Identification of Fungicidal 2,6-Disubstituted Quinolines with Activity against Candida Biofilms

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Screening and Hit Identification

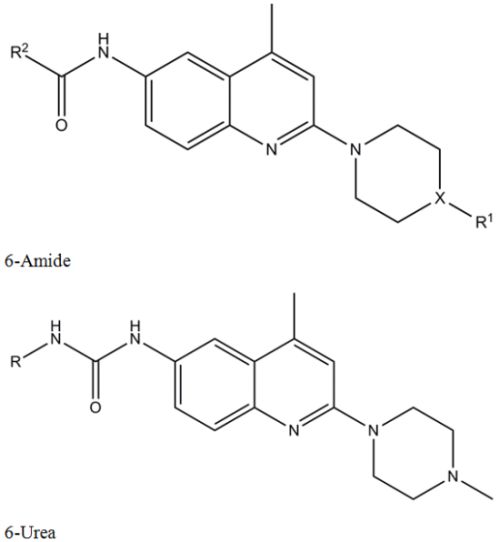{kind=link}
{kind=link}
{kind=link}
(a)  | |||
| Compound | R1 | X | R2 |
| 1 | ethyl | N | 4-tert-butylphenyl |
| 2 | ethyl | N | 4-butylphenyl |
| 3 | 2-pyrimidyl | N | 4-tert-butylphenyl |
| 4 | methyl | CH | 4-tert-butylphenyl |
| 5 | methyl | N | 4-butylphenyl |
| 6 | methyl | N | 4-tert-butylcyclohexyl |
| 7 | methyl | N | 5-chloro-2-methoxyphenyl |
| 8 | methyl | N | 2,4-dimethoxyphenyl |
| 9 | methyl | N | 4-tert-butylphenyl |
(b)  | |||
| Compound | R | ||
| 10 | 3-chlorophenyl | ||
| 11 | phenyl | ||
| 12 | 4-chlorophenyl | ||
| 13 | 3,5-dimethylphenyl | ||
| 14 | 3,4-dimethylphenyl | ||
| 15 | 4-ethylphenyl | ||
| 16 | 2-methylphenyl | ||
| 17 | 2-chlorophenyl | ||
| 18 | 4-bromophenyl | ||
2.2. Structure-Activity Relationship
| Compound | C. albicans MFC [a] (µM) | C. glabrata MFC [a] (µM) | C. albicans BEC50 [b] (µM) |
|---|---|---|---|
| 1 | 12.5 | 25 | 25 |
| 2 | 25 | >50 | 25 |
| 3 | >50 | >50 | >50 |
| 4 | >50 | >50 | >50 |
| 5 | 6.25 | 25 | 25 |
| 6 | 12.5 | >50 | 25 |
| 7 | 50 | >50 | >50 |
| 8 | 50 | >50 | >50 |
| 9 | 50 | >50 | >50 |
| 10 | 50 | >50 | 25 |
| 11 | >50 | >50 | >50 |
| 12 | >50 | >50 | 50 |
| 13 | >50 | >50 | 50 |
| 14 | 50 | >50 | >50 |
| 15 | >50 | >50 | 50 |
| 16 | >50 | >50 | >50 |
| 17 | 25 | >50 | >50 |
| 18 | >50 | >50 | >50 |
| AmB | <3.25 | <12.5 | <12.5 |
| Mico | >50 | >50 | 25 |
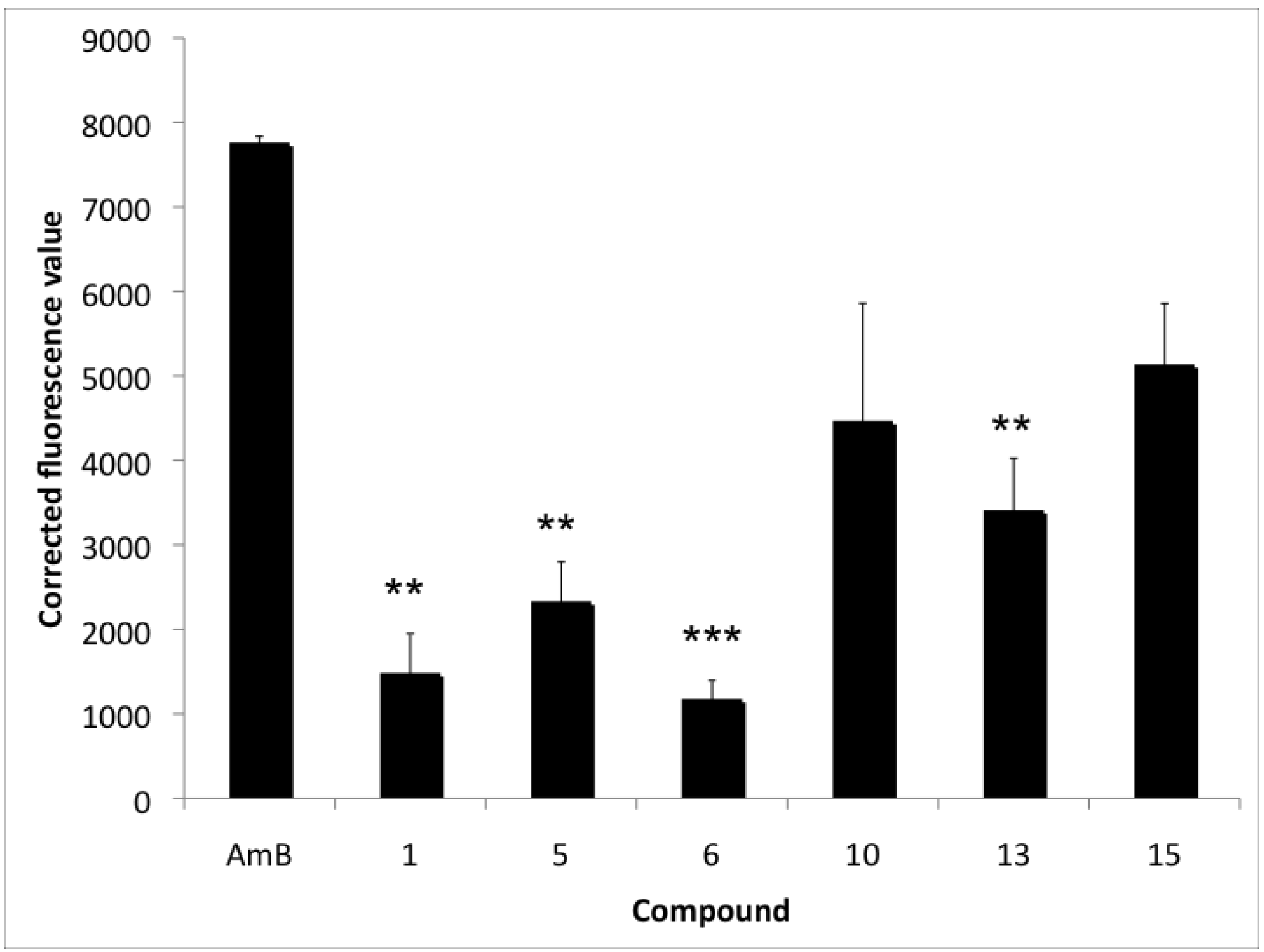
2.3. Accumulation of Endogenous ROS

3. Experimental
3.1. Chemical Compounds
3.2. Strains and Media
3.3. Fungicidal Activity
3.4. Antibiofilm Activity
3.5. Measurement of ROS Accumulation
4. Conclusions
Acknowledgments
- Sample Availability: Not available.
References
- Marr, K.A. Invasive Candida infections: The changing epidemiology. Oncology (Williston Park) 2004, 18, 9–14. [Google Scholar]
- Fanning, S.; Mitchell, A.P. Fungal biofilms. PLoS Pathog. 2012, 8, e1002585. [Google Scholar] [CrossRef]
- Ramage, G.; Mowat, E.; Jones, B.; Williams, C.; Lopez-Ribot, J. Our current understanding of fungal biofilms. Crit. Rev. Microbiol. 2009, 35, 340–355. [Google Scholar]
- Kuhn, D.M.; Ghannoum, M.A. Candida biofilms: Antifungal resistance and emerging therapeutic options. Curr. Opin. Investig. Drugs 2004, 5, 186–197. [Google Scholar]
- Klepser, M.E. Candida resistance and its clinical relevance. Pharmacotherapy 2006, 26, 68–75. [Google Scholar] [CrossRef]
- Chamilos, G.; Kontoyiannis, D.P. Update on antifungal drug resistance mechanisms of Aspergillus fumigatus. Drug Resist. Updat. 2005, 8, 344–358. [Google Scholar] [CrossRef]
- Odds, F.C. Genomics, molecular targets and the discovery of antifungal drugs. Rev. Iberoam. Micol. 2005, 22, 229–237. [Google Scholar] [CrossRef]
- Thevissen, K.; Hillaert, U.; Meert, E.M.; Chow, K.K.; Cammue, B.P.; Van Calenbergh, S.; Francois, I.E. Fungicidal activity of truncated analogues of dihydrosphingosine. Bioorg. Med. Chem. Lett. 2008, 18, 3728–3730. [Google Scholar]
- Vandenbosch, D.; Braeckmans, K.; Nelis, H.J.; Coenye, T. Fungicidal activity of miconazole against Candida spp. biofilms. J. Antimicrob. Chemother. 2010, 65, 694–700. [Google Scholar] [CrossRef]
- Al-Dhaheri, R.S.; Douglas, L.J. Apoptosis in Candida biofilms exposed to amphotericin B. J. Med. Microbiol. 2010, 59, 149–157. [Google Scholar] [CrossRef]
- Lanza, T.J.; Durette, P.L.; Rollins, T.; Siciliano, S.; Cianciarulo, D.N.; Kobayashi, S.V.; Caldwell, C.G.; Springer, M.S.; Hagmann, W.K. Substituted 4,6-diaminoquinolines as inhibitors of C5a receptor binding. J. Med. Chem. 1992, 35, 252–258. [Google Scholar] [CrossRef]
- Foley, M.; Tilley, L. Quinoline antimalarials: mechanisms of action and resistance and prospects for new agents. Pharmacol. Ther. 1998, 79, 55–87. [Google Scholar] [CrossRef]
- Taramelli, D.; Tognazioli, C.; Ravagnani, F.; Leopardi, O.; Giannulis, G.; Boelaert, J.R. Inhibition of intramacrophage growth of Penicillium marneffei by 4-aminoquinolines. Antimicrob. Agents Chemother. 2001, 45, 1450–1455. [Google Scholar] [CrossRef]
- Ulven, T.; Frimurer, T.M.; Receveur, J.M.; Little, P.B.; Rist, O.; Norregaard, P.K.; Hogberg, T. 6-Acylamino-2-aminoquinolines as potent melanin-concentrating hormone 1 receptor antagonists. Identification, structure-activity relationship, and investigation of binding mode. J. Med. Chem. 2005, 48, 5684–5697. [Google Scholar]
- Thevissen, K.; Marchand, A.; Chaltin, P.; Meert, E.M.; Cammue, B.P. Antifungal carbazoles. Curr. Med. Chem. 2009, 16, 2205–2211. [Google Scholar] [CrossRef]
- Kobayashi, D.; Kondo, K.; Uehara, N.; Otokozawa, S.; Tsuji, N.; Yagihashi, A.; Watanabe, N. Endogenous reactive oxygen species is an important mediator of miconazole antifungal effect. Antimicrob. Agents Chemother. 2002, 46, 3113–3117. [Google Scholar] [CrossRef]
- Thevissen, K.; Ayscough, K.R.; Aerts, A.M.; Du, W.; De Brucker, K.; Meert, E.M.; Ausma, J.; Borgers, M.; Cammue, B.P.; Francois, I.E. Miconazole induces changes in actin cytoskeleton prior to reactive oxygen species induction in yeast. J. Biol. Chem. 2007, 282, 21592–21597. [Google Scholar]
- Francois, I.E.; Thevissen, K.; Pellens, K.; Meert, E.M.; Heeres, J.; Freyne, E.; Coesemans, E.; Viellevoye, M.; Deroose, F.; Martinez Gonzalez, S.; et al. Design and synthesis of a series of piperazine-1-carboxamidine derivatives with antifungal activity resulting from accumulation of endogenous reactive oxygen species. ChemMedChem 2009, 4, 1714–1721. [Google Scholar] [CrossRef]
- Fonzi, W.A.; Irwin, M.Y. Isogenic strain construction and gene mapping in Candida albicans. Genetics 1993, 134, 717–728. [Google Scholar]
- Kaur, R.; Ma, B.; Cormack, B.P. A family of glycosylphosphatidylinositol-linked aspartyl proteases is required for virulence of Candida glabrata. Proc. Natl. Acad. Sci. USA 2007, 104, 7628–7633. [Google Scholar]
- Aerts, A.M.; Carmona-Gutierrez, D.; Lefevre, S.; Govaert, G.; Francois, I.E.; Madeo, F.; Santos, R.; Cammue, B.P.; Thevissen, K. The antifungal plant defensin RsAFP2 from radish induces apoptosis in a metacaspase independent way in Candida albicans. FEBS Lett. 2009, 583, 2513–2516. [Google Scholar] [CrossRef]
- Bink, A.; Vandenbosch, D.; Coenye, T.; Nelis, H.; Cammue, B.P.; Thevissen, K. Superoxide dismutases are involved in Candida albicans biofilm persistence against miconazole. Antimicrob. Agents Chemother. 2011, 55, 4033–4037. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Delattin, N.; Bardiot, D.; Marchand, A.; Chaltin, P.; De Brucker, K.; Cammue, B.P.A.; Thevissen, K. Identification of Fungicidal 2,6-Disubstituted Quinolines with Activity against Candida Biofilms. Molecules 2012, 17, 12243-12251. https://doi.org/10.3390/molecules171012243
Delattin N, Bardiot D, Marchand A, Chaltin P, De Brucker K, Cammue BPA, Thevissen K. Identification of Fungicidal 2,6-Disubstituted Quinolines with Activity against Candida Biofilms. Molecules. 2012; 17(10):12243-12251. https://doi.org/10.3390/molecules171012243
Chicago/Turabian StyleDelattin, Nicolas, Dorothée Bardiot, Arnaud Marchand, Patrick Chaltin, Katrijn De Brucker, Bruno P. A. Cammue, and Karin Thevissen. 2012. "Identification of Fungicidal 2,6-Disubstituted Quinolines with Activity against Candida Biofilms" Molecules 17, no. 10: 12243-12251. https://doi.org/10.3390/molecules171012243
APA StyleDelattin, N., Bardiot, D., Marchand, A., Chaltin, P., De Brucker, K., Cammue, B. P. A., & Thevissen, K. (2012). Identification of Fungicidal 2,6-Disubstituted Quinolines with Activity against Candida Biofilms. Molecules, 17(10), 12243-12251. https://doi.org/10.3390/molecules171012243