Ultraviolet Radiation, Aging and the Skin: Prevention of Damage by Topical cAMP Manipulation

{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
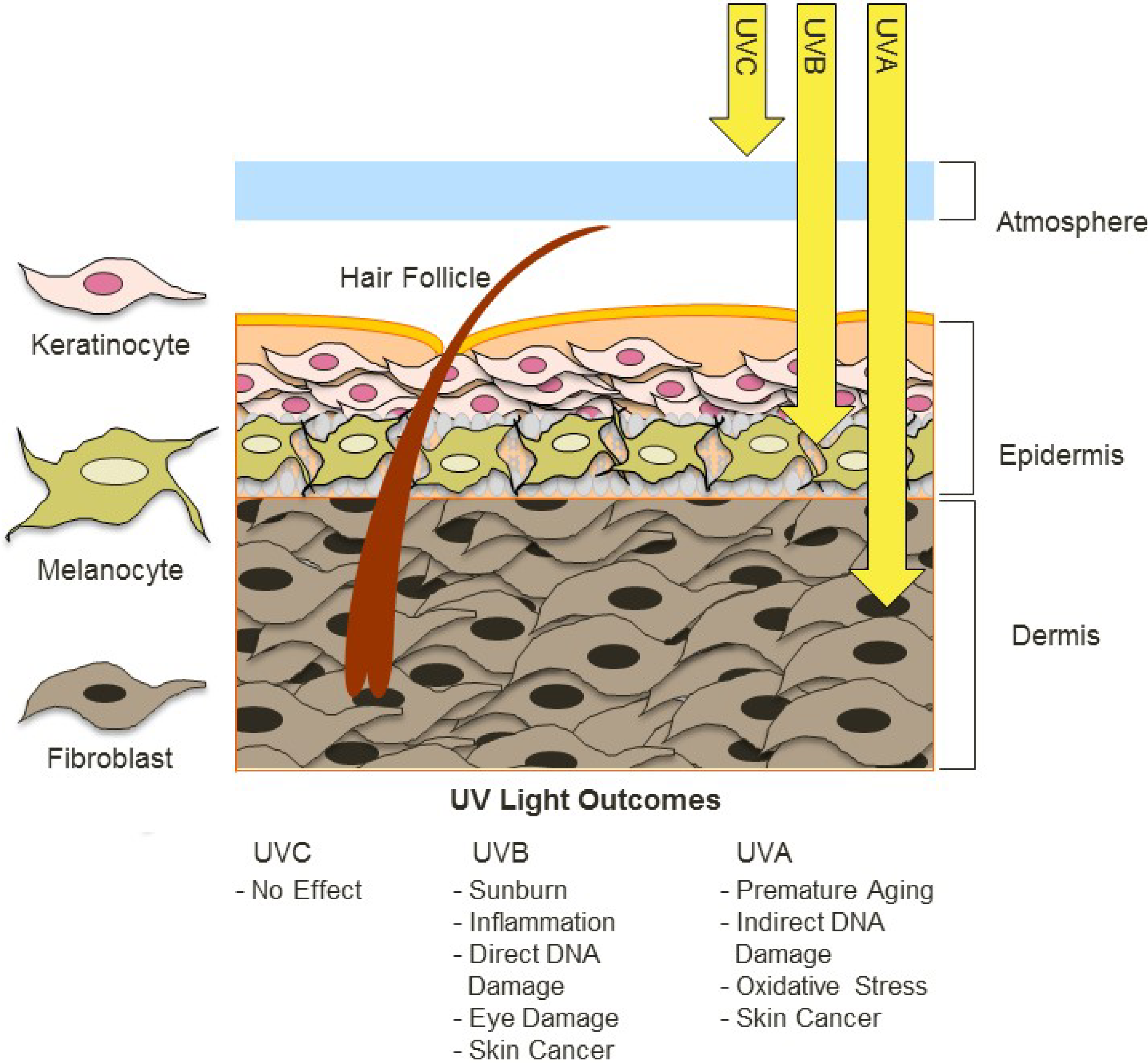
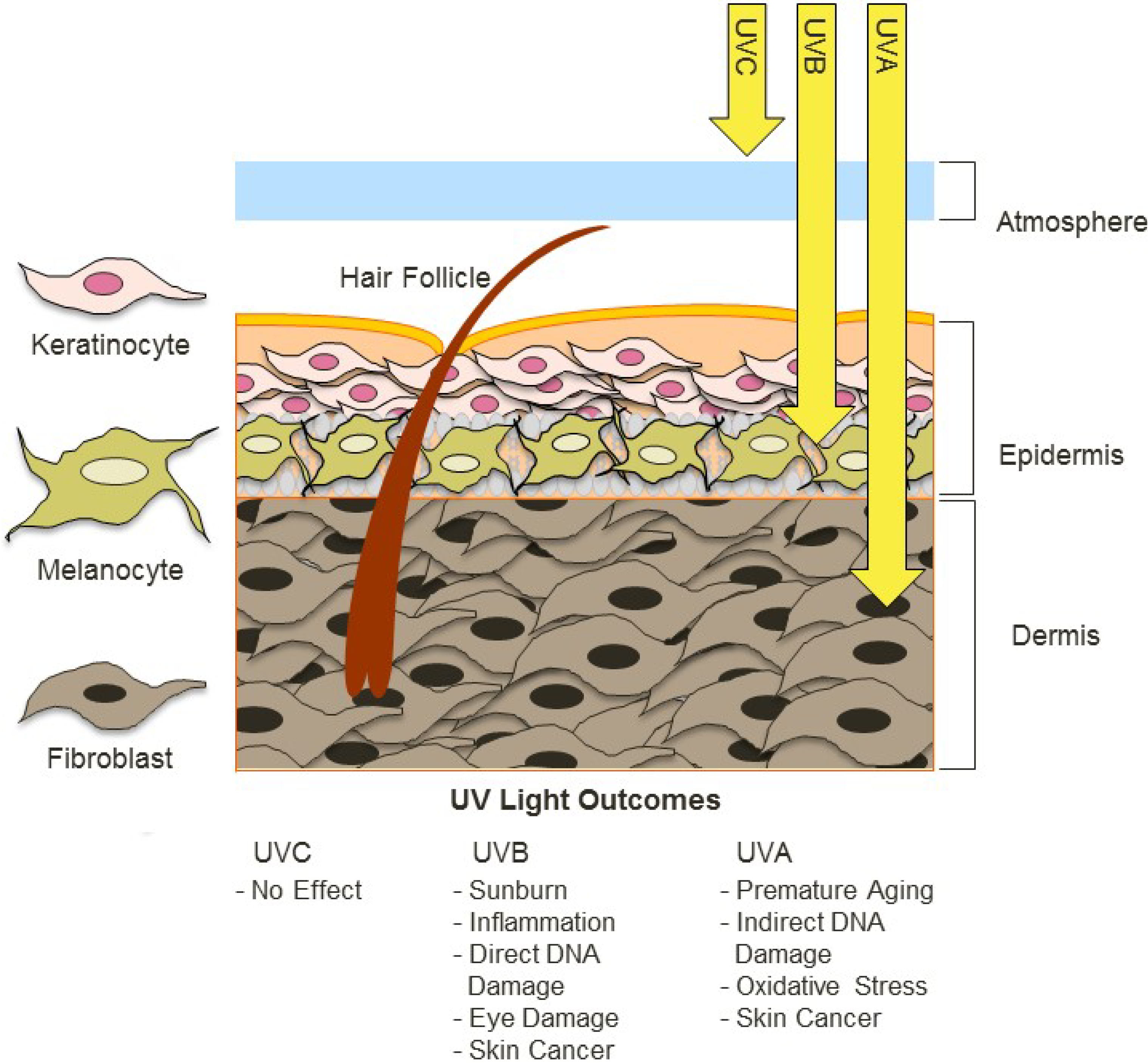
2. Factors Contributing to UV Exposure
3. Age
4. Artificial UV and Tanning Beds
5. Sunburns and Melanoma
6. The Melanocortin 1 Receptor (MC1R) and the Tanning Response
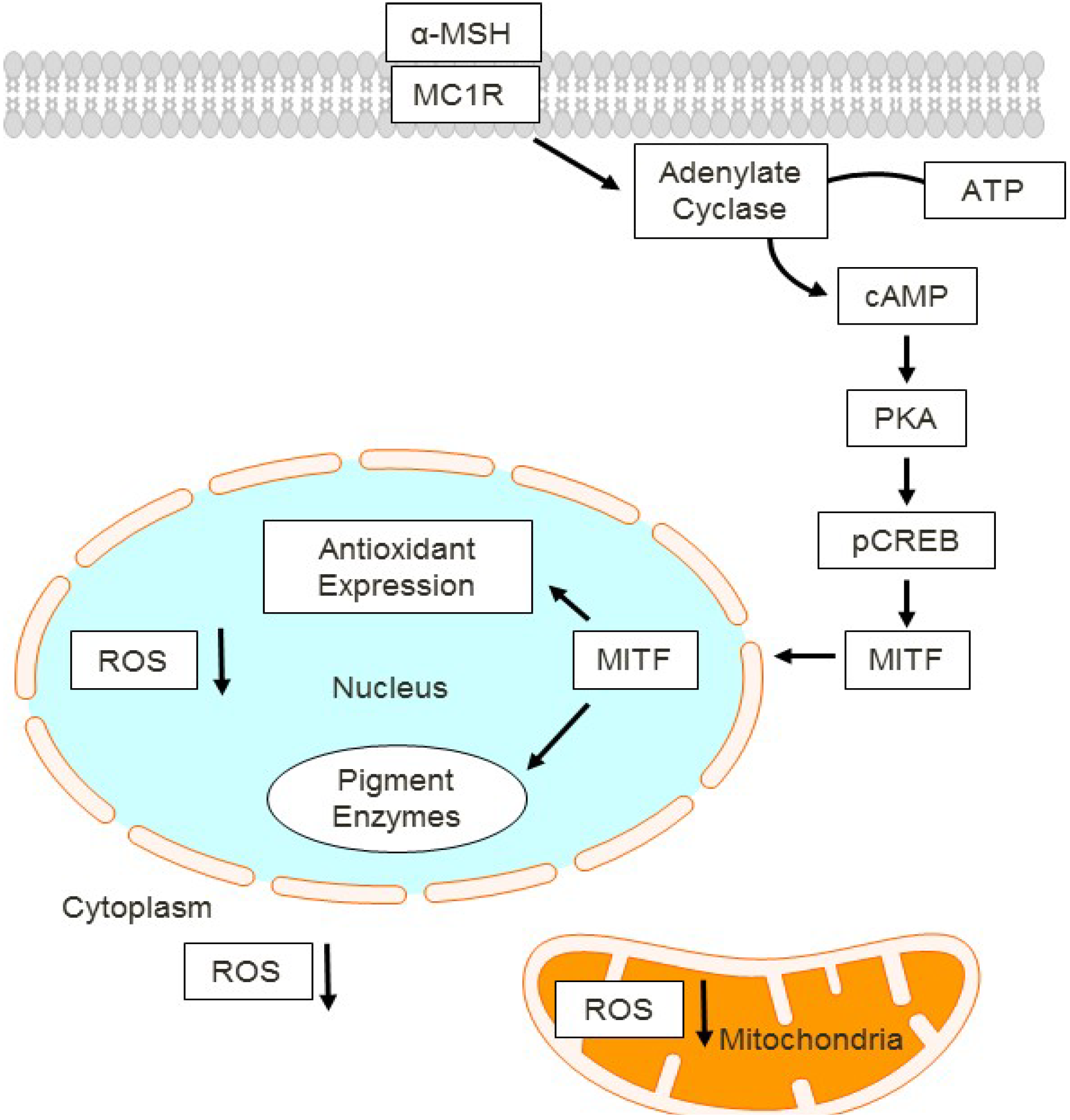
7. Pigmentation Phenotype Depends on MC1R Signaling
8. Forskolin Rescues cAMP Deficient Signaling
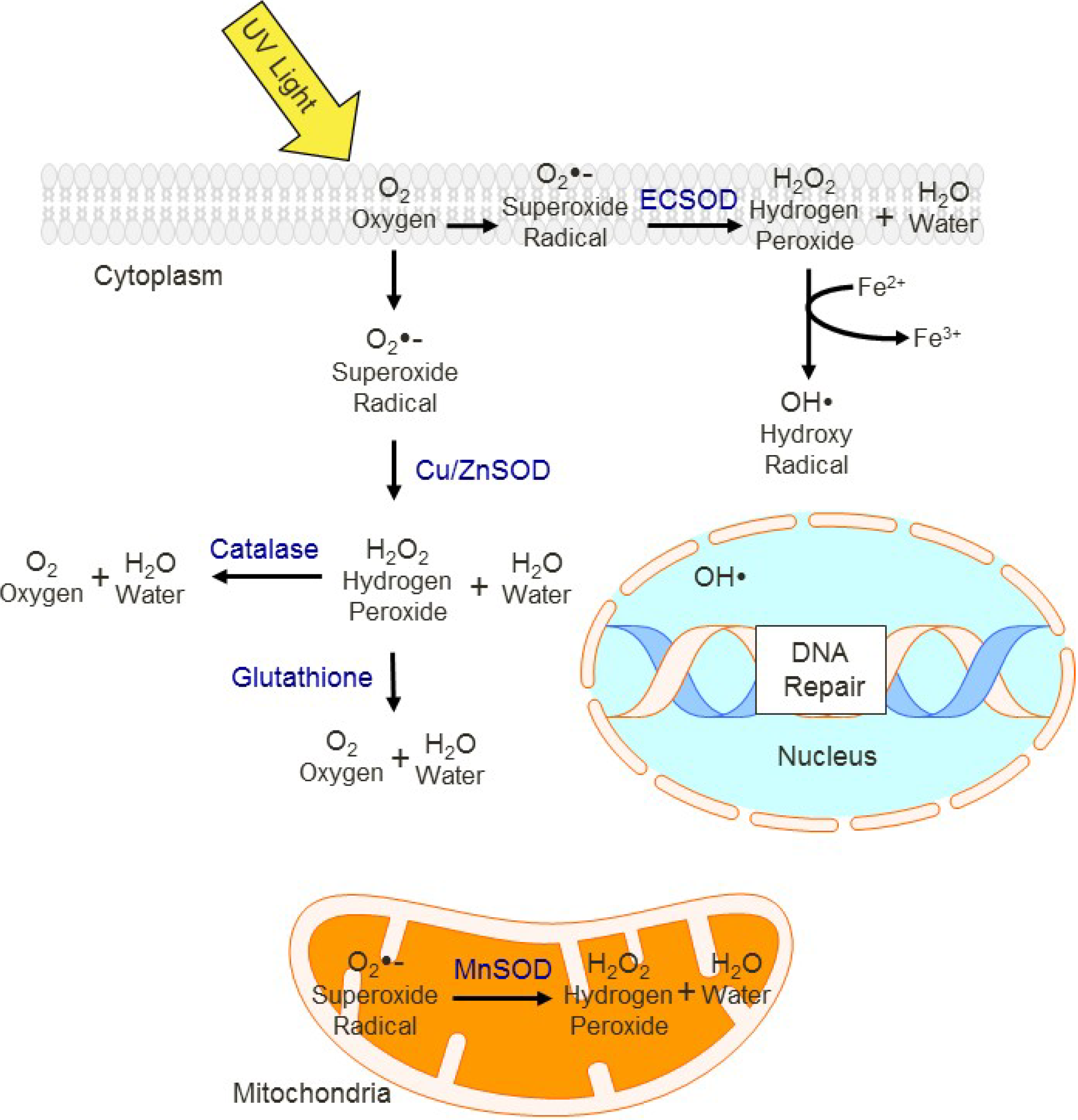
9. Oxidative Stress and Aging
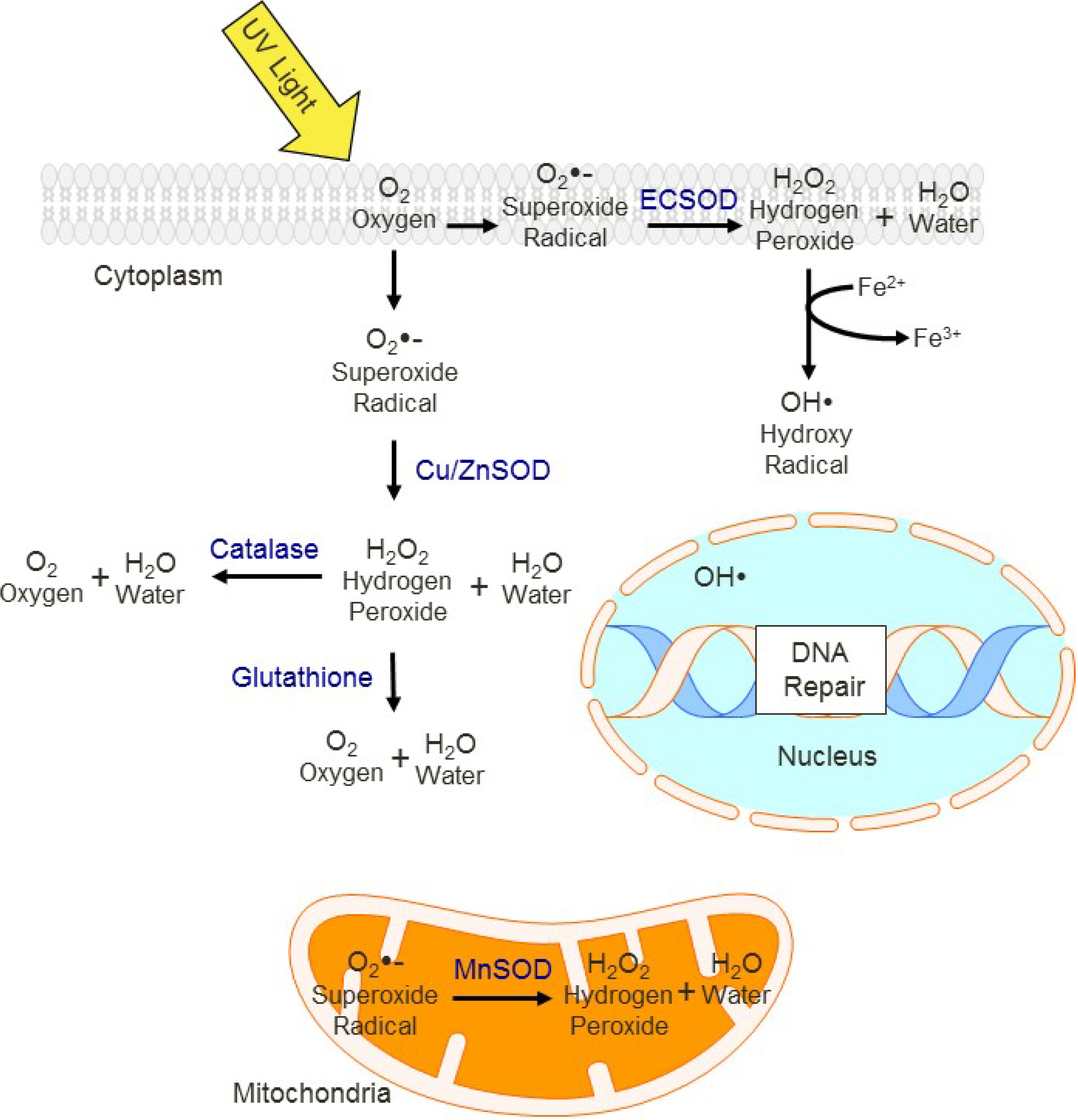
10. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Elwood, J.M.; Jopson, J. Melanoma and sun exposure: An overview of published studies. Int. J. Cancer 1997, 73, 198–203. [Google Scholar] [CrossRef]
- Godar, D.E. UV doses worldwide. Photochem. Photobiol. 2005, 81, 736–749. [Google Scholar] [CrossRef]
- Wei, Q.; Lee, J.E.; Gershenwald, J.E.; Ross, M.I.; Mansfield, P.F.; Strom, S.S.; Wang, L.E.; Guo, Z.; Qiao, Y.; Amos, C.I.; et al. Repair of UV light-induced DNA damage and risk of cutaneous malignant melanoma. J. Natl. Cancer Instit. 2003, 95, 308–315. [Google Scholar] [CrossRef]
- Cleaver, J.E.; Crowley, E. UV damage, DNA repair and skin carcinogenesis. Front. Biosci. 2002, 7, d1024–d1043. [Google Scholar] [CrossRef]
- Krutmann, J.; Morita, A.; Chung, J.H. Sun exposure: What molecular photodermatology tells us about its good and bad sides. J. Investig. Dermatol. 2012, 132, 976–984. [Google Scholar] [CrossRef]
- Sarasin, A. The molecular pathways of ultraviolet-induced carcinogenesis. Mutat. Res. 1999, 428, 5–10. [Google Scholar] [CrossRef]
- Hoeijmakers, J.H. DNA damage, aging, and cancer. N. Engl. J. Med. 2009, 361, 1475–1485. [Google Scholar] [CrossRef]
- Meyskens, F.L., Jr.; Farmer, P.; Fruehauf, J.P. Redox regulation in human melanocytes and melanoma. Pigment Cell Res. 2001, 14, 148–154. [Google Scholar]
- Schulz, I.; Mahler, H.C.; Boiteux, S.; Epe, B. Oxidative DNA base damage induced by singlet oxygen and photosensitization: Recognition by repair endonucleases and mutagenicity. Mutat. Res. 2000, 461, 145–156. [Google Scholar] [CrossRef]
- Norval, M.; Lucas, R.M.; Cullen, A.P.; de Gruijl, F.R.; Longstreth, J.; Takizawa, Y.; van der Leun, J.C. The human health effects of ozone depletion and interactions with climate change. Photochem. Photobiol. Sci. 2011, 10, 199–225. [Google Scholar] [CrossRef]
- Rigel, D.S.; Russak, J.; Friedman, R. The evolution of melanoma diagnosis: 25 years beyond the ABCDs. CA Cancer J. Clin. 2010, 60, 301–316. [Google Scholar] [CrossRef]
- Garbe, C.; Leiter, U. Melanoma epidemiology and trends. Clin. Dermatol. 2009, 27, 3–9. [Google Scholar] [CrossRef]
- Armstrong, B.K.; Kricker, A. The epidemiology of UV induced skin cancer. J. Photochem. Photobiol. B Biol. 2001, 63, 8–18. [Google Scholar]
- Diffey, B.L. The future incidence of cutaneous melanoma within the UK. Br. J. Dermatol. 2004, 151, 868–872. [Google Scholar] [CrossRef]
- Grant, W.B. The effect of solar UVB doses and vitamin D production, skin cancer action spectra, and smoking in explaining links between skin cancers and solid tumours. Eur. J. Cancer 2008, 44, 12–15. [Google Scholar] [CrossRef]
- Paunel, A.N.; Dejam, A.; Thelen, S.; Kirsch, M.; Horstjann, M.; Gharini, P.; Murtz, M.; Kelm, M.; de Groot, H.; Kolb-Bachofen, V.; et al. Enzyme-independent nitric oxide formation during UVA challenge of human skin: Characterization, molecular sources, and mechanisms. Free Radic. Biol. Med. 2005, 38, 606–615. [Google Scholar] [CrossRef]
- Venditti, E.; Bruge, F.; Astolfi, P.; Kochevar, I.; Damiani, E. Nitroxides and a nitroxide-based UV filter have the potential to photoprotect UVA-irradiated human skin fibroblasts against oxidative damage. J. Dermatol. Sci. 2011, 63, 55–61. [Google Scholar] [CrossRef]
- Akhalaya, M.Y.; Maksimov, G.V.; Rubin, A.B.; Lademann, J.; Darvin, M.E. Molecular action mechanisms of solar infrared radiation and heat on human skin. Ageing Res. Rev. 2014. [Google Scholar] [CrossRef]
- Yin, B.; Jiang, X. Telomere shortening in cultured human dermal fibroblasts is associated with acute photodamage induced by UVA irradiation. Postepy Dermatol. I Alergol. 2013, 30, 13–18. [Google Scholar]
- Atillasoy, E.S.; Seykora, J.T.; Soballe, P.W.; Elenitsas, R.; Nesbit, M.; Elder, D.E.; Montone, K.T.; Sauter, E.; Herlyn, M. UVB induces atypical melanocytic lesions and melanoma in human skin. Am. J. Pathol. 1998, 152, 1179–1186. [Google Scholar]
- Wendt, J.; Schanab, O.; Binder, M.; Pehamberger, H.; Okamoto, I. Site-dependent actinic skin damage as risk factor for melanoma in a central European population. Pigm. Cell Melanoma Res. 2012, 25, 234–242. [Google Scholar]
- Flament, F.; Bazin, R.; Laquieze, S.; Rubert, V.; Simonpietri, E.; Piot, B. Effect of the sun on visible clinical signs of aging in Caucasian skin. Clin. Cosmet. Investig. Dermatol. 2013, 6, 221–232. [Google Scholar]
- Breger, J.; Baeva, L.; Agrawal, A.; Shindell, E.; Godar, D.E. UVB-induced inflammatory cytokine release, DNA damage and apoptosis of human oral compared with skin tissue equivalents. Photochem. Photobiol. 2013, 89, 665–670. [Google Scholar] [CrossRef]
- Haluza, D.; Simic, S.; Moshammer, H. Temporal and spatial melanoma trends in Austria: An ecological study. Int. J. Environ. Res. Public Health 2014, 11, 734–748. [Google Scholar]
- Aceituno-Madera, P.; Buendia-Eisman, A.; Olmo, F.J.; Jimenez-Moleon, J.J.; Serrano-Ortega, S. Melanoma, altitude, and UV-B radiation. Actas Dermo-Sifiliograficas 2011, 102, 199–205. (In Spanish) [Google Scholar] [CrossRef]
- Hammar, N.; Linnersjo, A.; Alfredsson, L.; Dammstrom, B.G.; Johansson, M.; Eliasch, H. Cancer incidence in airline and military pilots in Sweden 1961–1996. Aviat. Space Environ. Med. 2002, 73, 2–7. [Google Scholar]
- Lichte, V.; Dennenmoser, B.; Dietz, K.; Hafner, H.M.; Schlagenhauff, B.; Garbe, C.; Fischer, J.; Moehrle, M. Professional risk for skin cancer development in male mountain guides–A cross-sectional study. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 797–804. [Google Scholar]
- Eide, M.J.; Weinstock, M.A. Association of UV index, latitude, and melanoma incidence in nonwhite populations–US Surveillance, Epidemiology, and End Results (SEER) Program, 1992 to 2001. Arch. Dermatol. 2005, 141, 477–481. [Google Scholar]
- Hu, S.; Ma, F.; Collado-Mesa, F.; Kirsner, R.S. UV radiation, latitude, and melanoma in US Hispanics and blacks. Arch. Dermatol. 2004, 140, 819–824. [Google Scholar]
- Cicarma, E.; Juzeniene, A.; Porojnicu, A.C.; Bruland, O.S.; Moan, J. Latitude gradient for melanoma incidence by anatomic site and gender in Norway 1966–2007. J. Photochem. Photobiol. B Biol. 2010, 101, 174–178. [Google Scholar] [CrossRef]
- Chang, Y.M.; Barrett, J.H.; Bishop, D.T.; Armstrong, B.K.; Bataille, V.; Bergman, W.; Berwick, M.; Bracci, P.M.; Elwood, J.M.; Ernstoff, M.S.; et al. Sun exposure and melanoma risk at different latitudes: A pooled analysis of 5700 cases and 7216 controls. Int. J. Epidemiol. 2009, 38, 814–830. [Google Scholar] [CrossRef]
- Qureshi, A.A.; Laden, F.; Colditz, G.A.; Hunter, D.J. Geographic variation and risk of skin cancer in US women. Differences between melanoma, squamous cell carcinoma, and basal cell carcinoma. Arch. Intern. Med. 2008, 168, 501–507. [Google Scholar] [CrossRef]
- Armstrong, B.K. Epidemiology of malignant melanoma: Intermittent or total accumulated exposure to the sun? J. Dermatol. Surg. Oncol. 1988, 14, 835–849. [Google Scholar] [CrossRef]
- Sharp, L.; Donnelly, D.; Hegarty, A.; Carsin, A.E.; Deady, S.; McCluskey, N.; Gavin, A.; Comber, H. Risk of several cancers is higher in urban areas after adjusting for socioeconomic status. results from a two-country population-based study of 18 common cancers. J. Urban Health 2014, in press. [Google Scholar]
- Brouse, C.H.; Hillyer, G.C.; Basch, C.E.; Neugut, A.I. Geography, facilities, and promotional strategies used to encourage indoor tanning in New York City. J. Commun. Health 2011, 36, 635–639. [Google Scholar] [CrossRef]
- Garbe, C.; Orfanos, C.E. Epidemiology of malignant melanoma in central Europe: Risk factors and prognostic predictors. Results of the Central Malignant Melanoma Registry of the German Dermatological Society. Pigm. Cell Res. 1992, 2, 285–294. [Google Scholar] [CrossRef]
- Bishop, J.N.; Bataille, V.; Gavin, A.; Lens, M.; Marsden, J.; Mathews, T.; Wheelhouse, C. The prevention, diagnosis, referral and management of melanoma of the skin: Concise guidelines. Clin. Med. 2007, 7, 283–290. [Google Scholar] [CrossRef]
- Volkmer, B.; Greinert, R. UV and children’s skin. Progr. Biophys. Mol. Biol. 2011, 107, 386–388. [Google Scholar] [CrossRef]
- Cust, A.E.; Jenkins, M.A.; Goumas, C.; Armstrong, B.K.; Schmid, H.; Aitken, J.F.; Giles, G.G.; Kefford, R.F.; Hopper, J.L.; Mann, G.J. Early-life sun exposure and risk of melanoma before age 40 years. Cancer Causes Control 2011, 22, 885–897. [Google Scholar] [CrossRef]
- Crump, C.; Sundquist, K.; Sieh, W.; Winkleby, M.A.; Sundquist, J. Season of birth and other perinatal risk factors for melanoma. Int. J. Epidemiol. 2014, in press. [Google Scholar]
- Purdue, M.P.; Freeman, L.E.; Anderson, W.F.; Tucker, M.A. Recent trends in incidence of cutaneous melanoma among US Caucasian young adults. J. Investig. Dermatol. 2008, 128, 2905–2908. [Google Scholar] [CrossRef]
- Wehner, M.R.; Chren, M.M.; Nameth, D.; Choudhry, A.; Gaskins, M.; Nead, K.T.; Boscardin, W.J.; Linos, E. International prevalence of indoor tanning: A systematic review and meta-analysis. JAMA Dermatol. 2014. [Google Scholar] [CrossRef]
- Heckman, C.J.; Cohen-Filipic, J.; Darlow, S.; Kloss, J.D.; Manne, S.L.; Munshi, T. Psychiatric and addictive symptoms of young adult female indoor tanners. Am. J. Health Promot. 2014, 28, 168–174. [Google Scholar] [CrossRef]
- Dore, J.F.; Chignol, M.C. Tanning salons and skin cancer. Photochem. Photobiol. Sci. 2012, 11, 30–37. [Google Scholar] [CrossRef]
- Petit, A.; Karila, L.; Chalmin, F.; Lejoyeux, M. Phenomenology and psychopathology of excessive indoor tanning. Int. J. Dermatol. 2014. [Google Scholar] [CrossRef]
- Petit, A.; Lejoyeux, M.; Reynaud, M.; Karila, L. Excessive indoor tanning as a behavioral addiction: A literature review. Curr. Pharmaceut. Des. 2013, in press. [Google Scholar]
- Lazovich, D.; Vogel, R.I.; Berwick, M.; Weinstock, M.A.; Anderson, K.E.; Warshaw, E.M. Indoor tanning and risk of melanoma: A case-control study in a highly exposed population. Cancer Epidemiol. Biomark. Prev. 2010, 19, 1557–1568. [Google Scholar] [CrossRef]
- Pfahlberg, A.; Kolmel, K.F.; Gefeller, O.; Febim Study, G. Timing of excessive ultraviolet radiation and melanoma: Epidemiology does not support the existence of a critical period of high susceptibility to solar ultraviolet radiation-induced melanoma. Br. J. Dermatol. 2001, 144, 471–475. [Google Scholar] [CrossRef]
- Centers for Disease, C. Prevention, sunburn and sun protective behaviors among adults aged 18–29 years–United States, 2000–2010. MMWR. Morbid. Mortal. Wkl. Rep. 2012, 61, 317–322. [Google Scholar]
- Behrens, C.L.; Thorgaard, C.; Philip, A.; Bentzen, J. Sunburn in children and adolescents: Associations with parents’ behaviour and attitudes. Scand. J. Public Health 2013, 41, 302–310. [Google Scholar] [CrossRef]
- Suzuki, I.; Cone, R.D.; Im, S.; Nordlund, J.; Abdel-Malek, Z.A. Binding of melanotropic hormones to the melanocortin receptor MC1R on human melanocytes stimulates proliferation and melanogenesis. Endocrinology 1996, 137, 1627–1633. [Google Scholar]
- Spry, M.L.; Vanover, J.C.; Scott, T.; Abona-Ama, O.; Wakamatsu, K.; Ito, S.; D’Orazio, J.A. Prolonged treatment of fair-skinned mice with topical forskolin causes persistent tanning and UV protection. Pigm. Cell Melanoma Res. 2009, 22, 219–229. [Google Scholar] [CrossRef]
- D’Orazio, J.A.; Nobuhisa, T.; Cui, R.; Arya, M.; Spry, M.; Wakamatsu, K.; Igras, V.; Kunisada, T.; Granter, S.R.; Nishimura, E.K.; et al. Topical drug rescue strategy and skin protection based on the role of Mc1r in UV-induced tanning. Nature 2006, 443, 340–344. [Google Scholar] [CrossRef]
- Cui, R.; Widlund, H.R.; Feige, E.; Lin, J.Y.; Wilensky, D.L.; Igras, V.E.; D’Orazio, J.; Fung, C.Y.; Schanbacher, C.F.; Granter, S.R.; et al. Central role of p53 in the suntan response and pathologic hyperpigmentation. Cell 2007, 128, 853–864. [Google Scholar] [CrossRef]
- Amaro-Ortiz, A.; Vanover, J.C.; Scott, T.L.; D’Orazio, J.A. Pharmacologic induction of epidermal melanin and protection against sunburn in a humanized mouse model. J. Vis. Exp. 2013. [Google Scholar] [CrossRef]
- Tolleson, W.H. Human melanocyte biology, toxicology, and pathology. J. Environ. Sci. Health Part C Environ. Carcinogen. Ecotoxicol. Rev. 2005, 23, 105–161. [Google Scholar] [CrossRef]
- Scott, M.C.; Wakamatsu, K.; Ito, S.; Kadekaro, A.L.; Kobayashi, N.; Groden, J.; Kavanagh, R.; Takakuwa, T.; Virador, V.; Hearing, V.J.; et al. Human melanocortin 1 receptor variants, receptor function and melanocyte response to UV radiation. J. Cell Sci. 2002, 115, 2349–2355. [Google Scholar]
- Kadekaro, A.L.; Kanto, H.; Kavanagh, R.; Abdel-Malek, Z. Significance of the melanocortin 1 receptor in regulating human melanocyte pigmentation, proliferation, and survival. Ann. N. Y. Acad. Sci. 2003, 994, 359–365. [Google Scholar] [CrossRef]
- Rouzaud, F.; Kadekaro, A.L.; Abdel-Malek, Z.A.; Hearing, V.J. MC1R and the response of melanocytes to ultraviolet radiation. Mutat. Res. 2005, 571, 133–152. [Google Scholar] [CrossRef]
- Bohm, M.; Wolff, I.; Scholzen, T.E.; Robinson, S.J.; Healy, E.; Luger, T.A.; Schwarz, T.; Schwarz, A. alpha-Melanocyte-stimulating hormone protects from ultraviolet radiation-induced apoptosis and DNA damage. J. Biol. Chem. 2005, 280, 5795–5802. [Google Scholar] [CrossRef]
- Hauser, J.E.; Kadekaro, A.L.; Kavanagh, R.J.; Wakamatsu, K.; Terzieva, S.; Schwemberger, S.; Babcock, G.; Rao, M.B.; Ito, S.; Abdel-Malek, Z.A. Melanin content and MC1R function independently affect UVR-induced DNA damage in cultured human melanocytes. Pigm. Cell Res. 2006, 19, 303–314. [Google Scholar] [CrossRef]
- Abdel-Malek, Z.; Suzuki, I.; Tada, A.; Im, S.; Akcali, C. The melanocortin-1 receptor and human pigmentation. Ann. N. Y. Acad. Sci. 1999, 885, 117–133. [Google Scholar]
- Abdel-Malek, Z.; Scott, M.C.; Suzuki, I.; Tada, A.; Im, S.; Lamoreux, L.; Ito, S.; Barsh, G.; Hearing, V.J. The melanocortin-1 receptor is a key regulator of human cutaneous pigmentation. Pigm. Cell Res. 2000, 13, 156–162. [Google Scholar]
- Abdel-Malek, Z.A.; Knittel, J.; Kadekaro, A.L.; Swope, V.B.; Starner, R. The melanocortin 1 receptor and the UV response of human melanocytes–A shift in paradigm. Photochem. Photobiol. 2008, 84, 501–508. [Google Scholar]
- Bastiaens, M.T.; ter Huurne, J.A.; Kielich, C.; Gruis, N.A.; Westendorp, R.G.; Vermeer, B.J.; Bavinck, J.N.; Leiden Skin Cancer Study, T. Melanocortin-1 receptor gene variants determine the risk of nonmelanoma skin cancer independently of fair skin and red hair. Am. J. Hum. Genet. 2001, 68, 884–894. [Google Scholar] [CrossRef]
- Gruis, N.A.; van Doorn, R. Melanocortin 1 receptor function: Shifting gears from determining skin and nevus phenotype to fetal growth. J. Investig. Dermatol. 2012, 132, 1953–1955. [Google Scholar] [CrossRef]
- Quint, K.D.; van der Rhee, J.I.; Gruis, N.A.; Ter Huurne, J.A.; Wolterbeek, R.; van der Stoep, N.; Bergman, W.; Kukutsch, N.A. Melanocortin 1 receptor (MC1R) variants in high melanoma risk patients are associated with specific dermoscopic ABCD features. Acta Dermato-Venereol. 2012, 92, 587–592. [Google Scholar] [CrossRef]
- Prunier, C.; Masson-Genteuil, G.; Ugolin, N.; Sarrazy, F.; Sauvaigo, S. Aging and photo-aging DNA repair phenotype of skin cells-evidence toward an effect of chronic sun-exposure. Mutat. Res. 2012, 736, 48–55. [Google Scholar] [CrossRef]
- Pullar, C.E.; Isseroff, R.R. Cyclic AMP mediates keratinocyte directional migration in an electric field. J. Cell Sci. 2005, 118, 2023–2034. [Google Scholar] [CrossRef]
- Spindler, V.; Vielmuth, F.; Schmidt, E.; Rubenstein, D.S.; Waschke, J. Protective endogenous cyclic adenosine 5'-monophosphate signaling triggered by pemphigus autoantibodies. J. Immunol. 2010, 185, 6831–6838. [Google Scholar] [CrossRef]
- De Vries, G.W.; Amdahl, L.D.; Lowe, N.; Wheeler, L.A. Effect of forskolin on beta-adrenergic hyporesponsiveness in skin. Skin Pharmacol. 1988, 1, 106–114. [Google Scholar] [CrossRef]
- Harmon, C.S.; Nevins, T.D. Evidence that activation of protein kinase A inhibits human hair follicle growth and hair fibre production in organ culture and DNA synthesis in human and mouse hair follicle organ culture. Br. J. Dermatol. 1997, 136, 853–858. [Google Scholar] [CrossRef]
- Michelet, J.F.; Gautier, B.; Gaillard, O.; Bernard, B.A.; Benech, F. Human hair follicle pigmentary unit as a direct target for modulators of melanogenesis, as studied by [14C]-2-thiouracil incorporation. Exp. Dermatol. 2009, 18, 414–416. [Google Scholar] [CrossRef]
- Kadekaro, A.L.; Chen, J.; Yang, J.; Chen, S.; Jameson, J.; Swope, V.B.; Cheng, T.; Kadakia, M.; Abdel-Malek, Z. Alpha-melanocyte-stimulating hormone suppresses oxidative stress through a p53-mediated signaling pathway in human melanocytes. Mol. Cancer Res. 2012, 10, 778–786. [Google Scholar] [CrossRef]
- Passeron, T.; Namiki, T.; Passeron, H.J.; le Pape, E.; Hearing, V.J. Forskolin protects keratinocytes from UVB-induced apoptosis and increases DNA repair independent of its effects on melanogenesis. J. Investig. Dermatol. 2009, 129, 162–166. [Google Scholar] [CrossRef]
- Scott, T.L.; Christian, P.A.; Kesler, M.V.; Donohue, K.M.; Shelton, B.; Wakamatsu, K.; Ito, S.; D'Orazio, J. Pigment-independent cAMP-mediated epidermal thickening protects against cutaneous UV injury by keratinocyte proliferation. Exp. Dermatol. 2012, 21, 771–777. [Google Scholar]
- Al-Ayadhi, L.Y.; Korish, A.A.; Al-Tuwaijri, A.S. The effect of vitamin E, L-arginine, N-nitro L-arginine methyl ester and forskolin on endocrine and metabolic changes of rats exposed to acute cold stress. Saudi Med. J. 2006, 27, 17–22. [Google Scholar]
- Mishima, K.; Baba, A.; Matsuo, M.; Itoh, Y.; Oishi, R. Protective effect of cyclic AMP against cisplatin-induced nephrotoxicity. Free Radic. Biol. Med. 2006, 40, 1564–1577. [Google Scholar] [CrossRef]
- Zastrow, L.; Groth, N.; Klein, F.; Kockott, D.; Lademann, J.; Renneberg, R.; Ferrero, L. The missing link--light-induced (280–1600 nm) free radical formation in human skin. Skin Pharmacol. Physiol. 2009, 22, 31–44. [Google Scholar] [CrossRef]
- Afanas’ev, I.B. Signaling by reactive oxygen and nitrogen species in skin diseases. Curr. Drug MeTable 2010, 11, 409–414. [Google Scholar] [CrossRef]
- Bossi, O.; Gartsbein, M.; Leitges, M.; Kuroki, T.; Grossman, S.; Tennenbaum, T. UV irradiation increases ROS production via PKCdelta signaling in primary murine fibroblasts. J. Cell. Biochem. 2008, 105, 194–207. [Google Scholar] [CrossRef]
- Choi, Y.J.; Uehara, Y.; Park, J.Y.; Chung, K.W.; Ha, Y.M.; Kim, J.M.; Song, Y.M.; Chun, P.; Park, J.W.; Moon, H.R.; et al. Suppression of melanogenesis by a newly synthesized compound, MHY966 via the nitric oxide/protein kinase G signaling pathway in murine skin. J. Dermatol. Sci. 2012, 68, 164–171. [Google Scholar] [CrossRef]
- Gabe, Y.; Osanai, O.; Takema, Y. The relationship between skin aging and steady state ultraweak photon emission as an indicator of skin oxidative stress in vivo. Skin Res. Technol 2013. [Google Scholar] [CrossRef]
- Lan, C.C.; Wu, C.S.; Yu, H.S. Solar-simulated radiation and heat treatment induced metalloproteinase-1 expression in cultured dermal fibroblasts via distinct pathways: Implications on reduction of sun-associated aging. J. Dermatol. Sci. 2013, 72, 290–295. [Google Scholar] [CrossRef]
- Poljsak, B.; Dahmane, R. Free radicals and extrinsic skin aging. Dermatol. Res. Pract. 2012, 2012, 135206:1–135206:4. [Google Scholar]
- Langton, A.K.; Sherratt, M.J.; Griffiths, C.E.; Watson, R.E. A new wrinkle on old skin: The role of elastic fibres in skin ageing. Int. J. Cosmet. Sci. 2010. [Google Scholar] [CrossRef]
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef]
- Mahmood, T.; Akhtar, N.; Khan, B.A.; Shoaib Khan, H.M.; Saeed, T. Changes in skin mechanicalproperties after long-term application of cream containing green tea extract. Aging Clin. Exp. Res. 2011, 23, 333–336. [Google Scholar] [CrossRef]
- Lee, C.W.; Park, N.H.; Kim, J.W.; Um, B.H.; Shpatov, A.V.; Shults, E.E.; Sorokina, I.V.; Popov, S.A. Study of skin anti-ageing and anti-inflammatory effects of dihydroquercetin, natural triterpenoids, and their synthetic derivatives. Bioorg. Khim. 2012, 38, 374–381. [Google Scholar]
- Stohs, S.J. The role of free radicals in toxicity and disease. J. Basic Clin. Physiol. Pharmacol. 1995, 6, 205–228. [Google Scholar] [CrossRef]
- Liu, L.; Xie, H.; Chen, X.; Shi, W.; Xiao, X.; Lei, D.; Li, J. Differential response of normal human epidermal keratinocytes and HaCaT cells to hydrogen peroxide-induced oxidative stress. Clin. Exp. Dermatol. 2012, 37, 772–780. [Google Scholar] [CrossRef]
- Yun, J.S.; Pahk, J.W.; Lee, J.S.; Shin, W.C.; Lee, S.Y.; Hong, E.K. Inonotus obliquus protects against oxidative stress-induced apoptosis and premature senescence. Mol. Cells 2011, 31, 423–429. [Google Scholar] [CrossRef]
- Velarde, M.C.; Flynn, J.M.; Day, N.U.; Melov, S.; Campisi, J. Mitochondrial oxidative stress caused by Sod2 deficiency promotes cellular senescence and aging phenotypes in the skin. Aging 2012, 4, 3–12. [Google Scholar]
- Sakura, M.; Chiba, Y.; Kamiya, E.; Furukawa, A.; Kawamura, N.; Niwa, M.; Takeuchi, M.; Hosokawa, M. Spontaneous occurrence of photoageing-like phenotypes in the dorsal skin of old SAMP1 mice, an oxidative stress model. Exp. Dermatol. 2013, 22, 62–64. [Google Scholar] [CrossRef]
- Makrantonaki, E.; Zouboulis, C.C.; William, J. Cunliffe Scientific Awards. Characteristics and pathomechanisms of endogenously aged skin. Dermatology 2007, 214, 352–360. [Google Scholar] [CrossRef]
- Davis, T.; Wyllie, F.S.; Rokicki, M.J.; Bagley, M.C.; Kipling, D. The role of cellular senescence in Werner syndrome: Toward therapeutic intervention in human premature aging. Ann. N. Y. Acad. Sci. 2007, 1100, 455–469. [Google Scholar] [CrossRef]
- Yokoo, S.; Furumoto, K.; Hiyama, E.; Miwa, N. Slow-down of age-dependent telomere shortening is executed in human skin keratinocytes by hormesis-like-effects of trace hydrogen peroxide or by anti-oxidative effects of pro-vitamin C in common concurrently with reduction of intracellular oxidative stress. J. Cell. Biochem. 2004, 93, 588–597. [Google Scholar] [CrossRef]
- Kashino, G.; Kodama, S.; Nakayama, Y.; Suzuki, K.; Fukase, K.; Goto, M.; Watanabe, M. Relief of oxidative stress by ascorbic acid delays cellular senescence of normal human and Werner syndrome fibroblast cells. Free Radic. Biol. Med. 2003, 35, 438–443. [Google Scholar] [CrossRef]
- Kong, S.Z.; Shi, X.G.; Feng, X.X.; Li, W.J.; Liu, W.H.; Chen, Z.W.; Xie, J.H.; Lai, X.P.; Zhang, S.X.; Zhang, X.J.; et al. Inhibitory effect of hydroxysafflor yellow a on mouse skin photoaging induced by ultraviolet irradiation. Rejuven. Res. 2013, 16, 404–143. [Google Scholar] [CrossRef]
- Felippi, C.C.; Oliveira, D.; Stroher, A.; Carvalho, A.R.; van Etten, E.A.; Bruschi, M.; Raffin, R.P. Safety and efficacy of antioxidants-loaded nanoparticles for an anti-aging application. J. Biomed. Nanotechnol. 2012, 8, 316–321. [Google Scholar] [CrossRef]
- Cornacchione, S.; Sadick, N.S.; Neveu, M.; Talbourdet, S.; Lazou, K.; Viron, C.; Renimel, I.; de Queral, D.; Kurfurst, R.; Schnebert, S.; et al. In vivo skin antioxidant effect of a new combination based on a specific Vitis vinifera shoot extract and a biotechnological extract. J. Drugs Dermatol. 2007, 6, s8–s13. [Google Scholar]
- Raschke, T.; Koop, U.; Dusing, H.J.; Filbry, A.; Sauermann, K.; Jaspers, S.; Wenck, H.; Wittern, K.P. Topical activity of ascorbic acid: From in vitro optimization to in vivo efficacy. Skin Pharmacol. Physiol. 2004, 17, 200–206. [Google Scholar] [CrossRef]
- Imokawa, G. Mechanism of UVB-induced wrinkling of the skin: Paracrine cytokine linkage between keratinocytes and fibroblasts leading to the stimulation of elastase. J. Investig. Dermatol. Symp. Proc. 2009, 14, 36–43. [Google Scholar] [CrossRef]
- Shin, M.H.; Seo, J.E.; Kim, Y.K.; Kim, K.H.; Chung, J.H. Chronic heat treatment causes skin wrinkle formation and oxidative damage in hairless mice. Mechan. Ageing Dev. 2012, 133, 92–98. [Google Scholar] [CrossRef]
- Gonzalez, S.; Pathak, M.A.; Cuevas, J.; Villarrubia, V.G.; Fitzpatrick, T.B. Topical or oral administration with an extract of Polypodium leucotomos prevents acute sunburn and psoralen-induced phototoxic reactions as well as depletion of Langerhans cells in human skin. Photodermatol. Photoimmunol. Photomed. 1997, 13, 50–60. [Google Scholar] [CrossRef]
- Kalka, K.; Mukhtar, H.; Turowski-Wanke, A.; Merk, H. Biomelanin antioxidants in cosmetics: Assessment based on inhibition of lipid peroxidation. Skin Pharmacol. Appl. Skin Physiol. 2000, 13, 143–149. [Google Scholar] [CrossRef]
- Cesarini, J.P.; Michel, L.; Maurette, J.M.; Adhoute, H.; Bejot, M. Immediate effects of UV radiation on the skin: Modification by an antioxidant complex containing carotenoids. Photodermatol. Photoimmunol. Photomed. 2003, 19, 182–189. [Google Scholar] [CrossRef]
- Oresajo, C.; Stephens, T.; Hino, P.D.; Law, R.M.; Yatskayer, M.; Foltis, P.; Pillai, S.; Pinnell, S.R. Protective effects of a topical antioxidant mixture containing vitamin C, ferulic acid, and phloretin against ultraviolet-induced photodamage in human skin. J. Cosmet. Dermatol. 2008, 7, 290–297. [Google Scholar] [CrossRef]
- Kimura, Y.; Sumiyoshi, M. French maritime pine bark (Pinus maritima Lam.) extract (Flavangenol) prevents chronic UVB radiation-induced skin damage and carcinogenesis in melanin-possessing hairless mice. Photochem. Photobiol. 2010, 86, 955–963. [Google Scholar] [CrossRef]
- Veratti, E.; Rossi, T.; Giudice, S.; Benassi, L.; Bertazzoni, G.; Morini, D.; Azzoni, P.; Bruni, E.; Giannetti, A.; Magnoni, C. 18beta-glycyrrhetinic acid and glabridin prevent oxidative DNA fragmentation in UVB-irradiated human keratinocyte cultures. Anticancer Res. 2011, 31, 2209–2215. [Google Scholar]
- Valacchi, G.; Pecorelli, A.; Mencarelli, M.; Maioli, E.; Davis, P.A. Beta-carotene prevents ozone-induced proinflammatory markers in murine skin. Toxicol. Ind. Health 2009, 25, 241–247. [Google Scholar] [CrossRef]
- Afaq, F.; Zaid, M.A.; Khan, N.; Dreher, M.; Mukhtar, H. Protective effect of pomegranate-derivedproducts on UVB-mediated damage in human reconstituted skin. Exp. Dermatol. 2009, 18, 553–561. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Tulipani, S.; Gonzales-Paramas, A.M.; Santos-Buelga, C.; Bompadre, S.; Quiles, J.L.; Mezzetti, B.; Battino, M. Photoprotective potential of strawberry (Fragaria x ananassa) extract against UV-A irradiation damage on human fibroblasts. J. Agric. Food Chem. 2012, 60, 2322–2327. [Google Scholar] [CrossRef]
- Lademann, J.; Schanzer, S.; Meinke, M.; Sterry, W.; Darvin, M.E. Interaction between carotenoids and free radicals in human skin. Skin Pharmacol. Physiol. 2011, 24, 238–244. [Google Scholar] [CrossRef]
- Haugen, L.; Bjornson, T. Beta Carotene: Dietary Sources, Cancer and Cognition; Nova Biomedical Books: New York, NY, USA, 2009; p. 358. [Google Scholar]
- Darvin, M.E.; Richter, H.; Ahlberg, S.; Haag, S.F.; Meinke, M.C.; le Quintrec, D.; Doucet, O.; Lademann, J. Influence of sun exposure on the cutaneous collagen/elastin fibers and carotenoids: Negative effects can be reduced by application of sunscreen. J. Biophoton. 2014. [Google Scholar] [CrossRef]
- Henri, P.; Beaumel, S.; Guezennec, A.; Poumes, C.; Stoebner, P.E.; Stasia, M.J.; Guesnet, J.; Martinez, J.; Meunier, L. MC1R expression in HaCaT keratinocytes inhibits UVA-induced ROS production via NADPH oxidase- and cAMP-dependent mechanisms. J. Cell. Physiol. 2012, 227, 2578–2585. [Google Scholar] [CrossRef]
- Song, X.; Mosby, N.; Yang, J.; Xu, A.; Abdel-Malek, Z.; Kadekaro, A.L. Alpha-MSH activates immediate defense responses to UV-induced oxidative stress in human melanocytes. Pigm. Cell Melanoma Res. 2009, 22, 809–818. [Google Scholar] [CrossRef]
© 2014 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Amaro-Ortiz, A.; Yan, B.; D'Orazio, J.A. Ultraviolet Radiation, Aging and the Skin: Prevention of Damage by Topical cAMP Manipulation. Molecules 2014, 19, 6202-6219. https://doi.org/10.3390/molecules19056202
Amaro-Ortiz A, Yan B, D'Orazio JA. Ultraviolet Radiation, Aging and the Skin: Prevention of Damage by Topical cAMP Manipulation. Molecules. 2014; 19(5):6202-6219. https://doi.org/10.3390/molecules19056202
Chicago/Turabian StyleAmaro-Ortiz, Alexandra, Betty Yan, and John A. D'Orazio. 2014. "Ultraviolet Radiation, Aging and the Skin: Prevention of Damage by Topical cAMP Manipulation" Molecules 19, no. 5: 6202-6219. https://doi.org/10.3390/molecules19056202