Expression and Functional Studies on the Noncoding RNA, PRINS

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
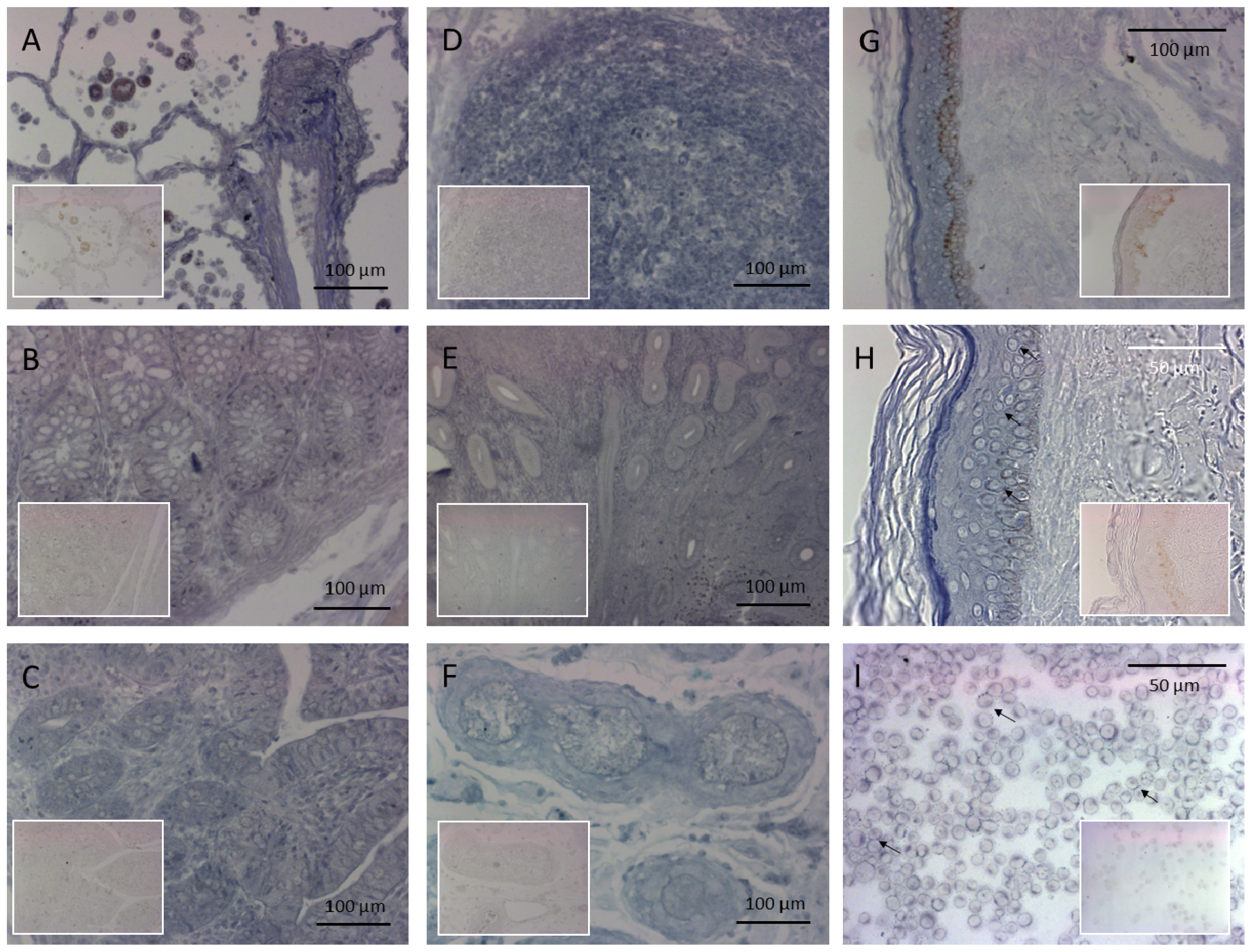
2.1. The Histological and Cellular Distribution of PRINS
2.2. PRINS Expression in Healthy and Psoriatic Skin
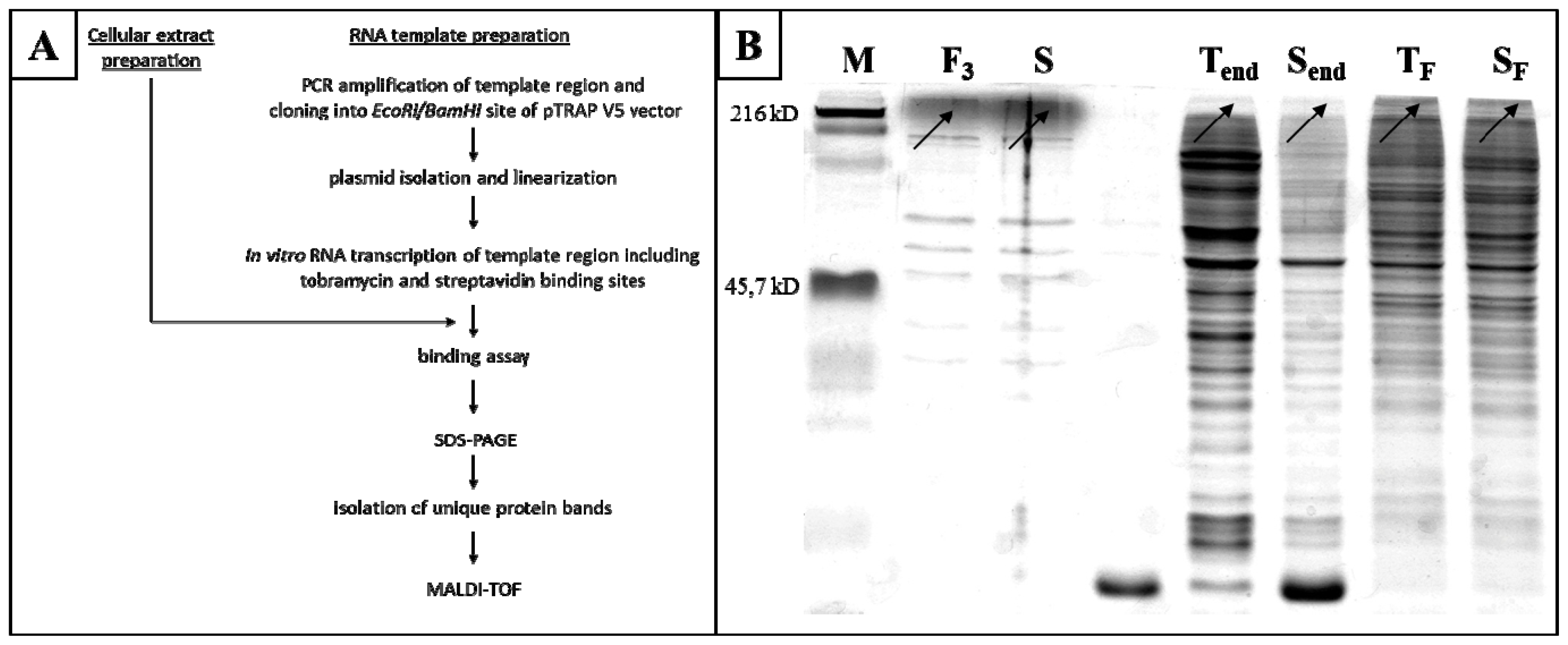
2.3. In Vitro Identification of NPM as a PRINS-Interacting Protein
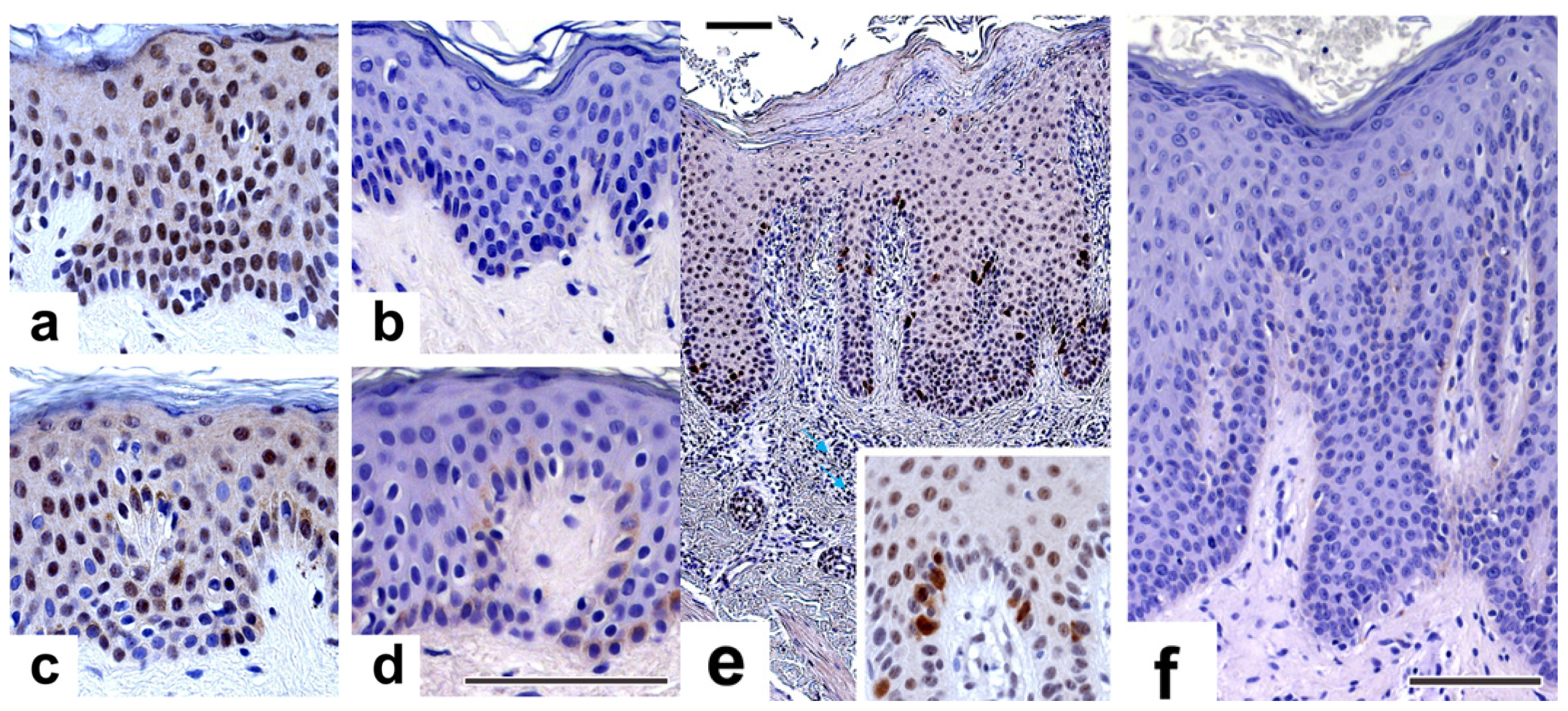
2.4. NPM Is Overexpressed in Psoriatic Involved Epidermis
2.5. NPM Expression Decreases as Keratinocytes Differentiate
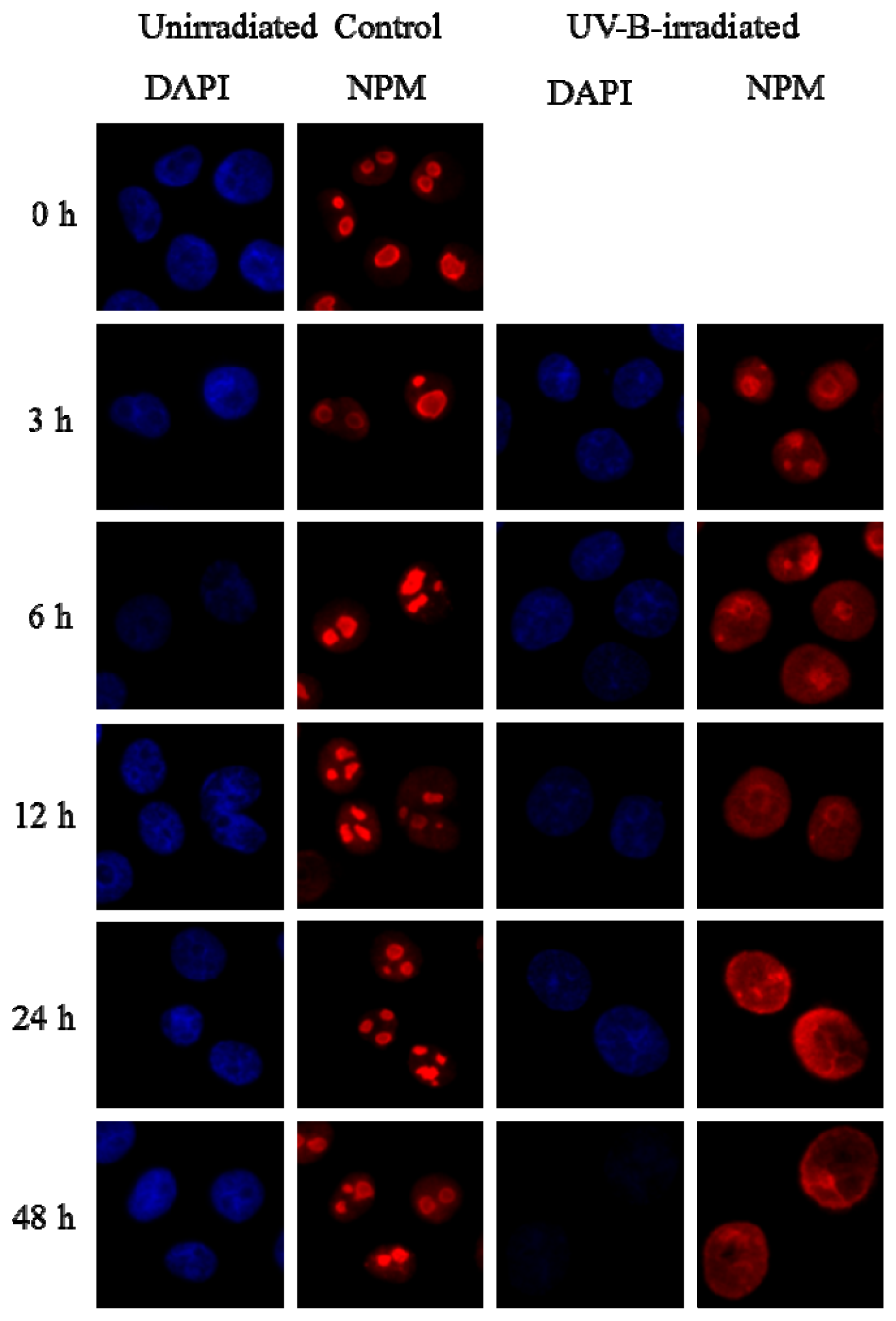
2.6. Intracellular Localization of NPM Protein in Keratinocytes
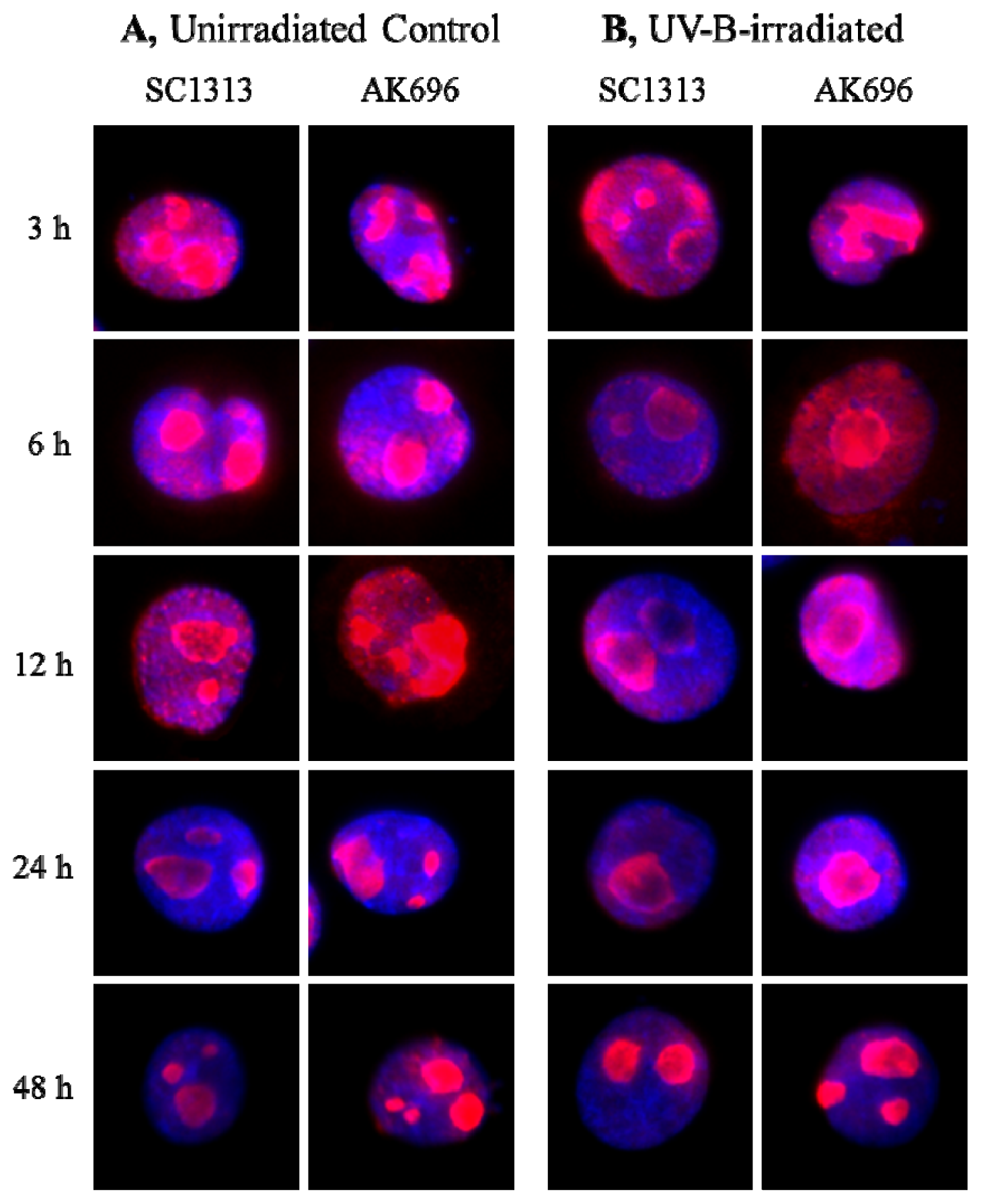
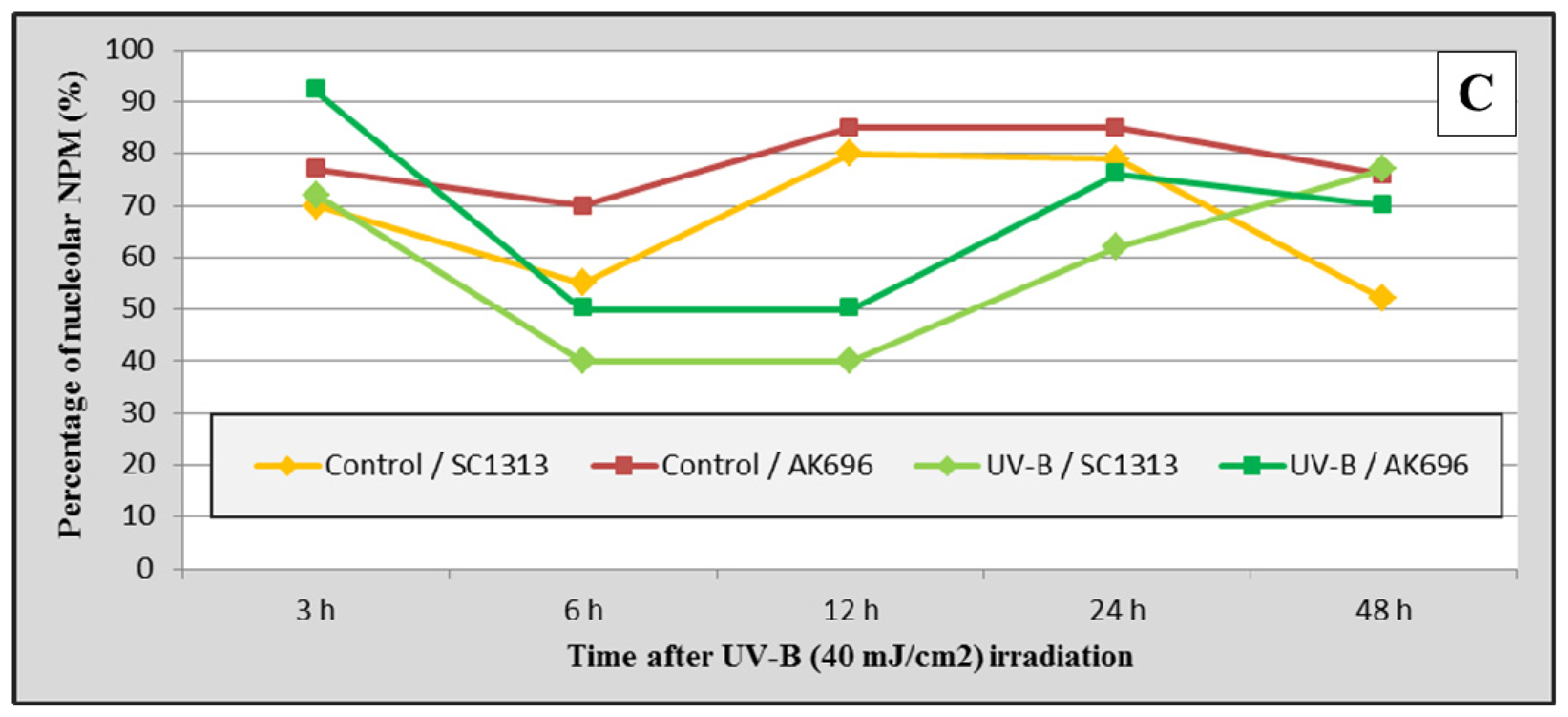
2.7. PRINS Modifies the UV-B Irradiation-Induced Intracellular Shuttling of NPM
2.8. In Silico Prediction and Gene-Specific Silencing Experiments Suggest that PRINS Might Be Part of the Transcription Regulatory CTCF/NPM Complex and Acts as a Negative Regulator of Cell Cycle Progression
3. Experimental Section
3.1. Cell Cultures
3.2. In Situ Hybridization
3.3. In Vitro Binding Assay
3.4. Real-Time RT-PCR
3.5. Western Blot Analysis
3.6. Immunohistochemistry
3.7. Gene-Specific Silencing
3.8. Immunocytochemistry
3.9. In Silico Prediction of PRINS-Binding Proteins
4. Conclusions
Supplementary Information
ijms-14-00205-s001.pdfAcknowledgments
- Conflict of InterestThe authors declare no conflict of interest.
References
- Lebwohl, M. Psoriasis. Lancet 2003, 361, 1197–1204. [Google Scholar]
- Bowcock, A.M.; Krueger, J.G. Getting under the skin: The immunogenetics of psoriasis. Nat. Rev. Immunol 2005, 5, 699–711. [Google Scholar]
- Duffy, D.L.; Spelman, L.S.; Martin, N.G. Psoriasis in Australian twins. J. Am. Acad. Dermatol 1993, 29, 428–434. [Google Scholar]
- Bos, J.D.; De Rie, M.A.; Teunissen, M.B.; Piskin, G. Psoriasis: Ddysregulation of innate immunity. Br. J. Dermatol 2005, 152, 1098–1107. [Google Scholar]
- Bos, J.D.; Hulsebosch, H.J.; Krieg, S.R.; Bakker, P.M.; Cormane, R.H. Immunocompetent cells in psoriasis. In situ immunophenotyping by monoclonal antibodies. Arch. Dermatol. Res 1983, 275, 181–189. [Google Scholar]
- Bata-Csorgo, Z.; Hammerberg, C.; Voorhees, J.J.; Cooper, K.D. Kinetics and regulation of human keratinocyte stem cell growth in short-term primary ex vivo culture. Cooperative growth factors from psoriatic lesional T lymphocytes stimulate proliferation among psoriatic uninvolved, but not normal, stem keratinocytes. J. Clin. Invest 1995, 95, 317–327. [Google Scholar]
- Bata-Csorgo, Z.; Cooper, K.D.; Ting, K.M.; Voorhees, J.J.; Hammerberg, C. Fibronectin and alpha5 integrin regulate keratinocyte cell cycling. A mechanism for increased fibronectin potentiation of T cell lymphokine-driven keratinocyte hyperproliferation in psoriasis. J. Clin. Invest 1998, 101, 1509–1518. [Google Scholar]
- Stoeckman, A.K.; Baechler, E.C.; Ortmann, W.A.; Behrens, T.W.; Michet, C.J.; Peterson, E.J. A distinct inflammatory gene expression profile in patients with psoriatic arthritis. Genes Immun 2006, 7, 583–591. [Google Scholar]
- Carlen, L.M.; Sanchez, F.; Bergman, A.C.; Becker, S.; Hirschberg, D.; Franzen, B.; Coffey, J.; Jornvall, H.; Auer, G.; Alaiya, A.A.; et al. Proteome analysis of skin distinguishes acute guttate from chronic plaque psoriasis. J. Invest. Dermatol 2005, 124, 63–69. [Google Scholar]
- Zhou, X.; Krueger, J.G.; Kao, M.C.; Lee, E.; Du, F.; Menter, A.; Wong, W.H.; Bowcock, A.M. Novel mechanisms of T-cell and dendritic cell activation revealed by profiling of psoriasis on the 63,100-element oligonucleotide array. Physiol. Genomics 2003, 13, 69–78. [Google Scholar]
- Nomura, I.; Gao, B.; Boguniewicz, M.; Darst, M.A.; Travers, J.B.; Leung, D.Y. Distinct patterns of gene expression in the skin lesions of atopic dermatitis and psoriasis: A gene microarray analysis. J. Allergy Clin. Immunol 2003, 112, 1195–1202. [Google Scholar]
- Bowcock, A.M.; Shannon, W.; Du, F.; Duncan, J.; Cao, K.; Aftergut, K.; Catier, J.; Fernandez-Vina, M.A.; Menter, A. Insights into psoriasis and other inflammatory diseases from large-scale gene expression studies. Hum. Mol. Genet 2001, 10, 1793–1805. [Google Scholar]
- Sonkoly, E.; Bata-Csorgo, Z.; Pivarcsi, A.; Polyanka, H.; Kenderessy-Szabo, A.; Molnar, G.; Szentpali, K.; Bari, L.; Megyeri, K.; Mandi, Y.; et al. Identification and characterization of a novel, psoriasis susceptibility-related noncoding RNA gene, PRINS. J. Biol. Chem 2005, 280, 24159–24167. [Google Scholar]
- Tiedge, H.; Fremeau, R.T., Jr; Weinstock, P.H.; Arancio, O.; Brosius, J. Dendritic location of neural BC1 RNA. Proc. Natl. Acad. Sci. USA 1991, 88, 2093–2097. [Google Scholar]
- Mattick, J.S. Challenging the dogma: The hidden layer of non-protein-coding RNAs in complex organisms. Bioessays 2003, 25, 930–939. [Google Scholar]
- Zalfa, F.; Adinolfi, S.; Napoli, I.; Kuhn-Holsken, E.; Urlaub, H.; Achsel, T.; Pastore, A.; Bagni, C. Fragile X mental retardation protein (FMRP) binds specifically to the brain cytoplasmic RNAs BC1/BC200 via a novel RNA-binding motif. J. Biol. Chem 2005, 280, 33403–33410. [Google Scholar]
- Krueger, B.J.; Jeronimo, C.; Roy, B.B.; Bouchard, A.; Barrandon, C.; Byers, S.A.; Searcey, C.E.; Cooper, J.J.; Bensaude, O.; Cohen, E.A.; et al. LARP7 is a stable component of the 7SK snRNP while P-TEFb, HEXIM1 and hnRNP A1 are reversibly associated. Nucleic Acids Res 2008, 36, 2219–2229. [Google Scholar]
- Rinn, J.L.; Kertesz, M.; Wang, J.K.; Squazzo, S.L.; Xu, X.; Brugmann, S.A.; Goodnough, L.H.; Helms, J.A.; Farnham, P.J.; Segal, E.; et al. Functional demarcation of active and silent chromatin domains in human HOX loci by noncoding RNAs. Cell 2007, 129, 1311–1323. [Google Scholar]
- Szell, M.; Bata-Csorgo, Z.; Koreck, A.; Pivarcsi, A.; Polyanka, H.; Szeg, C.; Gaal, M.; Dobozy, A.; Kemeny, L. Proliferating keratinocytes are putative sources of the psoriasis susceptibility-related EDA (extra domain A of fibronectin) oncofetal fibronectin. J. Invest. Dermatol 2004, 123, 537–546. [Google Scholar]
- Ting, K.M.; Rothaupt, D.; McCormick, T.S.; Hammerberg, C.; Chen, G.; Gilliam, A.C.; Stevens, S.; Culp, L.; Cooper, K.D. Overexpression of the oncofetal Fn variant containing the EDA splice-in segment in the dermal-epidermal junction of psoriatic uninvolved skin. J. Invest. Dermatol 2000, 114, 706–711. [Google Scholar]
- Szegedi, K.; Sonkoly, E.; Nagy, N.; Nemeth, I.B.; Bata-Csorgo, Z.; Kemeny, L.; Dobozy, A.; Szell, M. The anti-apoptotic protein G1P3 is overexpressed in psoriasis and regulated by the non-coding RNA, PRINS. Exp. Dermatol 2010, 19, 269–278. [Google Scholar]
- Spector, D.L.; Ochs, R.L.; Busch, H. Silver staining, immunofluorescence, and immunoelectron microscopic localization of nucleolar phosphoproteins B23 and C23. Chromosoma 1984, 90, 139–148. [Google Scholar]
- Feuerstein, N.; Spiegel, S.; Mond, J.J. The nuclear matrix protein, numatrin (B23), is associated with growth factor-induced mitogenesis in Swiss 3T3 fibroblasts and with T lymphocyte proliferation stimulated by lectins and anti-T cell antigen receptor antibody. J. Cell Biol 1988, 107, 1629–1642. [Google Scholar]
- Feuerstein, N.; Chan, P.K.; Mond, J.J. Identification of numatrin, the nuclear matrix protein associated with induction of mitogenesis, as the nucleolar protein B23. Implication for the role of the nucleolus in early transduction of mitogenic signals. J. Biol. Chem 1988, 263, 10608–10612. [Google Scholar]
- Yogev, O.; Saadon, K.; Anzi, S.; Inoue, K.; Shaulian, E. DNA damage-dependent translocation of B23 and p19 ARF is regulated by the Jun N-terminal kinase pathway. Cancer Res 2008, 68, 1398–1406. [Google Scholar]
- Wu, M.H.; Chang, J.H.; Yung, B.Y. Resistance to UV-induced cell-killing in nucleophosmin/B23 over-expressed NIH 3T3 fibroblasts: Enhancement of DNA repair and up-regulation of PCNA in association with nucleophosmin/B23 over-expression. Carcinogenesis 2002, 23, 93–100. [Google Scholar]
- Lee, C.; Smith, B.A.; Bandyopadhyay, K.; Gjerset, R.A. DNA damage disrupts the p14ARF-B23(nucleophosmin) interaction and triggers a transient subnuclear redistribution of p14ARF. Cancer Res 2005, 65, 9834–9842. [Google Scholar]
- Kurki, S.; Peltonen, K.; Latonen, L.; Kiviharju, T.M.; Ojala, P.M.; Meek, D.; Laiho, M. Nucleolar protein NPM interacts with HDM2 and protects tumor suppressor protein p53 from HDM2-mediated degradation. Cancer Cell 2004, 5, 465–475. [Google Scholar]
- Szymanski, M.; Barciszewska, M.Z.; Zywicki, M.; Barciszewski, J. Noncoding RNA transcripts. J. Appl. Genet 2003, 44, 1–19. [Google Scholar]
- Palaniswamy, V.; Moraes, K.C.; Wilusz, C.J.; Wilusz, J. Nucleophosmin is selectively deposited on mRNA during polyadenylation. Nat. Struct. Mol. Biol 2006, 13, 429–435. [Google Scholar]
- Tarapore, P.; Shinmura, K.; Suzuki, H.; Tokuyama, Y.; Kim, S.H.; Mayeda, A.; Fukasawa, K. Thr199 phosphorylation targets nucleophosmin to nuclear speckles and represses pre-mRNA processing. FEBS Lett 2006, 580, 399–409. [Google Scholar]
- Lindstrom, M.S.; Zhang, Y. B23 and ARF: Friends or foes? Cell Biochem. Biophys 2006, 46, 79–90. [Google Scholar]
- Zaric, B.L.; Kambach, C. Reconstitution of recombinant human LSm complexes for biochemical, biophysical, and cell biological studies. Methods Enzymol 2008, 448, 57–74. [Google Scholar]
- Moll, I.; Leitsch, D.; Steinhauser, T.; Blasi, U. RNA chaperone activity of the Sm-like Hfq protein. EMBO Rep 2003, 4, 284–289. [Google Scholar]
- Papadopoulou, C.; Ganou, V.; Patrinou-Georgoula, M.; Guialis, A. HuR-hnRNP interactions and the effect of cellular stress. Mol. Cell Biochem 2013, 372, 137–147. [Google Scholar]
- Beylot-Barry, M.; Lamant, L.; Vergier, B.; de Muret, A.; Fraitag, S.; Delord, B.; Dubus, P.; Vaillant, L.; Delaunay, M.; MacGrogan, G.; et al. Detection of t(2;5)(p23;q35) translocation by reverse transcriptase polymerase chain reaction and in situ hybridization in CD30-positive primary cutaneous lymphoma and lymphomatoid papulosis. Am. J. Pathol. 1996, 149, 483–492. [Google Scholar]
- Amin, M.A.; Matsunaga, S.; Uchiyama, S.; Fukui, K. Depletion of nucleophosmin leads to distortion of nucleolar and nuclear structures in HeLa cells. Biochem. J 2008, 415, 345–351. [Google Scholar]
- Széll, M.; Szegedi, K.; Antal, M.; Németh, I.B.; Sonkoly, E.; Bata-Csorgo, Z.; Dobozy, A.; Kemény, L. PRINS, the Psoriasis Susceptibility Related Non-Coding RNA Interacts with Nucleophosmin and Regulates the Gene Expression of G1P3, an Anti-Apoptotic, Interferon Inducible Protein. Medimond, International Proceeding, Berlin, Germany; 2009; pp. 187–192. [Google Scholar]
- Pivarcsi, A.; Szell, M.; Kemeny, L.; Dobozy, A.; Bata-Csorgo, Z. Serum factors regulate the expression of the proliferation- related genes alpha5 integrin and keratin 1, but not keratin 10, in HaCaT keratinocytes. Arch. Dermatol. Res 2001, 293, 206–213. [Google Scholar]
- Di Carlo, E.; Meazza, R.; Basso, S.; Rosso, O.; Comes, A.; Gaggero, A.; Musiani, P.; Santi, L.; Ferrini, S. Dissimilar anti-tumour reactions induced by tumour cells engineered with the interleukin-2 or interleukin-15 gene in nude mice. J. Pathol 2000, 191, 193–201. [Google Scholar]
- Peter, M.; Nakagawa, J.; Doree, M.; Labbe, J.C.; Nigg, E.A. Identification of major nucleolar proteins as candidate mitotic substrates of cdc2 kinase. Cell 1990, 60, 791–801. [Google Scholar]
- Feuerstein, N. Phosphorylation of numatrin and other nuclear proteins by cdc2 containing CTD kinase cdc2/p58. J. Biol. Chem 1991, 266, 16200–16206. [Google Scholar]
- Takemura, M.; Ohta, N.; Furuichi, Y.; Takahashi, T.; Yoshida, S.; Olson, M.O.; Umekawa, H. Stimulation of calf thymus DNA polymerase alpha activity by nucleolar protein B23. Biochem. Biophys. Res. Commun 1994, 199, 46–51. [Google Scholar]
- Van Hooser, A.A.; Yuh, P.; Heald, R. The perichromosomal layer. Chromosoma 2005, 114, 377–388. [Google Scholar]
- Hernandez-Verdun, D.; Gautier, T. The chromosome periphery during mitosis. Bioessays 1994, 16, 179–185. [Google Scholar]
- Feuerstein, N.; Mond, J.J.; Kinchington, P.R.; Hickey, R.; Karjalainen Lindsberg, M.L.; Hay, I.; Ruyechan, W.T. Evidence for DNA binding activity of numatrin (B23), a cell cycle-regulated nuclear matrix protein. Biochim. Biophys. Acta 1990, 1087, 127–136. [Google Scholar]
- Borer, R.A.; Lehner, C.F.; Eppenberger, H.M.; Nigg, E.A. Major nucleolar proteins shuttle between nucleus and cytoplasm. Cell 1989, 56, 379–390. [Google Scholar]
- Polyanka, H.; Szabo, K.; Tax, G.; Vilmos, T.; Kusz, E.; Ujfaludi, Z.; Imre, B.; Bata-Csorgo, Z.; Lajos, K.; Szell, M. Primary characterization of a novel HPV-E6 oncogene immortalized keratinocyte cell line. J. Invest Dermatol 2011, 131, S70. [Google Scholar]
- Yusufzai, T.M.; Tagami, H.; Nakatani, Y.; Felsenfeld, G. CTCF tethers an insulator to subnuclear sites, suggesting shared insulator mechanisms across species. Mol. Cell 2004, 13, 291–298. [Google Scholar]
- Belso, N.; Szell, M.; Pivarcsi, A.; Kis, K.; Kormos, B.; Kenderessy, A.S.; Dobozy, A.; Kemeny, L.; Bata-Csorgo, Z. Differential expression of D-type cyclins in HaCaT keratinocytes and in psoriasis. J. Invest. Dermatol 2008, 128, 634–642. [Google Scholar]
- Pivarcsi, A.; Bodai, L.; Rethi, B.; Kenderessy-Szabo, A.; Koreck, A.; Szell, M.; Beer, Z.; Bata-Csorgoo, Z.; Magocsi, M.; Rajnavolgyi, E.; et al. Expression and function of toll-like receptors 2 and 4 in human keratinocytes. Int. Immunol 2003, 15, 721–730. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar]
- Chao, A.; Lin, C.Y.; Tsai, C.L.; Hsueh, S.; Lin, Y.Y.; Lin, C.T.; Chou, H.H.; Wang, T.H.; Lai, C.H.; Wang, H.S. Estrogen stimulates the proliferation of human endometrial cancer cells by stabilizing nucleophosmin/B23 (NPM/B23). J. Mol. Med. 2012. [Google Scholar] [CrossRef]
- Wang, Q.Q.; Zhang, Z.Y.; Xiao, J.Y.; Yi, C.; Li, L.Z.; Huang, Y.; Yun, J.P. Knockdown of nucleophosmin induces S-phase arrest in HepG2 cells. Chin. J. Cancer 2011, 30, 853–860. [Google Scholar]
- Itahana, K.; Bhat, K.P.; Jin, A.; Itahana, Y.; Hawke, D.; Kobayashi, R.; Zhang, Y. Tumor suppressor ARF degrades B23, a nucleolar protein involved in ribosome biogenesis and cell proliferation. Mol. Cell 2003, 12, 1151–1164. [Google Scholar]



© 2013 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Szegedi, K.; Göblös, A.; Bacsa, S.; Antal, M.; Németh, I.B.; Bata-Csörgő, Z.; Kemény, L.; Dobozy, A.; Széll, M. Expression and Functional Studies on the Noncoding RNA, PRINS. Int. J. Mol. Sci. 2013, 14, 205-225. https://doi.org/10.3390/ijms14010205
Szegedi K, Göblös A, Bacsa S, Antal M, Németh IB, Bata-Csörgő Z, Kemény L, Dobozy A, Széll M. Expression and Functional Studies on the Noncoding RNA, PRINS. International Journal of Molecular Sciences. 2013; 14(1):205-225. https://doi.org/10.3390/ijms14010205
Chicago/Turabian StyleSzegedi, Krisztina, Anikó Göblös, Sarolta Bacsa, Mária Antal, István Balázs Németh, Zsuzsanna Bata-Csörgő, Lajos Kemény, Attila Dobozy, and Márta Széll. 2013. "Expression and Functional Studies on the Noncoding RNA, PRINS" International Journal of Molecular Sciences 14, no. 1: 205-225. https://doi.org/10.3390/ijms14010205