Effects of Ursolic Acid Derivatives on Caco-2 Cells and Their Alleviating Role in Streptozocin-Induced Type 2 Diabetic Rats

Abstract
:
1. Introduction
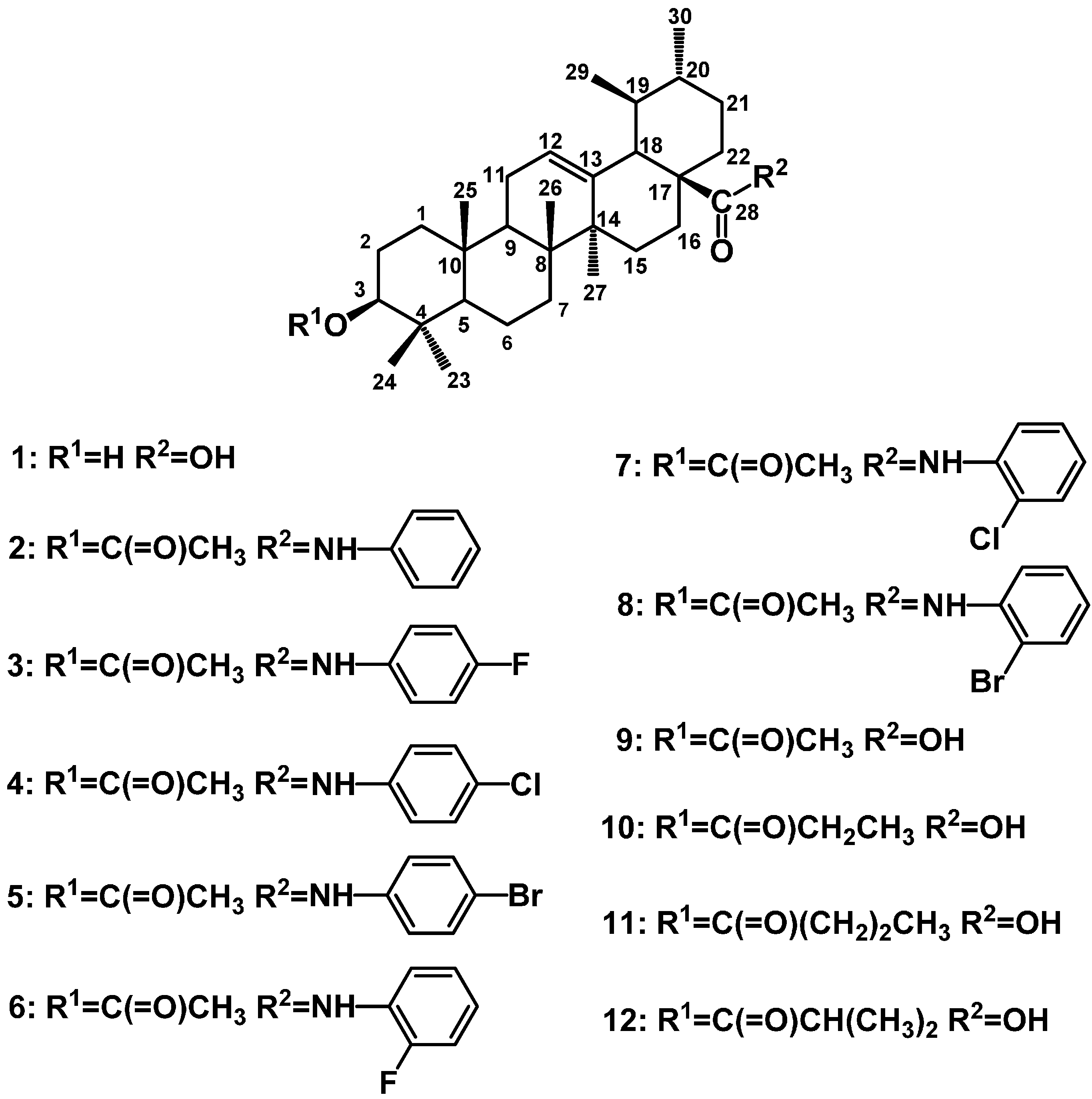
2. Results and Discussion
2.1. Cell Toxicity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Inhibition Rate % | Compound | Inhibition Rate % |
|---|---|---|---|
| 1 | 9.88 ± 6.51 | 8 | 9.86 ± 2.68 |
| 2 | 5.02 ± 2.38 | 9 | 4.40 ± 3.09 |
| 3 | 8.65 ± 3.49 | 10 | 7.84 ± 4.15 |
| 4 | 8.53 ± 5.69 | 11 | 2.29 ± 6.08 |
| 5 | 8.72 ± 1.18 | 12 | 7.67 ± 5.36 |
| 6 | 13.88 ± 4.52 | Phlorizin | 7.70 ± 3.91 |
| 7 | 8.14 ± 3.81 | Phloretin | 4.70 ± 2.59 |
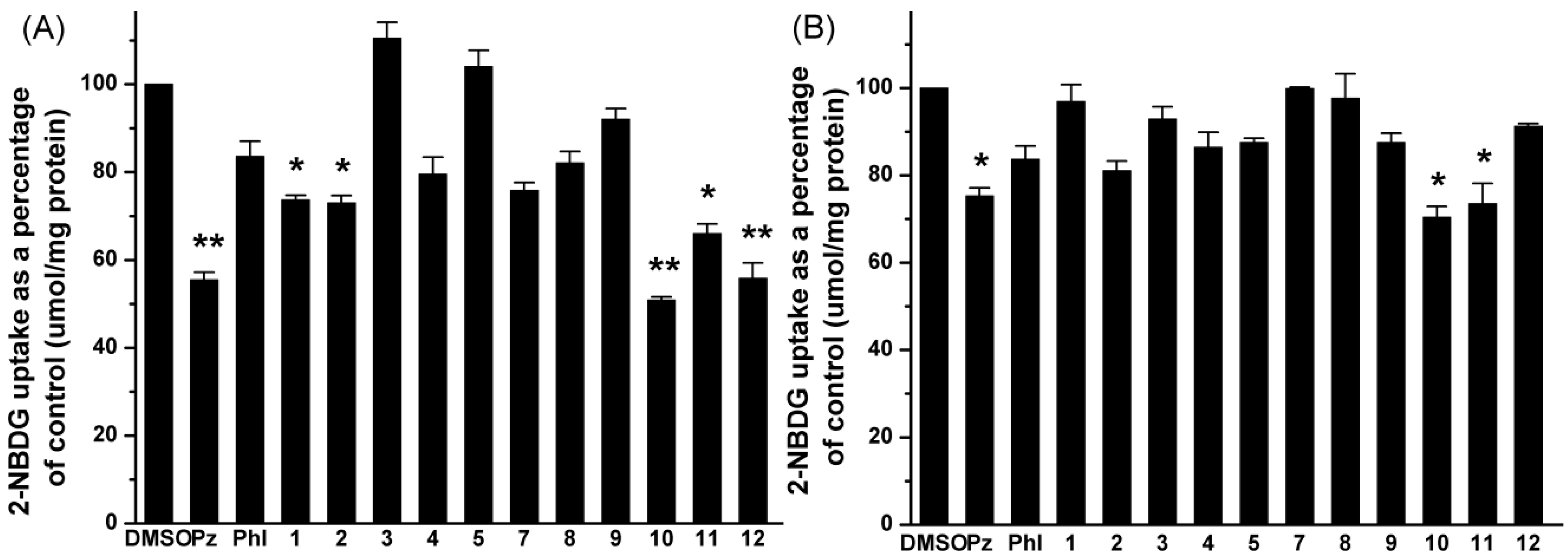
2.2. Effects of UA and Its Derivatives on Glucose Uptake by Caco-2 Cells
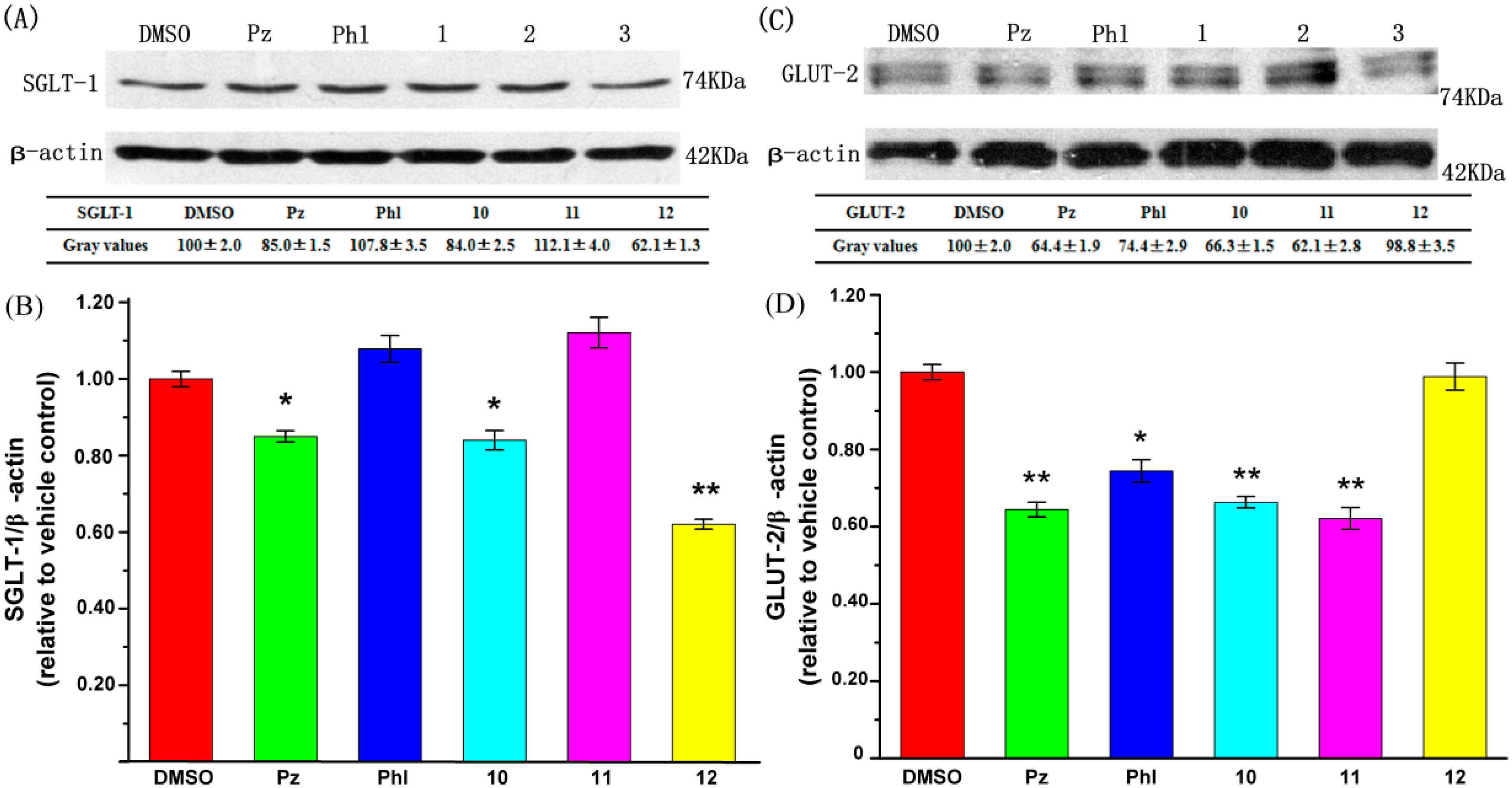
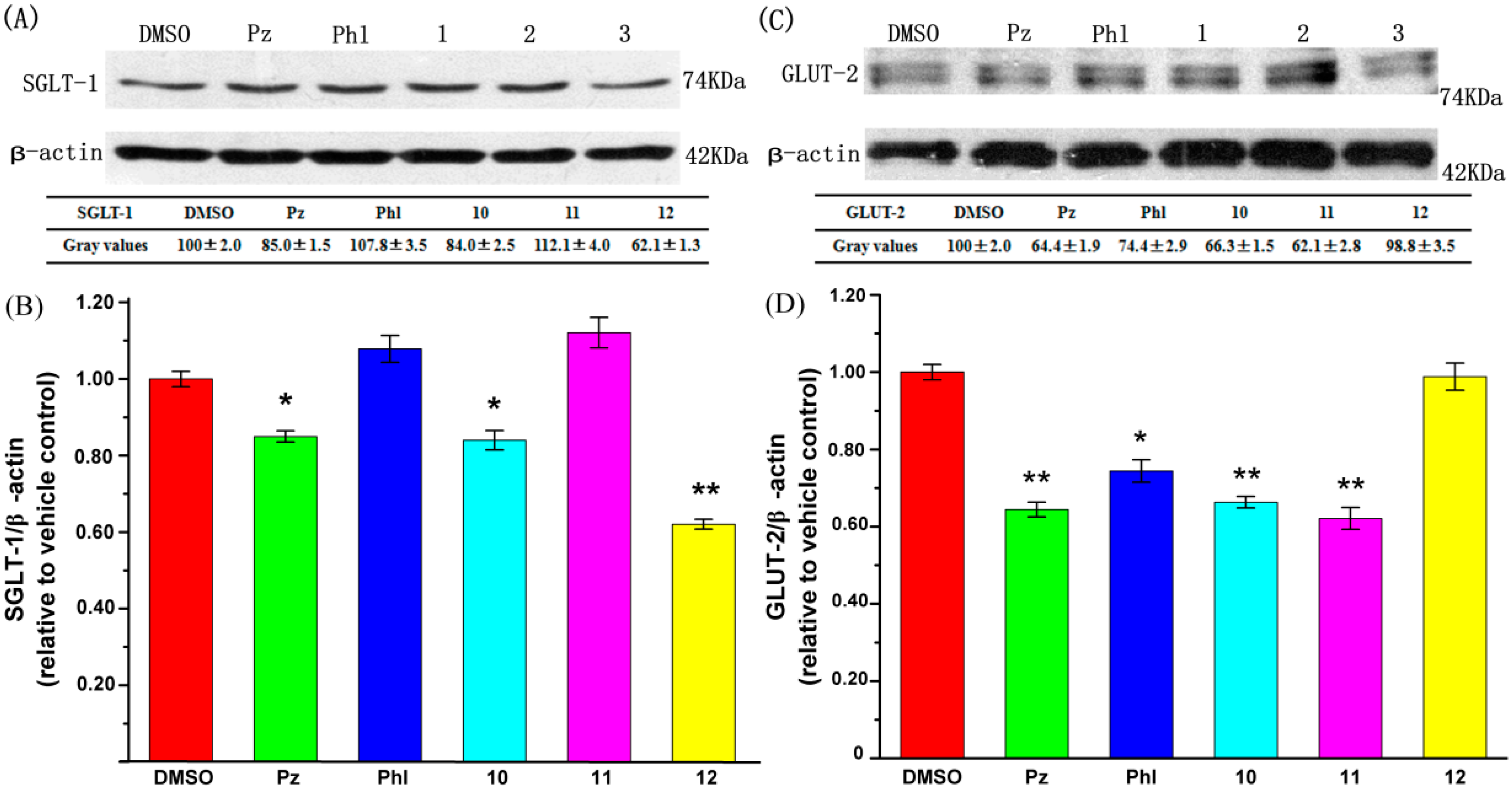
2.3. Effect of UA and Its Derivatives on Glucose Transporter Protein Expression
2.4. Acute Oral Toxicity Study of UA and Its Derivatives
2.5. Effects of UA and Its Derivatives on Fasting Blood Glucose Levels
| Group | Fasting Blood Glucose Level (mg/dL) | ||||
|---|---|---|---|---|---|
| Week 0 | Week 1 | Week 2 | Week 3 | Week 4 | |
| Normal control | 73.80 ± 4.50 | 75.60 ± 7.20 | 72.00 ± 6.30 | 72.00 ± 5.40 | 81.00 ± 2.16 |
| Diabetic control | 320.40 ± 20.52 | 401.40 ± 22.50 | 428.40 ± 50.40 | 433.80 ± 39.06 | 446.40 ± 31.68 |
| Glibenclamide (10 mg/kg) | 338.40 ± 16.38 | 271.80 ± 18.36 * | 183.60 ± 11.70 * | 216.00 ± 9.72 * | 203.40 ± 13.50 * |
| Compound 1 (100 mg/kg) | 342.00 ± 19.08 | 405.00 ± 17.46 | 392.40 ± 13.32 | 379.80 ± 19.44 | 396.00 ± 16.92 |
| Compound 10 (100 mg/kg) | 320.40 ± 12.96 | 270.00 ± 16.74 * | 286.20 ± 18.90 | 234.00 ± 14.58 * | 225.00 ± 11.70 * |
| Compound 11 (100 mg/kg) | 318.60 ± 17.64 | 365.40 ± 20.34 | 352.80 ± 27.36 | 324.00 ± 23.04 | 351.00 ± 26.64 |
| Compound 12 (100 mg/kg) | 325.80 ± 14.58 | 392.40 ± 36.54 | 379.80 ± 30.60 | 408.60 ± 20.52 | 432.00 ± 28.26 |
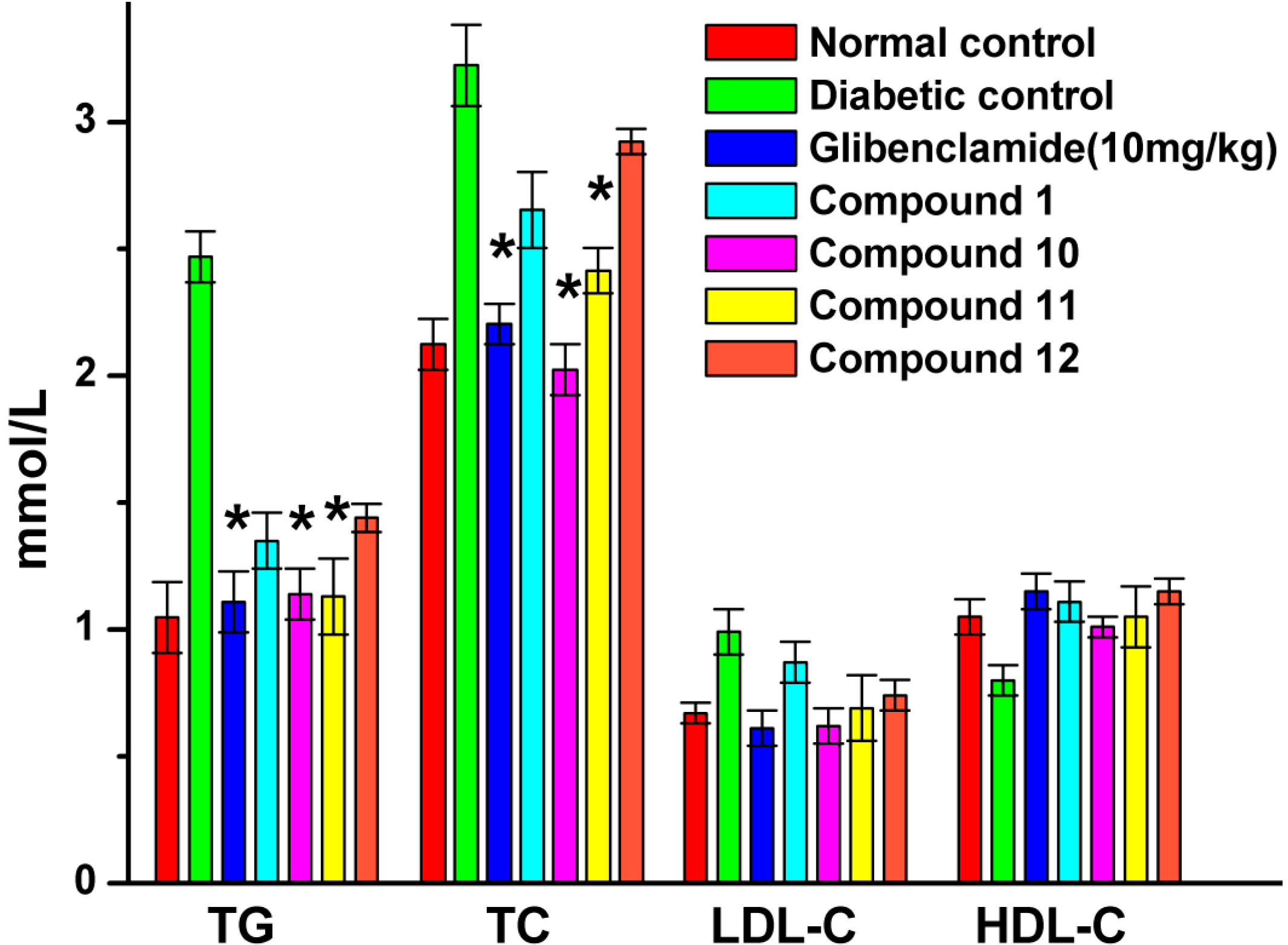
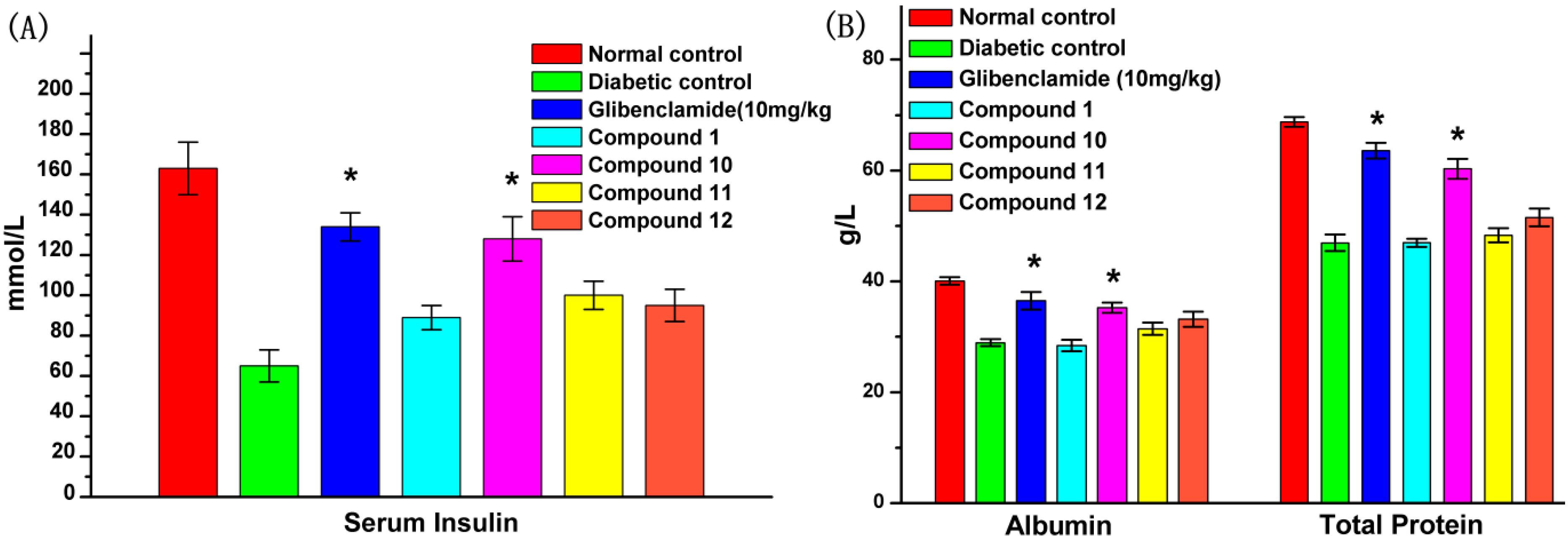
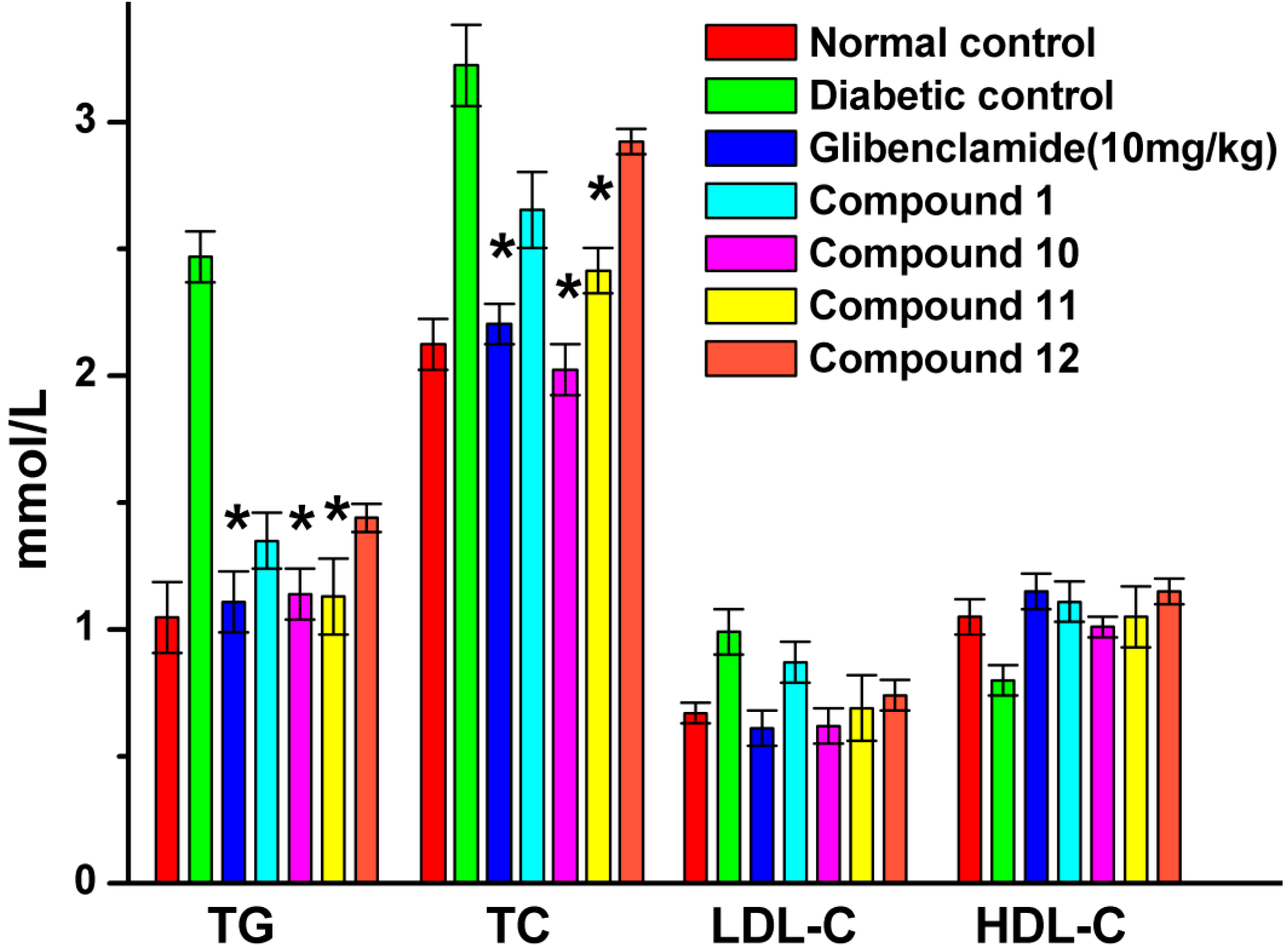
2.6. Effect of UA and Its Derivatives on Serum Biochemical Parameters
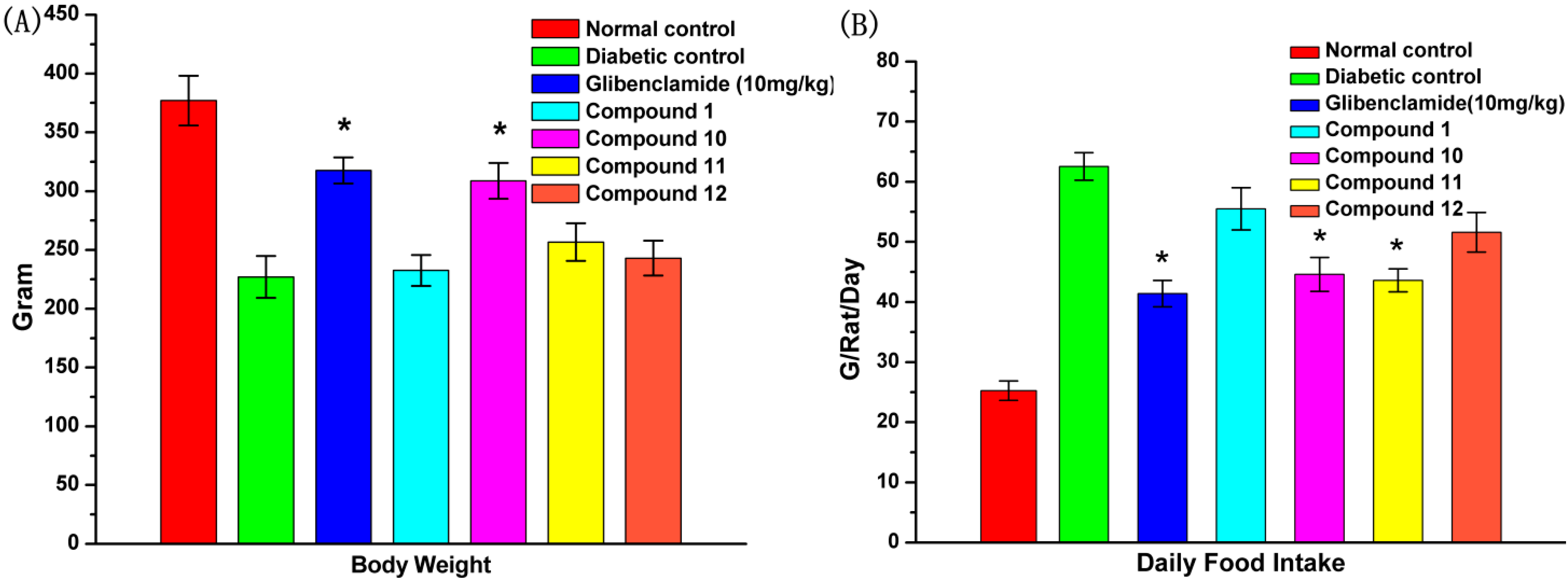
2.7. Effect of UA and Its Derivatives on Body Weight and Food Intake
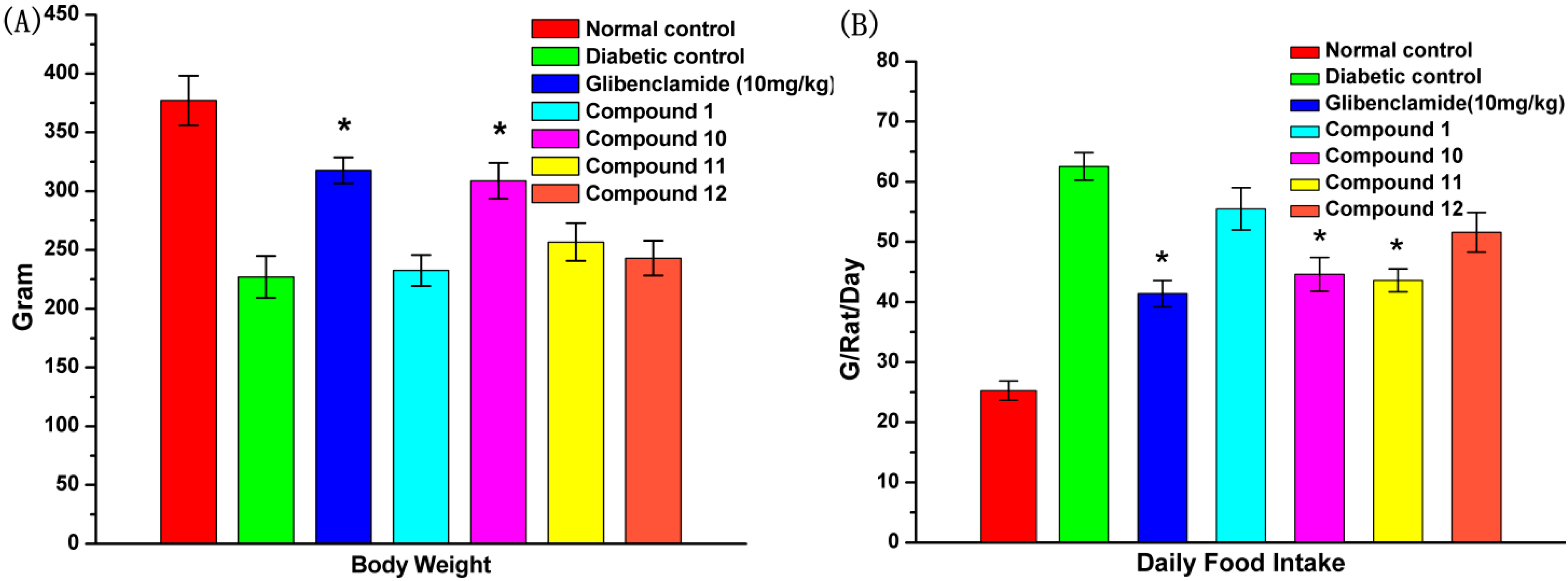
2.8. Effect of UA and Its Derivatives on Hyperlipidemia
2.9. Effect of UA and Its Derivatives on Oxidative Stress
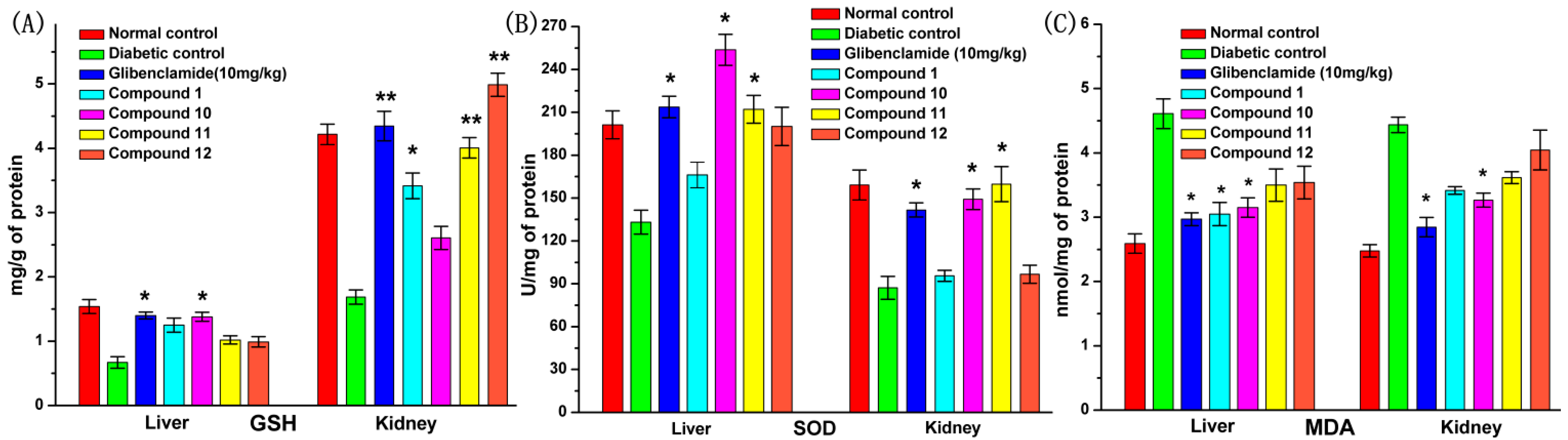
2.10. Discussion
3. Experimental Section
3.1. Preparation of UA and Its Derivatives
3.2. In vitro Cell Culture
3.3. Cytotoxicity Assay
3.4. Glucose Uptake by Caco-2 Cell Monolayers
3.5. Western Blot Analysis
3.6. In Vivo Experimental Animals
3.7. Oral Acute Toxicity Studies
3.8. Induction of Diabetes
3.9. Experimental Procedure
- Group I: Normal rats treated with 0.5% CMC-Na (2 mL/200 g body weight).
- Group II: Diabetic rats treated with 0.5% CMC-Na.
- Group III: Diabetic rats treated with 10 mg/kg of glibenclamide.
- Group IV: Diabetic rats treated with 100 mg/kg of compound 1 (UA).
- Group V: Diabetic rats treated with 100 mg/kg of compound 10.
- Group VI: Diabetic rats treated with 100 mg/kg of compound 11.
- Group VII: Diabetic rats treated with 100 mg/kg of compound 12.
3.10. Biochemical Analysis
3.11. Assessment of Oxidative Stress Markers
3.12. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| 2-NBDG | 2-[N-(7-nitrobenz-2-oxa-1, 3-diazol-4-yl) amino]-2-deoxyglucose |
| SGLT-1 | Na+-dependent glucose transporter |
| GLUT-2 | Na+-independent glucose transporter |
| DM | diabetes mellitus |
| UA | ursolic acid |
| STZ | streptozotocin |
| ROS | reactive oxygen species |
| GSH | glutathione |
| SOD | superoxide dismutase |
| MDA | malondialdehyde |
| TG | triglyceride |
| TC | total cholesterol |
| LDL-C | low density lipoprotein cholesterol |
| HDL-C | high density lipoprotein cholesterol |
Conflicts of Interest
References
- WHO. “Fact Sheet # 312” to Be Found under. Available online: http://www.who.int/mediacentre/factsheets/fs312/en/ (accessed on 15 December 2013).
- Tuomilehto, J.; Lindström, J.; Eriksson, J.G.; Valle, T.T.; Hämäläinen, H.; Ilanne-Parikka, P.; Keinänen-Kiukaanniemi, S.; Laakso, M.; Louheranta, A.; Rastas, M. Prevention of type 2 diabetes mellitus by changes in lifestyle among subjects with impaired glucose tolerance. N. Engl. J. Med. 2001, 344, 1343–1350. [Google Scholar] [CrossRef]
- Zimmet, P.; Alberti, K.G.; Shaw, J. Global and societal implications of the diabetes epidemic. Nature 2001, 414, 782–787. [Google Scholar] [CrossRef]
- Lopez-Candales, A. Metabolic syndrome X: A comprehensive review of the pathophysiology and recommended therapy. J. Med. 2001, 32, 283–300. [Google Scholar]
- Hays, N.P.; Galassetti, P.R.; Coker, R.H. Prevention and treatment of type 2 diabetes: Current role of lifestyle, natural product, and pharmacological interventions. Pharmacol. Ther. 2008, 118, 181–191. [Google Scholar] [CrossRef]
- Hung, H.Y.; Qian, K.; Morris-Natschke, S.L.; Hsu, C.S.; Lee, K.H. Recent discovery of plant-derived anti-diabetic natural products. Nat. Prod. Rep. 2012, 29, 580–606. [Google Scholar] [CrossRef]
- Jia, W.; Gao, W.; Tang, L. Antidiabetic herbal drugs officially approved in China. Phytother. Res. 2003, 17, 1127–1134. [Google Scholar] [CrossRef]
- Roglic, G.; Unwin, N.; Bennett, P.H.; Mathers, C.; Tuomilehto, J.; Nag, S.; Connolly, V.; King, H. The burden of mortality attributable to diabetes: Realistic estimates for the year 2000. Diabetes Care 2005, 28, 2130–2135. [Google Scholar] [CrossRef]
- Harmand, P.O.; Duval, R.; Delage, C.; Simon, A. Ursolic acid induces apoptosis through mitochondrial intrinsic pathway and caspase-3 activation in M4Beu melanoma cells. Int. J. Cancer 2005, 114, 1–11. [Google Scholar] [CrossRef]
- Kim, D.K.; Baek, J.H.; Kang, C.M.; Yoo, M.; Sung, J.W.; Kim, D.K.; Chung, H.Y.; Kim, N.D.; Choi, Y.H.; Lee, S.H. Apoptotic activity of ursolic acid may correlate with the inhibition of initiation of DNA replication. Int. J. Cancer 2000, 87, 629–636. [Google Scholar]
- Baricevic, D.; Sosa, S.; Della, L.R.; Tubaro, A.; Simonovska, B.; Krasna, A.; Zupancic, A. Topical anti-inflammatory activity of Salvia officinalis L. leaves: The relevance of ursolic acid. J. Ethnopharmacol. 2001, 75, 125–132. [Google Scholar] [CrossRef]
- Kazmi, I.; Rahman, M.; Afzal, M.; Gupta, G.; Saleem, S.; Afzal, O.; Shaharyar, M.A.; Nautiyal, U.; Ahmed, S.; Anwar, F. Anti-diabetic potential of ursolic acid stearoyl glucoside: A new triterpenic gycosidic ester from Lantana camara. Fitoterapia 2012, 83, 142–146. [Google Scholar] [CrossRef]
- Ullevig, S.L.; Zhao, Q.; Zamora, D.; Asmis, R. Ursolic acid protects diabetic mice against monocyte dysfunction and accelerated atherosclerosis. Atherosclerosis 2011, 219, 409–416. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, J.S.; Zhang, X.; Wu, Y.J.; Huang, K.; Zheng, L. Ursolic acid inhibits early lesions of diabetic nephropathy. Int. J. Mol. Med. 2010, 26, 565–570. [Google Scholar]
- Zou, C.; Wang, Y.; Shen, Z. 2-NBDG as a fluorescent indicator for direct glucose uptake measurement. J. Biochem. Biophys. Methods 2005, 64, 207–215. [Google Scholar] [CrossRef]
- Yamada, K.; Saito, M.; Matsuoka, H.; Inagaki, N. A real-time method of imaging glucose uptake in single, living mammalian cells. Nat. Protoc. 2007, 2, 753–762. [Google Scholar] [CrossRef]
- Hassanein, M.; Weidow, B.; Koehler, E.; Bakane, N.; Garbett, S.; Shyr, Y.; Quaranta, V. Development of high-throughput quantitative assays for glucose uptake in cancer cell lines. Mol. Imaging Biol. 2011, 13, 840–852. [Google Scholar] [CrossRef]
- Nistor, B.L.; Martineau, L.C.; Benhaddou-Andaloussi, A.; Arnason, J.T.; Levy, E.; Haddad, P.S. Inhibition of intestinal glucose absorption by anti-diabetic medicinal plants derived from the James Bay Cree traditional pharmacopeia. J. Ethnopharmacol. 2010, 132, 473–482. [Google Scholar] [CrossRef]
- Kim, H.K.; Baek, S.S.; Cho, H.Y. Inhibitory effect of pomegranate on intestinal sodium dependent glucose uptake. Am. J. Chin. Med. 2011, 39, 1015–1027. [Google Scholar] [CrossRef]
- Haidara, M.A.; Yassin, H.Z.; Rateb, M.; Ammar, H.; Zorkani, M.A. Role of oxidative stress in development of cardiovascular complications in diabetes mellitus. Curr. Vasc. Pharmacol. 2006, 4, 215–227. [Google Scholar] [CrossRef]
- Pitozzi, V.; Giovannelli, L.; Bardini, G.; Rotella, C.M.; Dolara, P. Oxidative DNA damage in peripheral blood cells in type 2 diabetes mellitus: Higher vulnerability of polymorphonuclear leukocytes. Mutat. Res. 2003, 529, 129–133. [Google Scholar] [CrossRef]
- Evans, J.L. Antioxidants: Do they have a role in the treatment of insulin resistance? Indian J. Med. Res. 2007, 125, 355–372. [Google Scholar]
- Wu, P.P.; Zhang, K.; Lu, Y.J.; He, P.; Zhao, S.Q. In vitro and in vivo evaluation of the antidiabetic activity of ursolic acid derivatives. Eur. J. Med. Chem. 2014, 80, 502–508. [Google Scholar] [CrossRef]
- Sambuy, Y.; de Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell. Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef]
- Delie, F.; Rubas, W. A human colonic cell line sharing similarities with enterocytes as a model to examine oral absorption: advantages and limitations of the Caco-2 model. Drug Carrier Syst. 1997, 14, 221–286. [Google Scholar]
- Mahraoui, L.; Rodolosse, A.; Barbat, A.; Dussaulx, E.; Zweibaum, A.; Rousset, M.; Brot-Laroche, E. Presence and differential expression of SGLT1, GLUT1, GLUT2, GLUT3 and GLUT5 hexose-transporter mRNAs in Caco-2 cell clones in relation to cell growth and glucose consumption. Biochem. J. 1994, 298, 629–633. [Google Scholar]
- Jang, S.M.; Yee, S.T.; Choi, J.; Choi, M.S.; Do, G.M.; Jeon, S.M.; Yeo, J.; Kim, M.J.; Seo, K.I.; Lee, M.K. Ursolic acid enhances the cellular immune system and pancreatic beta-cell function in streptozotocin-induced diabetic mice fed a high-fat diet. Int. Immunopharmacol. 2009, 9, 113–119. [Google Scholar] [CrossRef]
- Jang, S.M.; Kim, M.J.; Choi, M.S.; Kwon, E.Y.; Lee, M.K. Inhibitory effects of ursolic acid on hepatic polyol pathway and glucose production in streptozotocin-induced diabetic mice. Metabolism 2010, 59, 512–519. [Google Scholar] [CrossRef]
- Arya, A.; Looi, C.Y.; Cheah, S.C.; Mustafa, M.R.; Mohd, M.A. Anti-diabetic effects of Centratherum anthelminticum seeds methanolic fraction on pancreatic cells, beta-TC6 and its alleviating role in type 2 diabetic rats. J. Ethnopharmacol. 2012, 144, 22–32. [Google Scholar] [CrossRef]
- Poitout, V.; Stout, L.E.; Armstrong, M.B.; Walseth, T.F.; Sorenson, R.L.; Robertson, R.P. Morphological and functional characterization of beta TC-6 cells-an insulin-secreting cell line derived from transgenic mice. Diabetes 1995, 44, 306–313. [Google Scholar] [CrossRef]
- Yamada, K.; Nakata, M.; Horimoto, N.; Saito, M.; Matsuoka, H.; Inagaki, N. Measurement of glucose uptake and intracellular calcium concentration in single, living pancreatic beta-cells. J. Biol. Chem. 2000, 275, 22278–22283. [Google Scholar] [CrossRef]
- Masumoto, S.; Akimoto, Y.; Oike, H.; Kobori, M. Dietary phloridzin reduces blood glucose levels and reverses Sglt1 expression in the small intestine in streptozotocin-induced diabetic mice. J. Agric. Food. Chem. 2009, 57, 4651–4656. [Google Scholar] [CrossRef]
- Goto, T.; Horita, M.; Nagai, H.; Nagatomo, A.; Nishida, N.; Matsuura, Y.; Nagaoka, S. Tiliroside, a glycosidic flavonoid, inhibits carbohydrate digestion and glucose absorption in the gastrointestinal tract. Mol. Nutr. Food Res. 2012, 56, 435–445. [Google Scholar] [CrossRef]
- Kellett, G.L.; Brot-Laroche, E.; Mace, O.J.; Leturque, A. Sugar absorption in the intestine: The role of GLUT2. A. Leturque. Annu. Rev. Nutr. 2008, 28, 35–54. [Google Scholar] [CrossRef]
- Brown, G.K. Glucose transporters: Structure, function and consequences of deficiency. Metab. Dis. 2000, 23, 237–246. [Google Scholar]
- Manzano, S.; Williamson, G. Polyphenols and phenolic acids from strawberry and apple decrease glucose uptake and transport by human intestinal Caco-2 cells. Mol. Nutr. Food Res. 2010, 54, 1773–1780. [Google Scholar] [CrossRef]
- Kellett, G.L.; Brot-Laroche, E. Apical GLUT2: A major pathway of intestinal sugar absorption. Diabetes 2005, 54, 3056–3062. [Google Scholar] [CrossRef]
- Almdal, T.P.; Vilstrup, H. Strict insulin therapy normalises organ nitrogen contents and the capacity of urea nitrogen synthesis in experimental diabetes in rats. Diabetologia 1988, 31, 114–118. [Google Scholar] [CrossRef]
- Knopp, R.H.; Retzlaff, B.; Aikawa, K.; Kahn, S.E. Management of patients with diabetic hyperlipidemia. Am. J. Cardiol. 2003, 91 (Suppl. 1), 24–28. [Google Scholar] [CrossRef]
- Kavalali, G.; Tuncel, H.; Goksel, S.; Hatemi, H.H. Hypoglycemic activity of Urtica pilulifera in streptozotocin-diabetic rats. J. Ethnopharmacol. 2003, 84, 241–245. [Google Scholar] [CrossRef]
- Cemek, M.; Kaga, S.; Simsek, N.; Buyukokuroglu, M.E.; Konuk, M. Antihyperglycemic and antioxidative potential of Matricaria chamomilla L. in streptozotocin-induced diabetic rats. J. Nat. Med. 2008, 62, 284–293. [Google Scholar] [CrossRef]
- Fang, Y.Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Griesmacher, A.; Kindhauser, M.; Andert, S.E.; Schreiner, W.; Toma, C.; Knoebl, P.; Pietschmann, P.; Prager, R.; Schnack, C.; Schernthaner, G.; et al. Enhanced serum levels of thiobarbituric-acid-reactive substances in diabetes mellitus. Am. J. Med. 1995, 98, 469–475. [Google Scholar] [CrossRef]
- Dewanjee, S.; Das, A.K.; Sahu, R.; Gangopadhyay, M. Antidiabetic activity of Diospyros peregrina fruit: effect on hyperglycemia, hyperlipidemia and augmented oxidative stress in experimental type 2 diabetes. Food Chem. Toxicol. 2009, 47, 2679–2685. [Google Scholar] [CrossRef]
- Blake, D.R.; Allen, R.E.; Lunec, J. Free radicals in biological systems-A review orientated to inflammatory processes. Br. Med. Bull. 1987, 43, 371–385. [Google Scholar]
- Lin, Y.F.; Tsai, H.L.; Lee, Y.C.; Chang, S.J. Maternal vitamin E supplementation affects the antioxidant capability and oxidative status of hatching chicks. J. Nutr. 2005, 135, 2457–2461. [Google Scholar]
- Carr, M.C.; Brunzell, J.D. Abdominal obesity and dyslipidemia in the metabolic syndrome: Importance of type 2 diabetes and familial combined hyperlipidemia in coronary artery disease risk. J. Clin. Endocrinol. Metab. 2004, 89, 2601–2607. [Google Scholar] [CrossRef]
- Yoshihara, T.; Kumashiro, N.; Kanazawa, Y.; Mita, T.; Sakurai, Y.; Kawai, J.; Abe, M.; Motojima, K.; Hara, K.; Yamazaki, Y.; et al. Therapeutic efficacy of mitiglinide combined with once daily insulin glargine after switching from multiple daily insulin regimen of aspart insulin and glargine in patients with type 2 diabetes mellitus. Endocr. J. 2006, 53, 67–72. [Google Scholar] [CrossRef]
- Burnette, W.N. “Western blotting”: Electrophoretic transfer of proteins from sodium dodecyl sulfate-Polyacrylamide gels to unmodified nitrocellulose and radiographic detection with antibody and radioiodinated protein A. Anal. Biochem. 1981, 112, 195–203. [Google Scholar] [CrossRef]
- Shirwaikar, A.; Rajendran, K.; Barik, R. Effect of aqueous bark extract of Garuga pinnata Roxb. in streptozotocin-nicotinamide induced type-II diabetes mellitus. J. Ethnopharmacol. 2006, 107, 285–290. [Google Scholar] [CrossRef]
- Ghosh, M.N. Fundamentals of experimental pharmacology. Indian J. Pharmacol. 2007, 39, 216. [Google Scholar] [CrossRef]
- Turner, R.A.; Hebborn, P. Screening Methods in Pharmacology; Academic Press: New York, NY, USA, 1965; Volume 1, pp. 129–135. [Google Scholar]
- Sarkar, S.; Pranava, M.; Marita, R. Demonstration of the hypoglycemic action of Momordica charantia in a validated animal model of diabetes. Pharmacol. Res. 1996, 33, 1–4. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. Malondialdehyde determination as index of lipid peroxidation. Method. Enzymol. 1990, 186, 421–431. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are available from the authors.
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open ccess article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wu, P.; He, P.; Zhao, S.; Huang, T.; Lu, Y.; Zhang, K. Effects of Ursolic Acid Derivatives on Caco-2 Cells and Their Alleviating Role in Streptozocin-Induced Type 2 Diabetic Rats. Molecules 2014, 19, 12559-12576. https://doi.org/10.3390/molecules190812559
Wu P, He P, Zhao S, Huang T, Lu Y, Zhang K. Effects of Ursolic Acid Derivatives on Caco-2 Cells and Their Alleviating Role in Streptozocin-Induced Type 2 Diabetic Rats. Molecules. 2014; 19(8):12559-12576. https://doi.org/10.3390/molecules190812559
Chicago/Turabian StyleWu, Panpan, Ping He, Suqing Zhao, Tianming Huang, Yujing Lu, and Kun Zhang. 2014. "Effects of Ursolic Acid Derivatives on Caco-2 Cells and Their Alleviating Role in Streptozocin-Induced Type 2 Diabetic Rats" Molecules 19, no. 8: 12559-12576. https://doi.org/10.3390/molecules190812559
APA StyleWu, P., He, P., Zhao, S., Huang, T., Lu, Y., & Zhang, K. (2014). Effects of Ursolic Acid Derivatives on Caco-2 Cells and Their Alleviating Role in Streptozocin-Induced Type 2 Diabetic Rats. Molecules, 19(8), 12559-12576. https://doi.org/10.3390/molecules190812559