by the nitric oxide signalling pathway. J. Pharm. Pharmacol. 2013, 65, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Chaves Lde, S.; Nicolau, L.A.; Silva, R.O.; Barros, F.C.; Freitas, A.L.; Aragao, K.S.; Ribeiro Rde, A.; Souza, M.H.; Barbosa, A.L.; Medeiros, J.V. Antiinflammatory and antinociceptive effects in mice of a sulfated polysaccharide fraction extracted from the marine red algae Gracilaria caudata. Immunopharmacol. Immunotoxicol. 2013, 35, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Pereira, J.G.; Mesquita, J.X.; Aragao, K.S.; Franco, A.X.; Souza, M.H.; Brito, T.V.; Dias, J.M.; Silva, R.O.; Medeiros, J.V.; Oliveira, J.S.; et al. Polysaccharides isolated from Digenea simplex inhibit inflammatory and nociceptive responses. Carbohydr. Polym. 2014, 108, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. Consorzio Interuniversitario Nazionale per la Bio-Oncologia, I. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic, and antiadhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [CrossRef] [PubMed]
- Panagos, C.G.; Thomson, D.S.; Moss, C.; Hughes, A.D.; Kelly, M.S.; Liu, Y.; Chai, W.; Venkatasamy, R.; Spina, D.; Page, C.P.; et al. Fucosylated chondroitin sulfates from the body wall of the sea cucumber Holothuria forskali: Conformation, selectin binding, and biological activity. J. Biol. Chem. 2014, 289, 28284–28298. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, J.G.; Bitencourt, F.S.; Cunha, T.M.; Luz, P.B.; Nascimento, K.S.; Mota, M.R.; Sampaio, A.H.; Cavada, B.S.; Cunha, F.Q.; Alencar, N.M. Agglutinin isolated from the red marine alga Hypnea cervicornis J. Agardh reduces inflammatory hypernociception: Involvement of nitric oxide. Pharmacol. Biochem. Behav. 2010, 96, 371–377. [Google Scholar] [CrossRef] [PubMed]
- Nagano, C.S.; Debray, H.; Nascimento, K.S.; Pinto, V.P.; Cavada, B.S.; Saker-Sampaio, S.; Farias, W.R.; Sampaio, A.H.; Calvete, J.J. HCA and HML isolated from the red marine algae Hypnea cervicornis and Hypnea musciformis define a novel lectin family. Protein Sci. 2005, 14, 2167–2176. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.M.; Lima, V.; Holanda, M.L.; Pinheiro, P.G.; Rodrigues, J.A.; Lima, M.E.; Benevides, N.M. Antinociceptive and anti-inflammatory activities of lectin from marine red alga Pterocladiella capillacea. Biol. Pharm. Bull. 2010, 33, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Vanderlei, E.S.; Patoilo, K.K.; Lima, N.A.; Lima, A.P.; Rodrigues, J.A.; Silva, L.M.; Lima, M.E.; Lima, V.; Benevides, N.M. Antinociceptive and anti-inflammatory activities of lectin from the marine green alga Caulerpa cupressoides. Int. Immunopharmacol. 2010, 10, 1113–1118. [Google Scholar] [CrossRef] [PubMed]
- Moura Rda, M.; Aragao, K.S.; de Melo, A.A.; Carneiro, R.F.; Osorio, C.B.; Luz, P.B.; de Queiroz, A.F.; Dos Santos, E.A.; de Alencar, N.M.; Cavada, B.S. Holothuria grisea agglutinin (HGA): The first invertebrate lectin with anti-inflammatory effects. Fundam. Clin. Pharmacol. 2013, 27, 656–668. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Ferreira, M.; Magalhaes, G.S.; Fernandez, J.H.; Junqueira-de-Azevedo Ide, L.; Le Ho, P.; Lima, C.; Valente, R.H.; Moura-da-Silva, A.M. Structural and biological characterization of Nattectin, a new C-type lectin from the venomous fish Thalassophryne nattereri. Biochimie 2011, 93, 971–980. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Mahmud, T.; Tajima, H.; Wang, W.; Aoki, S.; Nakagawa, S.; Mayumi, T.; Kitagawa, I. Marine natural products. XXXVI. Biologically active polyacetylenes, adociacetylenes A, B, C, and D, from an Okinawan Marine sponge of Adocia sp. Chem. Pharm. Bull. 1996, 44, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.P.; Huang, S.Y.; Lin, Y.Y.; Wang, H.M.; Jean, Y.H.; Wu, S.F.; Duh, C.Y.; Wen, Z.H. Soft coral-derived lemnalol alleviates monosodium urate-induced gouty arthritis in rats by inhibiting leukocyte infiltration and iNOS, COX-2 and c-Fos protein expression. Mar. Drugs 2013, 11, 99–113. [Google Scholar] [CrossRef] [PubMed]
- Bavington, C.D.; Lever, R.; Mulloy, B.; Grundy, M.M.; Page, C.P.; Richardson, N.V.; McKenzie, J.D. Anti-adhesive glycoproteins in echinoderm mucus secretions. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Vorobjeva, N.V.; Pinegin, B.V. Neutrophil extracellular traps: Mechanisms of formation and role in health and disease. Biochemistry 2014, 79, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Pinegin, B.; Vorobjeva, N.; Pinegin, V. Neutrophil extracellular traps and their role in the development of chronic inflammation and autoimmunity. Autoimmun. Rev. 2015, 14, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Barnado, A.; Crofford, L.J.; Oates, J.C. At the Bedside: Neutrophil extracellular traps (NETs) as targets for biomarkers and therapies in autoimmune diseases. J. Leukoc. Biol. 2016, 99, 265–278. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Menegazzo, L.; Scattolini, V.; Gintoli, M.; Albiero, M.; Avogaro, A. A perspective on NETosis in diabetes and cardiometabolic disorders. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liaw, P.C.; Ito, T.; Iba, T.; Thachil, J.; Zeerleder, S. DAMP and DIC: The role of extracellular DNA and DNA-binding proteins in the pathogenesis of DIC. Blood Rev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Warnatsch, A.; Ioannou, M.; Wang, Q.; Papayannopoulos, V. Inflammation. Neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science 2015, 349, 316–320. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Sources | Activities * | Ref. |
|---|---|---|---|
| Excavatoid E | Briareum excavatum | E | [20] |
| Excavatoid F | Briareum excavatum | E | [20] |
| Excavatoid L | Briareum excavatum | ROS, E | [21] |
| Excavatoid O | Briareum excavatum | ROS, E | [22] |
| Excavatoid P | Briareum excavatum | ROS, E | [22] |
| Carijoside A | Carijoa sp. | ROS, E | [23] |
| Hirsutocospiro A | Cladiella hirsuta | ROS, E | [24] |
| Hirsutalin N | Cladiella hirsuta | E | [25] |
| Hirsutalin S | Cladiella hirsuta | ROS, E | [26] |
| Krempfielin K | Cladiella krempfi | E | [27] |
| Krempfielin M | Cladiella krempfi | E | [27] |
| Krempfielin N | Cladiella krempfi | E | [28] |
| Krempfielin P | Cladiella krempfi | ROS, E | [28] |
| Krempfielin Q | Cladiella krempfi | ROS, E | [29] |
| Krempfielin R | Cladiella krempfi | ROS, E | [29] |
| 6-epi-cladieunicellin F | Cladiella sp. | ROS, E | [30] |
| Cladielloide B | Cladiella sp. | ROS, E | [31] |
| Cladieunicellin C | Cladiella sp. | ROS | [32] |
| Cladieunicellin H | Cladiella sp. | ROS, E | [33] |
| (–)-solenopodin C | Cladiella sp. | ROS, E | [34] |
| 6-epi-Yonarasterol B | Echinomuricea sp. | ROS, E | [35] |
| Echinoclerodane A | Echinomuricea sp. | ROS, E | [36] |
| Echinohalimane A | Echinomuricea sp. | E | [37] |
| 7β-Hydroperoxycholesterol | Eudistoma sp. | E | [38] |
| Dihydroxyicosanoids | Farlowia mollis | ROS, E | [39] |
| Methylfarnesylquinone | Homoeostrichus formosana | ROS, E | [40] |
| Frajunolide E | Junceella fragilis | ROS, E | [41] |
| Frajunolide J | Junceella fragilis | ROS, E | [41] |
| Junceellolide K | Junceella fragilis | E | [42] |
| (−)-11β,20β-Epoxy-4-deacetoxyjunceellolide D | Junceella fragilis | E | [42] |
| Junceol E | Junceella juncea | ROS | [43] |
| Junceol F | Junceella juncea | ROS | [43] |
| Junceol G | Junceella juncea | ROS | [43] |
| Junceol H | Junceella juncea | ROS | [43] |
| Klymollin M | Klyxum molle | ROS, E | [44] |
| Lobocrassin B | Lobophytum crassum | ROS, E | [45] |
| Menelloide D | Menella sp. | E | [46] |
| (−)-Hydroxylindestrenolide | Menella sp. | ROS | [47] |
| Pseudoalteromone B | Pseudoalteromonas sp. | E | [48] |
| 2β-Acetoxyclovan-9α-ol | Rumphella antipathies | ROS, E | [49] |
| 9α-Acetoxyclovan-2β-ol | Rumphella antipathies | ROS, E | [49] |
| Rumphellaoic acid A | Rumphella antipathies | E | [50] |
| Rumphellaone C | Rumphella antipathies | ROS, E | [51] |
| Rumphellol A | Rumphella antipathies | ROS, E | [52] |
| Rumphellol B | Rumphella antipathies | ROS, E | [52] |
| Tortuosene A | Sarcophyton tortuosum. | ROS | [53] |
| Sinularbol B | Sinularia arborea | ROS | [54] |
| Flexibilin B | Sinularia flexibilis | E | [55] |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, C.-Y.; Tsai, Y.-F.; Chang, W.-Y.; Yang, S.-C.; Hwang, T.-L. Marine Natural Product Inhibitors of Neutrophil-Associated Inflammation. Mar. Drugs 2016, 14, 141. https://doi.org/10.3390/md14080141
Chen C-Y, Tsai Y-F, Chang W-Y, Yang S-C, Hwang T-L. Marine Natural Product Inhibitors of Neutrophil-Associated Inflammation. Marine Drugs. 2016; 14(8):141. https://doi.org/10.3390/md14080141
Chicago/Turabian StyleChen, Chun-Yu, Yung-Fong Tsai, Wen-Yi Chang, Shun-Chin Yang, and Tsong-Long Hwang. 2016. "Marine Natural Product Inhibitors of Neutrophil-Associated Inflammation" Marine Drugs 14, no. 8: 141. https://doi.org/10.3390/md14080141
APA StyleChen, C.-Y., Tsai, Y.-F., Chang, W.-Y., Yang, S.-C., & Hwang, T.-L. (2016). Marine Natural Product Inhibitors of Neutrophil-Associated Inflammation. Marine Drugs, 14(8), 141. https://doi.org/10.3390/md14080141