Disruption of the Expression of a Non-Coding RNA Significantly Impairs Cellular Differentiation in Toxoplasma gondii


Abstract
:1. Introduction
2. Results and Discussion
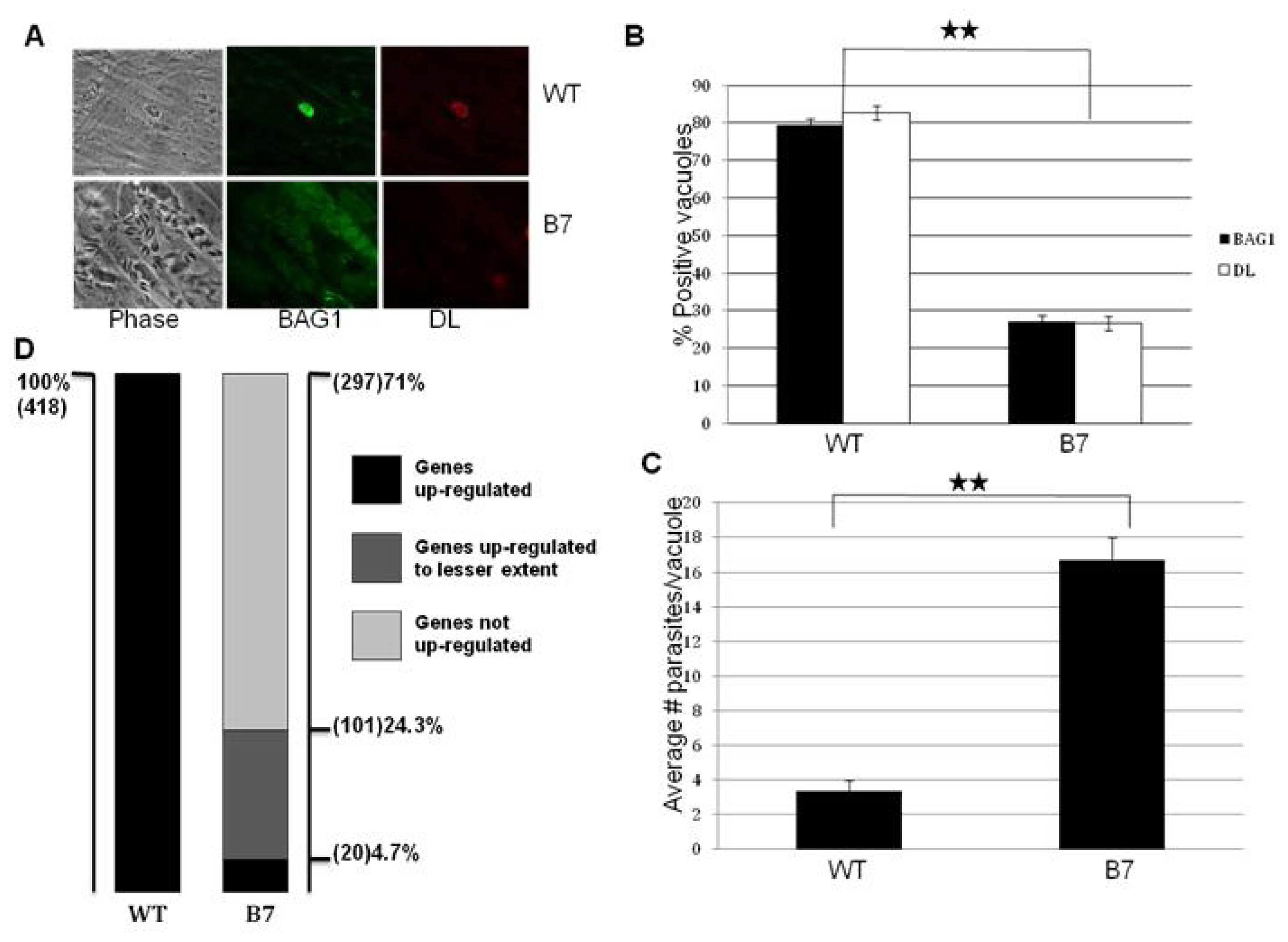
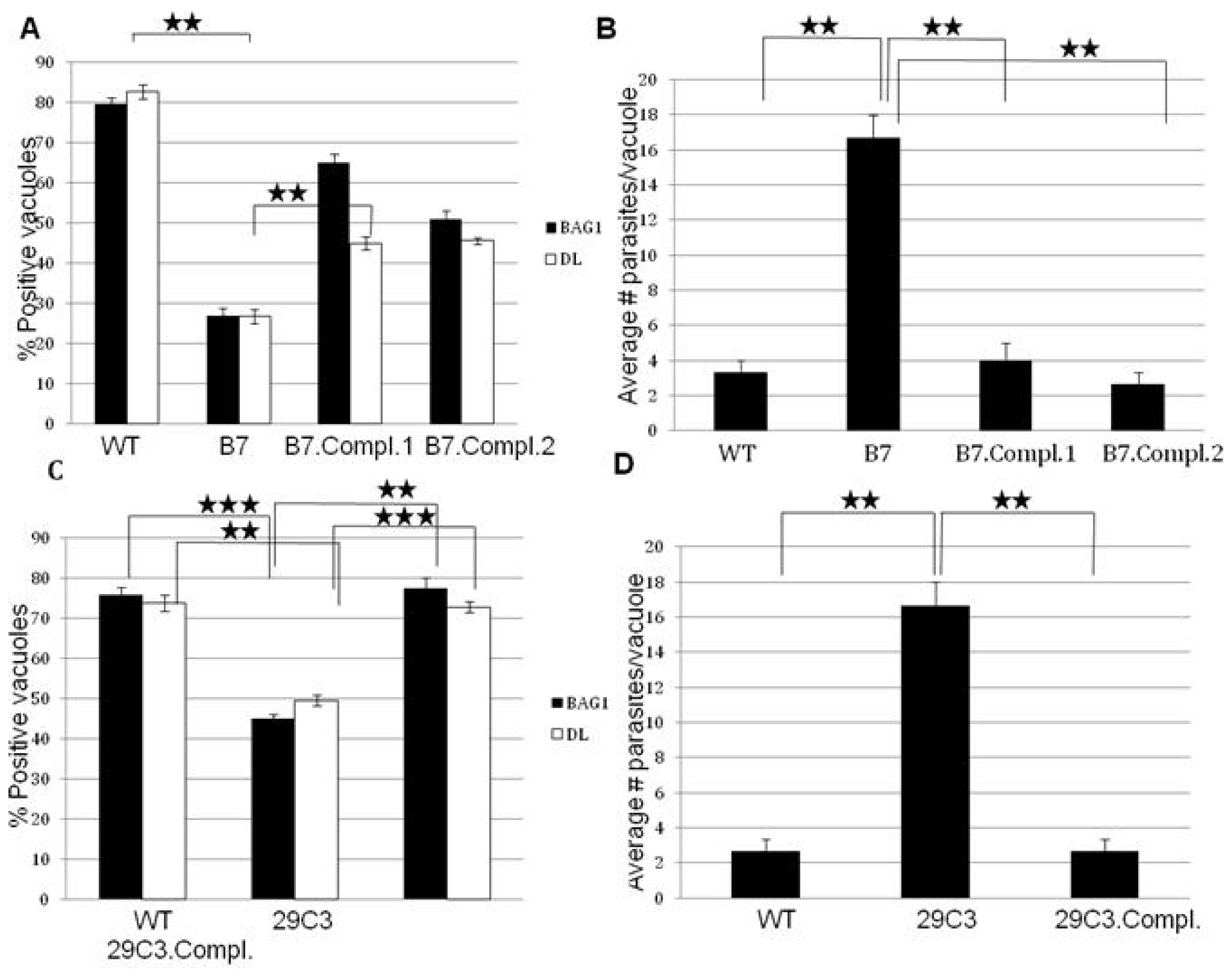
2.1. Mutant B7 Is Severely Impaired to Form Bradyzoites
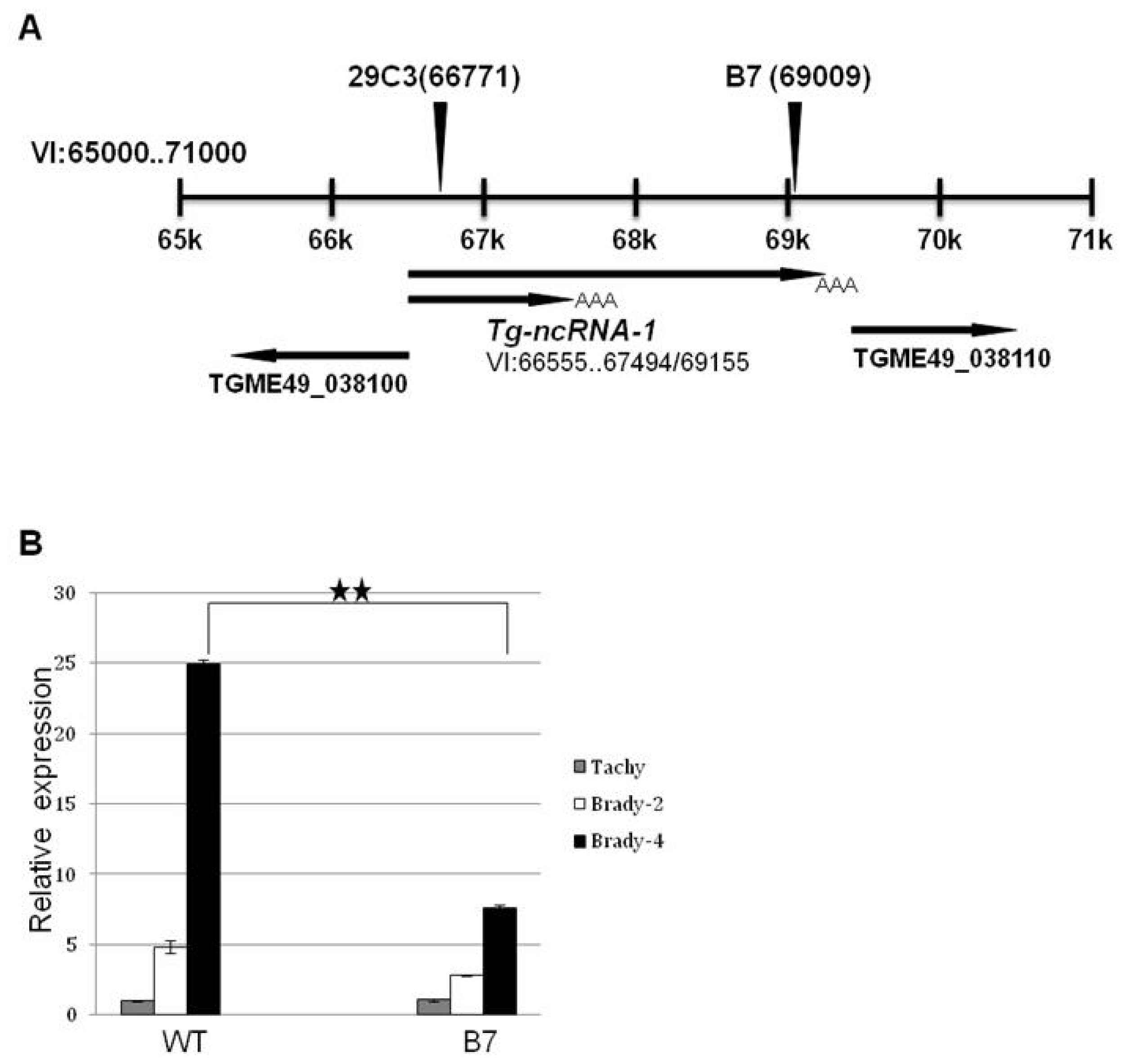
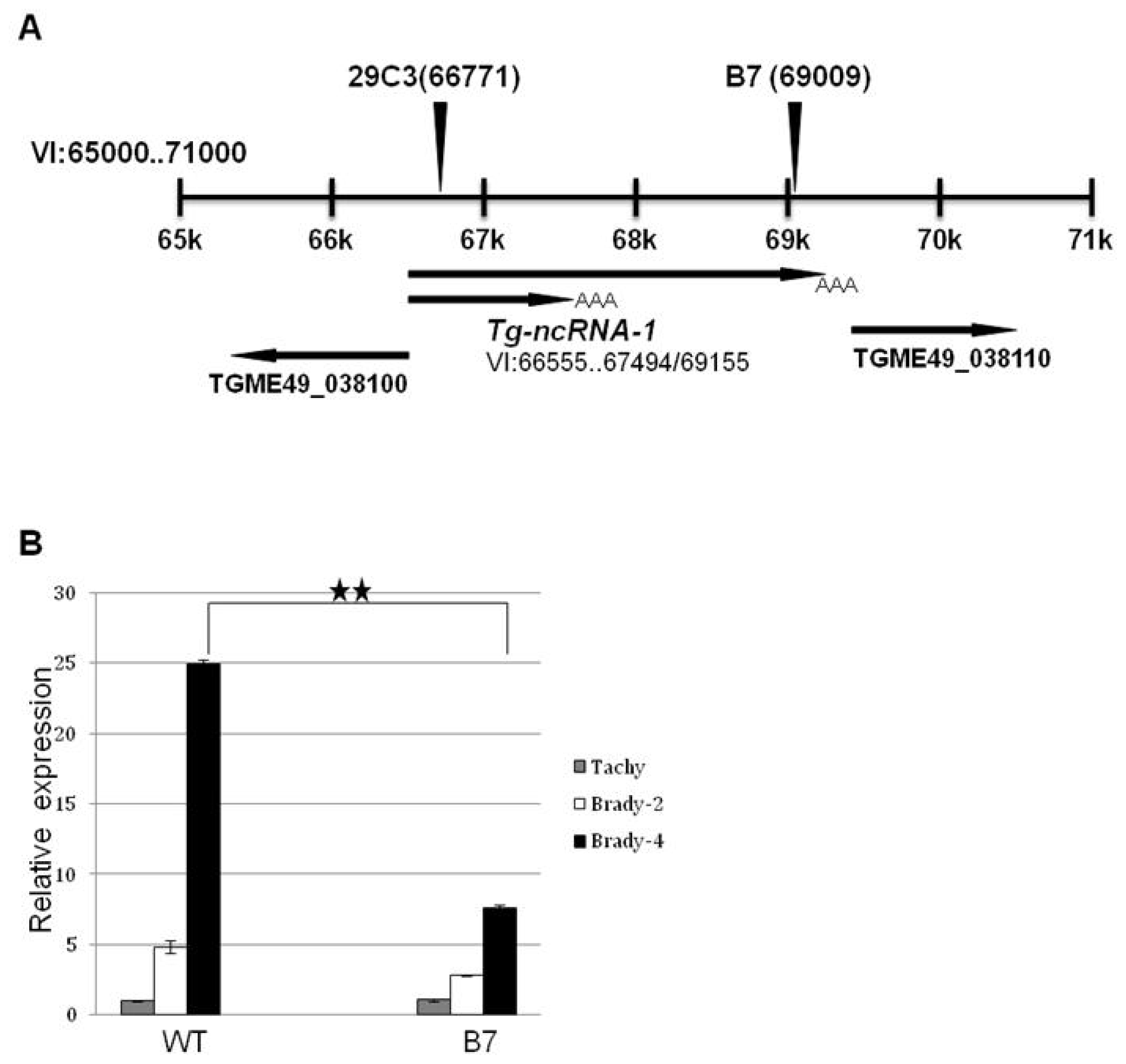
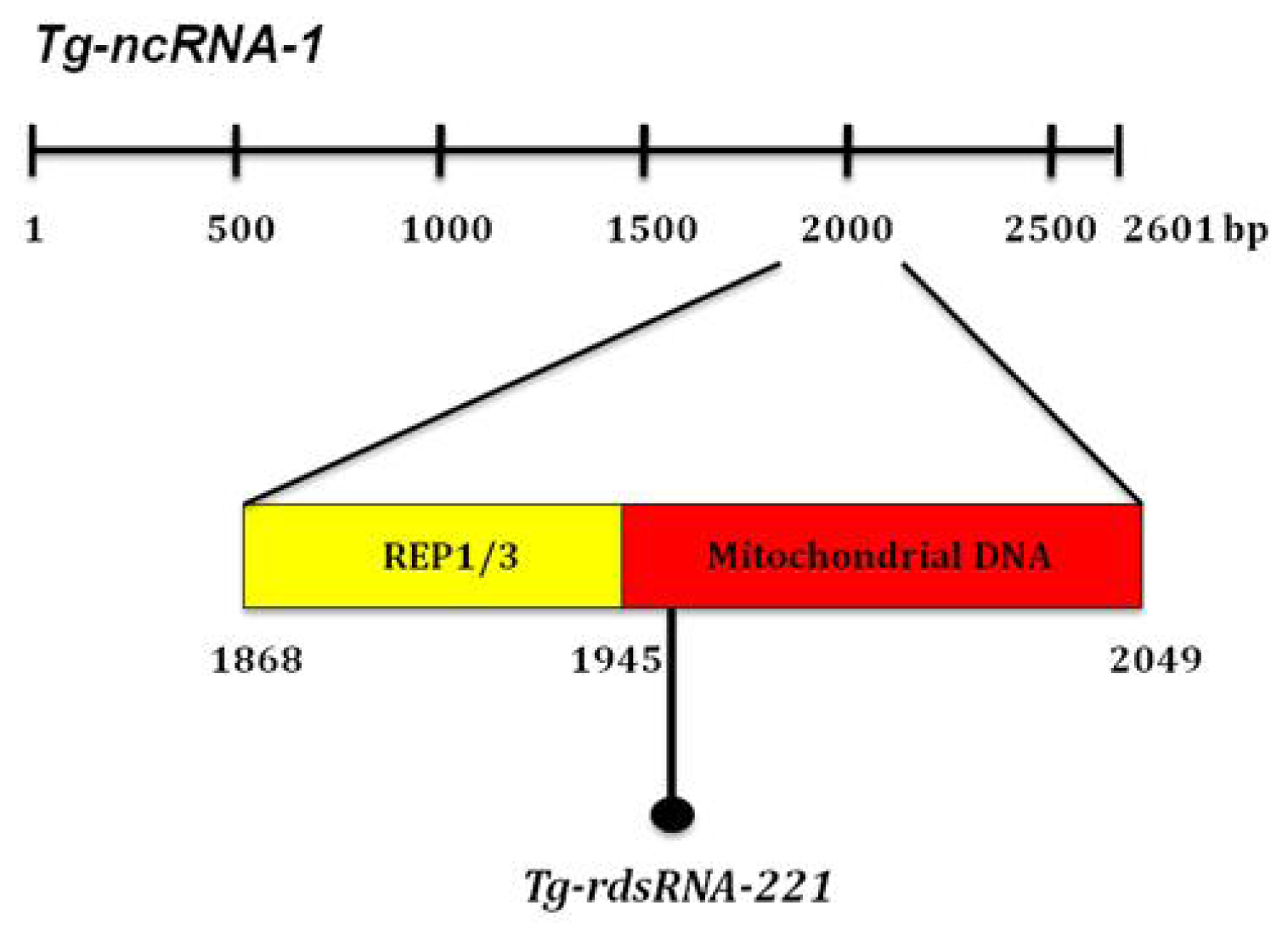
2.2. The Locus Disrupted in Mutant B7 Encodes a Non-Coding RNA
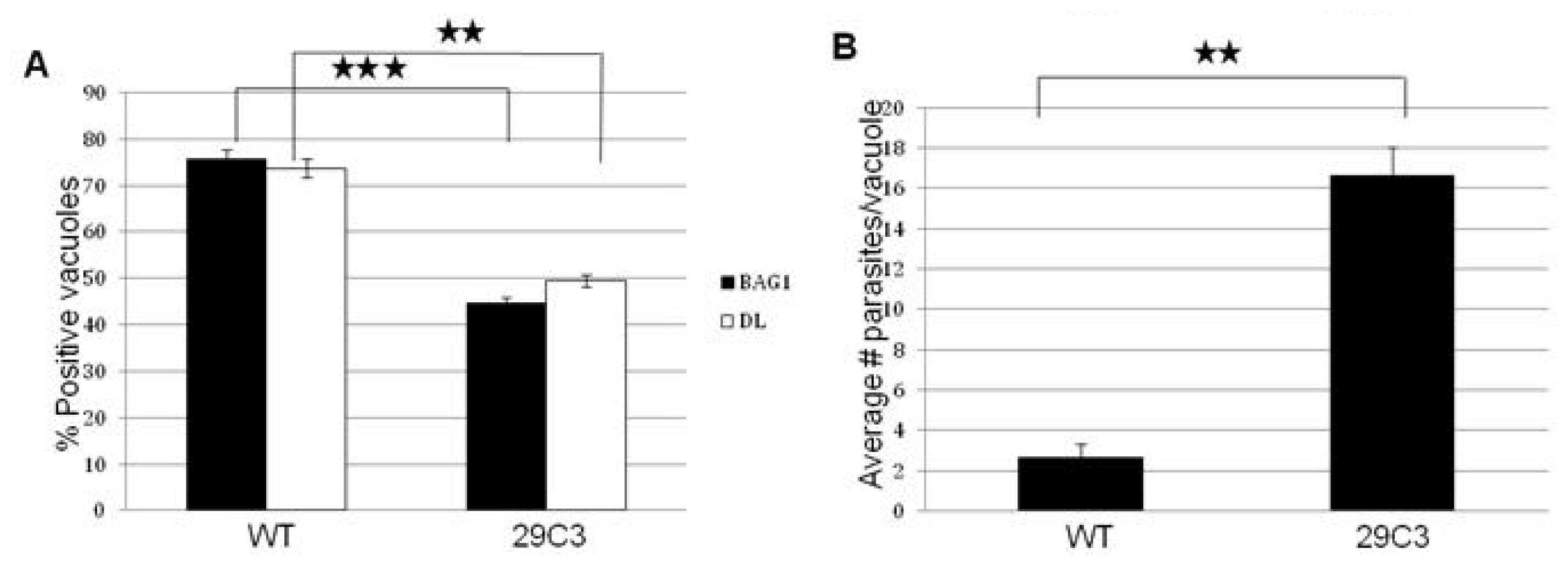
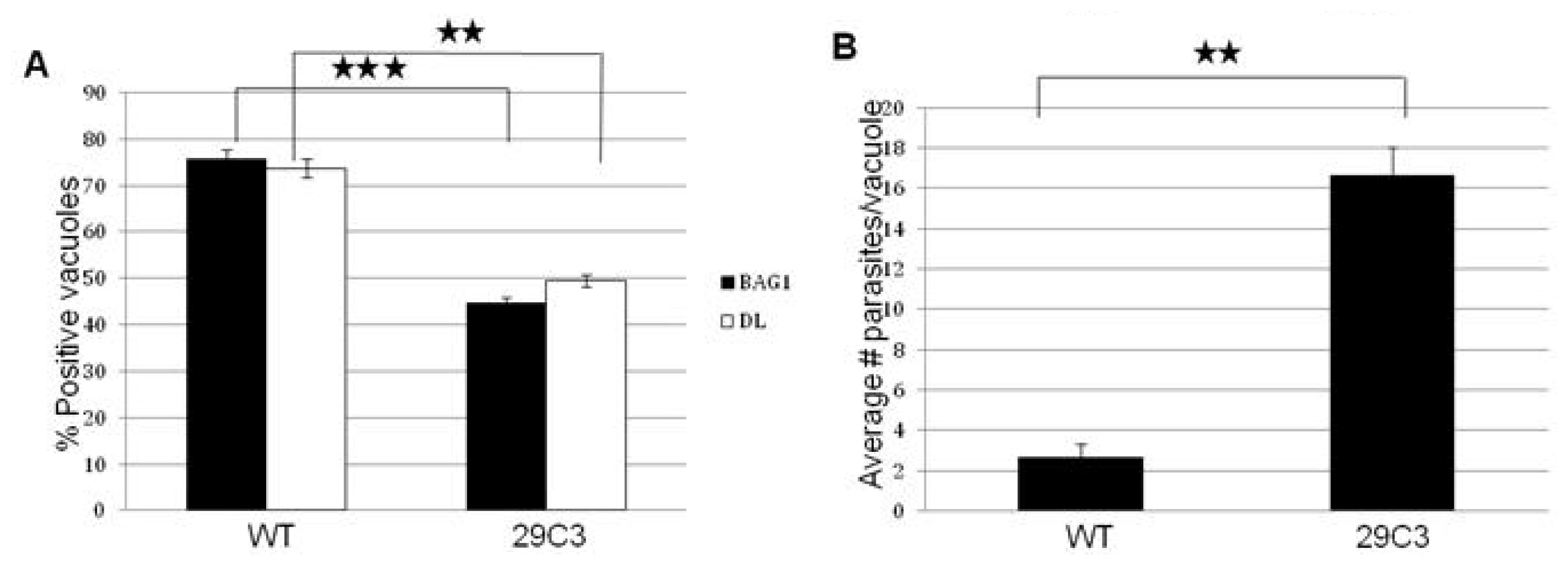
2.3. Another Mutant (29C3) That Exhibits a Defect in Bradyzoite Differentiation also Has an Insertion within the Gene Tg-ncRNA-1
2.4. Complementation of Mutants B7 and 29C3 Restores the WT Phenotype
3. Experimental Section
3.1. Parasite Growth and Differentiation
3.2. Immunofluorescence Assays
3.3. Transfection and Bleomycin (BLE) Selection
3.4. Quantitative Real Time RT PCR (TaqMan)
4. Conclusions
Acknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Wong, S.Y.; Remington, J.S. Biology of Toxoplasma gondii. AIDS 1993, 7, 299–316. [Google Scholar]
- Montoya, J.G.; Liesenfeld, O. Toxoplasmosis. Lancet 2004, 363, 1965–1976. [Google Scholar]
- Boothroyd, J.C.; Black, M.; Bonnefoy, S.; Hehl, A.; Knoll, L.J.; Manger, I.D.; Ortega-Barria, E.; Tomavo, S. Genetic and biochemical analysis of development in Toxoplasma gondii. Philos. Trans. R. Soc. Lond. B 1997, 352, 1347–1354. [Google Scholar]
- Weiss, L.M.; Kim, K. The Development and biology of bradyzoites of Toxoplasma gondii. Front. Biosci 2000, 5, D391–D405. [Google Scholar]
- Soête, M.; Camus, D.; Dubremetz, J.F. Experimental induction of bradyzoite-specific antigen expression and Cyst formation by the RH strain of Toxoplasma gondii in vitro. Exp. Parasitol 1994, 78, 361–370. [Google Scholar]
- Soete, M.; Fortier, B.; Camus, D.; Dubremetz, J.F. Toxoplasma gondii: Kinetics of bradyzoite-tachyzoite interconversion in vitro. Exp. Parasitol 1993, 76, 259–264. [Google Scholar]
- Weiss, L.M.; Laplace, D.; Takvorian, P.M.; Tanowitz, H.B.; Cali, A.; Wittner, M.A. Cell culture system for study of the development of Toxoplasma gondii bradyzoites. J. Eukaryot. Microbiol 1995, 42, 150–157. [Google Scholar]
- Suzuki, Y.; Orellana, M.A.; Schreiber, R.D.; Remington, J.S. Interferon-gamma: The major mediator of resistance against Toxoplasma gondii. Science 1988, 240, 516–518. [Google Scholar]
- Subauste, C.S.; Remington, J.S. Role of gamma interferon in Toxoplasma gondii infection. Eur. J. Clin. Microbiol. Infect. Dis 1991, 10, 58–67. [Google Scholar]
- Behnke, M.S.; Radke, J.B.; Smith, A.T.; Sullivan, W.J., Jr; White, M.W. The Transcription of bradyzoite genes in Toxoplasma gondii is controlled by autonomous promoter elements. Mol. Microbiol. 2008, 68, 1502–1518. [Google Scholar]
- Lescault, P.J.; Thompson, A.B.; Patil, V.; Lirussi, D.; Burton, A.; Margarit, J.; Bond, J.; Matrajt, M. Genomic data reveal Toxoplasma gondii differentiation mutants are also impaired with respect to switching into a novel extracellular tachyzoite state. PLoS One 2010, 5, e14463. [Google Scholar]
- Matrajt, M.; Donald, R.G.; Singh, U.; Roos, D.S. Identification and characterization of differentiation mutants in the protozoan parasite Toxoplasma gondii. Mol. Microbiol 2002, 44, 735–747. [Google Scholar]
- Singh, U.; Brewer, J.L.; Boothroyd, J.C. Genetic analysis of tachyzoite to bradyzoite differentiation mutants in Toxoplasma gondii reveals a hierarchy of gene induction. Mol. Microbiol 2002, 44, 721–733. [Google Scholar]
- Anderson, M.Z.; Brewer, J.; Singh, U.; Boothroyd, J.C. A pseudouridine synthase homologue is critical to cellular differentiation in Toxoplasma gondii. Eukaryot. Cell 2009, 8, 398–409. [Google Scholar]
- Vanchinathan, P.; Brewer, J.L.; Harb, O.S.; Boothroyd, J.C.; Singh, U. Disruption of a locus encoding a nucleolar zinc finger protein decreases tachyzoite-to-bradyzoite differentiation in Toxoplasma gondii. Infect. Immun 2005, 73, 6680–6688. [Google Scholar]
- Bohne, W.; Gross, U.; Ferguson, D.J.; Heesemann, J. Cloning and characterization of a bradyzoite-specifically expressed gene (hsp30/bag1) of Toxoplasma gondii, related to genes encoding small heat-shock proteins of plants. Mol. Microbiol 1995, 16, 1221–1230. [Google Scholar]
- Fichera, M.E.; Bhopale, M.K.; Roos, D.S. In vitro assays elucidate peculiar kinetics of clindamycin action against Toxoplasma gondii. Antimicrob. Agents Chemother 1995, 39, 1530–1537. [Google Scholar]
- Matrajt, M. Non-coding RNA in apicomplexan parasites. Mol. Biochem. Parasitol 2010, 174, 1–7. [Google Scholar]
- Kong, L.; Zhang, Y.; Ye, Z.Q.; Liu, X.Q.; Zhao, S.Q.; Wei, L.; Gao, G. CPC: Assess the protein-coding potential of transcripts using sequence features and support vector machine. Nucleic Acids Res 2007, 35, W345–W349. [Google Scholar]
- Braun, L.; Cannella, D.; Ortet, P.; Barakat, M.; Sautel, C.F.; Kieffer, S.; Garin, J.; Bastien, O.; Voinnet, O.; Hakimi, M.A. A complex small RNA repertoire is generated by a plant/fungal-like machinery and effected by a metazoan-like argonaute in the single-cell human parasite Toxoplasma gondii. PLoS Pathog 2010, 6, e1000920. [Google Scholar]
- Ossorio, P.N.; Sibley, D.L.; Boothroyd, J.C. Mitochondrial-like DNA sequences flanked by direct and inverted repeats in the nuclear genome of Toxoplasma gondii. J. Mol. Biol 1991, 222, 525–536. [Google Scholar]
- Frankel, M.B.; Mordue, D.G.; Knoll, L.J. Discovery of parasite virulence genes reveals a unique regulator of chromosome condensation 1 ortholog critical for efficient nuclear trafficking. Proc. Natl. Acad. Sci. USA 2007, 104, 10181–10186. [Google Scholar]
- Messina, M.; Niesman, I.; Mercier, C.; Sibley, L.D. Stable DNA Transformation of Toxoplasma gondii using phleomycin selection. Gene 1995, 165, 213–217. [Google Scholar]
- Soldati, D.; Kim, K.; Kampmeier, J.; Dubremetz, J.F.; Boothroyd, J.C. Complementation of Toxoplasma gondii ROP1 knock-out mutant using phleomycin selection. Mol. Biochem. Parasitol 1995, 74, 87–97. [Google Scholar]
- Roos, D.S.; Donald, R.G.; Morrissette, N.S.; Moulton, A.L.C. Molecular tools for genetic dissection of the protozoan parasite Toxoplasma gondii. Methods Cell Biol 1995, 45, 27–63. [Google Scholar]
- Mattick, J.S. A new paradigm for developmental biology. J. Exp. Biol 2007, 210, 1526–1547. [Google Scholar]
- Amaral, P.P.; Mattick, J.S. Non-coding RNA in development. Mamm. Genome 2008, 19, 454–492. [Google Scholar]
- Khalil, A.M.; Guttman, M.; Huarte, M.; Garber, M.; Raj, A.; Rivea Morales, D.; Thomas, K.; Presser, A.; Bernstein, B.E.; et al. Many human large intergenic non-coding RNAs associate with chromatin-modifying complexes and affect gene expression. Proc. Natl. Acad. Sci. USA 2009, 106, 11667–11672. [Google Scholar]
- Ullu, E.; Tschudi, C.; Chakraborty, T. RNA interference in protozoan parasites. Cell. Microbiol 2004, 6, 509–519. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence ID | C/NC | Coding potential score | Hit num | Hit score | Frame score | Coverage | Log-odds score | Type |
|---|---|---|---|---|---|---|---|---|
| B41(L) | Noncoding | −1.4067 | 0 | 0.0 | 0.0 | 8.07% | 20.32 | Full |
| B41(S) | Noncoding | −1.2738 | 0 | 0.0 | 0.0 | 22.34% | 20.32 | Full |
| TGGT1_087710 cGMP kinase | Coding | 2.69204 | 250 | 49.54 | 514.4 | 3.73% | 80.42 | Partial |
| TGME49_000320 HXGPRT | Coding | 1.39359 | 140 | 17.73 | 128.9 | 12.41% | 62.36 | Partial |
| Tsix_mus | Noncoding | −1.3004 | 0 | 0.0 | 0.0 | 3% | 27.5 | Full |
| AF282387 | Coding | 3.32462 | 150 | 25.00 | 208.4 | 99.43% | 109.4 | Full |
© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Patil, V.; Lescault, P.J.; Lirussi, D.; Thompson, A.B.; Matrajt, M. Disruption of the Expression of a Non-Coding RNA Significantly Impairs Cellular Differentiation in Toxoplasma gondii. Int. J. Mol. Sci. 2013, 14, 611-624. https://doi.org/10.3390/ijms14010611
Patil V, Lescault PJ, Lirussi D, Thompson AB, Matrajt M. Disruption of the Expression of a Non-Coding RNA Significantly Impairs Cellular Differentiation in Toxoplasma gondii. International Journal of Molecular Sciences. 2013; 14(1):611-624. https://doi.org/10.3390/ijms14010611
Chicago/Turabian StylePatil, Veerupaxagouda, Pamela J. Lescault, Dario Lirussi, Ann B. Thompson, and Mariana Matrajt. 2013. "Disruption of the Expression of a Non-Coding RNA Significantly Impairs Cellular Differentiation in Toxoplasma gondii" International Journal of Molecular Sciences 14, no. 1: 611-624. https://doi.org/10.3390/ijms14010611