Ricin Detection Using Phage Displayed Single Domain Antibodies


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
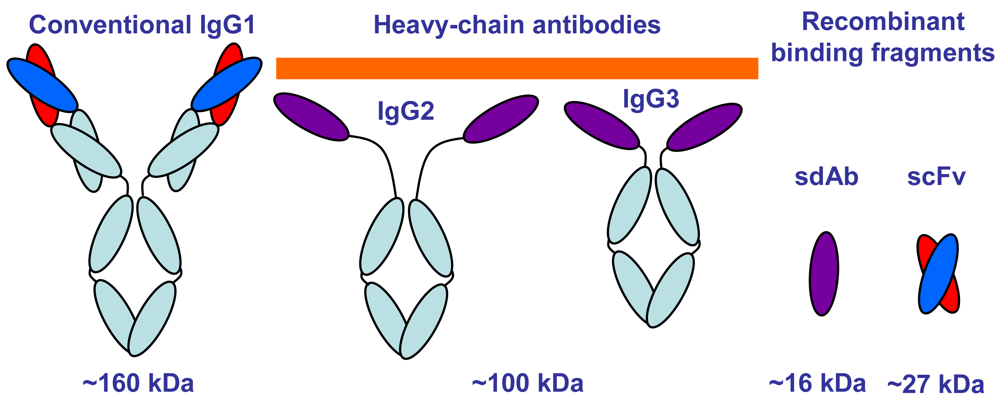
:1. Introduction
2. Results and Discussion
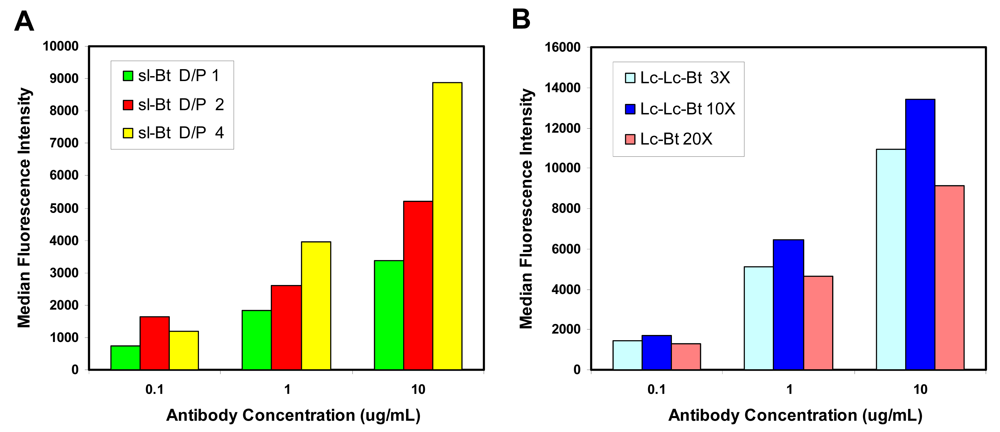
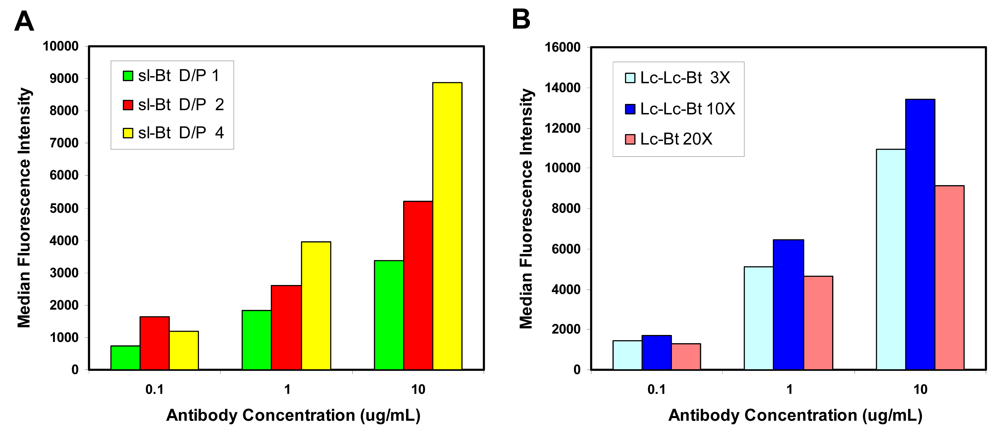
2.1. Evaluation of biotinylation reagents
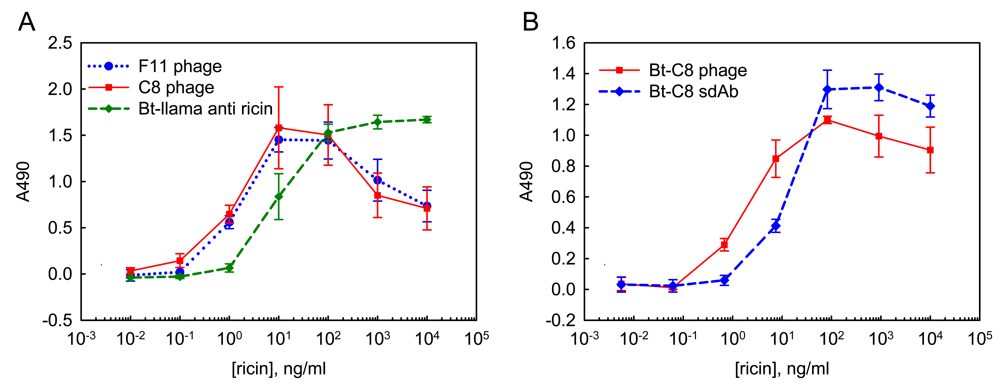
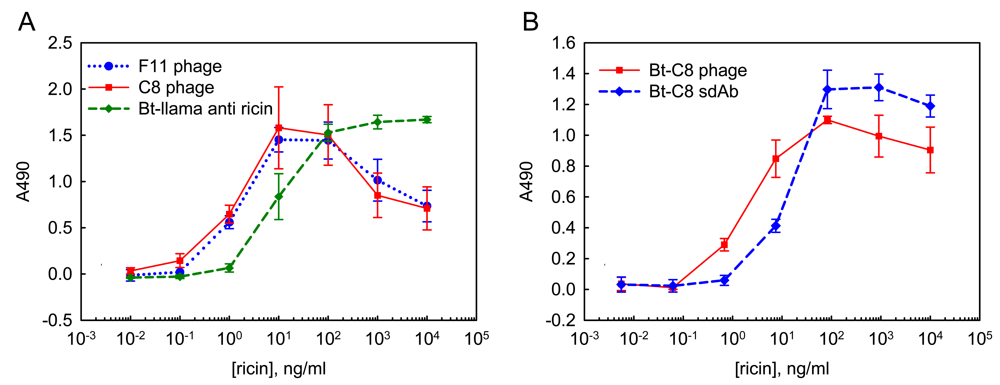
2.2. ELISA using soluble sdAb and phage-displayed sdAb
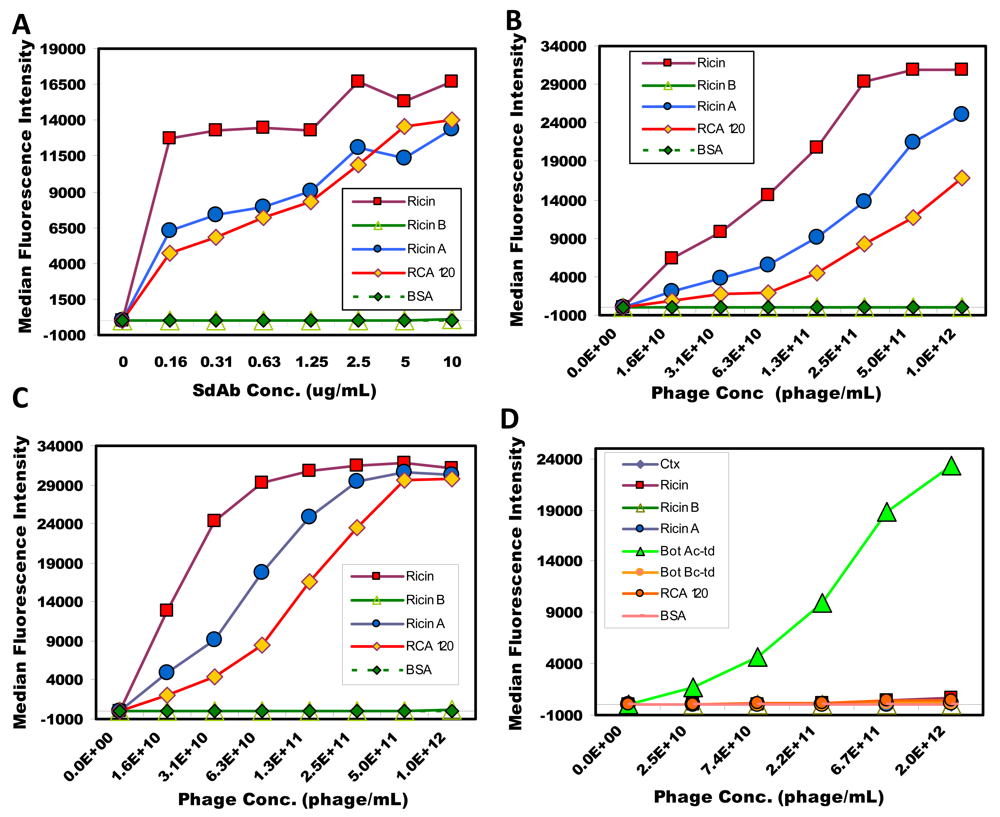
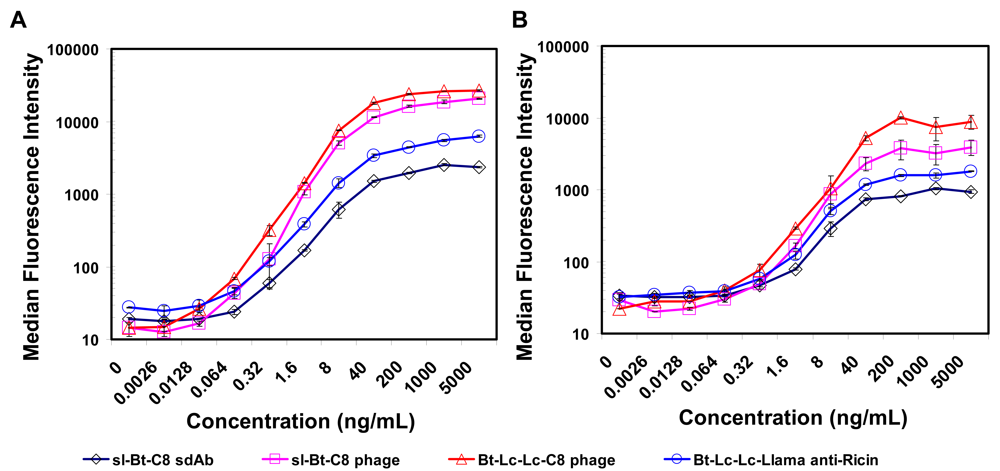
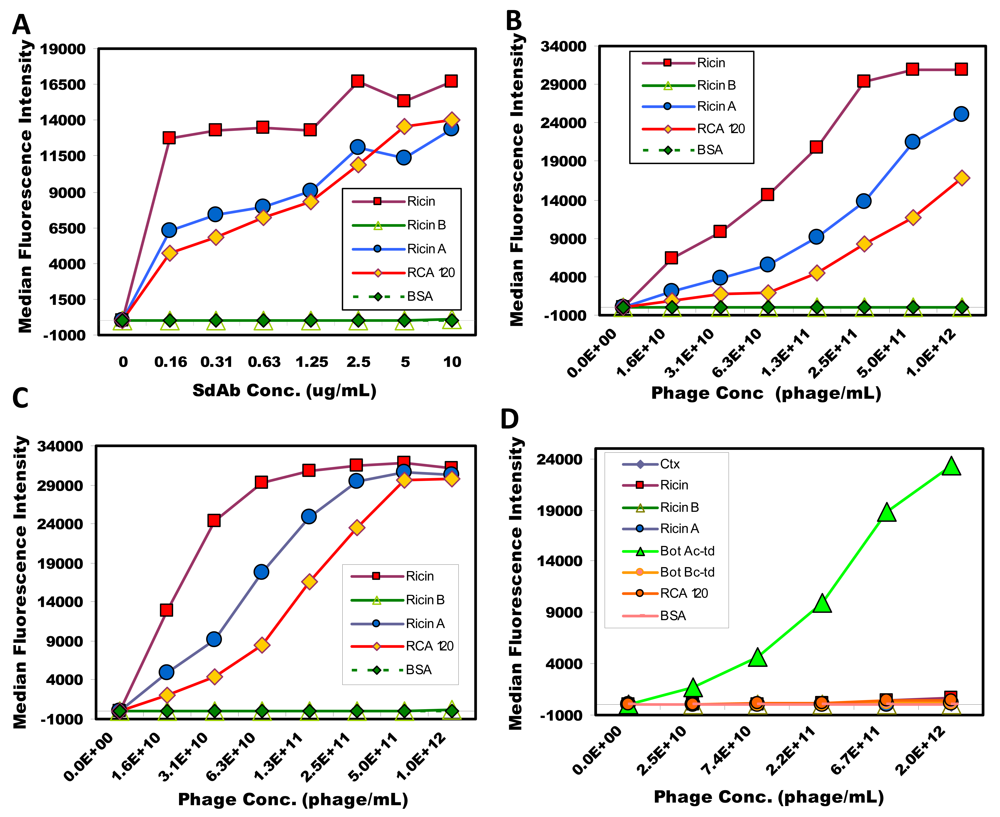
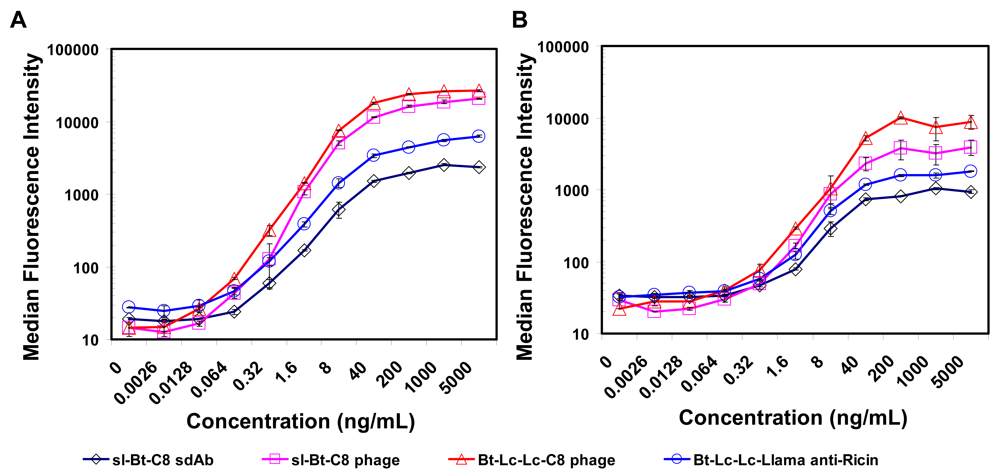
2.3. Luminex assays using soluble sdAb and phage-displayed sdAb
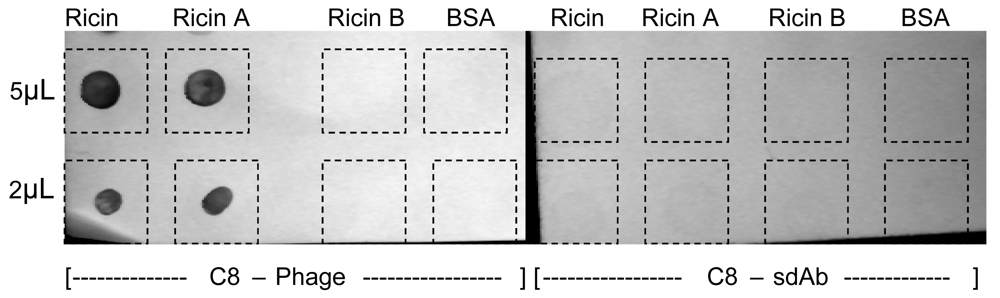
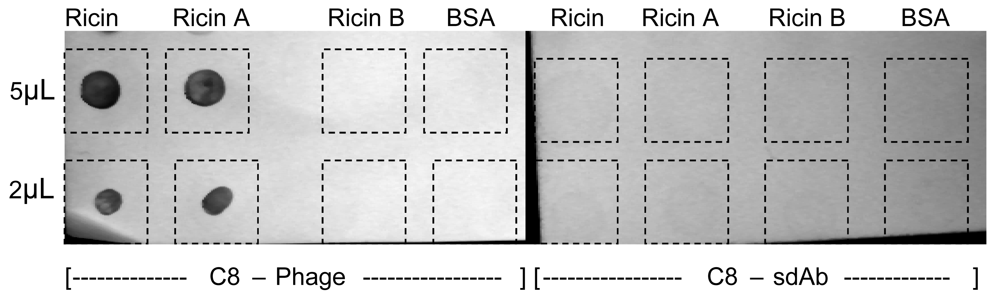
2.4. Phage as reporter reagents
3. Experimental Section
3.1. Reagents
3.2. Biotinylation and fluorphore labeling of sdAb, polyclonal antibodies, and phage
33. Preparation of soluble and phage-displayed sdAb reagents
3.4. ELISA assay protocols
3.5. Preparation of Luminex reagents and assay protocols
4. Conclusions
Acknowledgments
References and Notes
- Garber, E.A.E.; O'Brien, T.W. Detection of ricin in food using electrochemiluminescence-based technology. J. AOAC Int. 2008, 91, 376–382. [Google Scholar]
- Feltis, B.N.; Sexton, B.A.; Glenn, F.L.; Best, M.J.; Wilkins, M.; Davis, T.J. A hand-held surface plasmon resonance biosensor for the detection of ricin and other biological agents. Biosens. Bioelectron. 2008, 23, 1131–1136. [Google Scholar]
- Fulton, R.E.; Thompson, H.G. Fluorogenic hand-held immunoassay for the identification of ricin: Rapid analyte measurement platform. J. Immunoassay Immunochem. 2007, 28, 227–241. [Google Scholar]
- Huelseweh, B.; Ehricht, R.; Marschall, H.J. A simple and rapid protein array based method for the simultaneous detection of biowarfare agents. Proteomics 2006, 6, 2972–2981. [Google Scholar]
- Anderson, G.P.; Lingerfelt, B.M.; Taitt, C.R. Eight analyte detection using a four-channel optical biosensor. Sensor Lett. 2004, 2, 18–24. [Google Scholar]
- Taitt, C.R.; Golden, J.P.; Shubin, Y.S.; Shriver-Lake, L.C.; Sapsford, K.E.; Rasooly, A.; Ligler, F.S. A portable array biosensor for detecting multiple analytes in complex samples. Microbial Ecol. 2004, 47, 175–185. [Google Scholar]
- Bird, R.E.; Hardman, K.D.; Jacobson, J.W.; Johnson, S.; Kaufman, B.M.; Lee, S.M.; Lee, T.; Pope, S.H.; Riordan, G.S.; Whitlow, M. Single-chain Antigen-Binding proteins. Science 1988, 242, 423–426. [Google Scholar]
- Huston, J.S.; Levinson, D.; Mudgetthunter, M.; Tai, M.S.; Novotny, J.; Margolies, M.N.; Ridge, R.J.; Bruccoleri, R.E.; Haber, E.; Crea, R.; Oppermann, H. Protein engineering of antibody-binding sites- recovery of specific activity in an anti-digoxin single-chain FV analog produced in escheria-coli. Proc. Natl. Acad. Sci. U.S.A. 1988, 85, 5879–5883. [Google Scholar]
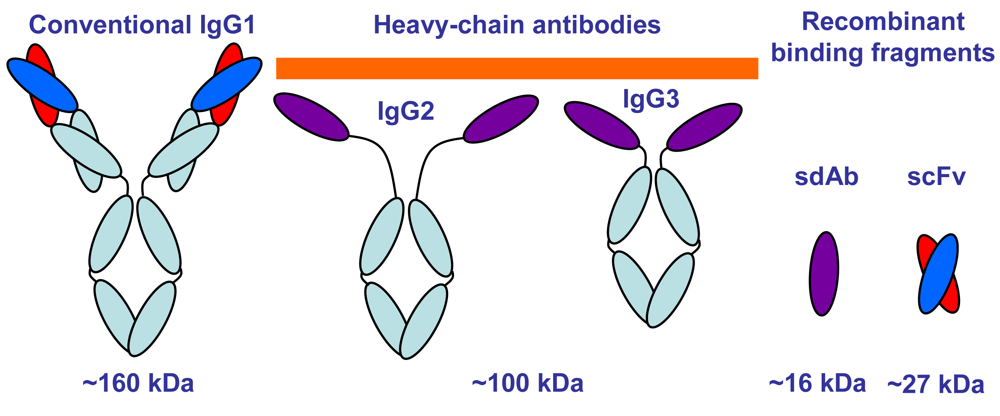
- Hamers-Casterman, C.; Atarhouch, T.; Muyldermans, S.; Robinson, G.; Hamers, C.; Songa, E.B.; Bendahman, N.; Hamers, R. Naturally-occurring antibodies devoid of light chains. Nature 1993, 363, 446–448. [Google Scholar]
- Maass, D.R.; Sepulveda, J.; Pernthaner, A.; Shoemaker, C.B. Alpaca (Lama pacos) as a convenient source of recombinant camelid heavy chain antibodies (VHHs). J. Immunol. Methods 2007, 324, 13–25. [Google Scholar]
- Ghahroudi, M.A.; Desmyter, A.; Wyns, L.; Hamers, R.; Muyldermans, S. Selection and identification of single domain antibody fragments from camel heavy-chain antibodies. FEBS Lett. 1997, 414, 521–526. [Google Scholar]
- Perez, J.M.J.; Renisio, J.G.; Prompers, J.J.; van Platerink, C.J.; Cambillau, C.; Darbon, H.; Frenken, L.G.J. Thermal unfolding of a llama antibody fragment: A two-state reversible process. Biochemistry 2001, 40, 74–83. [Google Scholar]
- Dumoulin, M.; Conrath, K.; Van Meirhaeghe, A.; Meersman, F.; Heremans, K.; Frenken, L.G.J.; Muyldermans, S.; Wyns, L.; Matagne, A. Single-domain antibody fragments with high conformational stability. Protein Sci. 2002, 11, 500–515. [Google Scholar]
- Verheesen, P.; ten Haaft, M.R.; Lindner, N.; Verrips, C.T.; de Haard, J.J.W. Beneficial properties of single-domain antibody fragments for application in immunoaffinity purification and immuno-perfasion chromatography. Biochim. Biophys. Acta 2003, 1624, 21–28. [Google Scholar]
- Tanha, J.; Dubuc, G.; Hirama, T.; Narang, S.A.; MacKenzie, C.R. Selection by phage display of llama conventional V-H fragments with heavy chain antibody VHH properties. J. Immunol. Methods 2002, 263, 97–109. [Google Scholar]
- Yau, K.Y.F.; Groves, M.A.T.; Li, S.; Sheedy, C.; Lee, H.; Tanha, J.; MacKenzie, C.R.; Jermutus, L.; Hall, J.C. Selection of hapten-specific single-domain antibodies from a non-immunized llama ribosome display library. J. Immunol. Methods 2003, 281, 161–175. [Google Scholar]
- Goldman, E.R.; Anderson, G.P.; Liu, J.L.; Delehanty, J.B.; Sherwood, L.J.; Osborn, L.E.; Cummins, L.B.; Hayhurst, A. Facile generation of heat stable antiviral and antitoxin single domain antibodies from a semi-synthetic llama library. Anal. Chem. 2006, 78, 8245–8255. [Google Scholar]
- Dekker, S.; Toussaint, W.; Panayotou, G.; de Wit, T.; Visser, P.; Grosveld, F.; Drabek, D. Intracellularly expressed single-domain antibody against p15 matrix protein prevents the production of porcine retroviruses. J. Virol. 2003, 77, 12132–12139. [Google Scholar]
- Goldman, E.R.; Anderson, G.P.; Conway, J.; Sherwood, L.J.; Fech, M.; Vo, B.H.; Liu, J.L.; Hayhurst, A. Thermostable llama single domain antibodies for detection of botulinum a neurotoxin complex. Anal. Chem. 2008, 80, 8583–8591. [Google Scholar]
- Zarebski, L.M.; Urrutia, M.; Goldbaum, F.A. Llama single domain antibodies as a tool for molecular mimicry. J. Mol. Biol. 2005, 349, 814–824. [Google Scholar]
- Doyle, P.J.; Arbabi-Ghahroudi, M.; Gaudette, N.; Furzer, G.; Savard, M.E.; Gleddie, S.; McLean, M.D.; Mackenzie, C.R.; Hall, J.C. Cloning, expression, and characterization of a single-domain antibody fragment with affinity for 15-acetyl-deoxynivalenol. Mol. Immunol. 2008, 45, 3703–3713. [Google Scholar]
- Scott, J.K.; Smith, G.P. Searching for peptide ligands with an epitope library. Science 1990, 249, 386–390. [Google Scholar]
- McCafferty, J.; Griffiths, A.; Winter, G.; Chiswell, D.J. Phage antibodies-filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar]
- Anderson, G.P.; Lui, J.L.; Hale, M.L.; Bernstein, R.D.; Moore, M.; Swain, M.D.; Goldman, E.R. Development of anti-Ricin single domain antibodies: Towards detection and therapeutic reagents. Anal. Chem. 2008, 80, 9604–9611. [Google Scholar]
- Anderson, G.P.; Ortiz-Vera, Y.A.; Hayhurst, A.; Czarnecki, J.; Dabbs, J.; Vo, B.H.; Goldman, E.R. Evaluation of llama anti-botulinum toxin heavy chain antibody. Botulinum J. 2008, 1, 100–115. [Google Scholar]
- Anderson, G.P.; Matney, R.; Liu, J.L.; Hayhurst, A.; Goldman, E.R. Multiplexed fluid array screening of phage displayed anti-ricin single domain antibodies for rapid assessment of specificity. BioTechniques 2007, 43, 806–811. [Google Scholar]
- Roberts, L.M.; Lamb, F.I.; Pappin, D.J.C.; Lord, J.M. The primary sequence of Ricinus communis agglutinin. Comparison with ricin. J. Biol. Chem. 1985, 260, 15682–15686. [Google Scholar]
- Hayhurst, A.; Happe, S.; Mabry, R.; Koch, Z.; Iverson, B.L.; Georgiou, G. Isolation and expression of recombinant antibody fragments to the biological warfare pathogen. Brucella Melitensis. J. Immunol. Methods 2003, 276, 185–196. [Google Scholar]
- Sherwood, L.J.; Osborn, L.E.; Carrion, R., Jr.; Patterson, J.L.; Hayhurst, A. Rapid assembly of sensitive antigen-capture assays for marburg virus, using in vitro selection of llama single-domain antibodies, at biosafety level 4. J. Infect. Dis. 2007, 196(Suppl. 2), S213–219. [Google Scholar]
- Henry, T.J.; Pratt, D. The Proteins of Bacteriophage M13. Proc. Natl. Acad. Sci. U.S.A. 1969, 62, 800–807. [Google Scholar]
- Smith, G.P.; Petrenko, V.A. Phage display. Chem. Rev. 1997, 97, 391–410. [Google Scholar]
- Zhou, B.; Wirsching, P.; Janda, K.D. Human antibodies against spores of the genus bacillus: a model study for detection of and protection against anthrax and the bioterrorist threat. Proc. Natl. Acad. Sci. U.S.A. 2002, 99, 5241–5246. [Google Scholar]
- Goldman, E.R.; Pazirandeh, M.P.; Mauro, J.M.; King, K.D.; Frey, J.C.; Anderson, G.P. Phage-displayed peptides as biosensor reagents. J. Mol. Recognit. 2000, 13, 382–387. [Google Scholar]
- Newton, J.R.; Kelly, K.A.; Mahmood, U.; Weissleder, R.; Deutscher, S.L. In Vivo selection of phage for the optical imaging of OC-3 human prostate carcinoma in mice. Neoplasia 2006, 8, 772–780. [Google Scholar]
- Li, Q.; Shivachandra, S.B.; Leppla, S.H.; Rao, V.B. Bacteriophage T4 capsid: a unique platform for efficient surface assembly of macromolecular complexes. J. Mol. Biol. 2006, 363, 577–588. [Google Scholar]
- Cerritelli, M.E.; Conway, J.F.; Cheng, N.; Trus, B.L.; Steven, A.C. Molecular mechanisms in bacteriophage T7 procapsid assembly, maturation, and DNA containment. Adv. Protein Chem. 2003, 64, 301–323. [Google Scholar]
- Soto, C.M.; Blum, A.S.; Vora, G.J.; Lebedev, N.; Meador, C.E.; Won, A.P.; Chatterji, A.; Johnson, J.E.; Ratna, B.R. Fluorescent signal amplification of carbocyanine dyes using engineered viral nanoparticles. J. Am. Chem. Soc. 2006, 128, 5184–5189. [Google Scholar]
- Sapsford, K.E.; Soto, C.M.; Blum, A.S.; Chatterji, A.; Lin, T.; Johnson, J.E.; Ligler, F.S.; Ratna, B.R. A cowpea mosaic virus nanoscaffold for multiplexed antibody conjugation: application as an immunoassay tracer. Biosens. Bioelectron. 2006, 21, 1668–1673. [Google Scholar]
- Nanduri, V.; Bhunia, A.K.; Tu, S.I.; Paoli, G.C.; Brewster, J.D. SPR biosensor for the detection of L-monocytogenes using phage-displayed antibody. Biosens. Bioelectron. 2007, 23, 248–252. [Google Scholar]
- Smith, G.P.; Scott, J.K. Libraries of peptides and proteins displayed on filamentous phage. Method. Enzymol. 1996, 217, 228–257. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Goldman, E.R.; Liu, J.L.; Bernstein, R.D.; Swain, M.D.; Mitchell, S.Q.; Anderson, G.P. Ricin Detection Using Phage Displayed Single Domain Antibodies. Sensors 2009, 9, 542-555. https://doi.org/10.3390/s90100542
Goldman ER, Liu JL, Bernstein RD, Swain MD, Mitchell SQ, Anderson GP. Ricin Detection Using Phage Displayed Single Domain Antibodies. Sensors. 2009; 9(1):542-555. https://doi.org/10.3390/s90100542
Chicago/Turabian StyleGoldman, Ellen R., Jinny L. Liu, Rachael D. Bernstein, Marla D. Swain, Stanley Q. Mitchell, and George P. Anderson. 2009. "Ricin Detection Using Phage Displayed Single Domain Antibodies" Sensors 9, no. 1: 542-555. https://doi.org/10.3390/s90100542