Cellulase-Assisted Extraction of Polysaccharides from White Hyacinth Bean: Characterization of Antioxidant Activity and Promotion for Probiotics Proliferation


Abstract
:1. Introduction
2. Results and Discussion
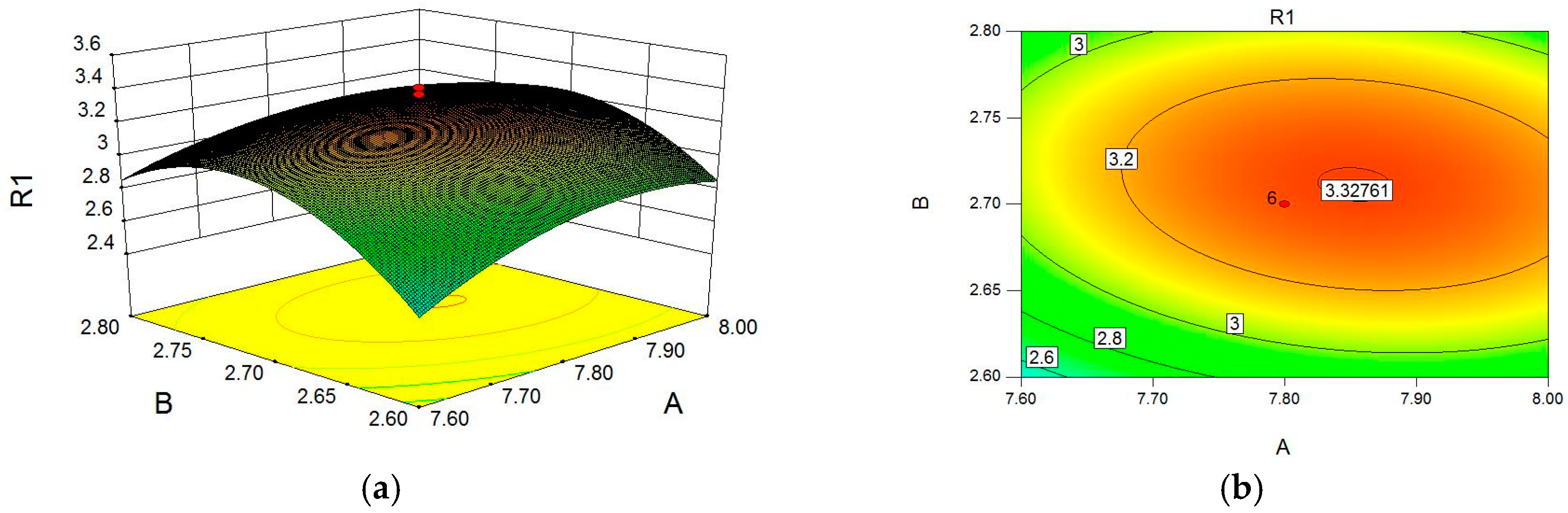
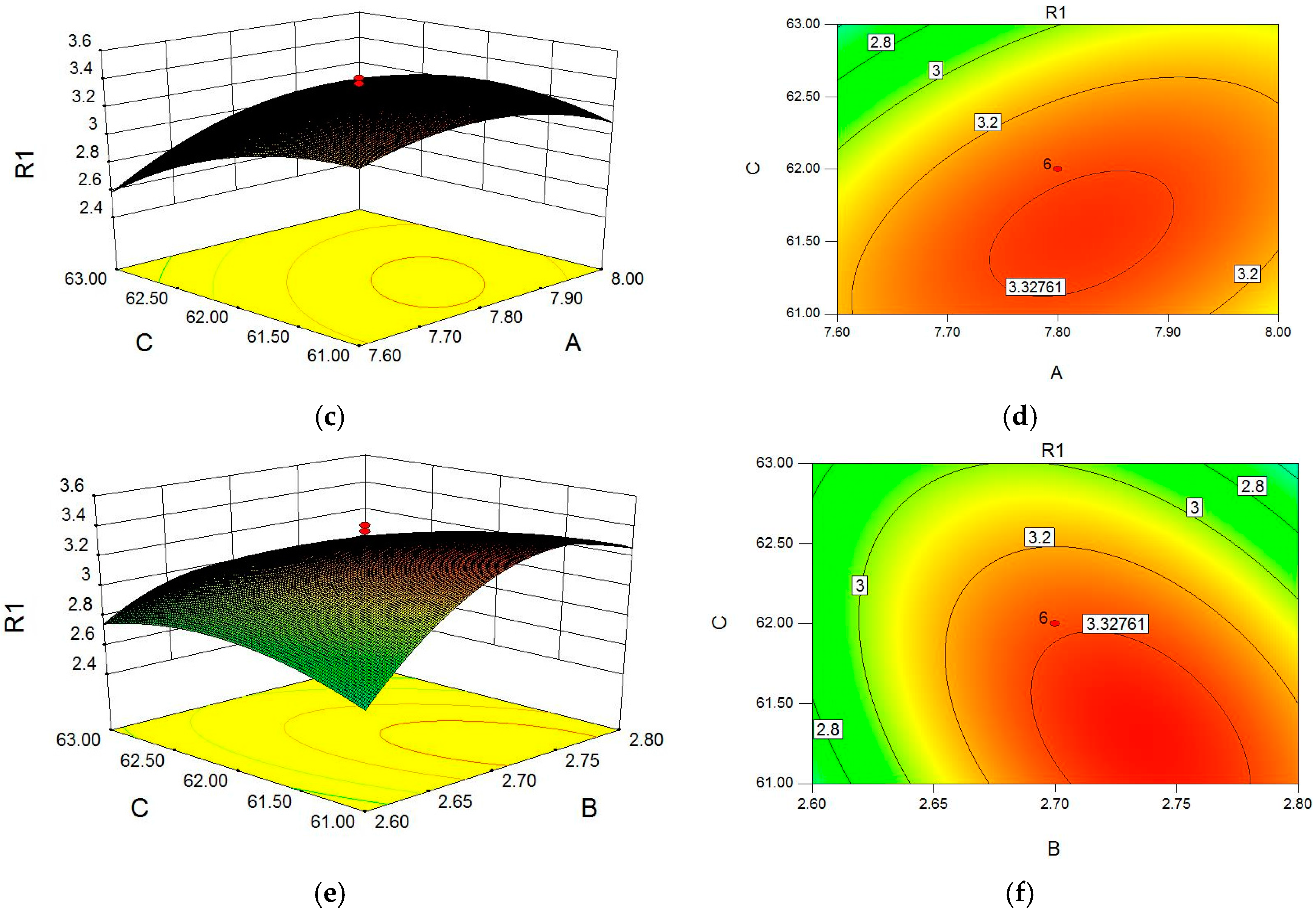
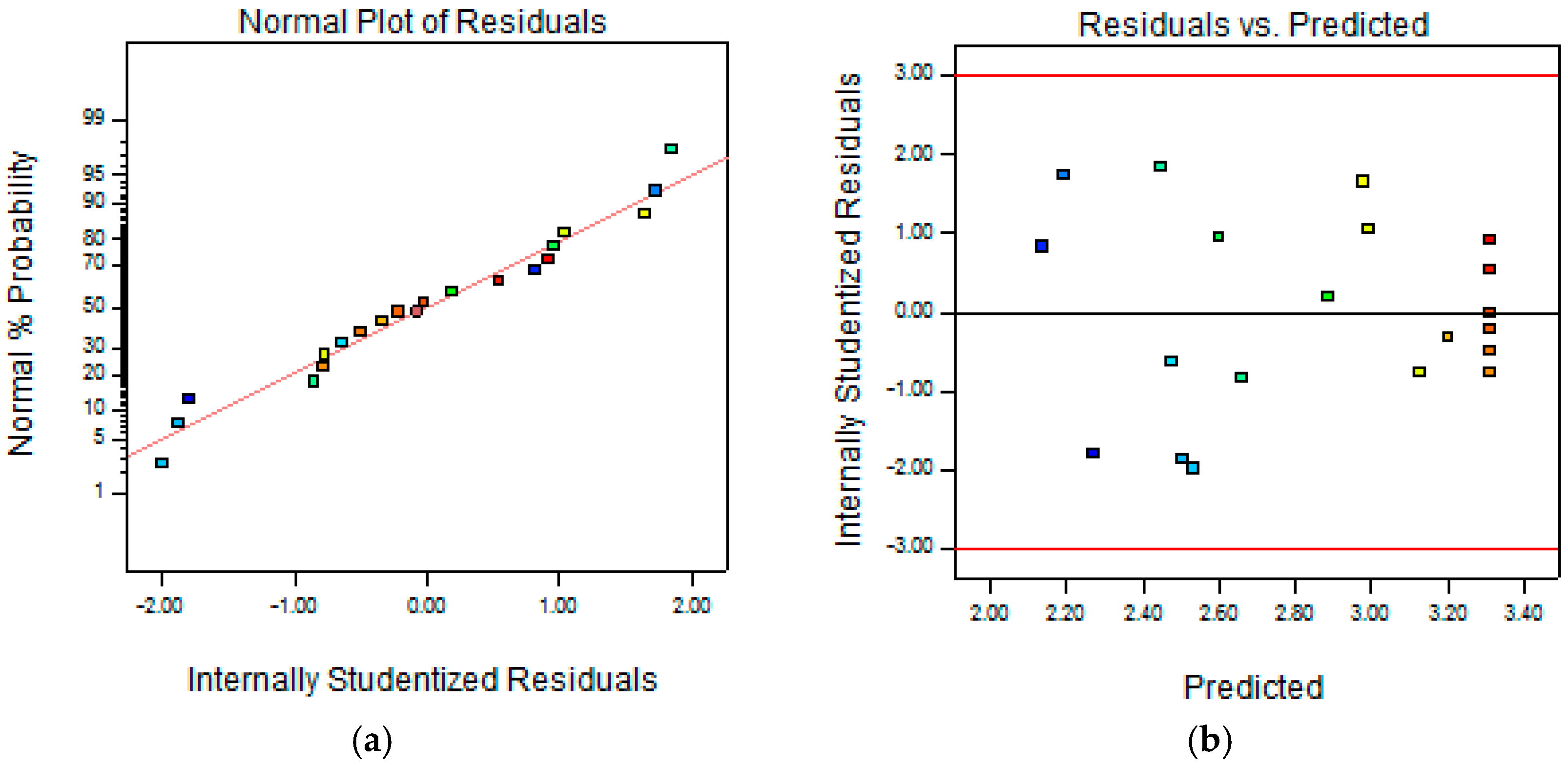
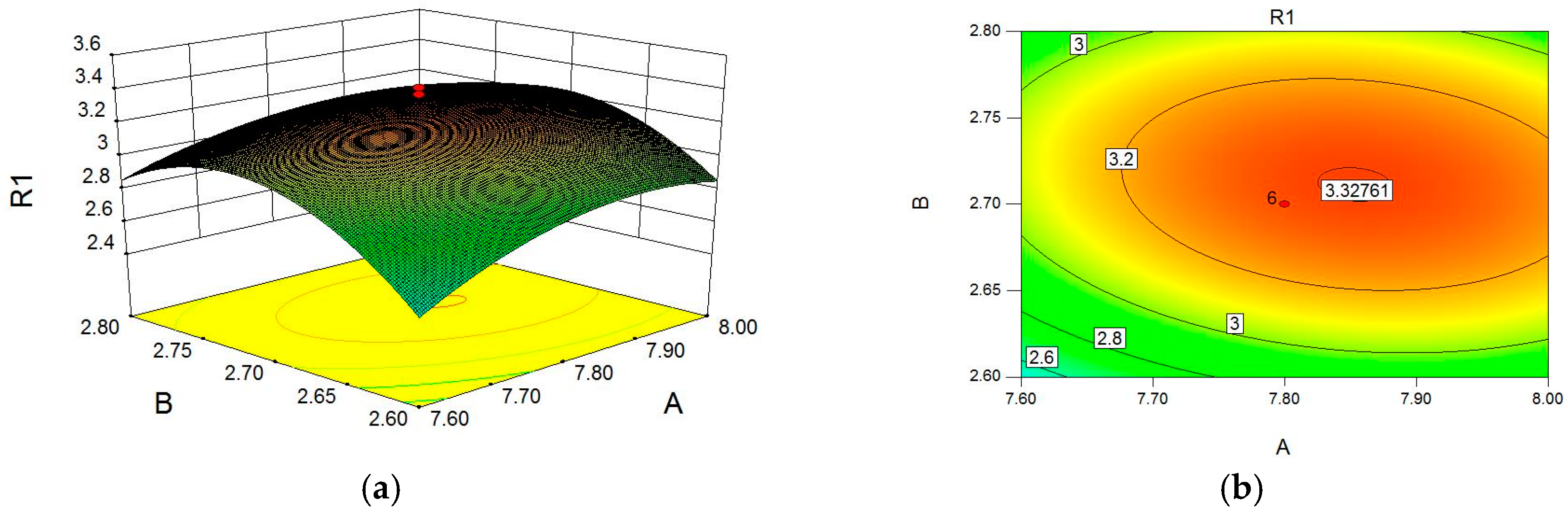
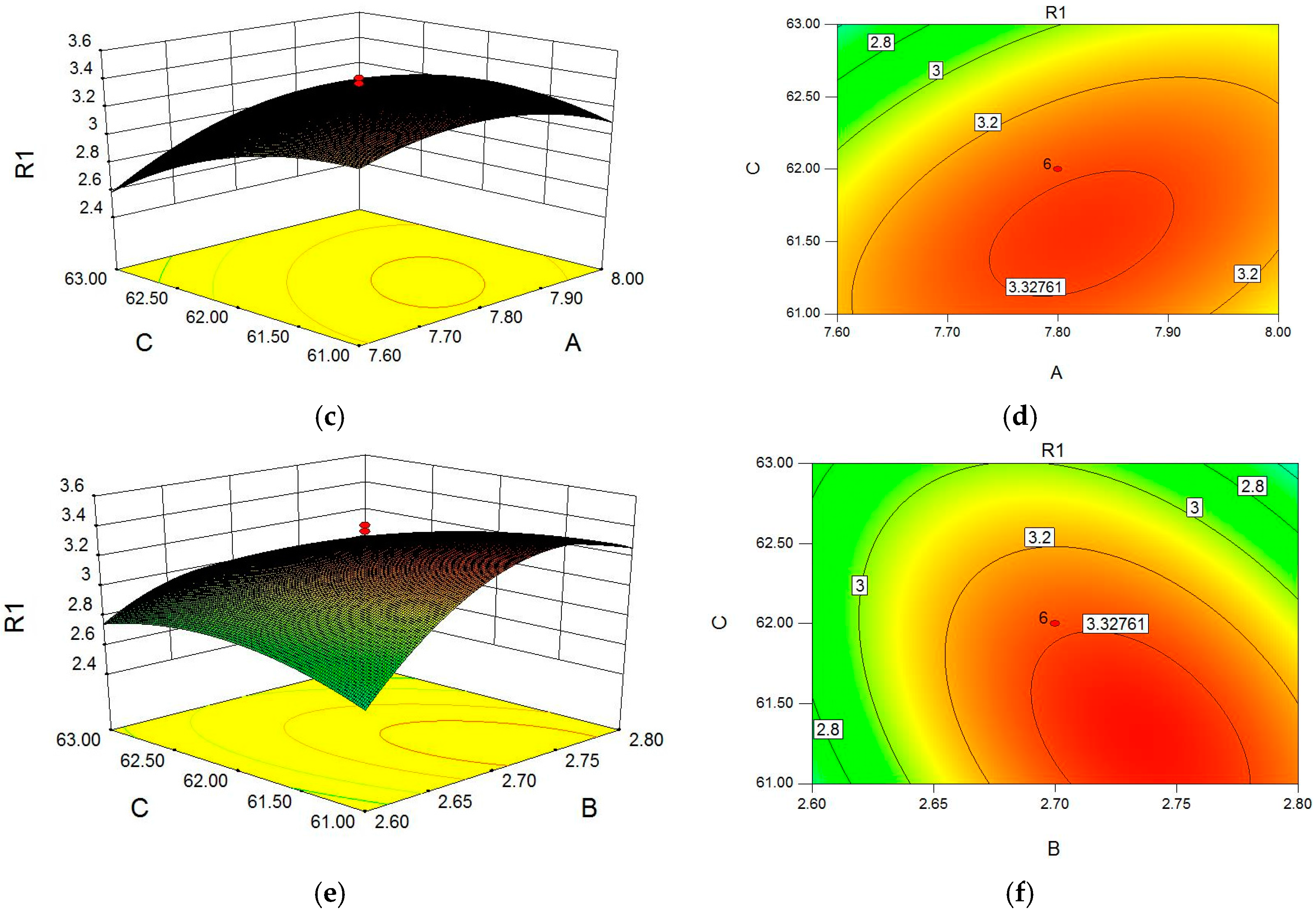
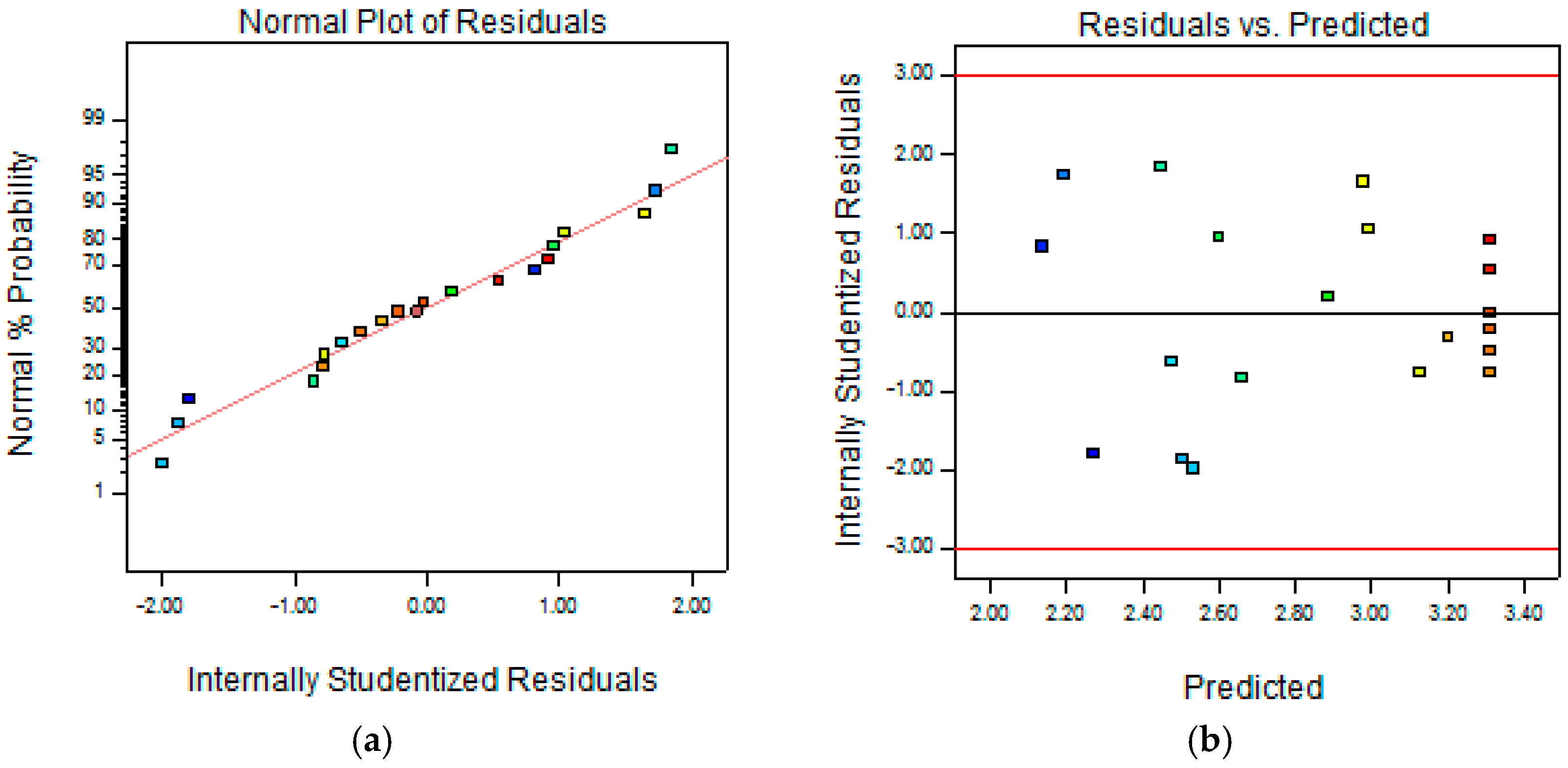
2.1. Optimization of Cellulase-Assisted Extraction of PWBs by Response Surface Methodology
2.2. Antioxidant Activities of PWBs
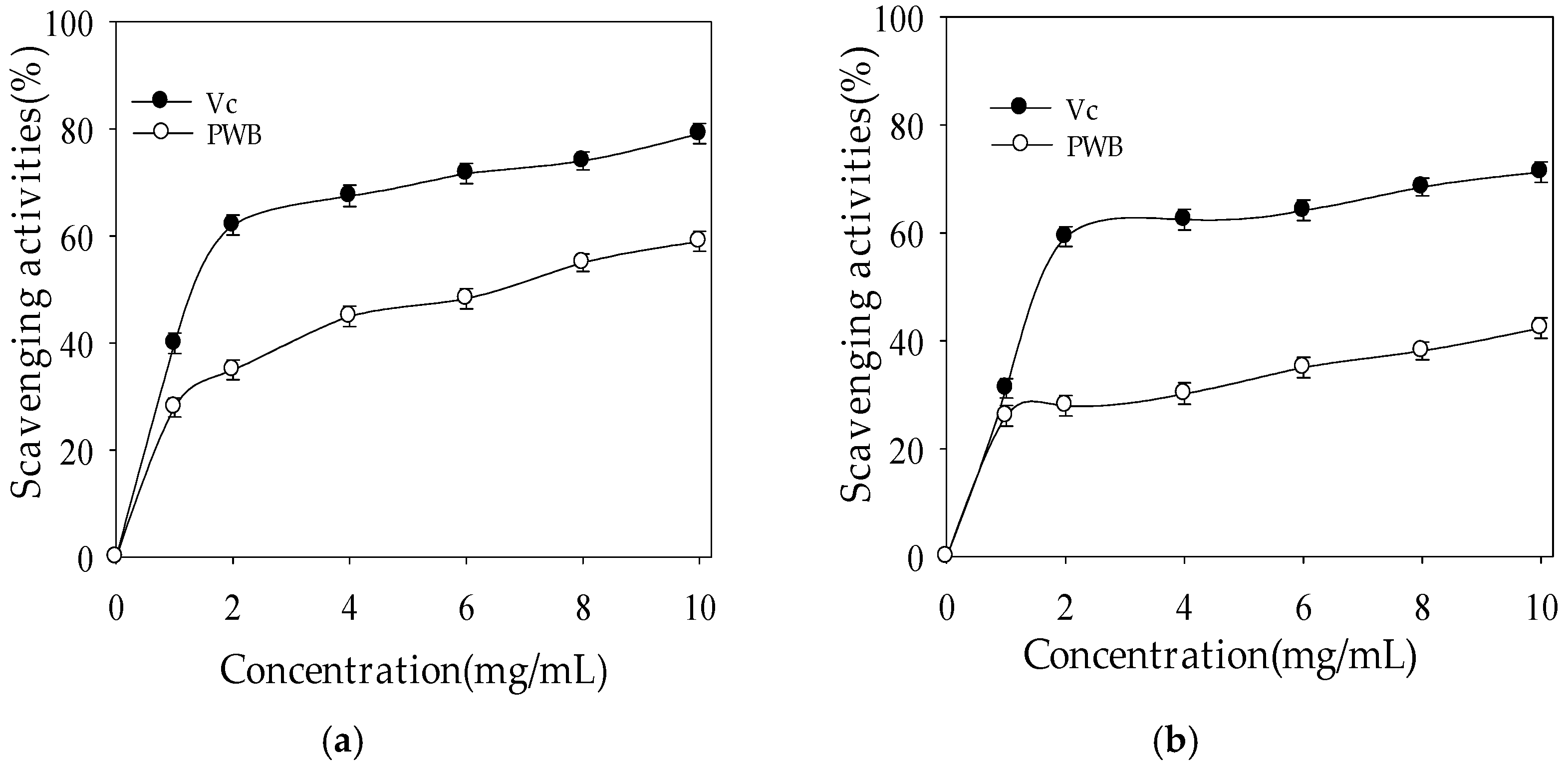
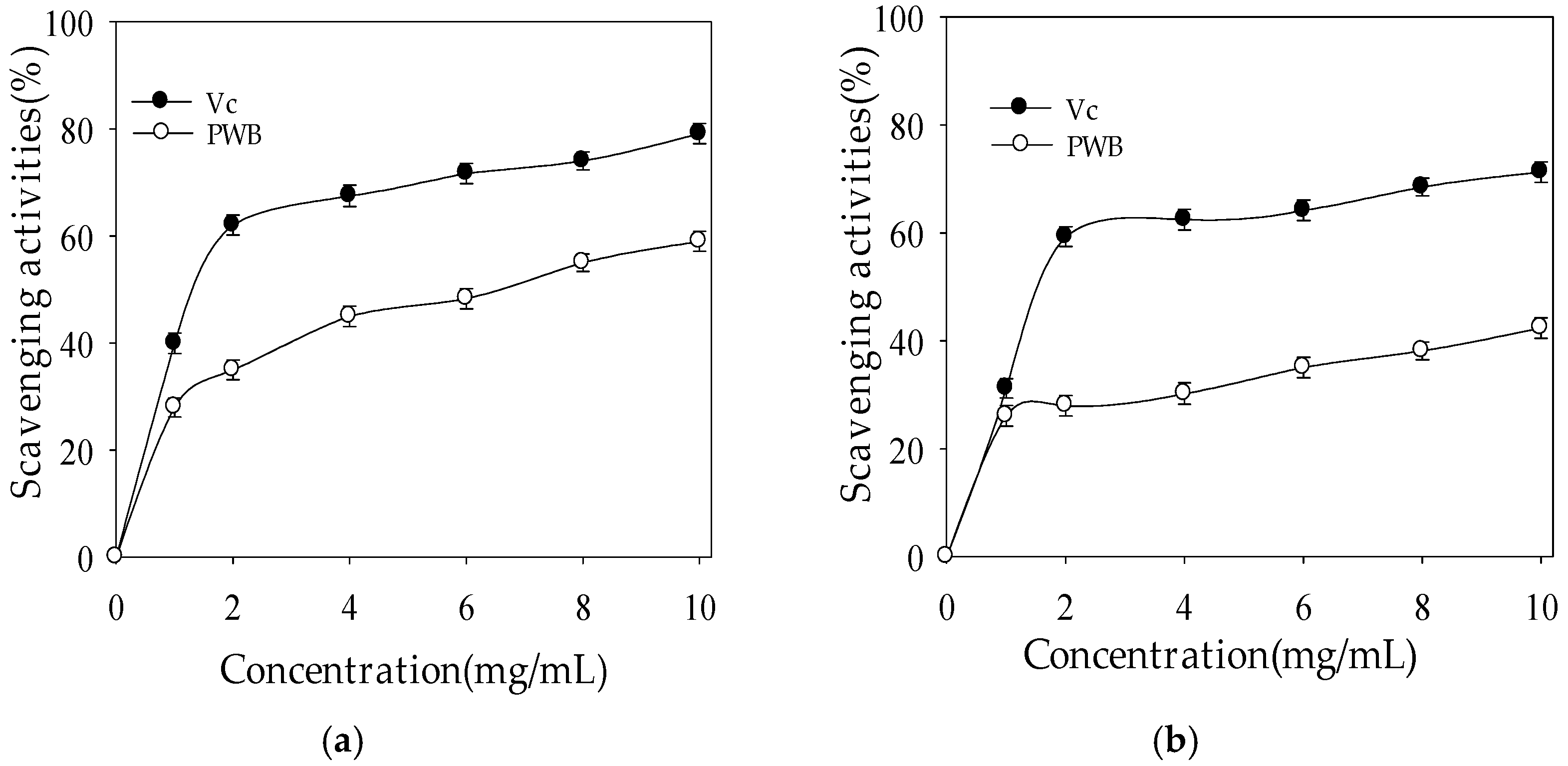
2.2.1. Hydroxyl Radical-Scavenging Activity
2.2.2. Superoxide Radical-Scavenging Activity
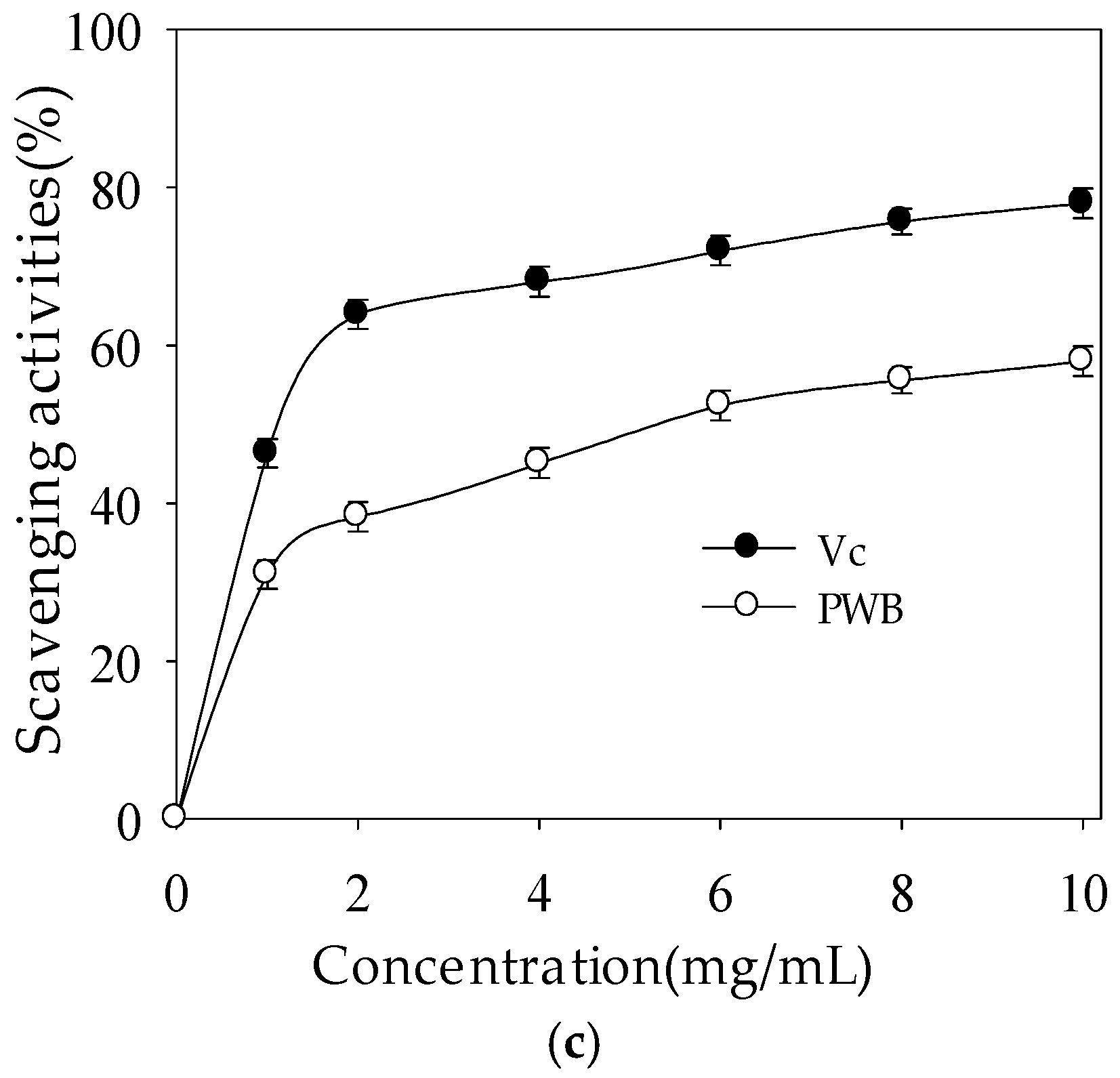
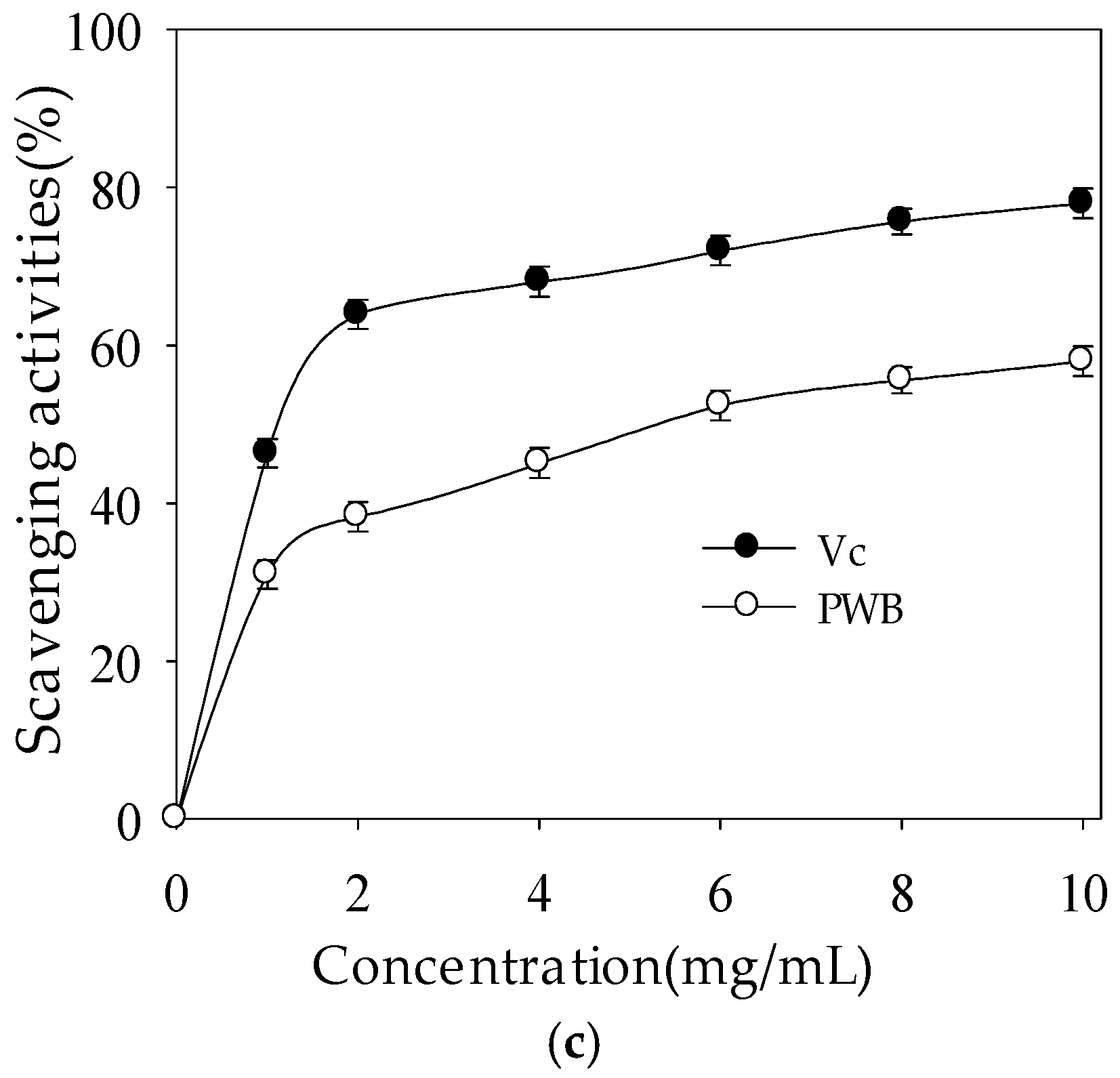
2.2.3. DPPH Radical-Scavenging Activity
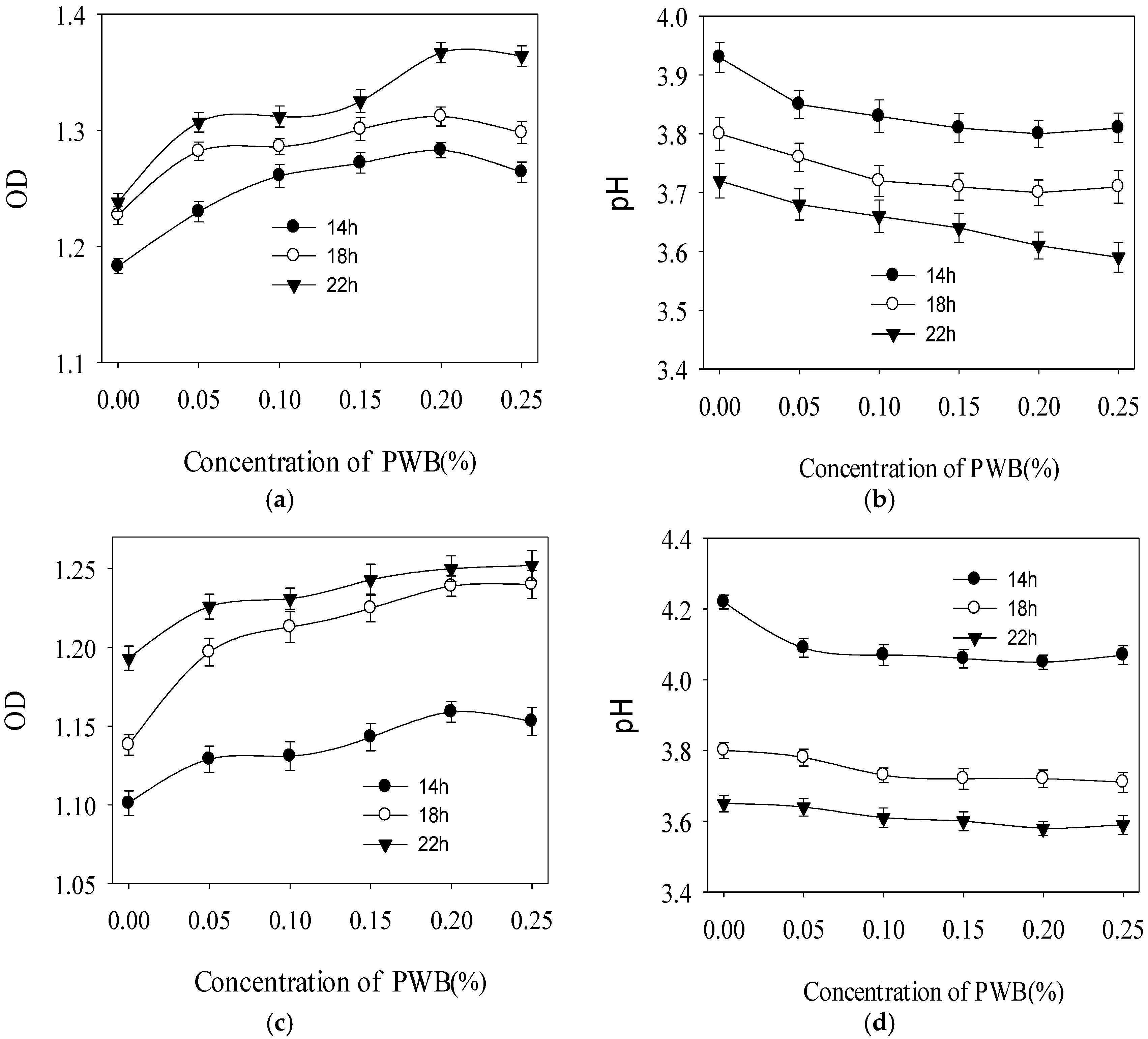
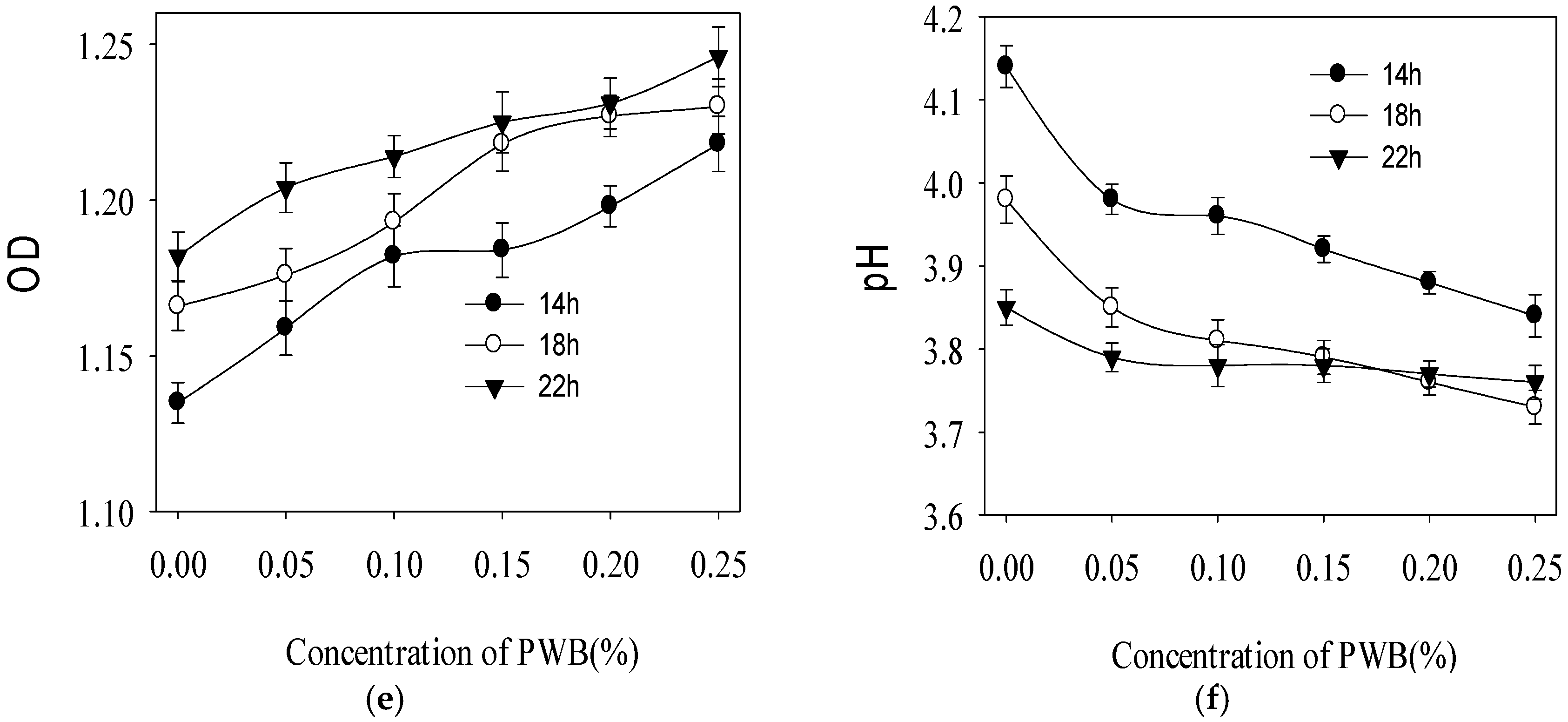
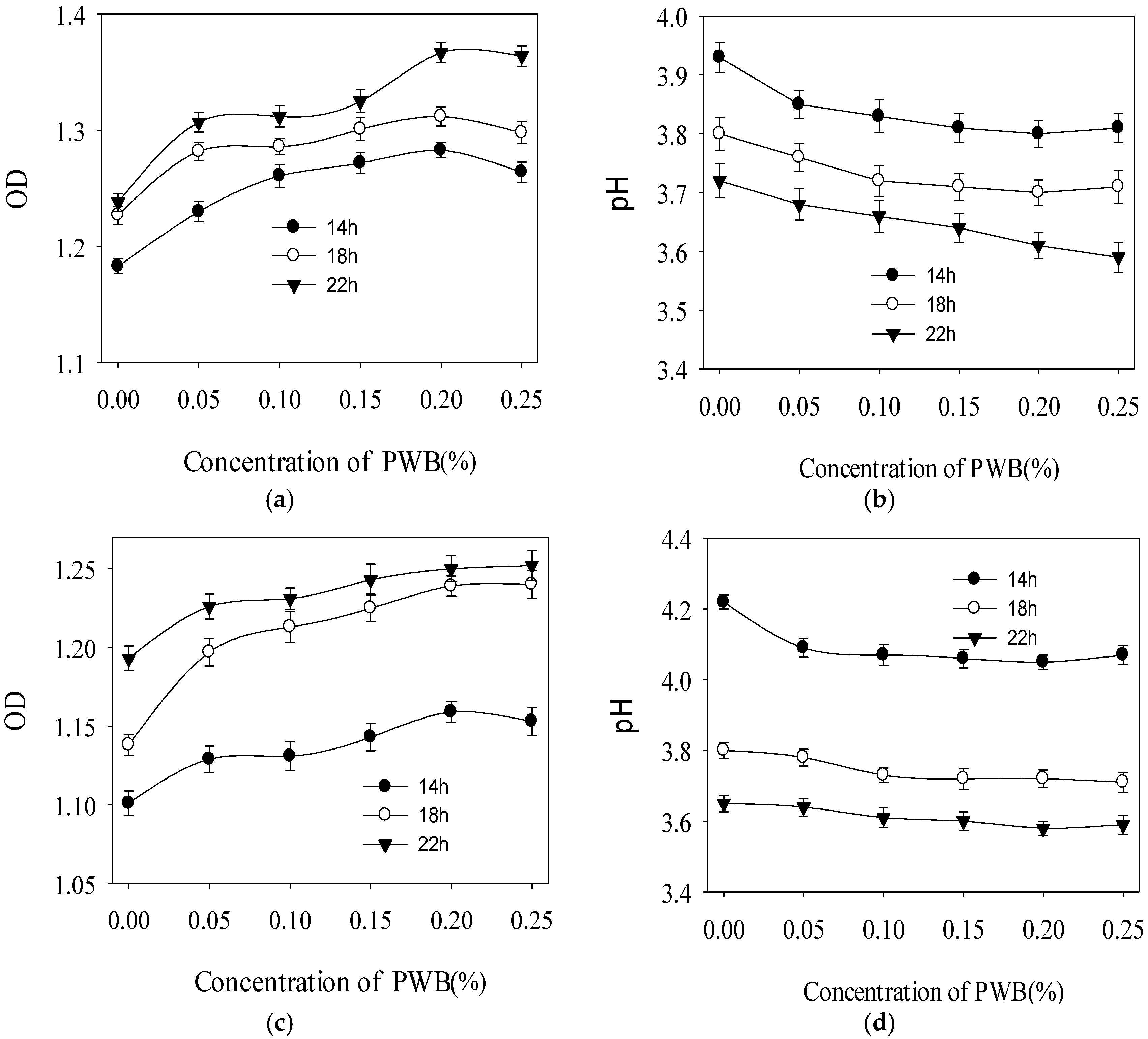
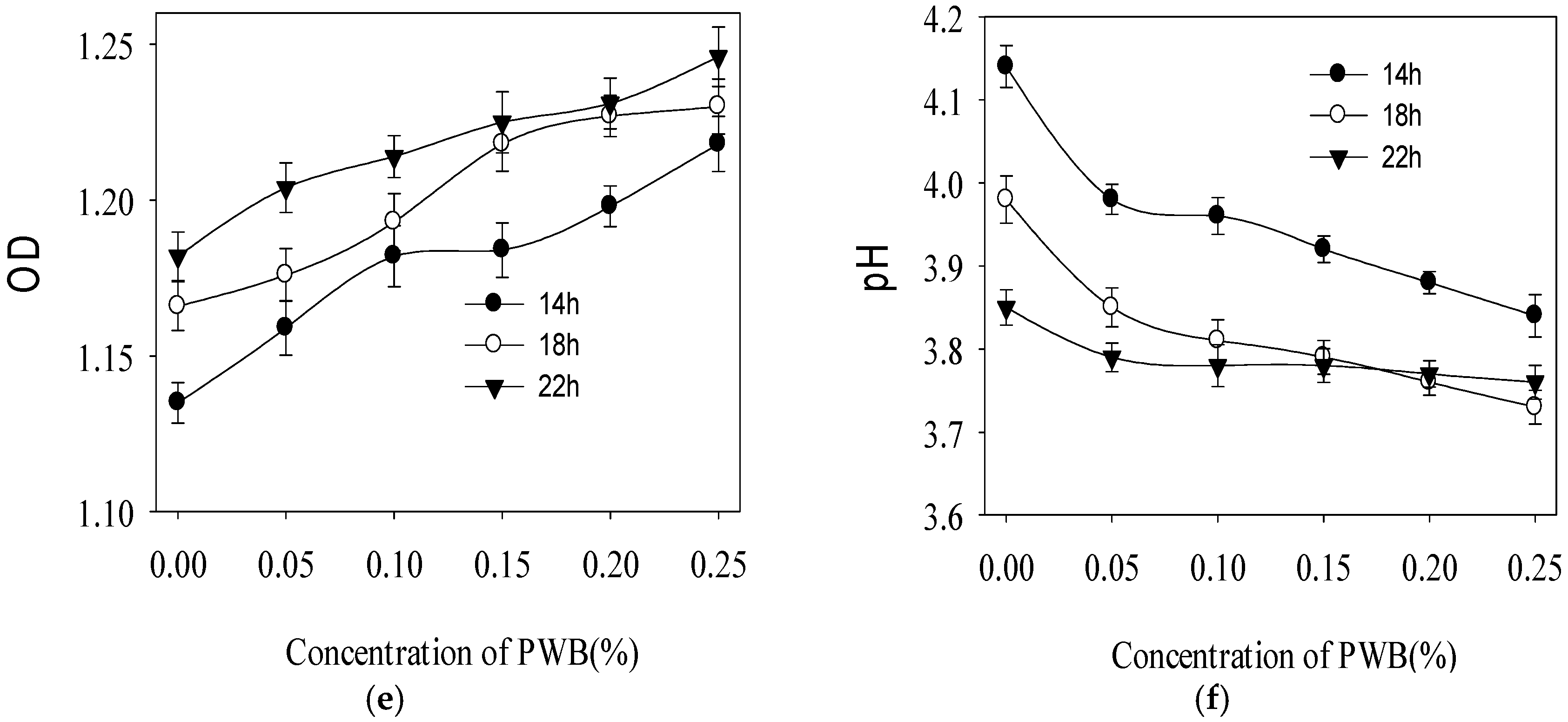
2.3. Effect of Polysaccharide on the Growth of Three Selected Probiotics
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Enzyme-Assisted Extraction of PWBs
3.3. Central Composite Design (CCD) and Statistical Analysis
3.4. Antioxidant Activity of Polysaccharides
3.4.1. An Assay of Scavenging Hydroxyl Radical Activity
3.4.2. Assay of Superoxide Radical Scavenging Activity
3.4.3. Assay of DPPH Radical Scavenging Activity
3.5. Effect of PWBs on the Growth of Three Selected Probiotics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Choudhury, F.K.; Rivero, R.M.; Blumwald, E.; Mittler, R. Reactive oxygen species, abiotic stress and stress combination. Plant J. 2017, 90, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Cadenas, E.; Davies, K.J. Mitochondrial free radical generation, oxidative stress, and aging. Free Radic. Biol. Med. 2000, 29, 222–230. [Google Scholar] [CrossRef]
- Ksouri, R.; Ksouri, W.M.; Jallali, I.; Debez, A.; Magné, C.; Hiroko, I. Medicinal halophytes: Potent source of health promoting biomolecules with medical, nutraceutical and food applications. Crit. Rev. Biotechnol. 2012, 32, 289–326. [Google Scholar] [CrossRef] [PubMed]
- Frankel, E.N. Antioxidants in lipid foods and their impact on food quality. Food Chem. 1996, 57, 51–55. [Google Scholar] [CrossRef]
- Moein, M.; Moein, S.; Fard, T.B.; Sabahi, Z. Scavenging evaluation of different free radicals by three species of ziziphus, and their fractions. Iran. J. Sci. Technol. A 2017, 41, 249–255. [Google Scholar] [CrossRef]
- Botterweck, A.A.; Verhagen, H.; Goldbohm, R.A.; Kleinjans, J.; Brandt, P.A. Intake of butylated hydroxyanisole and butylated hydroxytoluene and stomach cancer risk: Results from analyses in the netherlands cohort study. Food Chem. Toxicol. 2000, 38, 599–605. [Google Scholar] [CrossRef]
- Petera, B.; Delattre, C.; Pierre, G.; Wadouachi, A.; Elboutachfaiti, R.; Engel, E. Characterization of arabinogalactan-rich mucilage from cereus triangularis cladodes. Carbohydr. Polym 2015, 127, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, C.; Yue, Q. Extraction of polysaccharides from Phellinus nigricans, mycelia and their antioxidant activities in vitro. Carbohydr. Polym. 2014, 99, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Delattre, C.; Pierre, G.; Gardarin, C.; Traikia, M.; Elboutachfaiti, R.; Isogai, A. Antioxidant activities of a polyglucuronic acid sodium salt obtained from tempo-mediated oxidation of xanthan. Carbohydr. Polym. 2015, 116, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Zhang, F.; Luo, Y.; Ma, L.; Kou, X.; Huang, K. Antioxidant activity of a water-soluble polysaccharide purified from Pteridium Aquilinum. Carbohydr. Res. 2008, 344, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Kardosová, A.; Machová, E. Antioxidant activity of medicinal plant polysaccharides. Fitoterapia 2006, 77, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Sun, Y.; Yang, B.; Wang, Z.; Liu, Y.; Cao, Q. Optimization of polysaccharides extraction from seeds of Pharbitis nil and its anti-oxidant activity. Carbohydr. Polym. 2014, 102, 460–466. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Huang, J.; Mao, J.W.; Gong, J.Y.; Zhou, Y.F.; Huang, J.X. Optimization of total polysaccharide extraction from Herba Lophatheri using RSM and antioxidant activities. Int. J. Biol. Macromol. 2014, 67, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Subagio, A. Characterization of hyacinth bean (Lablab purpureus (L.) sweet) seeds from Indonesia and their protein isolate. Food Chem. 2006, 95, 65–70. [Google Scholar] [CrossRef]
- You, Q.; Yin, X.; Zhao, Y. Enzyme assisted extraction of polysaccharides from the fruit of Cornus officinalis. Carbohydr. Polym. 2013, 98, 607–610. [Google Scholar] [CrossRef] [PubMed]
- Singdevsachan, S.K.; Auroshree, P.; Mishra, J.; Baliyarsingh, B.; Tayung, K.; Thatoi, H. Mushroom polysaccharides as potential prebiotics with their antitumor and immunomodulating properties: A review. Bioact. Carbohydr. Diet. Fibre 2016, 7, 1–14. [Google Scholar] [CrossRef]
- Lei, N.; Chen, H.; Feng, J.H. Optimization of extraction of polysaccharides from white hyacinth bean using Plackett–Burman and steepest ascent experiment. Carpath. J. Food Sci. Technol. 2016, 8, 73–82. [Google Scholar]
- Li, X.; Han, L.; Li, Y.; Zhang, J.; Chen, J.; Lu, W. Protective effect of sinapine against hydroxyl radical-induced damage to mesenchymal stem cells and possible mechanisms. Chem. Pharm. Bull. 2016, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Gong, J.; Yang, Y.; Dong, J. Study on Antioxidant Activity In Vitro of Four Kinds of Herbal Polysaccharides. Henan Sci. 2009, 27, 1212–1215. [Google Scholar] [CrossRef]
- Mohammad, A.M.; Koji, Y.; Toshiki, M.; Yumi, N.; Katsumi, S.; Hiroaki, S. Superoxide anion radical scavenging activities of herbs and pastures in northern Japan determined using electron spin resonance spectrometry. Int. J. Biol. Sci. 2007, 3, 349–355. [Google Scholar]
- Chen, Y.; Xie, M.Y.; Nie, S.P.; Li, C.; Wang, Y.X. Purification, composition analysis and antioxidant activity of a polysaccharide from the fruiting bodies of Ganoderma atrum. Food Chem. 2008, 107, 231–241. [Google Scholar] [CrossRef]
- Krings, U.; Berger, R.G. Antioxidant activity of some roasted foods. Food Chem. 2001, 72, 223–229. [Google Scholar] [CrossRef]
- Li, P.; Zhou, L.; Yan, M.; Mao, Z. Extraction optimization of polysaccharide from Zanthoxylum bungeanum using RSM and its antioxidant activity. Int. J. Biol. Macromol. 2015, 72, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Dong, Y.; Chen, G.; Hu, Q. Extraction, purification, characterization and antitumor activity of polysaccharides from Ganoderma lucidum. Carbohydr. Polym. 2010, 80, 783–789. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, M.; Qu, Z.; Xie, B. Antioxidant activities of different fractions of polysaccharide conjugates from green tea (Camellia Sinensis). Food Chem. 2008, 106, 559–563. [Google Scholar] [CrossRef]
- Ramnani, P.; Chitarrari, R.; Tuohy, K. In vitro fermentation and prebiotic potential of novel low molecular weight polysaccharides derived from agar and alginate seaweeds. Anaerobe 2012, 18, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mueller, M.; Čavarkapa, A.; Unger, F.M.; Viernstein, H.; Praznik, W. Prebiotic potential of neutral oligo- and polysaccharides from seed mucilage of Hyptis suaveolens. Food Chem. 2017, 221, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Madhukumar, M.; Muralikrishna, G. Structural characterisation and determination of prebiotic activity of purified xylo-oligosaccharides obtained from Bengal gram husk (Cicer arietinum L.) and wheat bran (Triticum aestivum). Food Chem. 2010, 118, 215–223. [Google Scholar] [CrossRef]
- Hwang, P.A.; Phan, N.N.; Lu, W.J.; Hieu, B.T.N.; Lin, Y.C. Low-molecular-weight fucoidan and high-stability fucoxanthin from brown seaweed exert prebiotics and anti-inflammatory activities in Caco-2 cells. Food Nutr. Res. 2016, 60, 32033. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Wang, X.; Li, G.; Zhao, Y.; Zhang, J.; Niu, C. Antioxidant activity of prebiotic ginseng polysaccharides combined with potential probiotic Lactobacillus plantarum C88. Int. J. Food Sci. Technol. 2015, 50, 1673–1682. [Google Scholar] [CrossRef]
- Biedrzycka, E.; Bielecka, M. Prebiotic effectiveness of fructans of different degrees of polymerization. Trends Food Sci. Technol. 2004, 15, 170–175. [Google Scholar] [CrossRef]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The isolation of the components of streptoeoeeal nueleoproteins in serologieally active form. J. Biol. Chem. 1938, 124, 42–49. [Google Scholar]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Minjares-Fuentes, R.; Femenia, A.; Garau, M.C.; Candelas-Cadillo, M.G.; Simal, S.; Rosselló, C. Ultrasound-assisted extraction of hemicelluloses from grape pomace using response surface methodology. Carbohydr. Polym. 2016, 138, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, N.; Cumbes, Q.J. Hydroxyl radical scavenging activity of compatible solutes. Phytochemistry 1989, 28, 1057–1060. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef] [PubMed]
- Kazuko, S.; Kuniko, F.; Keiko, Y.; Takashi, N. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar]
- Yamaguchi, T.; Takamura, H.; Matoba, T.; Terao, J. HPLC method for evaluation of the free radical-scavenging activity of foods by using 1,1-diphenyl-2-picrylhydrazyl. Biosci. Biotechnol. Biochem. 2014, 62, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
Sample Availability: Samples of the compounds are available from the authors. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Coded Variable Levels | Polysaccharide Yield Y (%) | ||||
|---|---|---|---|---|---|---|
| A | B | C | Actual Value Y | Predicted Value Y’ | Residual Y−Y’ | |
| 1 | 1.682 | 0 | 0 | 3.07 | 2.99 | 0.076 |
| 2 | −1 | 1 | −1 | 3.18 | 3.20 | −0.022 |
| 3 | 0 | 0 | 0 | 3.23 | 3.31 | −0.082 |
| 4 | −1.682 | 0 | 0 | 2.60 | 2.66 | −0.062 |
| 5 | −1 | −1 | 1 | 2.15 | 2.27 | −0.12 |
| 6 | 1 | −1 | −1 | 2.38 | 2.50 | −0.12 |
| 7 | 0 | 0 | 0 | 3.41 | 3.31 | 0.098 |
| 8 | 0 | 0 | −1.682 | 3.07 | 3.13 | −0.056 |
| 9 | −1 | −1 | −1 | 2.57 | 2.45 | 0.12 |
| 10 | 0 | 0 | 0 | 3.26 | 3.31 | −0.052 |
| 11 | 1 | 1 | −1 | 3.09 | 2.98 | 0.11 |
| 12 | 1 | 1 | 1 | 2.40 | 2.53 | −0.13 |
| 13 | 0 | 0 | 0 | 3.37 | 3.31 | 0.058 |
| 14 | 0 | 0 | 0 | 3.31 | 3.21 | −0.0021 |
| 15 | 0 | 1.682 | 0 | 2.43 | 2.48 | −0.046 |
| 16 | 0 | −1.682 | 0 | 2.20 | 2.14 | 0.060 |
| 17 | 0 | 0 | 1.682 | 2.67 | 2.60 | 0.069 |
| 18 | 0 | 0 | 0 | 3.29 | 3.31 | −0.022 |
| 19 | 1 | −1 | 1 | 2.90 | 2.89 | 0.013 |
| 20 | −1 | 1 | 1 | 2.31 | 2.19 | 0.12 |
| Source | SS | DF | MS | F | Pr > F | Sig |
|---|---|---|---|---|---|---|
| Model | 3.409 | 9 | 0.379 | 28.109 | <0.0001 | *** |
| A | 0.134 | 1 | 0.134 | 9.909 | 0.0104 | * |
| B | 0.137 | 1 | 0.137 | 10.150 | 0.0097 | ** |
| C | 0.333 | 1 | 0.333 | 24.714 | 0.0006 | *** |
| AB | 0.039 | 1 | 0.039 | 2.909 | 0.1189 | |
| AC | 0.157 | 1 | 0.157 | 11.635 | 0.0066 | ** |
| BC | 0.344 | 1 | 0.344 | 25.559 | 0.0005 | *** |
| A2 | 0.422 | 1 | 0.422 | 31.299 | 0.0002 | *** |
| B2 | 1.815 | 1 | 1.815 | 134.713 | <0.0001 | *** |
| C2 | 0.363 | 1 | 0.363 | 26.935 | 0.0004 | *** |
| Residual error | 0.135 | 10 | 0.013 | |||
| Lack of fit | 0.112 | 5 | 0.022 | 4.889 | 0.0532 | |
| Pure error | 0.023 | 5 | 0.005 | |||
| Total | 3.544 | 19 |
| Variable | Level | ||||
|---|---|---|---|---|---|
| −1.682 | −1 | 0 | 1 | 1.682 | |
| A, pH value | 7.46 | 7.60 | 7.80 | 8.00 | 8.14 |
| B, cellulase (%) | 2.53 | 2.60 | 2.70 | 2.80 | 2.87 |
| C, Ratio (v/w) of water (mL) to material (g) | 60.32 | 61.00 | 62.00 | 63.00 | 63.68 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shu, G.-W.; He, Y.-X.; Lei, N.; Cao, J.-L.; Chen, H.; Chen, L. Cellulase-Assisted Extraction of Polysaccharides from White Hyacinth Bean: Characterization of Antioxidant Activity and Promotion for Probiotics Proliferation. Molecules 2017, 22, 1764. https://doi.org/10.3390/molecules22101764
Shu G-W, He Y-X, Lei N, Cao J-L, Chen H, Chen L. Cellulase-Assisted Extraction of Polysaccharides from White Hyacinth Bean: Characterization of Antioxidant Activity and Promotion for Probiotics Proliferation. Molecules. 2017; 22(10):1764. https://doi.org/10.3390/molecules22101764
Chicago/Turabian StyleShu, Guo-Wei, Yun-Xia He, Ni Lei, Ji-Li Cao, He Chen, and Li Chen. 2017. "Cellulase-Assisted Extraction of Polysaccharides from White Hyacinth Bean: Characterization of Antioxidant Activity and Promotion for Probiotics Proliferation" Molecules 22, no. 10: 1764. https://doi.org/10.3390/molecules22101764