
Promiscuous Effects of Some Phenolic Natural Products on Inflammation at Least in Part Arise from Their Ability to Modulate the Expression of Global Regulators, Namely microRNAs

Abstract
:1. Introduction
2. Phytochemicals and Inflammatory Factors
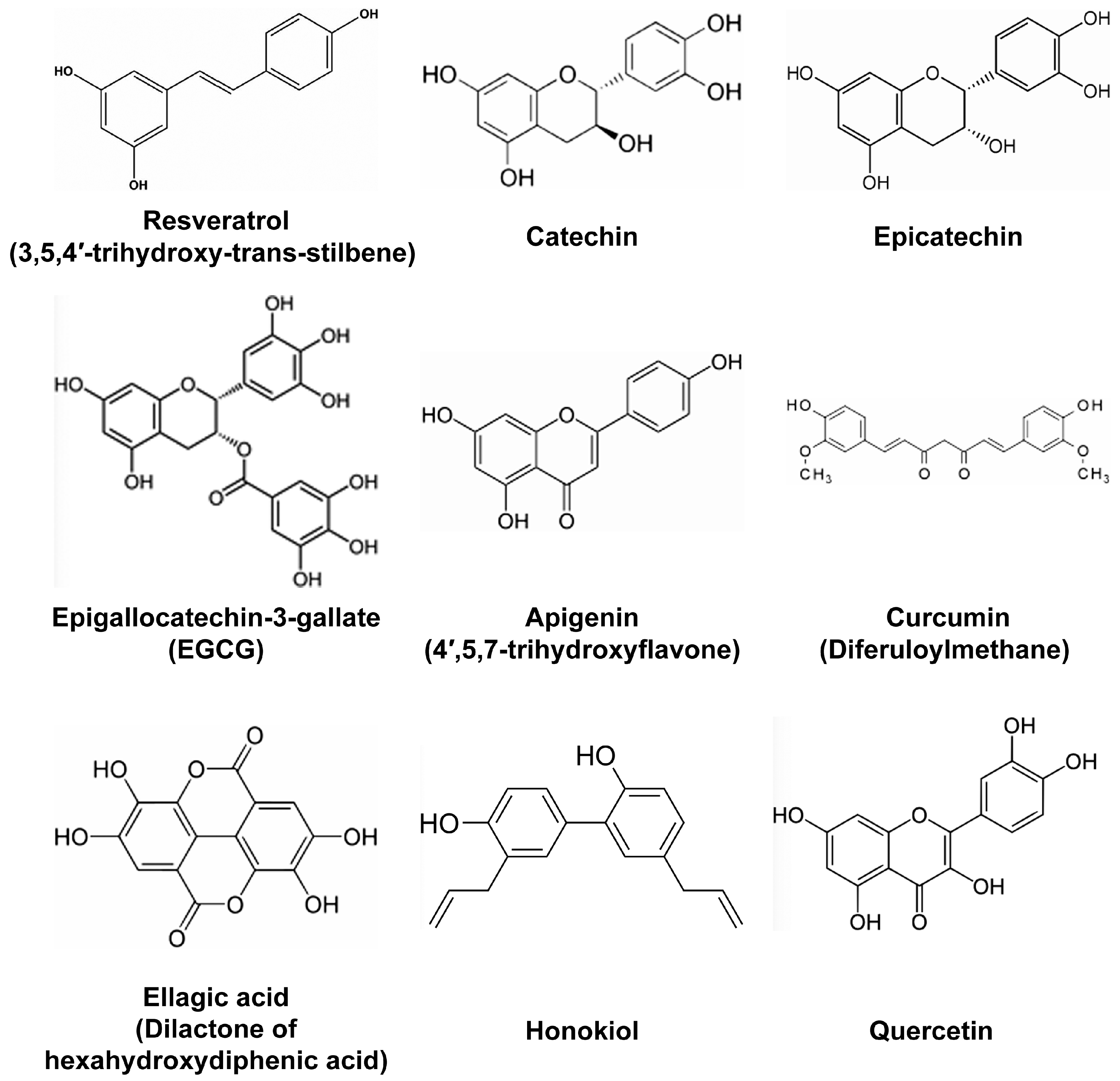
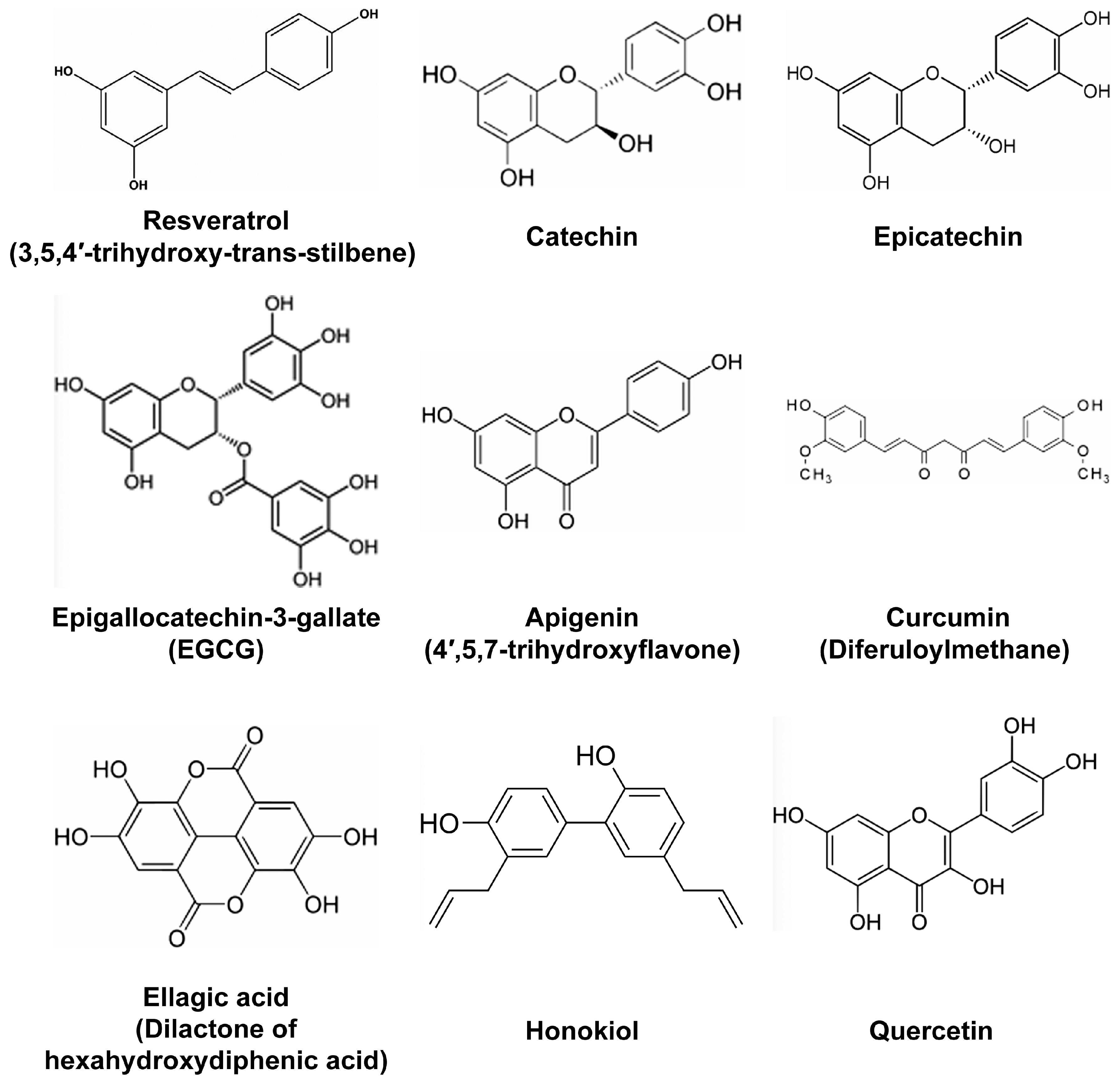
- Resveratrol (Figure 1) is probably the most studied polyphenol with anti-inflammatory properties. It has been proposed that in addition to being ingested with food derived from grapes and other berries, resveratrol could possibly be used as a cream for the treatment of epithelium lesions such as dermatitis. Indeed, in a murine model of atopic dermatitis-like lesions induced by the application of 2,4-dinitrofluorobenzene, systemic administration of resveratrol ameliorated the outcome by reducing the expression levels of epithelium derived cytokines, such as IL-25, IL-33, and thymic stromal lymphopoietin [17]. Reduced levels of these factors following resveratrol treatment was associated with the reduction of apoptotic epithelial cells as measured by the levels of caspase-3 and the increased thickness of the epithelium [17]. In another study, the polyphenols-rich fraction of grapes reduced the metabolic consequences of high-fat diet by decreasing adiposity and insulin resistance in addition to reducing inflammatory markers [14]. Interestingly, the same extract also impacted the gut microbiota in high fat-fed mice, as mice fed on these extracts showed alterations in gut microbiota groups. Another study found that resveratrol improved renal injury caused by micro-inflammation in a hypertensive renal damage rat model [18]. In this model, resveratrol exerted anti-fibrotic functions by reducing the levels of Plasminogen activator inhibitor (PAI)-1, TGF-β, and Fibronectin [18]. Of note, our group previously reported that TGF-β pathway is the primary target of resveratrol in human colon cancer cells [19]. We came to this conclusion, based on the fact that resveratrol changed the expression of different microRNAs targeting effectors of TGF-β signaling, including TGF-β itself [19]. Resveratrol anti-fibrotic effects in the hypertensive renal damage rat model were complemented with anti-inflammatory effects, as the expression of NF-κB, IL-6, Intercellular adhesion molecule (ICAM)-1 and Monocyte chemoattractant protein-1 (MCP) was significantly reduced [18]. Based on its anti-inflammatory properties, a combinatorial use of resveratrol with non-steroidal anti-inflammatory drug (NSAID) ibuprofen was tested for the ability to diminish innate immune response on lipopolysaccharide (LPS)-challenged RAW-264.7 mouse macrophages [20]. In parallel, different resveratrol-NSAID derivatives were tested in the same conditions, and one of these derivatives was found to present stronger anti-inflammatory properties than resveratrol, ibuprofen or their mixture. As mentioned earlier, resveratrol has been known to also reduce neuro-inflammation and might be of interest for slowing the progression of neuro-inflammatory pathologies. For example, resveratrol was tested for memory enhancing and its effects on cognitive disorders such as Alzheimer’s disease in a mouse model of this pathology [21]. In Alzheimer’s disease, Beta amyloid (Aβ) peptides are implicated in cognitive impairment, neuro-inflammation, and neuronal apoptosis [21]. Therefore, the effects of resveratrol were tested in mice microfused with Aβ peptides to induce learning/memory impairment, neuro-inflammation and apoptosis. Resveratrol was found to reverse Aβ1-42-induced expression of Phosphodiesterase 4A, 4B and 4D, and reverse Aβ1-42-induced decrease in the phosphorylation level of cAMP response-element binding protein (CREB), Brain derived neurotrophic factor (BDNF) and Anti-apoptotic factor B-cell CLL/lymphoma (Bcl)-2 [21].
- Curcuminoids, with curcumin (Figure 1) being the most studied, are known for their anti-inflammatory and free radical-scavenging activities. Among critical discoveries was the study that found curcumin supplementation to significantly reduce circulating levels of TNF in randomized clinical trials using meta-analysis [22]. In rats, curcumin was found to alleviate renal dysfunction and suppress inflammation in daunorubicin-induced nephrotoxicity by shifting macrophage polarization toward the M2 phenotype [23]. This shift of macrophage polarization to M2 phenotype in the presence of curcumin was due to the reduction of Extracellular signal–regulated kinase (ERK)1/2 and NF-κB p65 expression, concomitant to the upregulation of IL-10 expression [23]. In another study, curcumin significantly reduced oxidative stress and inflammation in a rat model of gastric antral ulceration [24]. In curcumin-fed rats, the inhibition of lipid peroxidation and naproxen-induced gastric ulcer was attributed to the effects of curcumin in increasing the activities of scavenging enzymes, such as Superoxide dismutase, Catalase, and Glutathione peroxidase [24]. The effects of polyphenols found in French Curcuma longa, were tested, in 3T3-L1 adipocytes under oxidative stress [25]. These polyphenols improved insulin-mediated lipid accumulation, and increased Peroxisome proliferator-activated receptor-gamma (PPARγ) gene expression and adiponectin secretion. The same polyphenols also impaired oxidative stress-induced production of IL-6, TNF, MCP-1, and NF-κB [25]. Expectedly, these polyphenols also reduced intracellular levels of ROS and modulated the expression of genes encoding Superoxide dismutase (SOD) and Catalase antioxidant enzymes [25]. Additionally, curcumin, was tested, for its fungicidal effects on Sporothrix schenckii fungus [26]. This study found that sublethal curcumin treatments of S. schenckii activate protein kinase C (PKC) and increase the expression of Chitin synthase 1 and 3, resulting in increased chitin content on the S. schcenckii conidial cell wall [26]. As a result, mice infected with curcumin-treated conidia showed reduced fungal burden. Thus, like resveratrol, curcumin presents with a wide range of anti-inflammatory effects, suggesting that both compounds might affect the expression of microRNAs involved in immune response and inflammation.
- Tannin-rich fruits tested for their anti-inflammatory effects, in a preclinical model of colitis, showed protective effects against ulcerative colitis. Beverages prepared from mango and pomegranate (Punica granatum) rich in gallotannins and ellagitannins, respectively, were tested in a rat sulfate-induced colitis model [27]. While both beverages reduced intestinal inflammation and the levels of pro-inflammatory cytokines in mucosa and serum, they also showed specific effects. Thus, mango beverage impaired the strong activation of Insulin-like growth factor (IGF)-1/AKT/rapamycin (mTOR) pathway, by reducing the expression of IGF1 and Insulin receptor (INSR) genes, while pomegranate reduced the levels of Ribosomal protein S6 kinase beta-1 (p70S6K), Ribosomal protein S6 (RPS6), as well as those of transcripts encoding Ribosomal protein S6 kinase A2 (RPS6KA2), Mitogen-activated protein kinase kinase 2 (MAP2K2), and MAPK1 [27]. Finally, polyphenolic extracts prepared from these two beverages inhibited IGF-1R/AKT/mTOR pathway (mango) or ERK1/2-mTOR pathway (pomegranate), respectively [27]. In addition, in silico analysis showed the presence of a high binding-dock of gallic acid to the catalytic domain of IGF-1R, while ellagic acid could dock effectively into Epidermal growth factor receptor (EGFR) and IGF-1R, opening the possibility to use gallic acid to specifically impair EGFR pathway [27]. These data point to specific effects of these two-tannin rich fruits in colitis, and suggest that a combination of both extracts potentially could have complementary protective effects against ulcerative colitis. This study supports the hypothesis that active compounds found in our diet, although highly similar, might have specific effects on cellular signaling and pathways they modulate in the cell. In addition, their activity depends on the “healthy” state of the cell, so that we cannot expect phenolic NPs to have identical effects in normal versus pathological conditions on a given cell.
- EGCG, the most abundant polyphenol of green tea, has widely been studied for its anti-oxidant, anti-cancer and anti-inflammatory effects. The anti-inflammatory effects of EGCG are of particular interest for treatment of inflammatory bowel disease (IBD) [28]. In inflamed colon, EGCG reduces myeloperoxidase activity and production of pro-inflammatory factors, and provides a general protection of gut mucosa [29]. From a metabolic point of view, green tea phytochemicals reduce body mass index and waist circumference and improve lipid metabolism [30]. In another study, EGCG reduced inflammation and pain caused by intervertebral disc degeneration [31]. Interestingly, in addition to reducing inflammation, EGCG enhanced the survival of primary human disc cells under oxidative stress. Furthermore, EGCG enhanced the survival of the same cells in lethal oxidative stress conditions through activation of pro-survival PI3K/AKT pathway. It will be important to determine parameters that allow the tested compounds to be either pro-apoptotic (thus being beneficial for cancer treatment) or anti-apoptotic (thus protecting cells from death induced by ischemic or oxidative stress).
- Anti-inflammatory effects of polyphenols found in cocoa extracts were tested on mouse RAW-264.7 macrophages challenged with LPS to induce innate immune response [32]. The reaction of RAW-264.7 macrophages to LPS was significantly attenuated by cocoa extracts polyphenols, as measured by reduced levels of pro-inflammatory enzymes such as 5-Lipoxigenase (5-LOX), prostaglandin E2 (PGE2), reactive oxygen species (ROS), nitric oxide (NO), and TNF [32]. Analogous experiments with similar outcomes were conducted with phenolic and terpene extracts from the leaves of different species of Oregano [33]. Additionally, cocoa supplements, were reported to improve endothelial cell function [34], as did Chilean propolis [35], or resveratrol on human colon-derived CCD-18Co myofibroblast cells [36].
- The serum of mice pre-treated with banana peel polyphenols presented with reduced levels of pro-inflammatory cytokines such as TNF, IL-6, IL-1β, and IL-12 following treatment with carbon tetrachloride, a molecule used to induce hepatic injury [37]. Other inflammatory-related factors such as COX-2, and NO were also reduced. Banana peel polyphenols likewise reduced serum levels of Aspartate aminotransferase, Alanine aminotransferase and Lactate dehydrogenase enzymes. Overall, the capability of banana peel polyphenols to reduce the extent of carbon tetrachloride-induced liver injury and to protect liver cells, were attributed to their anti-inflammatory properties [37]. Phytochemicals present in methanol extracts of Algerian Hertia cheirifolia (rich in flavonoids and polyphenols) were tested for their anti-inflammatory effects in an experimental model of croton oil-induced ear edema in albino mice [38]. Topical application of these extracts in the ear of mice inhibited ear swelling caused by croton oil. This treatment also inhibited leucocyte migration and significantly reduced the release of TNF and IL-1β [38].
- Diets supplemented with ellagic acid (Figure 1), a polyphenol found in raspberry seed flour and other fruits or nuts, were effective in reducing high-fat high sucrose-induced metabolic stress by attenuating hepatic endoplasmic reticulum stress, ROS and adipocyte inflammation in mouse [39]. As high fat-high sucrose diet has become part of “western” diet and is the cause of high obesity in western countries, inclusion of ellagic acid either in diet (consummation of fruits and nuts) or as a supplement, potentially might be used to normalize metabolic insult triggered particularly by high sucrose diet. Other berries also deliver beneficial effects against metabolic stress, specifically diabetes mellitus. Thus, consumption of polyphenol-rich white mulberry (Morus alba) leaves attenuates diabetic retinopathy in rats [40]. Particularly, rats fed with M. alba extracts showed decreased levels of sorbitol, fructose, PKC, and pro-inflammatory cytokines, reduced apoptosis, reduction of Vascular endothelial growth factor (VEGF) expression, and attenuated oxidative stress markers in their retinas [40]. Even more interesting is the fact that M. alba-rich diet significantly attenuated hyperglycemia and weight loss. Considering that weight loss is a side effect of pancreatic cancer patients, it might be interesting to test these extracts on rodent models of cachexia. In high fat-fed mice, blood glucose concentration, hepatic metabolism and obesity-related gene expression signature were also improved by the intake of quercetin (Figure 1) and quercetin-containing apple and cherry extracts. In obese high-fat fed mice, phytochemicals from the same fruits also reduced hepatic lipid accumulation, and lowered blood glucose concentration after food deprivation [41]. Finally, extracts of Acai-berry inhibited osteoclastogenesis responsible for bone loss and known to be triggered by inflammation. RAW-264.7 macrophages challenged with Receptor activator of nuclear factor kappa-B ligand (RANKL) to induce osteoclastogenesis-like in vitro, showed attenuated osteoclast activity following treatment with Acai-berry extracts, as measured by tartrate-resistant acid phosphatase and hydroxyapatite resorption assay [42]. In parallel, Acai-berry extracts reduced the secretion of IL-1α, IL-6 and TNF, while increasing the secretion of IL-3, IL-4, IL-13 and interferon gamma (IFN-γ) [42].
- In human umbilical vein endothelial cells (HUVECs), polyphenols-enriched fraction from Chilean propolis was shown to inhibit activation of Hif-1α and Erk1/2 as well as expression of VEGFA [43]. Polyphenols are likewise tested for therapeutic prevention of malignant mesothelioma [44]. Malignant mesothelioma is caused by exposure to asbestos that triggers chronic inflammation of mesothelial cells. In addition, asbestos also reduces the ability of the immune system to mount a functional response to transformed mesothelial cells. Current knowledge suggests that polyphenols are able to reduce inflammation and also inhibit cancer cell growth, hence the consideration for using polyphenols for prevention of malignant mesothelioma [44].
3. Phytochemicals and microRNAs
- Polyphenols found in Chilean propolis were tested for their anti-angiogenic effects in atherosclerotic plaques from LDL receptor knockout mice [35] (Table 1). Propolis attenuated cell migration, capillary-like tube formation and sprouting by inhibiting the activation of Hypoxia-inducible factor 1-alpha (HIF-1α) and ERK1/2 kinases, as well as the expression of Vascular endothelial growth factor (VEGF) [35] (Table 1). Mango extracts that, as mentioned earlier, show anti-inflammatory and anti-oxidant properties suppressed the expression of NF-κB, PI3K (p85β), HIF-1α, p70S6K1, RPS6 and iNOS in LPS-treated CCD-18Co cells [72] (Table 1). Simultaneously, mango extracts had opposing effects on the expression of miR-126, with LPS reducing miR-126 expression and mango extracts increasing it [72]. MiR-126 was shown to target PI3K (p85β) transcripts. As most of the transcripts whose expression was changed by mango extracts encode members of the PI3K/AKT/mTOR pathway, it has been suggested that mango extracts impair the activation of this pathway by upregulating miR-126. It clearly remains to uncover the specific mechanisms of miR-126 upregulation in this setting, as miR-126 is crucial for the normal function of endothelial cells, and its expression is deregulated in several cardiovascular diseases, hence the interest of manipulating the levels of this microRNA [73]. MiR-126 is likewise the target of other polyphenols such as those derived from cowpea (Vigna unguiculata) or from pomegranate, both groups known for their anti-inflammatory properties [74]. Accordingly, the treatment of LPS-challenged colonic myofibroblasts with different polyphenols from cowpea plant impaired the robust induction by LPS of pro-inflammatory factors such as IL-8, TNF, VCAM-1, and NF-κB, while increasing the levels of miR-126, that itself targets VCAM [74] (Table 1). Likewise, pomegranate juice given to rats prior to inducing colorectal aberrant crypt foci using azoxymethane reduced azoxymethane effects and lowered the proliferation of mucosa cells by increasing the expression of miR-126 [75]. Pomegranate-induced miR-126 reduced the levels of VCAM-1 and PI3K p85β target transcripts as well as the expression of pro-inflammatory cytokines, NF-κB, iNOS and COX-2 [75] (Table 1). Another polyphenol-containing substance similarly acting as an anti-angiogenic factor is propolis. When LDL receptor knockout mice that are prone to atherosclerotic plaques were treated with propolis, atherosclerotic lesions were significantly attenuated and the expression of pro-angiogenic factor VEGF and of Hif-1α was reduced [35] (Table 1). Hif-1α, known to positively control the expression of VEGF at the transcriptional level, was expressed in the necrotic nucleus of atheromas. In addition to the transcriptional control of VEGF, propolis treatment significantly increased the expression of miR-181a, miR-106a and miR-20b [35]. These three microRNAs were found to target and reduce the expression of transcripts encoding both VEGF and Hif-1α. Thus, anti-angiogenic effects of propolis are at least through these three microRNAs in addition to its effects on protein coding genes.
- MiR-34 has been an attractive microRNA for its therapeutic potentials against cancer, and miR-34-based therapeutics are now on clinical trials [76,77,78]. Honokiol (Figure 1), a polyphenol present in Magnolia grandiflora, was found to upregulate the expression of miR-34a, which in turn targets and inhibits the Wnt-1-Metastasis-associated protein (MTA)1-β-catenin pathway (Table 1), thus counteracting oncogenic effects of leptin on breast cancer growth and metastatic properties of human breast cancer cell lines MCF7, MDA-MB-231, MDA-MB-468 and T47D [79]. Based on a phospho-kinase screening array, honokiol was found to inhibit the phosphorylation and activation of factors involved in Leptin-signaling. Accordingly, honokiol inhibited the phosphorylation of Stat3, a transcription factor that binds to and represses the expression of miR-34 [79]. Of note, in breast cancer, Wnt-1-MTA1-β-catenin pathway is activated by Leptin, and is considered a critical therapeutic target. Other polyphenols derived from pomegranate also induced the expression of miR-34a, this time in EJ bladder cancer cells, where miR-34a was reported to target c-Myc and CD44 transcripts [80] (Table 1).
- Another microRNA involved in inflammation and cancer is miR-181b [81]. MiR-181b binds to the 3′-UTR of transcripts encoding pro-inflammatory chemokines (C-X-C motif) ligand (CXCL)1 and CXCL2 and simultaneously reduces their expression in metastatic breast cancer cells [82] (Table 1). Curcumin had similar effects on these two transcripts, suggesting that its effects on breast cancer cells are potentially due to its upregulation of miR-181b expression [82]. Indeed microRNA microarray analysis of curcumin-treated breast cancer cells showed that this phytochemical upregulates the expression of miR-181b and reduces metastatic progression. Curcumin also significantly reduced the growth of flank murine melanoma tumors by substantially upregulating the expression of miR-205-5p [83] (Table 1). Of interest to note is also the fact that curcumin altered the expression of microRNAs whose corresponding putative target transcripts are implicated in O-glycan biosynthesis and endoplasmic reticulum protein maturation and/or processing. These results suggest testing curcumin in other types of cancer where miR-205-5p and the above-mentioned mechanisms are involved.
- As mentioned earlier, the flavonoid apigenin has anti-viral activity [15]. This activity was attributed to apigenin acting as an inhibitor of microRNA maturation, especially that of miR-122, previously shown to positively regulate hepatitis C virus replication [15] (Table 1). In human liver Huh7 cells, apigenin blocked microRNA maturation by preventing phosphorylation and consequently activation of TAR RNA binding protein (TRBP) enzyme, which is involved in microRNA maturation [15]. The impairment of microRNA maturation was associated with the reduction of hepatitis C virus replication. This finding is of great interest for patients chronically infected by hepatitis C virus. In (miR103)-overexpressing transgenic mice presenting with glucose intolerance, apigenin improved this pathogenic status likely by decreasing matured miR103 expression levels through inhibition of TRBP phosphorylation [84] (Table 1). Given the above results, it would certainly be of interest to look for additional compounds with anti-bacterial, anti-viral, or anti-parasite activity that might impact affect the expression of microRNAs capable to inhibit the survival and/or the functions of pathogens.
- With regard to phytochemicals and oxidative stress, it did not come as a surprise that different phytochemicals induce the expression of miR-210, the most critical microRNA of oxidative stress response [62,85]. This induction was primarily at the transcription level, as it was found that phytochemicals such as EGCG upregulate the expression of HIF-1α, the transcription factor for miR-210 [86] (Table 1).
- MiR-21 is an oncogenic microRNA upregulated during the immune response and highly expressed in nearly all solid and liquid malignancies. Targeted downregulation of miR-21 therefore is of upmost interest in the field of cancer [45]. Opposite to cancer cells, miR-21 expression is significantly reduced in ischemic heart, thus restoring its expression in these conditions would be desirable [87]. Therefore, identifying compounds that could either increase or reduce miR-21 expression is critical. Certainly, phytochemicals should have specific effects based on the cell type, cell condition, cell function, and depending primarily on the cell transcriptome, metabolome, and proteome. Therefore, it is most likely that cell-specific conditions affecting phytochemicals effects would also impact phytochemicals effects on miR-21 expression. This microRNA has often been associated with the activity of NF-κB [45]. Resveratrol treatment of U251 brain tumor cells induces apoptosis and decreases miR-21 expression and NF-κB activity [88] (Table 1). On the other hand, miR-21 was shown to protect the heart from ischemia through its anti-apoptotic activity. In cardiomyocytes, miR-21 targets and consequently decreases the levels of transcripts encoding Fas ligand, a pro-apoptotic factor [89]. In addition to miR-21, another microRNA involved in neuroblastoma is miR-7-1 [90]. The expression of this microRNA increases in malignant neuroblastoma cell lines following treatment with either EGC or EGCG [90]. Upon its upregulation by EGCG or EGC treatment, miR-7-1 contributes to the induction of cell apoptosis [90]. The pro-apoptotic activity of EGCG and EGC also arises from their effects on the expression of other microRNAs, including the downregulation of oncogenic microRNAs from the miR-17-92 cluster family: specifically, miR-92, miR-93 and miR-106b [90] (Table 1). This downregulation is probably through modulation of transcription factors that control the expression of this cluster, such as GAM [91].
- A combinatorial-correlative computer based analysis based on a mouse tobacco carcinogen-induced lung tumor model led to the identification of a signature transcripts/microRNAs/pathways associated with EGCG treatments [92], with NF-κB, AKT, and MAP kinase pathways being the major pathways affected by this compound. This study found twelve microRNAs upregulated and nine microRNAs downregulated by EGCG (Table 1).
4. The Gut Microbiota and the Bioavailability of Phytochemicals
5. Closing Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Kotas, M.E.; Medzhitov, R. Homeostasis, inflammation, and disease susceptibility. Cell 2015, 60, 816–827. [Google Scholar] [CrossRef] [PubMed]
- Varoni, E.M.; Lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer Molecular Mechanisms of Resveratrol. Front. Nutr. 2016, 3, 8. [Google Scholar] [CrossRef] [PubMed]
- Lançon, A.; Kaminski, J.; Tili, E.; Michaille, J.-J.; Latruffe, N. Control of microRNA expression as a new way for resveratrol to deliver its beneficial effects. J. Agric. Food. Chem. 2012, 60, 8783–8739. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Safer, A.M.; Menon, M. Green tea polyphenols and their potential role in health and disease. Inflammopharmacology 2015, 23, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Banerjee, S.; Sil, P.C. The beneficial role of curcumin on inflammation, diabetes and neurodegenerative disease: A recent update. Food. Chem. Toxicol. 2015, 83, 111–124. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Lee, Y.M.; Lee, C.K.; Jung, J.K.; Han, S.B.; Hong, J.T. Therapeutic applications of compounds in the Magnolia family. Pharmacol. Ther. 2011, 130, 157–176. [Google Scholar] [CrossRef] [PubMed]
- Bladé, C.; Aragonès, G.; Arola-Arnal, A.; Muguerza, B.; Bravo, F.I.; Salvadó, M.J.; Arola, L.; Suárez, M. Proanthocyanidins in health and disease. Biofactors 2016, 42, 5–12. [Google Scholar] [PubMed]
- Goya, L.; Martín, M.Á.; Sarriá, B.; Ramos, S.; Mateos, R.; Bravo, L. Effect of Cocoa and Its Flavonoids on Biomarkers of Inflammation: Studies of Cell Culture, Animals and Humans. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Leyva-López, N.; Gutierrez-Grijalva, E.P.; Ambriz-Perez, D.L.; Heredia, J.B. Flavonoids as Cytokine Modulators: A Possible Therapy for Inflammation-Related Diseases. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Chirumbolo, S. The role of quercetin, flavonols and flavones in modulating inflammatory cell function. Inflamm. Allergy Drug Targets 2010, 9, 263–285. [Google Scholar] [CrossRef] [PubMed]
- Korkina, L.G. Phenylpropanoids as naturally occurring antioxidants: From plant defense to human health. Cell. Mol. Biol. (Noisy-le-Grand) 2007, 53, 15–25. [Google Scholar]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Collins, B.; Hoffman, J.; Martinez, K.; Grace, M.; Lila, M.A.; Cockrell, C.; Nadimpalli, A.; Chang, E.; Chuang, C.C.; Zhong, W.; et al. A polyphenol-rich fraction obtained from table grapes decreases adiposity, insulin resistance and markers of inflammation and impacts gut microbiota in high-fat-fed mice. J. Nutr. Biochem. 2016, 31, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Shibata, C.; Ohno, M.; Otsuka, M.; Kishikawa, T.; Goto, K.; Muroyama, R.; Kato, N.; Yoshikawa, T.; Takata, A.; Koike, K. The flavonoid apigenin inhibits hepatitis C virus replication by decreasing mature microRNA122 levels. Virology 2014, 42, 462–463. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J. Resveratrol, MicroRNAs, Inflammation, and Cancer. J. Nucleic Acids 2011, 2011, 102431. [Google Scholar] [CrossRef] [PubMed]
- Caglayan Sozmen, S.; Karaman, M.; Cilaker Micili, S.; Isik, S.; Arikan Ayyildiz, Z.; Bagriyanik, A.; Uzuner, N.; Karaman, O. Resveratrol ameliorates 2,4-dinitrofluorobenzene-induced atopic dermatitis-like lesions through effects on the epithelium. PeerJ 2016, 4, e1889. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Li, Y.; Cao, Y.; Fan, Y.; Chen, X.; Huang, X.Z. Resveratrol ameliorates renal injury in spontaneously hypertensive rats by inhibiting renal micro-inflammation. Biosci. Rep. 2016. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Alder, H.; Volinia, S.; Delmas, D.; Latruffe, N.; Croce, C.M. Resveratrol modulates the levels of microRNAs targeting genes encoding tumor-suppressors and effectors of TGF beta signaling pathway in SW480 cells. Biochem. Pharmacol. 2010, 80, 2057–2065. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Ma, Y.Y.; Zhang, K.; Zhou, A.Y.; Zhang, Y.; Wang, H.; Du, Z.; Zhao, D.G. Synthesis and biological evaluation of novel resveratrol-NSAID derivatives as anti-inflammatory agents. Chem. Pharm. Bull. (Tokyo) 2016. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Chen, L.; Pan, X.; Chen, J.; Wang, L.; Wang, W.; Cheng, R.; Wu, F.; Feng, X.; Yu, Y.; et al. The effect of resveratrol on beta amyloid-induced memory impairment involves inhibition of phosphodiesterase-4 related signaling. Oncotarget 2016. [Google Scholar] [CrossRef] [PubMed]
- Sahebkar, A.; Cicero, A.F.; Simental-Mendía, L.E.; Aggarwal, B.B.; Gupta, S.C. Curcumin downregulates human tumor necrosis factor-α levels: A systematic review and meta-analysis of randomized controlled trials. Pharmacol. Res. 2016, 107, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Sreedhar, R.; Giridharan, V.V.; Afrin, R.; Harima, M.; Miyashita, S.; Hara, M.; Suzuki, K.; et al. Curcumin alleviates renal dysfunction and suppresses inflammation by shifting from M1 to M2 macrophage polarization in daunorubicin induced nephrotoxicity in rats. Cytokine 2016, 84, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jin, S.; Kwon, H.J.; Kim, B.W. Curcumin Blocks Naproxen-induced Gastric Antral Ulcerations through Inhibition of Lipid Peroxidation and Activation of Enzymatic Scavengers in Rats. J. Microbiol. Biotechnol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Septembre-Malaterre, A.; Le Sage, F.; Hatia, S.; Catan, A.; Janci, L.; Gonthier, M.P. Curcuma longa polyphenols improve insulin-mediated lipid accumulation and attenuate proinflammatory response of 3T3-L1 adipose cells during oxidative stress through regulation of key adipokines and antioxidant enzymes. Biofactors 2016. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhang, J.; Song, T.; Yuan, L.; Zhou, J.; Yin, H.; He, T.; Gao, W.; Sun, Y.; Hu, X.; et al. Antifungal curcumin promotes chitin accumulation associated with decreased virulence of Sporothrix schenckii. Int. Immunopharmacol. 2016, 34, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Banerjee, N.; Ivanov, I.; Pfent, C.M.; Prudhomme, K.R.; Bisson, W.H.; Dashwood, R.H.; Talcott, S.T.; Mertens-Talcott, S.U. Comparison of anti-inflammatory mechanisms of mango (Mangifera indica, L.) and pomegranate (Punica granatum, L.) in a preclinical model of colitis. Mol. Nutr. Food. Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Farzaei, M.H.; Rahimi, R.; Abdollahi, M. The role of dietary polyphenols in the management of inflammatory bowel disease. Curr. Pharm. Biotechnol. 2015, 16, 196–210. [Google Scholar] [CrossRef] [PubMed]
- Yeoh, B.S.; Aguilera Olvera, R.; Singh, V.; Xiao, X.; Kennett, M.J.; Joe, B.; Lambert, J.D.; Vijay-Kumar, M. Epigallocatechin-3-gallate inhibition of myeloperoxidase and its counter-regulation by Dietary Iron and Lipocalin 2 in Murine Model of Gut Inflammation. Am. J. Pathol. 2016, 186, 912–926. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016. [Google Scholar] [CrossRef] [PubMed]
- Krupkova, O.; Handa, J.; Hlavna, M.; Klasen, J.; Ospelt, C.; Ferguson, S.J.; Wuertz-Kozak, K. The natural polyphenol epigallocatechin gallate protects intervertebral disc cells from oxidative stress. Oxid. Med. Cell. Longev. 2016, 2016, 7031397. [Google Scholar] [CrossRef] [PubMed]
- Ranneh, Y.; Ali, F.; Al-Qubaisi, M.; Esa, N.M.; Ismail, A. The inhibitory activity of cocoa phenolic extract against pro-inflammatory mediators secretion induced by lipopolysaccharide in RAW 264.7 cells. SpringerPlus 2016, 5, 547. [Google Scholar] [CrossRef] [PubMed]
- Leyva-López, N.; Nair, V.; Bang, W.Y.; Cisneros-Zevallos, L.; Heredia, J.B. Protective role of terpenes and polyphenols from three species of Oregano (Lippia graveolens, Lippia palmeri and Hedeoma patens) on the suppression of lipopolysaccharide-induced inflammation in RAW 264.7 macrophage cells. J. Ethnopharmacol. 2016, 187, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Steffen, Y.; Schewe, T.; Sies, H. Myeloperoxidase-mediated LDL oxidation and endothelial cell toxicity of oxidized LDL: Attenuation by (−)-epicatechin. Free Radic. Res. 2006, 40, 1076–1085. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, A.; Saavedra, N.; Cavalcante, M.F.; Salazar, L.A.; Abdalla, D.S. Identification of microRNAs involved in the modulation of pro-angiogenic factors in atherosclerosis by a polyphenol-rich extract from propolis. Arch. Biochem. Biophys. 2014, 557, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Angel-Morales, G.; Noratto, G.; Mertens-Talcott, S. Red wine polyphenolics reduce the expression of inflammation markers in human colon-derived CCD-18Co myofibroblast cells: Potential role of microRNA-126. Food Funct. 2012, 3, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Feng, X.; Zhu, K.; Zhao, X.; Suo, H. Preventive activity of banana peel polyphenols on CCl4-induced experimental hepatic injury in Kunming mice. Exp. Ther. Med. 2016, 11, 1947–1954. [Google Scholar] [CrossRef] [PubMed]
- Bouriche, H.; Kada, S.; Assaf, A.M.; Senator, A.; Gül, F.; Dimertas, I. Phytochemical screening and anti-inflammatory properties of Algerian Hertia cheirifolia methanol extract. Pharm. Biol. 2016, 9, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kang, I.; Espín, J.C.; Carr, T.P.; Tomás-Barberán, F.A.; Chung, S. Raspberry seed flour attenuates high-sucrose diet-mediated hepatic stress and adipose tissue inflammation. J. Nutr. Biochem. 2016, 32, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Abd El-Twab, S.M.; Abdel-Reheim, E.S. Consumption of polyphenol-rich Morus alba leaves extract attenuates early diabetic retinopathy: The underlying mechanism. Eur. J. Nutr. 2016. [Google Scholar] [CrossRef] [PubMed]
- Snyder, S.M.; Zhao, B.; Luo, T.; Kaiser, C.; Cavender, G.; Hamilton-Reeves, J.; Sullivan, D.K.; Shay, N.F. Consumption of Quercetin and Quercetin-Containing Apple and Cherry Extracts Affects Blood Glucose Concentration, Hepatic Metabolism, and Gene Expression Patterns in Obese C57BL/6J High Fat-Fed Mice. J. Nutr. 2016, 146, 1001–1007. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.; Stavroullakis, A.T.; Ferreira, A.C.; Li, K.; Oliveira, T.; Nogueira-Filho, G.; Prakki, A. Extract of Acai-berry inhibits osteoclast differentiation and activity. Arch. Oral Biol. 2016, 68, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, A.; Saavedra, N.; Rudnicki, M.; Abdalla, D.S.; Salazar, L.A. ERK1/2 and HIF1α Are Involved in Antiangiogenic Effect of Polyphenols-Enriched Fraction from Chilean propolis. Evid. Based Complement. Altern. Med. 2015, 2015, 187575. [Google Scholar] [CrossRef] [PubMed]
- Benvenuto, M.; Mattera, R.; Taffera, G.; Giganti, M.G.; Lido, P.; Masuelli, L.; Modesti, A.; Bei, R. The Potential Protective Effects of Polyphenols in Asbestos-Mediated Inflammation and Carcinogenesis of Mesothelium. Nutrients 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Ambros, V. The functions of animal microRNAs. Nature 2004, 431, 350–355. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P.; Chen, C.Z. Micromanagers of gene expression: The potentially widespread influence of metazoan microRNAs. Nat. Rev. Genet. 2004, 5, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Stefani, G.; Slack, F.J. Small non-coding RNAs in animal development. Nat. Rev. Mol. Cell. Biol. 2008, 9, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Gandhi, V.; Plunkett, W.; Sampath, D.; Calin, G.A. miRNAs and their potential for use against cancer and other diseases. Future Oncol. 2007, 3, 521–537. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Costinean, S.; Croce, C.M. MicroRNAs, the immune system and rheumatic disease. Nat. Clin. Pract. Rheumatol. 2008, 4, 534–541. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.; Jung, S.; Keller, S.; Gregory, R.I.; Diederichs, S. Many roads to maturity: microRNA biogenesis pathways and their regulation. Nat. Cell. Biol. 2009, 11, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Tsang, J.; Zhu, J.; van Oudenaarden, A. MicroRNA-mediated feedback and feedforward loops are recurrent network motifs in mammals. Mol. Cell 2007, 26, 753–767. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, E.; Pivarcsi, A. Advances in microRNAs: Implications for immunity and inflammatory diseases. J. Cell. Mol. Med. 2009, 13, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Iborra, M.; Bernuzzi, F.; Invernizzi, P.; Danese, S. MicroRNAs in autoimmunity and inflammatory bowel disease: Crucial regulators in immune response. Autoimmun. Rev. 2012, 11, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Croce, C.M.; Michaille, J.-J. miR-155: On the crosstalk between inflammation and cancer. Int. Rev. Immunol. 2009, 28, 264–284. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Calin, G.A. Expression and function of micro-RNAs in immune cells during normal or disease state. Int. J. Med. Sci. 2008, 5, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Quinn, S.R.; O’Neill, L.A. A trio of microRNAs that control Toll-like receptor signalling. Int. Immunol. 2011, 23, 421–425. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Luo, Z.; Volinia, S.; Rassenti, L.Z.; Kipps, T.J.; Croce, C.M. The down-regulation of miR-125b in chronic lymphocytic leukemias leads to metabolic adaptation of cells to a transformed state. Blood 2012, 120, 2631–2638. [Google Scholar] [CrossRef] [PubMed]
- Kosaka, N.; Ochiya, T. Unraveling the Mystery of Cancer by Secretory microRNA: Horizontal microRNA Transfer between Living Cells. Front. Genet. 2012, 2, 97. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Adair, B.; Alder, H.; Limagne, E.; Taccioli, C.; Ferracin, M.; Delmas, D.; Latruffe, N.; Croce, C.M. Resveratrol decreases the levels of miR-155 by upregulating miR-663, a microRNA targeting JunB and JunD. Carcinogenesis 2010, 31, 1561–1566. [Google Scholar] [CrossRef] [PubMed]
- Yin, Q.; Wang, X.; McBride, J.; Fewell, C.; Flemington, E. B-cell receptor activation induces BIC/miR-155 expression through a conserved AP-1 element. J. Biol. Chem. 2008, 283, 2654–2662. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Croce, C.M. MicroRNAs play a central role in molecular dysfunctions linking inflammation with cancer. Immunol. Rev. 2013, 253, 167–184. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Chiabai, M.; Palmieri, D.; Brown, M.; Cui, R.; Fernandes, C.; Richmond, T.; Kim, T.; Sheetz, T.; Sun, H.L.; et al. Quaking and miR-155 interactions in inflammation and leukemogenesis. Oncotarget 2015, 6, 24599–24610. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Jun, M.; Ahn, M.R.; Kim, O.Y. Involvement of miR-Let7A in inflammatory response and cell survival/apoptosis regulated by resveratrol in THP-1 macrophage. Nutr. Res. Pract. 2016, 10, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Cimino, A.; Costinean, S.; Dumitru, C.D.; Adair, B.; Fabbri, M.; Alder, H.; Liu, C.G.; Calin, G.A.; et al. Modulation of miR-155 and miR-125b levels following lipopolysaccharide/TNF-alpha stimulation and their possible roles in regulating the response to endotoxin shock. J. Immunol. 2007, 179, 5082–5089. [Google Scholar] [CrossRef] [PubMed]
- Costinean, S.; Sandhu, S.K.; Pedersen, I.M.; Tili, E.; Trotta, R.; Perrotti, D.; Ciarlariello, D.; Neviani, P.; Harb, J.; Kauffman, L.R.; et al. Src homology 2 domain-containing inositol-5-phosphatase and CCAAT enhancer-binding protein beta are targeted by miR-155 in B cells of Emicro-MiR-155 transgenic mice. Blood 2009, 114, 1374–1382. [Google Scholar] [CrossRef] [PubMed]
- Androulidaki, A.; Iliopoulos, D.; Arranz, A.; Doxaki, C.; Schworer, S.; Zacharioudaki, V.; Margioris, A.N.; Tsichlis, P.N.; Tsatsanis, C. The kinase Akt1 controls macrophage response to lipopolysaccharide by regulating microRNAs. Immunity 2009, 31, 220–231. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Talcott, S.; Safe, S.; Mertens-Talcott, S.U. Cytotoxicity of pomegranate polyphenolics in breast cancer cells in vitro and vivo: Potential role of miRNA-27a and miRNA-155 in cell survival and inflammation. Breast Cancer Res. Treat. 2012, 136, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Wernicke, D.; Alder, H.; Costinean, S.; Volinia, S.; Croce, C.M. Mutator activity induced by microRNA-155 (miR-155) links inflammation and cancer. Proc. Natl. Acad. Sci. USA 2011, 108, 4908–4913. [Google Scholar] [CrossRef] [PubMed]
- Bala, S.; Petrasek, J.; Mundkur, S.; Catalano, D.; Levin, I.; Ward, J.; Alao, H.; Kodys, K.; Szabo, G. Circulating microRNAs in exosomes indicate hepatocyte injury and inflammation in alcoholic, drug-induced, and inflammatory liver diseases. Hepatology 2012, 56, 1946–1957. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, G.; Meng, C.; Guo, X.; Cheruku, P.S.; Shi, L.; Xu, H.; Li, H.; Wang, G.; Evans, A.R.; Safe, S.; et al. A novel regulator of macrophage activation: miR-223 in obesity-associated adipose tissue inflammation. Circulation 2012, 125, 2892–2903. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel-Duby, R.; Olson, E.N. The endothelial-specific microRNA miR-126 governs vascular integrity and angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Banerjee, N.; Barnes, R.C.; Pfent, C.M.; Talcott, S.T.; Dashwood, R.H.; Mertens-Talcott, S.U. Mango polyphenolics reduce inflammation in intestinal colitis-involvement of the miR-126/PI3K/AKT/mTOR axis in vitro and in vivo. Mol. Carcinog. 2016. [Google Scholar] [CrossRef] [PubMed]
- Ojwang, L.O.; Banerjee, N.; Noratto, G.D.; Angel-Morales, G.; Hachibamba, T.; Awika, J.M.; Mertens-Talcott, S.U. Polyphenolic extracts from cowpea (Vigna unguiculata) protect colonic myofibroblasts (CCD18Co cells) from lipopolysaccharide (LPS)-induced inflammation—Modulation of microRNA 126. Food. Funct. 2015, 6, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, N.; Kim, H.; Talcott, S.; Mertens-Talcott, S. Pomegranate polyphenolics suppressed azoxymethane-induced colorectal aberrant crypt foci and inflammation: Possible role of miR-126/VCAM-1 and miR-126/PI3K/AKT/mTOR. Carcinogenesis 2013, 34, 2814–2822. [Google Scholar] [CrossRef] [PubMed]
- Kasinski, A.L.; Slack, F.J. miRNA-34 prevents cancer initiation and progression in a therapeutically resistant K-ras and p53-induced mouse model of lung adenocarcinoma. Cancer Res. 2012, 72, 5576–5587. [Google Scholar] [CrossRef] [PubMed]
- Misso, G.; di Martino, M.T.; de Rosa, G.; Farooqi, A.A.; Lombardi, A.; Campani, V.; Zarone, M.R.; Gullà, A.; Tagliaferri, P.; Tassone, P.; et al. Mir-34: A new weapon against cancer? Mol. Ther. Nucleic Acids 2014, 3, e194. [Google Scholar] [CrossRef] [PubMed]
- Bader, A.G. miR-34—A microRNA replacement therapy is headed to the clinic. Front. Genet. 2012, 3, 120. [Google Scholar] [CrossRef] [PubMed]
- Avtanski, D.B.; Nagalingam, A.; Kuppusamy, P.; Bonner, M.Y.; Arbiser, J.L.; Saxena, N.K.; Sharma, D. Honokiol abrogates leptin-induced tumor progression by inhibiting Wnt1-MTA1-β-catenin signaling axis in a microRNA-34a dependent manner. Oncotarget 2015, 6, 16396–16410. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Yi, H.; Tan, J.; Wu, Y.; Liu, G.; Qiu, Z. Anti-proliferative effects of polyphenols from pomegranate rind (Punica granatum, L.) on EJ bladder cancer cells via regulation of p53/miR-34a axis. Phytother. Res. 2015, 29, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Sit, A.; Feinberg, M.W. Role of miR-181 family in regulating vascular inflammation and immunity. Trends Cardiovasc. Med. 2014, 24, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Kronski, E.; Fiori, M.E.; Barbieri, O.; Astigiano, S.; Mirisola, V.; Killian, P.H.; Bruno, A.; Pagani, A.; Rovera, F.; Pfeffer, U.; et al. miR181b is induced by the chemopreventive polyphenol curcumin and inhibits breast cancer metastasis via down-regulation of the inflammatory cytokines CXCL1 and -2. Mol. Oncol. 2014, 8, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Dahmke, I.N.; Backes, C.; Rudzitis-Auth, J.; Laschke, M.W.; Leidinger, P.; Menger, M.D.; Meese, E.; Mahlknecht, U. Curcumin intake affects miRNA signature in murine melanoma with mmu-miR-205–5p most significantly altered. PLoS ONE 2013, 8, e81122. [Google Scholar] [CrossRef] [PubMed]
- Ohno, M.; Shibata, C.; Kishikawa, T.; Yoshikawa, T.; Takata, A.; Kojima, K.; Akanuma, M.; Kang, Y.J.; Yoshida, H.; Otsuka, M.; et al. The flavonoid apigenin improves glucose tolerance through inhibition of microRNA maturation in miRNA103 transgenic mice. Sci. Rep. 2013, 3, 2553. [Google Scholar] [CrossRef] [PubMed]
- Ivan, M.; Harris, A.L.; Martelli, F.; Kulshreshtha, R. Hypoxia response and microRNAs: No longer two separate worlds. J. Cell. Mol. Med. 2008, 12, 1426–1431. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Bian, S.; Yang, C.S. Green tea polyphenol EGCG suppresses lung cancer cell growth through upregulating miR-210 expression caused by stabilizing HIF-1α. Carcinogenesis 2011, 32, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Perez-Polo, J.R.; Qian, J.; Birnbaum, Y. The role of microRNA in modulating myocardial ischemia-reperfusion injury. Physiol. Genom. 2011, 43, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Jia, Z.; Li, A.; Jenkins, G.; Yang, X.; Hu, J.; Guo, W. Resveratrol repressed viability of U251 cells by miR-21 inhibiting of NF-κB pathway. Mol. Cell. Biochem. 2013, 382, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Sayed, D.; He, M.; Hong, C.; Gao, S.; Rane, S.; Yang, Z.; Abdellatif, M. MicroRNA-21 is a downstream effector of AKT that mediates its antiapoptotic effects via suppression of Fas ligand. J. Biol. Chem. 2010, 285, 20281–20290. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; Ai, W.; Banik, N.L.; Ray, S.K. Overexpression of miR-7–1 increases efficacy of green tea polyphenols for induction of apoptosis in human malignant neuroblastoma SH-SY5Y and SK-N-DZ cells. Neurochem. Res. 2013, 38, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Tili, E.; Michaille, J.-J.; Liu, C.G.; Alder, H.; Taccioli, C.; Volinia, S.; Calin, G.A.; Croce, C.M. GAM/ZFp/ZNF512B is central to a gene sensor circuitry involving cell-cycle regulators, TGF{beta} effectors, Drosha and microRNAs with opposite oncogenic potentials. Nucleic Acids Res. 2010, 38, 7673–7688. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, J.X.; Yang, C.S.; Yang, M.Q.; Deng, Y.; Wang, H. Gene regulation mediated by microRNAs in response to green tea polyphenol EGCG in mouse lung cancer. BMC Genom. 2014, 15 (Suppl. 11). [Google Scholar] [CrossRef] [PubMed]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and evolutionary forces shaping microbial diversity in the human intestine. Cell 2006, 124, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, J.R.; Adams, D.H.; Fava, F.; Hermes, G.D.; Hirschfield, G.M.; Hold, G.; Quraishi, M.N.; Kinross, J.; Smidt, H.; Tuohy, K.M.; et al. The gut microbiota and host health: A new clinical frontier. Gut 2016, 65, 330–339. [Google Scholar] [CrossRef] [PubMed]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Neurath, M.F.; Wirtz, S. The Intestinal Microbiota in Inflammatory Bowel Disease. ILAR J. 2015, 56, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef] [PubMed]
- Morais, C.A.; de Rosso, V.V.; Estadella, D.; Pisani, L.P. Anthocyanins as inflammatory modulators and the role of the gut microbiota. J. Nutr. Biochem. 2016, 33, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Sela, D.A.; Xiao, J.; Boyacioglu, D.; Chen, F.; Capanoglu, E. The Reciprocal Interactions between Polyphenols and Gut Microbiota and Effects on Bioaccessibility. Nutrients 2016, 8, 78. [Google Scholar] [CrossRef] [PubMed]
- Mosele, J.I.; Macià, A.; Motilva, M.J. Metabolic and Microbial Modulation of the Large Intestine Ecosystem by Non-Absorbed Diet Phenolic Compounds: A Review. Molecules 2015, 20, 17429–17468. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A survey of modulation of gut microbiota by dietary polyphenols. Biomed. Res. Int. 2015, 2015, 850902. [Google Scholar] [CrossRef] [PubMed]
- Dueñas, M.; Cueva, C.; Muñoz-González, I.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. Studies on Modulation of Gut Microbiota by Wine Polyphenols: From Isolated Cultures to Omic Approaches. Antioxidants (Basel) 2015, 4, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Etxeberria, U.; Hijona, E.; Aguirre, L.; Milagro, F.I.; Bujanda, L.; Rimando, A.M.; Martínez, J.A.; Portillo, M.P. Pterostilbene-induced changes in gut microbiota composition in relation to obesity. Mol. Nutr. Food Res. 2016. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xu, J.; Rottinghaus, G.E.; Simonyi, A.; Lubahn, D.; Sun, G.Y.; Sun, A.Y. Resveratrol protects against global cerebral ischemic injury in gerbils. Brain Res. 2002, 958, 439–447. [Google Scholar] [CrossRef]
- Singleton, R.H.; Yan, H.Q.; Fellows-Mayle, W.; Dixon, C.E. Resveratrol attenuates behavioral impairments and reduces cortical and hippocampal loss in a rat controlled cortical impact model of traumatic brain injury. J. Neurotrauma 2010, 27, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Karthick, C.; Periyasamy, S.; Jayachandran, K.S.; Anusuyadevi, M. Intrahippocampal Administration of Ibotenic Acid Induced Cholinergic Dysfunction via NR2A/NR2B Expression: Implications of Resveratrol against Alzheimer Disease Pathophysiology. Front. Mol. Neurosci. 2016, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.I.; Shin, J.A.; Cho, S.; Kim, H.W.; Lee, J.Y.; Kang, J.L.; Park, E.M. Resveratrol attenuates peripheral and brain inflammation and reduces ischemic brain injury in aged female mice. Neurobiol. Aging 2016, 44, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Brasnyó, P.; Molnár, G.A.; Mohás, M.; Markó, L.; Laczy, B.; Cseh, J.; Mikolás, E.; Szijártó, I.A.; Mérei, A.; Halmai, R.; et al. Resveratrol improves insulin sensitivity, reduces oxidative stress and activates the Akt pathway in type 2 diabetic patients. Br. J. Nutr. 2011, 106, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Magyar, K.; Halmosi, R.; Palfi, A.; Feher, G.; Czopf, L.; Fulop, A.; Battyany, I.; Sumegi, B.; Toth, K.; Szabados, E. Cardioprotection by resveratrol: A human clinical trial in patients with stable coronary artery disease. Clin. Hemorheol. Microcirc. 2012, 50, 179–187. [Google Scholar] [PubMed]
- Tomé-Carneiro, J.; Larrosa, M.; González-Sarrías, A.; Tomás-Barberán, F.; García-Conesa, M.T.; Espín, J.C. Resveratrol and clinical trials: The crossroad from in vitro studies to human evidence. Curr. Pharm. Des. 2013, 19, 6064–6093. [Google Scholar]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; García-Almagro, F.J.; Avilés-Plaza, F.; Parra, S.; Yáñez-Gascón, M.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; et al. Consumption of a grape extract supplement containing resveratrol decreases oxidized LDL and ApoB in patients undergoing primary prevention of cardiovascular disease: A triple-blind, 6-month follow-up, placebo-controlled, randomized trial. Mol. Nutr. Food Res. 2012, 56, 810–821. [Google Scholar]
- Tomé-Carneiro, J.; Gonzálvez, M.; Larrosa, M.; Yáñez-Gascón, M.J.; García-Almagro, F.J.; Ruiz-Ros, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Espín, J.C. One-year consumption of a grape nutraceutical containing resveratrol improves the inflammatory and fibrinolytic status of patients in primary prevention of cardiovascular disease. Am. J. Cardiol. 2012, 110, 356–363. [Google Scholar] [CrossRef] [PubMed]
- Alfaras, I.; Pérez, M.; Juan, M.E.; Merino, G.; Prieto, J.G.; Planas, J.M.; Alvarez, A.I. Involvement of breast cancer resistance protein (BCRP1/ABCG2) in the bioavailability and tissue distribution of trans-resveratrol in knockout mice. J. Agric. Food Chem. 2010, 58, 4523–4528. [Google Scholar] [CrossRef] [PubMed]
- Van de Wetering, K.; Burkon, A.; Feddema, W.; Bot, A.; de Jonge, H.; Somoza, V.; Borst, P. Intestinal breast cancer resistance protein (BCRP)/Bcrp1 and multidrug resistance protein 3 (MRP3)/Mrp3 are involved in the pharmacokinetics of resveratrol. Mol. Pharmacol. 2009, 75, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Juan, M.E.; González-Pons, E.; Planas, J.M. Multidrug resistance proteins restrain the intestinal absorption of trans-resveratrol in rats. J. Nutr. 2010, 140, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Azorín-Ortuño, M.; Yáñez-Gascón, M.J.; Vallejo, F.; Pallarés, F.J.; Larrosa, M.; Lucas, R.; Morales, J.C.; Tomás-Barberán, F.A.; García-Conesa, M.T.; Espín, J.C. Metabolites and tissue distribution of resveratrol in the pig. Mol. Nutr. Food Res. 2011, 55, 1154–1168. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.; Steward, W.P.; Gescher, A.J.; Brown, K. Resveratrol in human cancer chemoprevention—Choosing the ‘right’ dose. Mol. Nutr. Food Res. 2012, 56, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.H.; Howe, P.R.; Buckley, J.D.; Coates, A.M.; Kunz, I.; Berry, N.M. Acute resveratrol supplementation improves flow-mediated dilatation in overweight/obese individuals with mildly elevated blood pressure. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 851–856. [Google Scholar] [CrossRef] [PubMed]
- Lesjak, M.; Hoque, R.; Balesaria, S.; Skinner, V.; Debnam, E.S.; Srai, S.K.; Sharp, P.A. Quercetin inhibits intestinal iron absorption and ferroportin transporter expression in vivo and in vitro. PLoS ONE 2014, 9, e102900. [Google Scholar] [CrossRef] [PubMed]
- Baselga-Escudero, L.; Blade, C.; Ribas-Latre, A.; Casanova, E.; Suárez, M.; Torres, J.L.; Salvadó, M.J.; Arola, L.; Arola-Arnal, A. Resveratrol and EGCG bind directly and distinctively to miR-33a and miR-122 and modulate divergently their levels in hepatic cells. Nucleic Acids Res. 2014, 42, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Baselga-Escudero, L.; Bladé, C.; Ribas-Latre, A.; Casanova, E.; Salvadó, M.J.; Arola, L.; Arola-Arnal, A. Grape seed proanthocyanidins repress the hepatic lipid regulators miR-33 and miR-122 in rats. Mol. Nutr. Food Res. 2012, 56, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.; Walters, M.A. Chemistry: Chemical con artists foil drug discovery. Nature 2014, 513, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.B. Feeling Nature’s PAINS: Natural Products, Natural Product Drugs, and Pan Assay Interference Compounds (PAINS). J. Nat. Prod. 2016, 79, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Bisson, J.; McAlpine, J.B.; Friesen, J.B.; Chen, S.N.; Graham, J.; Pauli, G.F. Can Invalid Bioactives Undermine Natural Product-Based Drug Discovery? J. Med. Chem. 2016, 59, 1671–1690. [Google Scholar] [CrossRef] [PubMed]
- Chango, A.; Pogribny, I.P. Considering maternal dietary modulators for epigenetic regulation and programming of the fetal epigenome. Nutrients 2015, 7, 2748–2470. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Wen, Q.; Jiang, J.; Li, H.L.; Tan, Y.F.; Li, Y.H.; Zeng, N.K. Could the gut microbiota reconcile the oral bioavailability conundrum of traditional herbs? J. Ethnopharmacol. 2016, 179, 253–264. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Molecules | Effects on microRNAs | Cells/Animals | Main Target Transcripts or Pathways | References |
|---|---|---|---|---|
| Resveratrol | miR-663 up | THP-1 cells | JunD and JunD; Impairs miR-155 up-regulation | [60] |
| Resveratrol | miR-663 up | SW480 cells | TGF-β1 transcripts and SMAD2/SMAD3/SMAD4 pathway | [19] |
| Resveratrol | miR-Let7A up | THP-1 cells | TNF, IL-6, IL-10, BDNF and ASK1 pathways | [64] |
| Resveratrol | miR-21 down | U251 brain tumor cells | NF-κB pathway | [88] |
| Pomegranate polyphenols | miR-155 down | MCF-10F and MCF-12F cells | PI3K/AKT and NF-κB pathways | [68] |
| Chilean propolis polyphenols | miR-20b, miR-106a, and miR-181a up | Atherosclerotic plaques of LDL receptor knockout mice | HIF-1α, ERK1/2 and VEGF pathways | [35] |
| Mango extracts | miR-126 up | CCD-18Co cells | PI3K(p85β) transcripts and PI3K/AKP/mTOR pathway | [72] |
| Cowpea (Vigna unguiculata) polyphenols | miR-126 up | CCD-18Co cells | VCAM-1 transcripts; TNF, IL8 and NF-κB pathways | [74] |
| Pomegranate juice polyphenols | miR-126 up | Colon mucosa of rats injected with azoxymethane (AOM) subcutaneously | PI3K(p85β) and VCAM-1 transcripts; iNOS, COX-2 and NF-κB pathways | [75] |
| Honokiol | miR-34a up | Human breast cancer cell lines | Wnt1 transcripts; Wnt1-MTA1-β-catenin pathway | [79] |
| Pomegranate rind polyphenols | miR-34a up | EJ bladder cancer cells | c-Myc and CD44 transcripts | [80] |
| Curcumin | miR-181b up | MDA-MB-231 human breast cancer cells | CXCL1 and CXCL2 transcripts | [82] |
| Curcumin | miR-205-5p and other microRNAs up | Murine melanoma | Proliferation and metastatic pathways; O-glycan biosynthesis; Endoplasmic reticulum protein maturation and/or processing | [83] |
| Apigenin | miR-122 down | Human Huh7 cells | miR-122 positively regulates hepatitis C replication in enhancing viral translation | [15] |
| Apigenin | miR-103 down | (miR103)-overexpressing transgenic mic presenting with glucose intolerance | TRBP phosphorylation; Improvement of pathogenic status | [84] |
| EGCG | miR-210 up | Mouse lung adenocarcinoma cell line CL13 | Pathways implicated in cell-proliferation and anchorage-independent growth | [86] |
| EGCG or EGC | miR-7-1 up, and miR-92, miR-93, and miR-106b down | SH-SY5Y and SK-N-DZ neuroblastoma cell lines | Apoptosis pathway | [90] |
| EGCG | 12 microRNAs up, and 9 microRNAs down | A/J mice lung adenoma | AKT, MAP kinases and NF-κB and cell cycle regulation pathways | [92] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tili, E.; Michaille, J.-J. Promiscuous Effects of Some Phenolic Natural Products on Inflammation at Least in Part Arise from Their Ability to Modulate the Expression of Global Regulators, Namely microRNAs. Molecules 2016, 21, 1263. https://doi.org/10.3390/molecules21091263
Tili E, Michaille J-J. Promiscuous Effects of Some Phenolic Natural Products on Inflammation at Least in Part Arise from Their Ability to Modulate the Expression of Global Regulators, Namely microRNAs. Molecules. 2016; 21(9):1263. https://doi.org/10.3390/molecules21091263
Chicago/Turabian StyleTili, Esmerina, and Jean-Jacques Michaille. 2016. "Promiscuous Effects of Some Phenolic Natural Products on Inflammation at Least in Part Arise from Their Ability to Modulate the Expression of Global Regulators, Namely microRNAs" Molecules 21, no. 9: 1263. https://doi.org/10.3390/molecules21091263