
Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation


1
Key Laboratory of Carbohydrate Chemistry and Biotechnology, Ministry of Education, School of Biotechnology, Jiangnan University, Wuxi 214122, China
2
School of Food and Biological Engineering, Jiangsu University, Zhenjiang 212013, China
3
National Engineering Laboratory for Cereal Fermentation Technology, Jiangnan University, Wuxi 214122, China
*
Author to whom correspondence should be addressed.
Molecules 2017, 22(6), 955; https://doi.org/10.3390/molecules22060955
Submission received: 22 April 2017
/
Accepted: 5 June 2017
/
Published: 13 June 2017
(This article belongs to the Special Issue Natural Polysaccharides)
Abstract
:Mushrooms are widely distributed around the world and are heavily consumed because of their nutritional value and medicinal properties. Polysaccharides (PSs) are an important component of mushrooms, a major factor in their bioactive properties, and have been intensively studied during the past two decades. Monosaccharide composition/combinations are important determinants of PS bioactivities. This review summarizes: (i) monosaccharide composition/combinations in various mushroom PSs, and their relationships with PS bioactivities; (ii) possible biosynthetic pathways of mushroom PSs and effects of key enzymes on monosaccharide composition; (iii) regulation strategies in PS biosynthesis, and prospects for controllable biosynthesis of PSs with enhanced bioactivities.
1. Introduction
Mushrooms, belonging to the class Basidiomycetes and sometimes Ascomycetes, are a widely distributed food resource on earth and have been consumed because of their nutritional value and medicinal properties for over 2000 years. In addition to their enjoyable flavor and taste, human health was improved by mushrooms due to their nutrients, including digestible proteins, carbohydrates, fiber, vitamins, minerals, and antioxidants [1,2]. A wide variety of bioactive compounds from medicinal mushrooms, which are widely used in eastern Asia, have been studied extensively, and these compounds including polysaccharides, lectins, lactones, terpenoids, and alkaloids have been reviewed [1,3,4]. Among these bioactive compounds, polysaccharides (PSs) with various activities are the main component for the bioactivities of some mushroom species. The diverse activities displayed by PSs include antitumor, immunomodulatory, anti-inflammatory, antinociceptive, antiviral, antioxidative, hypoglycemic, and hepatoprotective effects, as well as protection against chronic radiation stress [1,2,3,4,5].
PSs are divided into two distinct categories, homopolysaccharides (homo-PSs) and heteropolysaccharides (hetero-PSs), on the basis of monosaccharide composition. Both homo-PSs and hetero-PSs may possess homolinkages or heterolinkages in regard to configuration and/or linkage position. In addition to their differing types and sequences of monosaccharide units, hetero-PSs have various types and sequences of glycosidic linkages, resulting in practically limitless structural diversity [5]. Earlier studies suggested that homo-PSs, particularly glucans, are the only PSs with medicinal properties. However, hetero-PSs also display important biological activities [1], and both homo-PSs and hetero-PSs bioactivities are closely related to their specific structural characteristics [5]. The detailed mechanisms of bioactivities related to structure in hetero-PSs remain largely unexplored, although the essential roles of glucans and glycans have been well established. PS composition and combination are clearly correlated with pharmaceutical activities, and this relationship has been the subject of increasing research attention [6,7,8].
The production of chemically and biologically uniform PSs requires knowledge of PS synthetic pathways and regulatory mechanisms. The biosynthetic pathways of PS involve synthesis of nucleotide sugar precursors, assembly of repeating monosaccharide units, and the polymerization process. In PS biosynthetic pathways, enzymes involved in the biosynthetic pathways of nucleotide sugar precursors are closely related with production of mushroom PSs [9,10,11,12]. In submerged culture of Ganoderma lucidum (lingzhi mushroom), overexpression of homologous α-phosphoglucomutase and uridine diphosphate glucose pyrophosphorylase (UDP-glucose pyrophosphorylase) genes enhanced PS production [13,14]. Relationships between the monosaccharide composition of PSs and activities of related enzymes were recently described [15,16].
Numerous studies have addressed the structures and bioactivities of mushroom saccharides, and the relationship between them. Here, we review recent work in this field, with focus on the relationship between monosaccharide composition and the activities of mushroom PSs. We discuss: (i) monosaccharide composition/combinations in various mushroom PSs, and their relationships with bioactivities; (ii) possible biosynthetic pathways of mushroom PSs and effects of key pathway enzymes on monosaccharide composition; (iii) regulation strategies in PS biosynthesis, and prospects for controllable synthesis of PSs with enhanced bioactivities.
2. Monosaccharide Composition, Combinations, and Bioactivities of Mushroom Polysaccharides
Glucan is a well-studied PS and can be produced by different fungi with various terms, such as lentinan, ganoderan, grifolan, and schizophyllan [2,17]. Many studies have addressed the structure–bioactivity relationships of glucans, particularly lentinan [2]. Aside from glucans, no other homo-PSs have been reported. Many mushroom hetero-PSs have been reported in the past two decades, and attempts have been made to elucidate their structure–bioactivity relationships. Few of the PSs have been commercialized, apart from high production or purification costs and low or erratic PS yields, partly because of their unstable chemical characteristics, e.g., monosaccharide composition/combinations that give rise to great variability [17]. The most extensively studied monosaccharide compositions involve glucose (Glc), mannose (Man), galactose (Gal), xylose (Xyl), arabinose (Ara), rhamnose (Rha), and fucose (Fuc). Less frequently encountered components of mushroom PSs include fructose (Fru), ribose (Rib), glucuronic acid (GlcA), galacturonic acid (GalA), N-acetyl-glucosamine (GlcNac, GlcN), and N-acetyl-galactosamine (GalNac, GalN).
2.1. Monosaccharide Composition and Combinations
Monosaccharide compositions of PSs produced by Ascomycetes differ from those of PSs produced by Basidiomycetes. In Ascomycetes, the PSs are mainly hetero-PSs, and Glc, Man, Gal are often presented (Table 1). In a study of Cordyceps (Ophiocordyceps) sinensis PSs, Yan et al. reviewed that monosaccharide composition usually consisted of Glc, Man, and Gal in various molar ratios [4], and the latest research of Cordyceps sinensis also found the similar result [18,19]. In contrast, intracellular PSs (IPSs) from mycelia contain various polyglucans as well as Glc [20,21]. Compositions of other Cordyceps species were generally similar to those of C. sinensis [22,23,24,25], although some other monosaccharides, such as Rha, Xyl, Fru and Ara, were present in C. cicadae and C. kyushuensis [26,27]. In Morchella esculenta, another well-known ascomycete, the PSs contain four or more monosaccharides [28,29,30]. Zhao et al. isolated 52 PSs from fermentation systems and fruiting bodies of Tuber species, and found that they were all composed mainly of Man, Glc, and Gal, suggesting that Tuber PSs have identical chemical compositions [31]. On the other hand, two fractions of T. huidongense PSs were extracted from fruiting bodies, and Glc was the only monosaccharide detected in THWP-2 [32]. In T. rufum PS, Fuc was present in addition to Glc and Gal [33].
In comparison with Ascomycetes, PSs of Basidiomycetes are more complex, particularly in regard to monosaccharide composition and molar ratios of hetero-PSs. Many reviews have focused on chemical characterization of PSs from Basidiomycetes [1,2,3]. However, current knowledge of monosaccharide composition/combinations is not systematic. Table 2 summarizes results of studies during the past few decades of PSs from Ganoderma species, including monosaccharide compositions and molar ratios. The monosaccharide composition of PS from fruiting bodies, mycelia, or fermentation broth of a same fungal strain showed different. To facilitate interpretation of PS properties, Ruthes et al. have reviewed different PSs separately according to the main chain, and found most of the main chains consist solely of Glc, Man, or Gal; however, some PSs contain multiple main chains composed of two or more different monosaccharides in varying combinations, making description difficult [5].
2.2. Bioactivities of Mushroom PSs with Differing Monosaccharide Composition/Combinations
PSs are highly complex molecules, and detailed characterization of specific glycosidic linkages, degree of branching, monosaccharide composition, molecular weight, and chain conformation is essential for elucidating structure—biological activity relationships. Increasing attention has been paid during the past decade to the relationship between PS composition and biological activity. PSs with differing monosaccharide compositions differ in their biological activities and therapeutic effects. In some cases, PSs with high content/ratio monosaccharide, such as Man, Rha, and Fuc, appear to be responsible for the bioactivity. For examples, Man-rich exopolysaccharide (EPS) from Tremella mesenterica [47] and Rha-rich EPS from Lactobacillus rhamnosus [48] stimulate the immune system through receptors located on macrophage. l-Fuc-enriched EPS is used as a skin moisturizing agent in the cosmetic industry because l-Fuc displays anticancer and anti-inflammatory activities [49]. PSs containing high levels of uronic acid display significant antioxidant activity, and may be attributed to the presence of the functional group -COOH [7]. Functional groups that act as efficient electron or hydrogen donors are associated to the antioxidant activity of certain PSs [50].
Man is a typical component of many bioactive PSs. Most of these PSs also confer virulence against pathogenic species. Human cells are also able to recognize such carbohydrates through Man receptors, thus stimulating cytokine production [51,52]. The high molar proportion of Man in Inonotus obliquus PSs may enhance antioxidant activity [53,54]. Kim et al. reported an anti-cancer effect by endo-PS fractions from cultivated mycelia of I. obliquus with Man as the major component [55]. Chen et al. showed that the Man component of Tremella mesenterica EPS stimulated nitric oxide (NO) and cytokine (IL-6 and TNF-α) production in RAW 264.7 macrophage cells [47]. The major pharmacologically-active substance in Tremella is the PS glucuronoxylomannan, composed primarily of Man [56]. Biological activity of certain mushroom PSs is related to content of other monosaccharides such as rhamnose (Table 3). Rha as a major monosaccharide component may enhance antioxidant capacity, and play a role in radical-scavenging ability [34,57]. Biological activities displayed by active components in mushroom PSs are summarized in Table 3.
Many researchers have used a model, based on relationships between monosaccharide composition ratios and biological activities, to provide theoretical support for applications of PS from natural products. Lo et al. observed a strong correlation between monosaccharide composition and ferrous ion chelating ability, and showed that Rha level in PSs was the dominant component in modulation of the chelating response variable [6]. In another study, Lo et al. used statistical methods (multiple linear regression analysis, principal component analysis (PCA), factor analysis) to examine relationships between monosaccharide composition ratios and macrophage stimulatory activities of regionally different strains of Lentinula edodes [62]. Ara, Xyl, Man, and Gal were most strongly related to macrophage stimulatory activities. When additional PCA and factor analysis methods were applied to the same monosaccharide composition ratio data, Ara, Xyl, Man, and Gal compositions were again found to be the most important. Glc, although present as a major monosaccharide component in all strains and presumably forming the backbone of PS structures, was not a major determinant factor for structural characteristics or in vitro macrophage stimulatory activities [62]. Li et al. used multiple linear regression (MLR), support vector machine (SVM), and artificial neural network (ANN) methods to establish an optimum quantitative structure–activity relationship (QSAR) model for the antioxidant activity of PSs. In MLR and ANN models, Ara and GalA played major roles in determining EC50 of DPPH (1,1-Diphenyl-2-picrylhydrazyl)-scavenging activity, and Fuc, Rha, and uronic acid had the greatest effects on the hydroxyl radical scavenging activity of PSs. Such measurement methods are useful for predicting performance and evaluating relationships between PS properties and biological activities [7].
Monosaccharide composition/combinations clearly play an important role, and are strongly correlated with biological activity. Chemically and biologically uniform PSs with specific monosaccharide composition/combinations could be used for controllable biosynthesis. However, we know little about the detailed mechanisms of the bioactive effects relating to the monosaccharide composition of PS, although the bioactivities of PSs are well established.
3. Biosynthetic Pathways and Key Enzymes of Mushroom Polysaccharides
3.1. Biosynthetic Pathways
Fungal PSs are involved in a wide variety of cellular functions, including energy storage, cell wall structure, cell–cell interactions and signaling, host–pathogen interaction, and protein glycosylation [5]. The PSs are synthesized using intracellular nucleotide sugars at the membrane level to construct the main structure of cell walls or form a gel-like matrix on the hyphal surface. In submerged fermentation, gel-like PSs may be further excreted into the growth medium and are referred to as EPSs [63]. Currently, the biosynthetic pathways of mushroom PSs remain unclear because of the inadequate knowledge of relative enzymes and its functions in pathway, which should be proved by gene cloning and genetic transformation.
However, a simplified biosynthetic pathway of mushroom PSs has been constructed based on identification of intermediate compounds and activities of synthesis-related enzymes by various researchers [11,16,64]. Approaches such as gene expression [13,14] and RNAi-mediated gene silencing [64] have also been applied for study of the biosynthetic pathway. In Figure 1, we propose a more detailed biosynthetic pathway based on the previous publications. In this model, the biosynthetic pathway of nucleotide sugar precursors in mushroom PSs is similar to that in plants [65], providing a basis for analysis of other metabolic pathways in mushroom PSs. The genomes of Ganoderma lucidum [66], Antrodia cinnamomea [67], Lentinula edodes [68], and Lignosus rhinocerotis [69] have been published. These genomic studies provide insights into the genetic basis of reported medicinal properties of PSs, and a platform for further characterization of putative bioactive PSs and pathway enzymes.
3.2. Effects of Key Enzymes on PS Production and Monosaccharide Composition
Enzymes in the PS synthesis pathways of mushrooms play an important role in PS production and regulation of monosaccharide composition. These enzymes include Phosphoglucose isomerase, α-phosphoglucomutase, UDP-Glc pyrophosphorylase and so on. Phosphoglucose isomerase (PGI; EC 5.3.1.9) catalyzes the interconversion of Glc-6-phosphate and Fru-6-phosphate, and plays key roles in glycolysis and gluconeogenesis pathways. PGI catalyzes the reaction that generates Fru-6-phosphate, the precursor of GDP-Man in PS synthesis [16]. Thus, PGI activity level determines the direction of Glc-6-phosphate conversion. PGI may also control PS synthesis to some extent. Zhu et al. observed a strong correlation between PGI activity level and the amount of IPS produced [11]. Yang et al. showed that the addition of coix lacryma-jobi oil (CLO) to culture medium enhanced EPS and IPS production by directly affecting synthesis levels of PGI and α-phosphoglucomutase (PGM) at different stages [70]. In a study by Wei et al., higher sucrose level increased PGM activity, and also enhanced PS production through inhibition of PGI activity [12]. Similar results were reported by Tang and Zhong [71].
α-phosphoglucomutase (PGM; EC 5.4.2.2), a key enzyme in carbohydrate metabolism, catalyzes the reversible transfer of a phosphate group between C-1 and C-6. PGM converts Glc-6-P and Glc-1-P; Glc-6-phosphate enters a glycolysis pathway to yield energy, and Glc-1-phosphate is the precursor of sugar nucleotides (e.g., UDP-Glc, dTDP-Rha, UDP-Gal) in PSs [10,16]. In the study by Tang and Zhong, PGM activity was correlated with the amount of EPS produced by G. lucidum [71]. Xu et al. investigated the effects of PGM gene overexpression on PS production and transcription levels of three genes, and demonstrated that the PGM gene is an important regulatory gene in PS biosynthesis in G. lucidum [14]. PGM gene overexpression in submerged culture resulted in increased PS production and upregulation of downstream genes encoding the enzymes involved in PS biosynthesis, including PGM, UDP-Glc pyrophosphorylase (UGP), and β-1,3-glucan synthase (GLS) [14]. PGM activity was also affected by the addition of coixenolide during PS production in G. lucidum [10]. In a submerged culture of C. militaris, PGM was the key enzyme for regulation of IPS biosynthesis [11].
UDP-Glc pyrophosphorylase (UGP; also known as UGPase or UTP: Glc-1-phosphate uridylyltransferase; EC 2.7.7.9), a member of the Pfam glycosyl transferase family (PF01702), catalyzes the reversible interconversion of Glc-1-phosphate and UDP-Glc [64]. UGP is a key enzyme involved in carbohydrate metabolism and cell wall biosynthesis [72]. Studies by Liao et al. and Jiang et al. showed that the UGP gene (ugp) is associated with the synthesis of LPS and EPS surface structures, because its substrate/product UDP-Glc is the precursor of these PSs [73,74]. Li et al. cloned the ugp gene from G. lucidum, and examined the effect of RNAi silencing of ugp on PS synthesis. In liquid culture, EPS concentration in ugp-silenced strains was much lower (30–56%) than that in wild type (WT) strains. In contrast, IPS concentration did not differ significantly between ugp-silenced and WT strains [64]. Ji et al. showed that overexpression of homologous ugp in submerged culture G. lucidum enhanced PS production; maximal IPS and EPS levels in the ugp-overexpressing strains were respectively 42% and 36% higher than in WT strains [13]. Zhu et al. found that higher UGP activity was correlated with higher IPS biosynthesis rate in Cordyceps militaris [11], suggesting an important role of UGP in PS biosynthesis. However, a controversial conclusion was found by Zhou et al., Wei et al., and Yang et al.: that there was no significant association of UGP activity with PS biosynthesis [10,12,70], which may be attributed to the different strains and methods. Therefore, the exact reason still requires further research.
Enzyme activities of glucokinase (GK) [16], UDPG-dehydrogenase (UGDG) [11], and dTDP-Glc pyrophosphorylase (dTDP-GPP) [10,70] have also been investigated. However, these studies showed no notable association between PS biosynthesis and activities of these enzymes.
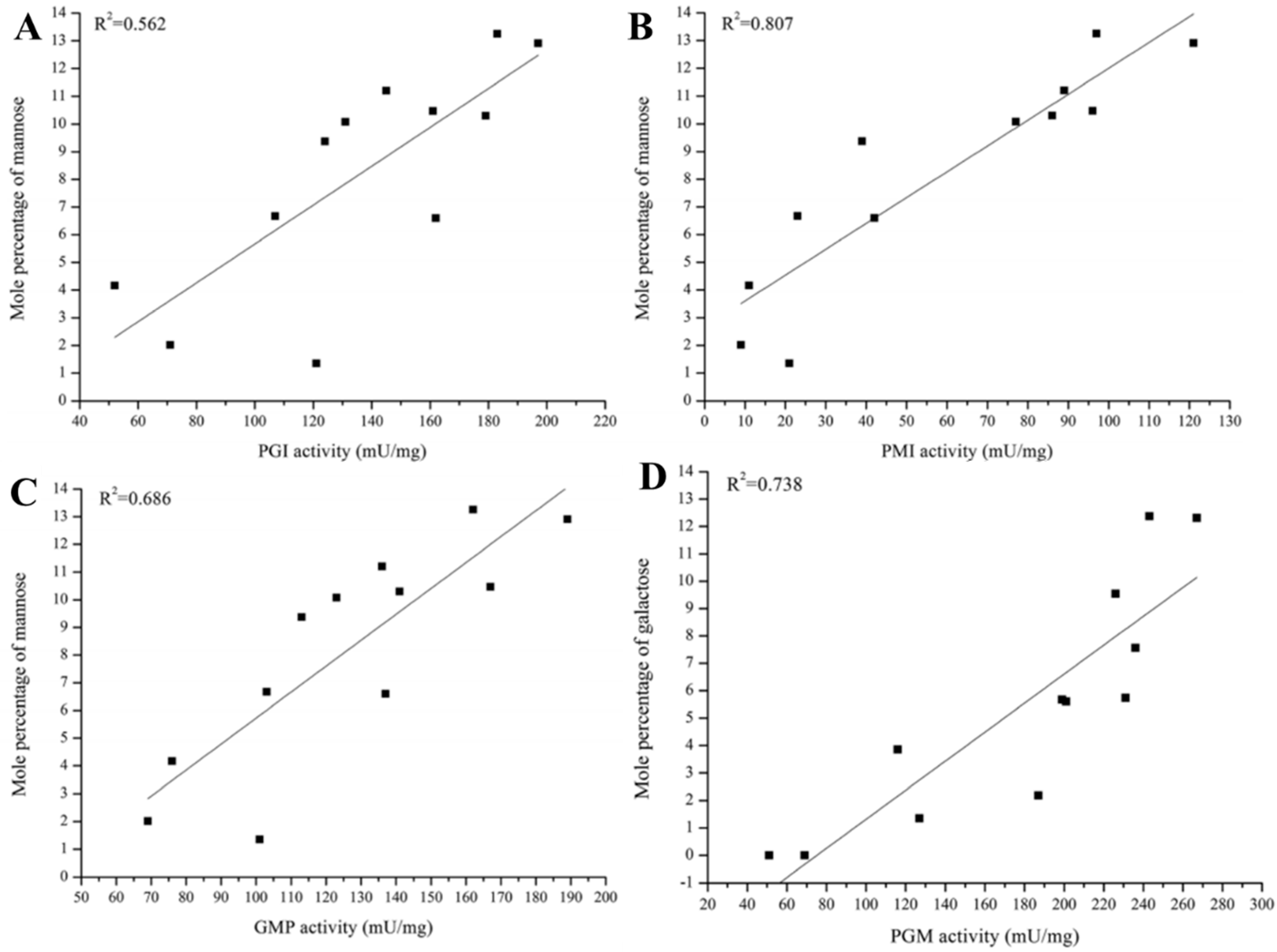
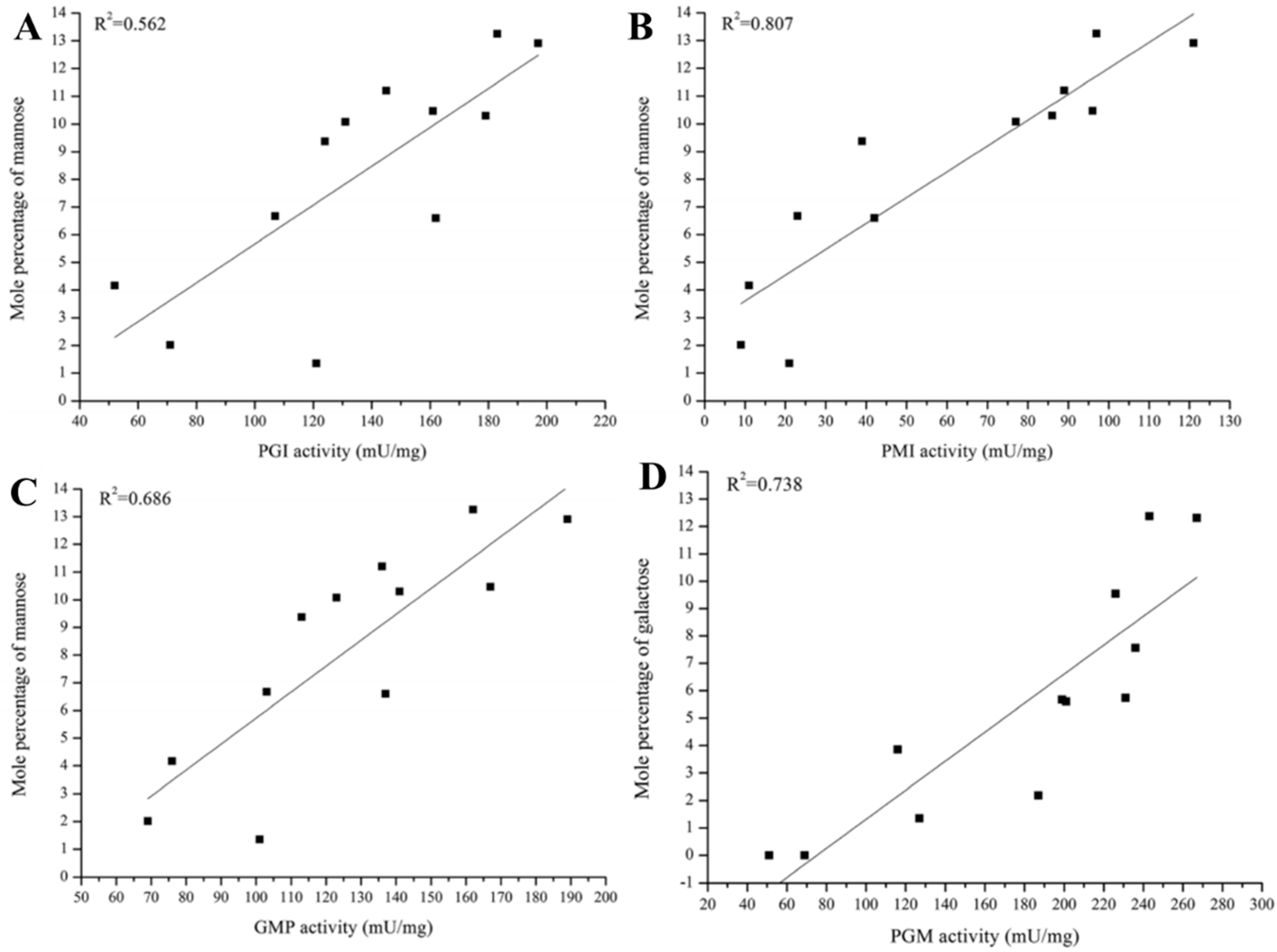
Relationships between the monosaccharide composition of PS and the activity of related enzymes were recently studied using a correlation coefficient (R2) based on data obtained under various culture and carbon source conditions [15,16]. Changes in PGM, PGI, and phosphomannose isomerase (PMI) activities were strongly correlated with Gal and Man mole percentages in G. lucidum EPS, based on R2 values obtained under various culture temperatures and initial pH values [15]. In contrast, under various carbon source conditions, PGI and PMI activities were correlated only with Man mole percentage, and PGM activity was correlated only with Gal mole percentage (Figure 2) [16]. GDP-Man pyrophosphorylase (GMP) activity, whose R2 with mole percentages of all monosaccharides under various culture conditions was low (<0.5), showed a stronger correlation with Man mole percentage under carbon source conditions. Taken together, results of these studies indicate that PGM, PGI, and PMI regulate the mole percentages of Gal and Man in EPS under various culture and carbon source conditions, whereas GMP regulates the mole percentage of Man only in response to carbon source. Key enzymes regulating monosaccharide synthetic pathways in G. lucidum EPS can thus be divided into two groups, based on responses to culture conditions and culture medium. Mole percentages of other monosaccharides (Glc, Ara, Fuc) and activities of other tested enzymes under the various conditions showed no significant correlations (R2 < 0.5) [16].
4. Factors Regulating Mushroom Polysaccharide Synthesis
Monosaccharide composition/combinations significantly affect PS bioactivity. Increasing research attention has been paid to regulation of synthesis of PS with stronger bioactivities. Variability in monosaccharide composition/combinations among mushroom PSs may result from strain variations, developmental stage, culture method and conditions, medium composition, extraction method, and even drying method [60,75,76,77,78,79]. In industrial-scale operations, submerged fermentation procedures have been successfully applied for mushroom cultivation during the past two decades. Submerged culture has advantages over traditional solid-state culture in terms of higher mycelial biomass production of bioactive metabolites in a compact space, shorter times, and reduced contamination [75,80]. This is a useful alternative approach for efficient production of IPS and EPS.
Structural parameters (monosaccharide composition, degree of branching, molecular weight, molecular weight distribution, chain conformation) are affected by the fermentation medium [81,82]. Among the various nutrients present in the culture medium, carbon source is an important factor determining the monosaccharide content of PS. In a study by Peng et al., the use of sugar as a carbon source was associated with an excess of the same monosaccharide in G. lucidum EPS. With mixed carbon sources, the main containing monosaccharides of G. lucidum EPS were Glc, Gal, and Man, and the combined mole percentage of EPS monosaccharides was the same as carbon sources used in culture medium. [16]. In Pleurotus pulmonarius submerged culture, each EPS sample had Man, Gal, and Glc as major monosaccharide components. Water-insoluble polysaccharides (IEPS) fractions obtained from Glc- and Xyl-based culture media (termed IEPS-Glc and IEPS-Xyl, respectively) contained Glc as the sole monosaccharide component. Surprisingly, these glucans were not isolated from Gal- and Ara-based culture media [83]. In Paecilomyces hepiali, a high percentage of Man in EPS was observed under Glc carbon source, and a high percentage of Glc was observed under Man carbon source [63]. These findings indicate that monosaccharide composition and mole percentages of PS are determined by carbon source in mushroom submerged culture. In a study of Cordyceps sinensis CS-HK1 mycelial cultures, the monosaccharide composition of EPS isolated on different days varied according to carbon source. The monosaccharides Glc, Gal, and Man were found in all EPS samples, with Glc being the major component (50–75% of total). The addition of Gal or Man to the culture medium resulted in no notable increase (in some cases, even a slight decrease) in their content in EPS [84].
In a study of Tuber melanosporum by Zhao et al., the addition of metal ions to culture medium affected IPS and EPS biosynthetic pathways. Gal, Glc, and Man were the major monosaccharides in both IPS and EPS. The addition of Mg2+ resulted in a maximal Man content (27.6%) in IPS, but no significant change in Gal or Glc content in IPS or EPS. Both IPS and EPS were acidic hetero-PSs containing Man, Gal, and Glc [81]. The metal ions may be a cofactor in enzymatic reactions in the PS biosynthesis pathway and alter some key enzyme activities, which lead to different molar ratios of the monosaccharide components [81]. Alternatively, addition of metal ions is an effective method to regulate the structural properties of polysaccharide.
5. Conclusions and Future Perspectives
A steadily increasing number of PSs have been extracted from a wide variety of mushroom species during the past decade. In many cases, their structural features and bioactivities have been elucidated through the application of advanced analytical techniques. However, there is no uniformity or predictability in the structural features or functional characteristics of bioactive PSs. The bioactivity mechanisms, biosynthetic pathways, and productivities of the PSs are also highly variable and confusing to researchers. Monosaccharide composition/combinations is an important characteristic of PSs, and there is a clear relationship between structure and bioactivity. Continued investigation along this line will help to identify the most effective structures, and lead to improved regulation strategies in PS biosynthesis and prospects for the controllable synthesis of PSs with enhanced bioactivities.
Acknowledgments
Funding for this study was provided by the National Natural Science Foundation of China (31271918), Open Project Program of Key Laboratory of Carbohydrate Chemistry and Biotechnology (KLCCB-KF201205) and Qing Lan Project. The authors are grateful to S. Anderson for English editing of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- He, X.R.; Wang, X.X.; Fang, J.C.; Chang, Y.; Ning, N.; Guo, H.; Huang, L.H.; Huang, X.Q.; Zhao, Z.F. Polysaccharides in Grifola frondosa mushroom and their health promoting properties: A review. Int. J. Biol. Macromol. 2017, 101, 910–921. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.F.; Yan, H.D.; Tang, J.; Chen, J.; Zhang, X.W. Polysaccharides in lentinus edodes: Isolation, structure, immunomodulating activity and future prospective. Crit. Rev. Food Sci. Nutr. 2014, 54, 474–487. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.C.; Heleno, S.A.; Reis, F.S.; Stojkovic, D.; Queiroz, M.J.; Vasconcelos, M.H.; Sokovic, M. Chemical features of Ganoderma polysaccharides with antioxidant, antitumor and antimicrobial activities. Phytochemistry 2015, 114, 38–55. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.K.; Wang, W.Q.; Wu, J.Y. Recent advances in Cordyceps sinensis polysaccharides: Mycelial fermentation, isolation, structure, and bioactivities: A review. J. Funct. Foods 2014, 6, 33–47. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. Mushroom heteropolysaccharides: A review on their sources, structure and biological effects. Carbohydr. Polym. 2016, 136, 358–375. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.T.; Cheng, A.C.; Chiu, K.H.; Tsay, P.K.; Jen, J.F. Correlation evaluation of antioxidant properties on the monosaccharide components and glycosyl linkages of polysaccharide with different measuring methods. Carbohydr. Polym. 2011, 86, 320–327. [Google Scholar] [CrossRef]
- Li, Z.M.; Nie, K.Y.; Wang, Z.J.; Luo, D.H. Quantitative structure activity relationship models for the antioxidant activity of polysaccharides. PLoS ONE 2016, 11, e0163536. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Hu, Y.; Quan, L.L. Production of bioactive polysaccharides by Inonotus obliquus under submerged fermentation supplemented with lignocellulosic biomass and their antioxidant activity. Bioprocess Biosyst. Eng. 2014, 37, 2483–2492. [Google Scholar] [CrossRef] [PubMed]
- Li, H.J.; Zhang, D.H.; Yue, T.H.; Jiang, L.X.; Yu, X.; Zhao, P.; Li, T.; Xu, J.W. Improved polysaccharide production in a submerged culture of Ganoderma lucidum by the heterologous expression of vitreoscilla hemoglobin gene. J. Biotechnol. 2016, 217, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.B.; Bi, P.Y.; Wu, X.T.; Huang, F.R.; Yang, H.L. Improved polysaccharide production in submerged culture of Ganoderma lucidum by the addition of coixenolide. Appl. Biochem. Biotechnol. 2014, 172, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.Y.; Liu, X.C.; Dong, F.Y.; Guo, M.Z.; Wang, X.T.; Wang, Z.; Zhang, Y.M. Influence of fermentation conditions on polysaccharide production and the activities of enzymes involved in the polysaccharide synthesis of Cordyceps militaris. Appl. Microbiol. Biotechnol. 2016, 100, 3909–3921. [Google Scholar] [CrossRef] [PubMed]
- Wei, Z.H.; Liu, L.; Guo, X.F.; Li, Y.J.; Hou, B.C.; Fan, Q.L.; Wang, K.X.; Luo, Y.; Zhong, J.J. Sucrose fed-batch strategy enhanced biomass, polysaccharide, and ganoderic acids production in fermentation of Ganoderma lucidum 5.26. Bioprocess Biosyst. Eng. 2016, 39, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Ji, S.L.; Liu, R.; Ren, M.F.; Li, H.J.; Xu, J.W. Enhanced production of polysaccharide through the overexpression of homologous uridine diphosphate glucose pyrophosphorylase gene in a submerged culture of lingzhi or reishi medicinal mushroom, Ganoderma lucidum (higher basidiomycetes). Int. J. Med. Mushrooms 2015, 17, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.W.; Ji, S.L.; Li, H.J.; Zhou, J.S.; Duan, Y.Q.; Dang, L.Z.; Mo, M.H. Increased polysaccharide production and biosynthetic gene expressions in a submerged culture of Ganoderma lucidum by the overexpression of the homologous α-phosphoglucomutase gene. Bioprocess Biosyst. Eng. 2015, 38, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Qiao, S.K.; Xu, Z.H.; Guang, F.; Ding, Z.Y.; Gu, Z.H.; Zhang, L.; Shi, G.Y. Effects of culture conditions on monosaccharide composition of Ganoderma lucidum exopolysaccharide and on activities of related enzymes. Carbohydr. Polym. 2015, 133, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, J.; Liu, Y.; Xu, Z.H.; Wu, J.Y.; Ding, Z.Y.; Gu, Z.H.; Zhang, L.; Shi, G.Y. Effects of mixed carbon sources on galactose and mannose content of exopolysaccharides and related enzyme activities in Ganoderma lucidum. RSC Adv. 2016, 6, 39284–39291. [Google Scholar] [CrossRef]
- Giavasis, I. Bioactive fungal polysaccharides as potential functional ingredients in food and nutraceuticals. Curr. Opin. Biotechnol. 2014, 26, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Q.; Nie, S.P.; Kan, L.J.; Chen, H.H.; Cui, S.W.; Phillips, A.O.; Phillips, G.O.; Xie, M.Y. Comparison of structural features and antioxidant activity of polysaccharides from natural and cultured Cordyceps sinensis. Food Sci. Biotechnol. 2017, 26, 55–62. [Google Scholar] [CrossRef]
- Cheong, K.L.; Meng, L.Z.; Chen, X.Q.; Wang, L.Y.; Wu, D.T.; Zhao, J.; Li, S.P. Structural elucidation, chain conformation and immuno-modulatory activity of glucogalactomannan from cultured Cordyceps sinensis fungus UM01. J. Funct. Foods 2016, 25, 174–185. [Google Scholar] [CrossRef]
- Yan, J.K.; Wang, W.Q.; Li, L.; Wu, J.Y. Physiochemical properties and antitumor activities of two α-glucans isolated from hot water and alkaline extracts of Cordyceps (CS-HK1) fungal mycelia. Carbohydr. Polym. 2011, 85, 753–758. [Google Scholar] [CrossRef]
- Wu, Y.L.; Sun, C.R.; Pan, Y.J. Structural analysis of a neutral (1→3), (1→4)-beta-d-glucan from the mycelia of Cordyceps sinensis. J. Nat. Prod. 2005, 68, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Smiderle, F.R.; Sassaki, G.L.; van Griensven, L.J.L.D.; Iacomini, M. Isolation and chemical characterization of a glucogalactomannan of the medicinal mushroom Cordyceps militaris. Carbohydr. Polym. 2013, 97, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.P.; Duan, Y.Q.; Yang, W.Y.; Zhang, H.H.; Li, C.Z.; Zhang, J.X. Structural elucidation and immunostimulatory activity of polysaccharide isolated by subcritical water extraction from Cordyceps militaris. Carbohydr. Polym. 2016, 157, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.C.; Zhu, Z.Y.; Tang, Y.L.; Wang, M.F.; Wang, Z.; Liu, A.J.; Zhang, Y.M. Structural properties of polysaccharides from cultivated fruit bodies and mycelium of Cordyceps militaris. Carbohydr. Polym. 2016, 142, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.K.; Gu, G.F.; Hao, L.M.; Jin, Z.N.; Wang, X. Characterization and in vitro antioxidant activity of a polysaccharide from Cordyceps sobolifera. J. Food Process. Preserv. 2016, 40, 447–452. [Google Scholar] [CrossRef]
- Sharma, S.K.; Gautam, N.; Atri, N.S. Optimized extraction, composition, antioxidant and antimicrobial activities of exo and intracellular polysaccharides from submerged culture of Cordyceps cicadae. BMC Complement. Altern. Med. 2015, 15, 446. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Yin, Q.; Han, T.; Zhao, Y.; Su, J.; Li, M.; Ling, J. Purification and antioxidant effect of novel fungal polysaccharides from the stroma of Cordyceps kyushuensis. Ind. Crops Prod. 2015, 69, 485–491. [Google Scholar] [CrossRef]
- Liu, C.; Sun, Y.H.; Mao, Q.; Guo, X.L.; Li, P.; Liu, Y.; Xu, N. Characteristics and antitumor activity of Morchella esculenta polysaccharide extracted by pulsed electric field. Int. J. Mol. Sci. 2016, 17, 986. [Google Scholar] [CrossRef] [PubMed]
- Li, S.H.; Gao, A.; Dong, S.; Chen, Y.; Sun, S.; Lei, Z.F.; Zhang, Z.Y. Purification, antitumor and immunomodulatory activity of polysaccharides from soybean residue fermented with Morchella esculenta. Int. J. Biol. Macromol. 2017, 96, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.L.; Chen, Y.; Wang, S.S.; Kai, G.Q.; Fang, Y.M. Isolation, partial characterisation and immunomodulatory activities of polysaccharide from Morchella esculenta. J. Sci. Food Agric. 2011, 91, 2180–2185. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, X.H.; Li, H.M.; Wang, S.H.; Chen, T.; Yuan, Z.P.; Tang, Y.J. Isolation and characterization of polysaccharides with the antitumor activity from Tuber fruiting bodies and fermentation system. Appl. Microbiol. Biotechnol. 2014, 98, 1991–2002. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.J.; Zhang, S.Q.; Ran, C.X.; Wang, L.S.; Kan, J.Q. Extraction, characterization and antioxidant activity of water-soluble polysaccharides from Tuber huidongense. Int. J. Biol. Macromol. 2016, 91, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Pattanayak, M.; Samanta, S.; Maity, P.; Manna, D.K.; Sen, I.K.; Nandi, A.K.; Panda, B.C.; Chattopadhyay, S.; Roy, S.; Sahoo, A.K. Polysaccharide of an edible Truffle tuber rufum: Structural studies and effects on human lymphocytes. Int. J. Biol. Macromol. 2016, 95, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.J.; Meng, G.Y.; Zhai, G.Y.; Yang, Y.H.; Zhao, H.J.; Jia, L. Extraction, characteristisation and antioxidant activity of polysaccharides of spent mushroom compost of Ganoderma lucidum. Int. J. Biol. Macromol. 2016, 82, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.W.; Feng, M.Y.; Zhai, X.F.; Hu, M.H.; You, L.J.; Luo, W.; Zhao, M.M. Optimization for the extraction of polysaccharides from Ganoderma lucidum and their antioxidant and antiproliferative activities. J. Taiwan Inst. Chem. Eng. 2013, 44, 886–894. [Google Scholar] [CrossRef]
- Liu, W.; Wang, H.Y.; Pang, X.B.; Yao, W.B.; Gao, X.D. Characterization and antioxidant activity of two low-molecular-weight polysaccharides purified from the fruiting bodies of Ganoderma lucidum. Int. J. Biol. Macromol. 2010, 46, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Zhang, J.S.; Tang, Q.J.; Yang, Y.; Guo, Q.B.; Wang, Q.; Wu, D.; Cui, S.W. Physicochemical characterization of a high molecular weight bioactive β-d-glucan from the fruiting bodies of Ganoderma lucidum. Carbohydr. Polym. 2014, 101, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Q.; Fang, L.; Zhang, K.C. Structure and bioactivities of a galactose rich extracellular polysaccharide from submergedly cultured Ganoderma lucidum. Carbohydr. Polym. 2007, 68, 323–328. [Google Scholar] [CrossRef]
- Ye, L.B.; Zhang, J.S.; Ye, X.J.; Tang, Q.J.; Liu, Y.F.; Gong, C.Y.; Du, X.J.; Pan, Y.J. Structural elucidation of the polysaccharide moiety of a glycopeptide (GLPCW-II) from Ganoderma lucidum fruiting bodies. Carbohydr. Res. 2008, 343, 746–752. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Wang, L.Q.; Chen, C.H.; Teng, B.S.; Wang, C.D.; Xu, Z.X.; Hu, B.W.; Zhou, P. Structure characterization of a novel neutral polysaccharide isolated from Ganoderma lucidum fruiting bodies. Food Chem. 2012, 135, 1097–1103. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zhang, Z.Y.; Yang, Y.N. Antioxidant and immunoregulatory activity of Ganoderma lucidum polysaccharide (GLP). Carbohydr. Polym. 2013, 95, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Nie, S.P.; Yin, J.Y.; Wang, Y.X.; Xie, M.Y. Structural characterization of a heterogalactan purified from fruiting bodies of Ganoderma atrum. Food Hydrocoll. 2014, 36, 339–347. [Google Scholar] [CrossRef]
- Zhang, H.; Li, W.J.; Nie, S.P.; Chen, Y.; Wang, Y.X.; Xie, M.Y. Structural characterisation of a novel bioactive polysaccharide from Ganoderma atrum. Carbohydr. Polym. 2012, 88, 1047–1054. [Google Scholar] [CrossRef]
- Han, X.Q.; Chan, B.C.L.; Yu, H.; Yang, Y.H.; Hu, S.Q.; Ko, C.H.; Dong, C.X.; Wong, C.K.; Shaw, P.C.; Fung, K.P.; et al. Structural characterization and immuno-modulating activities of a polysaccharide from Ganoderma sinense. Int. J. Biol. Macromol. 2012, 51, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.F.; Zhang, L.N.; Zeng, F.B.; Xu, Y.X. Structure and antitumor activity of extracellular polysaccharides from mycelium. Carbohydr. Polym. 2003, 54, 297–303. [Google Scholar] [CrossRef]
- Wang, C.L.; Pi, C.C.; Kuo, C.W.; Zhuang, Y.J.; Khoo, K.H.; Liu, W.H.; Chen, C.J. Polysaccharides purified from the submerged culture of Ganoderma formosanum stimulate macrophage activation and protect mice against listeria monocytogenes infection. Biotechnol. Lett. 2011, 33, 2271. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.Y.; Hsu, T.H.; Lin, F.Y.; Lai, H.H.; Wu, J.Y. Effects on cytokine-stimulating activities of eps from Tremella mesenterica with various carbon sources. Food Chem. 2006, 99, 92–97. [Google Scholar] [CrossRef]
- Chabot, S.; Yu, H.L.; Léséleuc, L.D.; Cloutier, D.; Calsteren, M.R.V.; Lessard, M.; Roy, D.; Lacroix, M.; Oth, D. Exopolysaccharides from Lactobacillus rhamnosus RW-9595M stimulate TNF, IL-6 and IL-12 in human and mouse cultured immunocompetent cells, and IFN-γ in mouse splenocytes. Dairy Sci. Technol. 2001, 81, 683–697. [Google Scholar] [CrossRef]
- Cescutti, P.; Kallioinen, A.; Impallomeni, G.; Toffanin, R.; Pollesello, P.; Leisola, M.; Eerikäinen, T. Structure of the exopolysaccharide produced by Enterobacter amnigenus. Carbohydr. Res. 2005, 340, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Li, W.; Rui, X.; Li, T.; Chen, X.H.; Jiang, M.; Dong, M.S. Chemical modification, characterization and bioactivity of a released exopolysaccharide (r-EPS1) from Lactobacillus plantarum 70810. Glycoconj. J. 2015, 32, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Netea, M.G.; Brown, G.D.; Kullberg, B.J.; Gow, N.A. An integrated model of the recognition of Candida albicans by the innate immune system. Nat. Rev. Microbiol. 2008, 6, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Ostrosky-Zeichner, L.; Vitale, R.G.; Nucci, M. New serological markers in medical mycology: (1,3)-β-d-glucan and aspergillus galactomannan. Infectio 2012, 16, 59–63. [Google Scholar] [CrossRef]
- Xiang, Y.L.; Xu, X.Q.; Li, J. Chemical properties and antioxidant activity of exopolysaccharides fractions from mycelial culture of Inonotus obliquus in a ground corn stover medium. Food Chem. 2012, 134, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Yan, M.C.; Zhu, J.W.; Xu, X.Q. Enhancement of exo-polysaccharide production and antioxidant activity in submerged cultures of Inonotus obliquus by lignocellulose decomposition. J. Ind. Microbiol. Biotechnol. 2011, 38, 291–298. [Google Scholar] [CrossRef] [PubMed]
- Yong, O.K.; Park, H.W.; Kim, J.H.; Lee, J.Y.; Moon, S.H.; Shin, C.S. Anti-cancer effect and structural characterization of endo-polysaccharide from cultivated mycelia of Inonotus obliquus. Life Sci. 2006, 79, 72–80. [Google Scholar]
- Reshetnikov, S.V.; Wasser, S.P.; Nevo, E.; Duckman, I.; Tsukor, K. Medicinal value of the genus Tremella pers. (heterobasidiomycetes) (review). Int. J. Med. Mushrooms 2000, 2, 26. [Google Scholar] [CrossRef]
- Zhao, M.; Chen, Z.; Xia, G.; Cui, F.Y.; Zhang, J.J.; Jia, M.S.; Jia, S.H.; Jia, L. Enzymatic and acidic degradation effect on intracellular polysaccharide of Flammulina velutipes SF-08. Int. J. Biol. Macromol. 2015, 73, 236–244. [Google Scholar]
- Liu, M.; Jing, H.J.; Zhang, J.J.; Che, G.; Zhou, M.; Gao, Z.; Li, S.S.; Ren, Z.Z.; Hao, L.; Liu, Y. Optimization of mycelia selenium polysaccharide extraction from Agrocybe cylindracea SL-02 and assessment of their antioxidant and anti-ageing activities. PLoS ONE 2016, 11, e0160799. [Google Scholar] [CrossRef] [PubMed]
- Im, S.; Wang, W.; Lee, C.K.; Lee, Y.N. Activation of macrophages by exopolysaccharide produced by MK1 bacterial strain isolated from neungee mushroom, sarcodon aspratus. Immune Netw. 2010, 10, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Lei, M.; Sun, S.S.; Li, R.; Shen, Z.P.; Wang, P.; Jiang, X.L. Antioxidant activity of polysaccharides produced by Hirsutella sp. And relation with their chemical characteristics. Carbohydr. Polym. 2015, 117, 452–457. [Google Scholar]
- He, P.X.; Li, F.L.; Huang, L.N.; Xue, D.D.; Liu, W.; Xu, C.P. Chemical characterization and antioxidant activity of polysaccharide extract from spent mushroom substrate of Pleurotus eryngii. J. Taiwan Inst. Chem. Eng. 2016, 69, 48–53. [Google Scholar] [CrossRef]
- Lo, T.C.T.; Yi, H.J.; Chao, A.L.J.; Cheng, A.C. Use of statistical methods to find the polysaccharide structural characteristics and the relationships between monosaccharide composition ratio and macrophage stimulatory activity of regionally different strains of Lentinula edodes. Anal. Chim. Acta 2007, 584, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.W.; Yang, Z.J.; Dan, G.; Fan, J.L.; Dai, Z.Q.; Wang, X.Q.; Bing, H.; Hong, Y.; Abid, M.; Zeng, X.X. Influences of carbon sources on the biomass, production and compositions of exopolysaccharides from Paecilomyces hepiali HN1. Biomass Bioenergy 2014, 67, 260–269. [Google Scholar] [CrossRef]
- Li, M.J.; Chen, T.X.; Gao, T.; Miao, Z.G.; Jiang, A.L.; Shi, L.; Ren, A.; Zhao, M.W. UDP-glucose pyrophosphorylase influences polysaccharide synthesis, cell wall components, and hyphal branching in Ganoderma lucidum via regulation of the balance between glucose-1-phosphate and UDP-glucose. Fungal Genet. Biol. 2015, 82, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.L.; Xu, J.; Liu, C.; Zhu, Y.J.; Nelson, D.R.; Zhou, S.G.; Li, C.F.; Wang, L.Z.; Guo, X.; Sun, Y.Z. Genome sequence of the model medicinal mushroom Ganoderma lucidum. Nat. Commun. 2011, 3, 913. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.Y.; Fan, W.L.; Wang, W.F.; Chen, T.; Tang, Y.C.; Chu, F.H.; Chang, T.T.; Wang, S.Y.; Li, M.Y.; Chen, Y.H. Genomic and transcriptomic analyses of the medicinal fungus Antrodia cinnamomea for its metabolite biosynthesis and sexual development. Proc. Natl. Acad. Sci. USA 2014, 111, 4743–4752. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.F.; Gong, Y.H.; Cai, Y.L.; Liu, W.; Zhou, Y.; Xiao, Y.; Xu, Z.Y.; Liu, Y.; Lei, X.Y.; Wang, G.Z. Genome sequence of the edible cultivated mushroom Lentinula edodes (shiitake) reveals insights into lignocellulose degradation. PLoS ONE 2016, 11, e0160336. [Google Scholar] [CrossRef] [PubMed]
- Yap, H.Y.Y.; Chooi, Y.H.; Firdausraih, M.; Fung, S.Y.; Ng, S.T.; Tan, C.S.; Tan, N.H. The genome of the tiger milk mushroom, Lignosus rhinocerotis, provides insights into the genetic basis of its medicinal properties. BMC Genom. 2014, 15, 635. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.L.; Min, W.H.; Bi, P.Y.; Zhou, H.B.; Huang, F.R. Stimulatory effects of coix lacryma-jobi oil on the mycelial growth and metabolites biosynthesis by the submerged culture of Ganoderma lucidum. Biochem. Eng. J. 2013, 76, 77–82. [Google Scholar] [CrossRef]
- Tang, Y.J.; Zhong, J.J. Exopolysaccharide biosynthesis and related enzyme activities of the medicinal fungus, Ganoderma lucidum, grown on lactose in a bioreactor. Biotechnol. Lett. 2002, 24, 1023–1026. [Google Scholar] [CrossRef]
- Chen, R.Z.; Zhao, X.; Shao, Z.; Wei, Z.; Wang, Y.Y.; Zhu, L.L.; Zhao, J.; Sun, M.X.; He, R.F.; He, G.C. Rice UDP-glucose pyrophosphorylase1 is essential for pollen callose deposition and its cosuppression results in a new type of thermosensitive genic male sterility. Plant Cell 2007, 19, 847–861. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.T.; Du, S.C.; Lo, H.H.; Hsiao, Y.M. The galu gene of Xanthomonas campestris pv. Campestris is involved in bacterial attachment, cell motility, polysaccharide synthesis, virulence, and tolerance to various stresses. Arch. Microbiol. 2014, 196, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.S.; Lin, T.Y.; Wang, W.B.; Liu, M.C.; Poren, H.; Shwujen, L. Characterization of udp-glucose dehydrogenase and UDP-glucose pyrophosphorylase mutants of proteus mirabilis: Defectiveness in polymyxin b resistance, swarming, and virulence. Antimicrob. Agents Chemother. 2010, 54, 2000–2009. [Google Scholar] [CrossRef] [PubMed]
- Diamantopoulou, P.; Papanikolaou, S.; Komaitis, M.; Aggelis, G.; Philippoussis, A. Patterns of major metabolites biosynthesis by different mushroom fungi grown on glucose-based submerged cultures. Bioprocess Biosyst. Eng. 2014, 37, 1385–1400. [Google Scholar] [CrossRef] [PubMed]
- Su, C.H.; Lai, M.N.; Lin, C.C.; Ng, L.T. Comparative characterization of physicochemical properties and bioactivities of polysaccharides from selected medicinal mushrooms. Appl. Microbiol. Biotechnol. 2016, 100, 4385–4393. [Google Scholar] [CrossRef] [PubMed]
- Chien, R.C.; Yen, M.T.; Tseng, Y.H.; Mau, J.L. Chemical characteristics and anti-proliferation activities of ganoderma tsugae polysaccharides. Carbohydr. Polym. 2015, 128, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.S.; Chen, H.X.; Zhu, W.C.; Wang, Z.S. Effect of different drying methods on physicochemical properties and antioxidant activities of polysaccharides extracted from mushroom inonotus obliquus. Food Res. Int. 2013, 50, 633–640. [Google Scholar] [CrossRef]
- Xu, X.Q; Li, J.; Hu, Y. Polysaccharides from inonotus obliquus sclerotia and cultured mycelia stimulate cytokine production of human peripheral blood mononuclear cells in vitro and their chemical characterization. Int. Immunopharmacol. 2014, 21, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.J.; Zhu, L.W.; Li, H.M.; Li, D.S. Submerged culture of mushrooms in bioreactors- challenges, current state-of-the-art, and future prospects. Food Technol. Biotechnol. 2006, 45, 221–229. [Google Scholar]
- Zhao, W.; Chai, D.D.; Li, H.M.; Chen, T.; Tang, Y.J. Significance of metal ion supplementation in the fermentation medium on the structure and anti-tumor activity of tuber polysaccharides produced by submerged culture of Tuber melanosporum. Process Biochem. 2014, 49, 2030–2038. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, L.N.; Chen, L.; Chen, Y.; Cheung, P.C.; Chen, L.G. Effect of culture media on the chemical and physical characteristics of polysaccharides isolated from Poria cocos mycelia. Carbohydr. Res. 2003, 338, 1507–1515. [Google Scholar] [CrossRef]
- Smiderle, F.R.; Olsen, L.M.; Ruthes, A.C.; Czelusniak, P.A.; Santana-Filho, A.P.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, M. Exopolysaccharides, proteins and lipids in Pleurotus pulmonarius submerged culture using different carbon sources. Carbohydr. Polym. 2012, 87, 368–376. [Google Scholar] [CrossRef]
- Chen, X.; Wu, J.Y.; Gui, X.T. Production and characterization of exopolysaccharides in mycelial culture of Cordyceps sinensis fungus CS-HK1 with different carbon sources. Chin. J. Chem. Eng. 2016, 24, 158–162. [Google Scholar] [CrossRef]
Figure 1.
Proposed/deduced/hypothesized biosynthetic pathways for mushroom PSs. 1. Glucokinase (GK); 2. Phosphoglucose isomerase (PGI); 3. Fructose-1,6-biphosphatase (FBPase); 4. Phosphofructokinase (FPK); 5. Phosphomannose isomerase (PMI); 6. Phosphomannose mutase (PMM); 7. GDP-Man pyrophosphorylase (GMP); 8. GDP-Man dehydratase (GMD); 9. GDP-4-keto-6-deoxymannose epimerase/reductase (GMER); 10. Phosphoglucose mutase (PGM); 11. dTDP-glucose pyrophosphorlase (dTDP-GPP); 12 and 13. dTDP-Rha synthase (dTRS); 14. UDP-Glc pyrophosphorylase (UGP); 15. UDP-Gal-4-epimerase (UGE); 16. UDP-Glc dehydrogenase (UGDG). 17. UDP-glucuronate decarboxylase/UDP-Xyl synthase (UXS); 18. UDP-Xyl-4-epimerase (UXE).
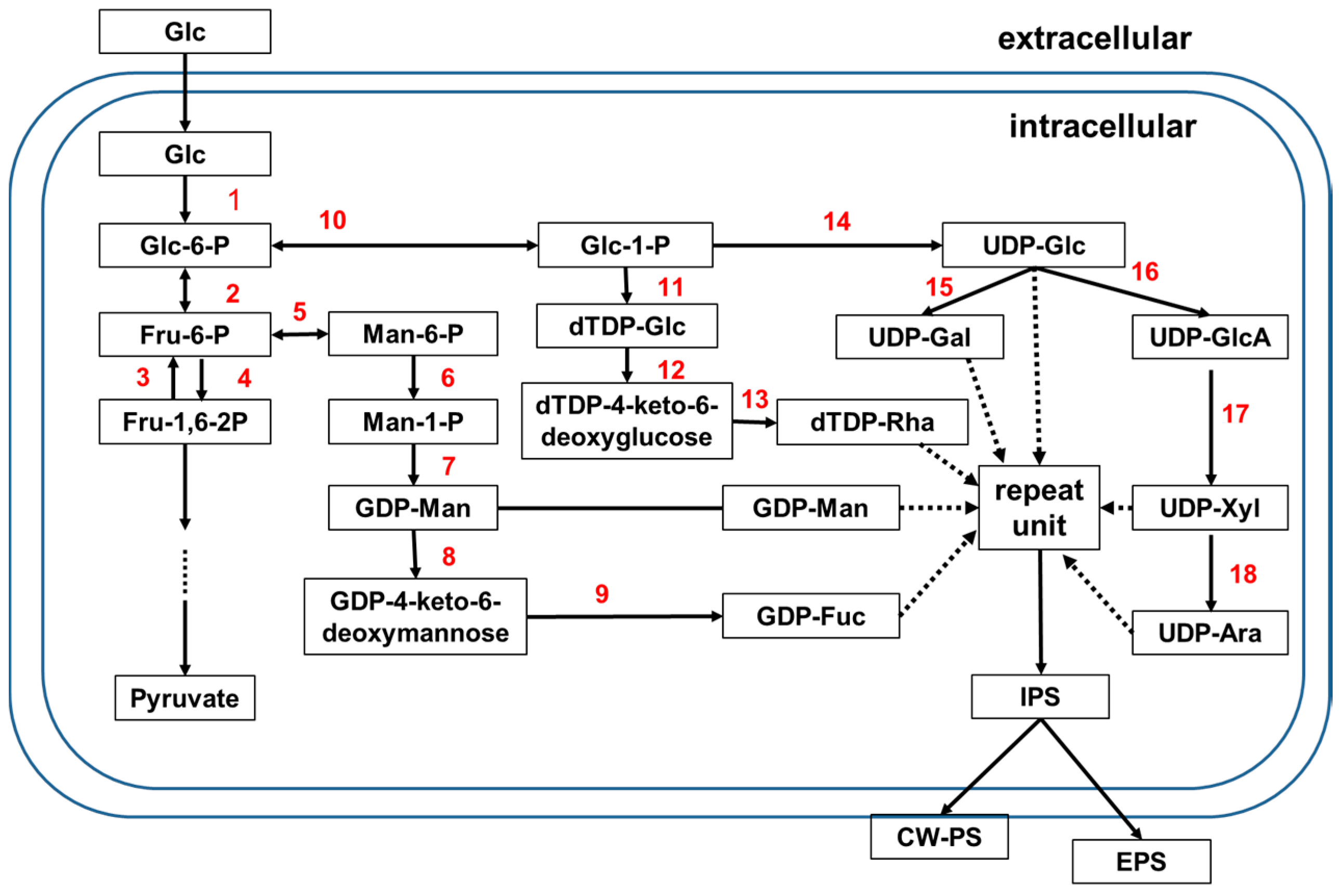
Figure 1.
Proposed/deduced/hypothesized biosynthetic pathways for mushroom PSs. 1. Glucokinase (GK); 2. Phosphoglucose isomerase (PGI); 3. Fructose-1,6-biphosphatase (FBPase); 4. Phosphofructokinase (FPK); 5. Phosphomannose isomerase (PMI); 6. Phosphomannose mutase (PMM); 7. GDP-Man pyrophosphorylase (GMP); 8. GDP-Man dehydratase (GMD); 9. GDP-4-keto-6-deoxymannose epimerase/reductase (GMER); 10. Phosphoglucose mutase (PGM); 11. dTDP-glucose pyrophosphorlase (dTDP-GPP); 12 and 13. dTDP-Rha synthase (dTRS); 14. UDP-Glc pyrophosphorylase (UGP); 15. UDP-Gal-4-epimerase (UGE); 16. UDP-Glc dehydrogenase (UGDG). 17. UDP-glucuronate decarboxylase/UDP-Xyl synthase (UXS); 18. UDP-Xyl-4-epimerase (UXE).
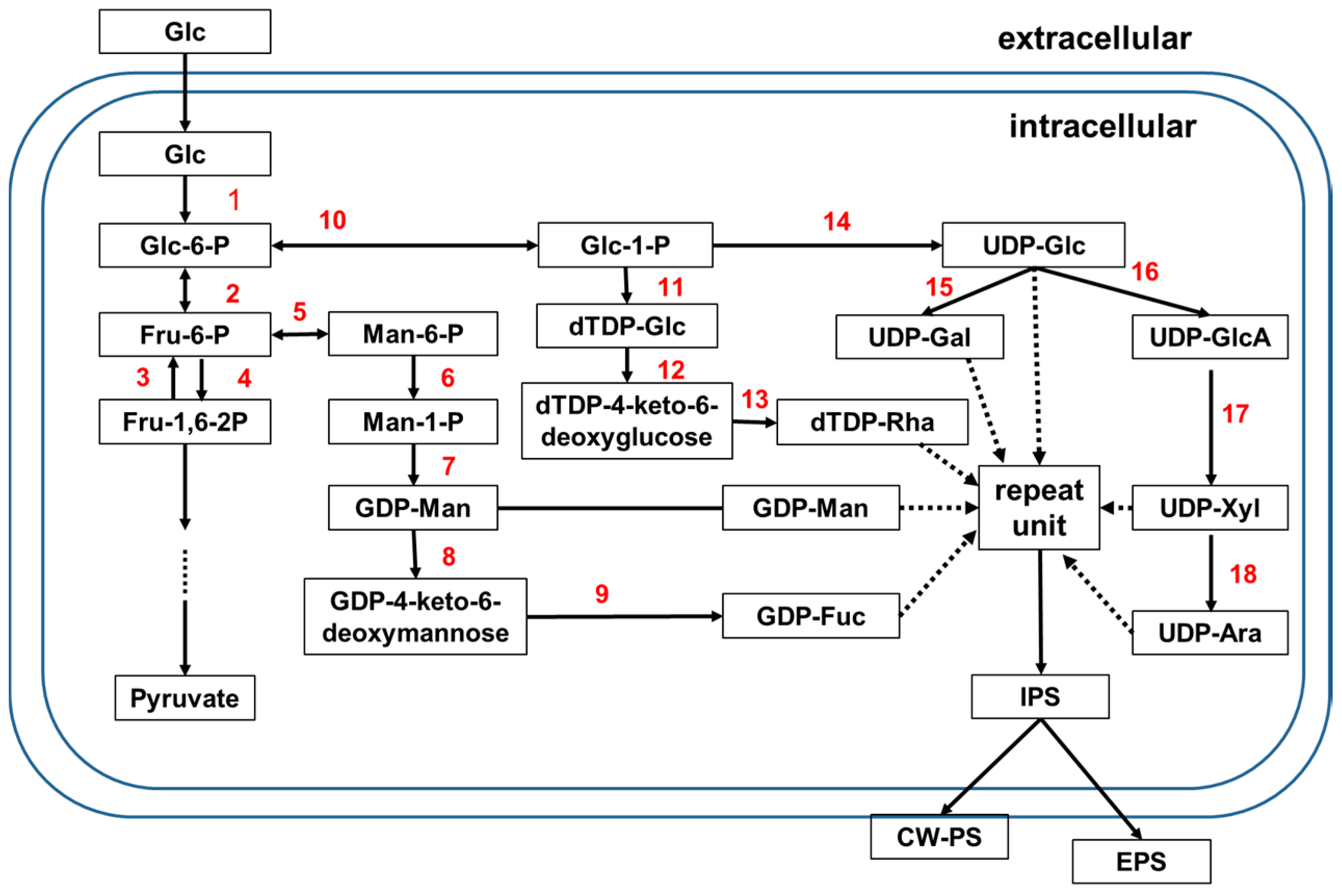
Figure 2.
Correlations (R2) between activities of exopolysaccharide (EPS) synthesis enzymes and monosaccharide mole percentages under various carbon source conditions. (A) Phosphoglucose isomerase (PGI) and Man; (B) Phosphomannose isomerase (PMI) and Man; (C) GDP-Man pyrophosphorylase (GMP) and Man; (D) Phosphoglucose mutase (PGM) and Gal.
Figure 2.
Correlations (R2) between activities of exopolysaccharide (EPS) synthesis enzymes and monosaccharide mole percentages under various carbon source conditions. (A) Phosphoglucose isomerase (PGI) and Man; (B) Phosphomannose isomerase (PMI) and Man; (C) GDP-Man pyrophosphorylase (GMP) and Man; (D) Phosphoglucose mutase (PGM) and Gal.

{kind=link}
{kind=link}
Table 1.
Monosaccharide composition and molar ratio of polysaccharides from Ascomycetes.
| Species | Source | Monosaccharide Composition | Molar Ratio | Reference |
|---|---|---|---|---|
| Cordyceps sinensis | mycelium | Man:Glc:Gal:GlcA | 9.6:4.0:4.4:1.0 | [19] |
| Cordyceps militaris | fruiting body | Man:Gal:Glc | 56.7:34.5:8.8 | [22] |
| NA | Man:Glc:Gal | 2.84:1:1.29 | [23] | |
| Man:Glc:Gal | 2.05:1:1.09 | |||
| fruiting body | Man:Glc:Gal | 1:28.63:1.41 | [24] | |
| Man:Glc:Gal | 1:12.41:0.74 | |||
| Cordyceps sobolifera | fruiting body | Man:Glc:Gal | 1.7:8.9:1 | [25] |
| Cordyceps cicadae | NA | Glc:Rha:Xyl:Man:Ara:Gal | 63.10:39.11:20.12:15.16:2.05:0.12 | [26] |
| Cordyceps kyushuensis | stroma | Fru:Man:Rha:GalN:Ara | 1.0:1.19:0.11:0.11:0.34 | [27] |
| Fru:Man:Rha:Glc:Ara | 1:1.29:0.14:0.07:0.32 | |||
| Morchella esculenta | fruiting body | Xyl:Glc:Man:Rha:Gal | 5.4:5.0:6.5:7.8:72.3 | [28] |
| fruiting body | Glc:Man:Gal:Ara:Xyl:Rha | 38.06:14.43:17.06:9.25:2.08:0.94 | [29] | |
| Glc:Man:Gal:Ara:Xyl:Rha | 27.04:28.66:11.12:9.07:6.71:3.22 | |||
| Glc:Man:Gal:Ara:Xyl:Rha | 24.69:20.46:10.22:7.91:4.05:2.83 | |||
| fermentation broth | Ara:Man:Glc:Gal | 0.7:2.8:24.8:1.0 | [30] | |
| Rha:Man:Glc:Gal | 1.8:3.1:21.4:1.0 | |||
| Tuber huidongense | fruiting body | Glc:Man:Gal | 60.56:20.12:19.32 | [32] |
| Glc | - | |||
| Tuber rufum | fruiting body | Glc:Gal:Fuc | 4:3:1 | [33] |
Glc: glucose; Man: mannose; Gal: galactose, Xyl: xylose; Ara: arabinose, Rha: rhamnose, Fuc: fucose; GlcA: glucuronic acid; GalN: N-acetyl-galactosamine.
Table 2.
Monosaccharide composition and molar ratio of polysaccharides from Ganoderma species.
| Species | Source | Monosaccharide Composition | Molar Ratio | Reference |
|---|---|---|---|---|
| Ganoderma lucidum | fruiting body | Rha:Ara:Xyl:Man:Glc:Gal | 11.4:30:27.1:8:9.1:14.3 | [34] |
| Rha:Ara:Xyl:Glc:Gal | 44.7:20.9:19.9:3.6:10.8 | |||
| NA | Man:Rha:Glc:Gal | 4.7:0.65:65.22:29.43 | [35] | |
| fruiting body | Glc | - | [36] | |
| Glc:Gal:Man | 29:1.8:1.0 | |||
| fruiting body | Glc | - | [37] | |
| fermentation broth | Gal:Man:Glc:Ara:Rha | 103:17:12:10:3 | [38] | |
| fruiting body | Glc:Fuc:Gal | 1.0:1.09:4.09 | [39] | |
| fruiting body | Gal:Rha:Glc | 1.00:1.15:3.22 | [40] | |
| fermented soybean curd residue | Ara:Rha:Xyl:Man:Glc | 4.66:1.23:3.1:0.61:1.29 | [41] | |
| Ara:Xyl:Glc | 2.82:1.33:0.87 | |||
| Ara:Rha:Xyl:Gal:Man:Glc | 5.09:0.52:1.07:1.29:0.48:2.76 | |||
| Ganoderma atrum | fruiting body | Gal:Fuc:Glc | 75.87:11.83:6.02 | [42] |
| fruiting body | Glc:Man:Gal:GalA | 4.91:1:1.28:0.71 | [43] | |
| Ganoderma sinense | fruiting body | Man:Glc:Gal | 4.7:27.1:1.0 | [44] |
| Ganoderma tsugae | mycelium | Fuc:Xyl:Man:Gal:Glc:GlcNac | 2.9:16.1:66.7:9.4:0.1:4.3 | [45] |
| Fuc:Xyl:Man:Gal:Glc:GlcNac | 4.5:1.3:58.0:24.5:3.8:7.9 | |||
| Ganoderma formosanum | fruiting body | Man:Gal:Glc:Ara:Fuc:Fru:Rha | 50.13:13.1:17.47:6.94:2.71:9.21:0.45 | [46] |
| Man:Gal:Glc:Ara:Fuc:Rha | 44.91:38.64:8.26:0.08:8.02:0.09 | |||
| Man:Gal:Glc:Ara:Fuc:Fru:Rha | 33.35:30.84:20.52:8.78:4.44:1.33:0.74 |
Glc: glucose; Man: mannose; Gal: galactose, Xyl: xylose; Ara: arabinose, Rha: rhamnose, Fuc: fucose; GalA: galacturonic acid; GlcNac: N-acetyl-glucosamine.
Table 3.
Biological activities of active components in mushroom PSs.
| Species | Biological Activity | Active Component | Reference |
|---|---|---|---|
| Inonotus obliquus | antioxidant and anticancer activity | Man | [53,54,55] |
| Tremella mesenterica | cytokine-stimulating activity | Man | [47] |
| Ganoderma lucidum | antioxidant activity | Rha | [34] |
| Flammulina velutipes | antioxidant activity | Rha | [57] |
| Agrocybe cylindracea | antioxidant activity | Glc, Gal | [58] |
| Sarcodon aspratus | macrophage activation | Man, Rha | [59] |
| Inonotus obliquus | antioxidant activity | Man, Rha, Xyl | [8] |
| Hirsutella sp. | antioxidant activity | Man, Glc | [60] |
| Pleurotus eryngii | antioxidant activity | Man, Rha, GalA | [61] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wang, Q.; Wang, F.; Xu, Z.; Ding, Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules 2017, 22, 955. https://doi.org/10.3390/molecules22060955
AMA Style
Wang Q, Wang F, Xu Z, Ding Z. Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation. Molecules. 2017; 22(6):955. https://doi.org/10.3390/molecules22060955
Chicago/Turabian StyleWang, Qiong, Feng Wang, Zhenghong Xu, and Zhongyang Ding. 2017. "Bioactive Mushroom Polysaccharides: A Review on Monosaccharide Composition, Biosynthesis and Regulation" Molecules 22, no. 6: 955. https://doi.org/10.3390/molecules22060955