Comparison of Cocoa Beans from China, Indonesia and Papua New Guinea

Abstract
:1. Introduction
2. Experimental Section
2.1. Materials
2.2. Measurement of Average Bean Weight and Fat
2.3. Measurement of Color and Absorbance at 420 nm
2.4. Measurement of Total Phenolic Content and Total Flavonoid Content
2.5. Measurement of Gallic Acid (GA) and Epicatechin (EC) Content
2.6. Measurement of Total Amino Acids
2.7. Measurement of Free Amino Acids
2.8. Measurement of Flavor with Electronic Nose (E-Nose)
2.9. Statistical Analysis
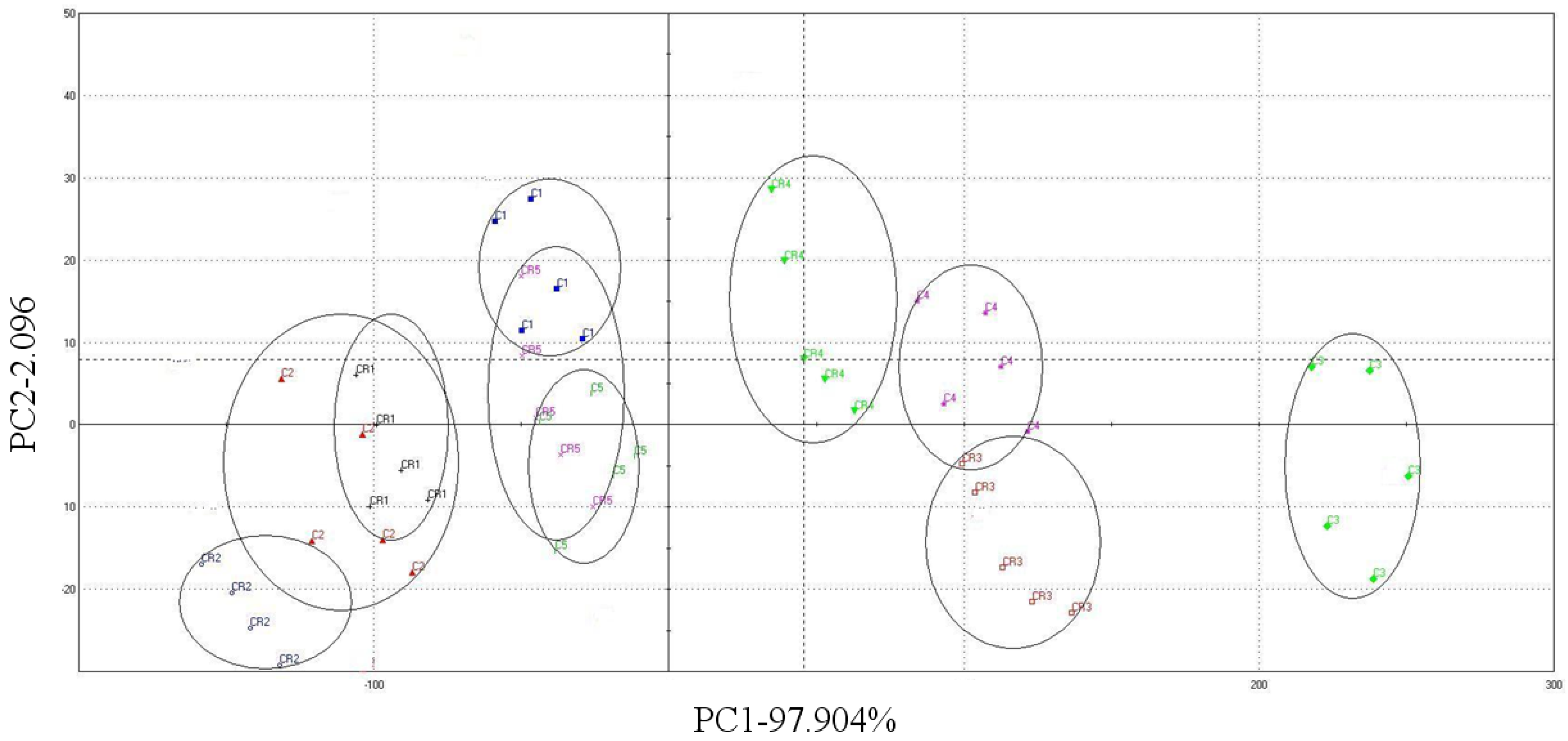
3. Results and Discussion
3.1. Average Bean Weight and Butter Content

{kind=link}
{kind=link}
{kind=link}
| Samples | Gross weight/g | Net weight/g | Butter content (%) |
|---|---|---|---|
| C1 | 1.2124 a,b,c ± 0.0572 | 1.0757 a,b,c ± 0.0346 | 45.86 a,c,d ± 5.42 |
| C2 | 1.3000 a,b,c ± 0.0143 | 1.1115 a,b,c ± 0.0261 | 53.67 a ± 4.81 |
| C3 | 1.3946 b ± 0.0049 | 1.1713 b ± 0.3148 | 49.85 a,b,d ± 3.18 |
| C4 | 1.1020 a,c ± 0.0997 | 1.0980 a,c ± 0.1582 | 39.24 c,d ± 4.25 |
| C5 | 1.0225 c ± 0.0741 | 1.0062 c ± 0.2653 | 43.44 b,c,d ± 5.13 |
3.2. Color and Absorbance at 420 nm
| Samples | A 420 nm | L* | a* | b* |
|---|---|---|---|---|
| C1 | 0.053 ± 0.021 | 42.22 a,b ± 0.243 | 5.240 a,b,c ± 1.165 | 3.327 a ± 1.183 |
| C2 | 0.058 ± 0.038 | 45.93 ± 1.043 | 6.793 c ± 0.902 | 7.420 d ± 1.121 |
| C3 | 0.887 a ± 0.022 | 47.42 c ± 0.904 | 4.940 a,b ± 0.242 | 6.783 c,d ± 0.193 |
| C4 | 0.161 b ± 0.005 | 41.30 a ± 0.667 | 3.787 a ± 0.191 | 3.327 a ± 0.424 |
| C5 | 0.195 c ± 0.005 | 41.52 a ± 1.187 | 4.633 a ± 1.166 | 2.323 a ± 1.020 |
| CR1 | 0.217 d ± 0.002 | 43.59 b ± 0.621 | 6.393 c,b ± 0.263 | 5.367 b,c ± 0.194 |
| CR2 | 0.218 d ± 0.004 | 43.67 b ± 0.501 | 7.457 ± 0.410 | 6.943 c,d ± 0.611 |
| CR3 | 0.087 e ± 0.003 | 49.31 d ± 1.165 | 6.807 c ± 0.564 | 9.357 ± 1.310 |
| CR4 | 0.048 ± 0.001 | 41.17 a ± 0.943 | 4.830 a,b ± 0.327 | 4.153 a,b ± 0.468 |
| CR5 | 0.144 b ± 0.004 | 40.59 a ± 0.861 | 5.363 a,b,c ± 0.631 | 3.835 a,b ± 0.190 |
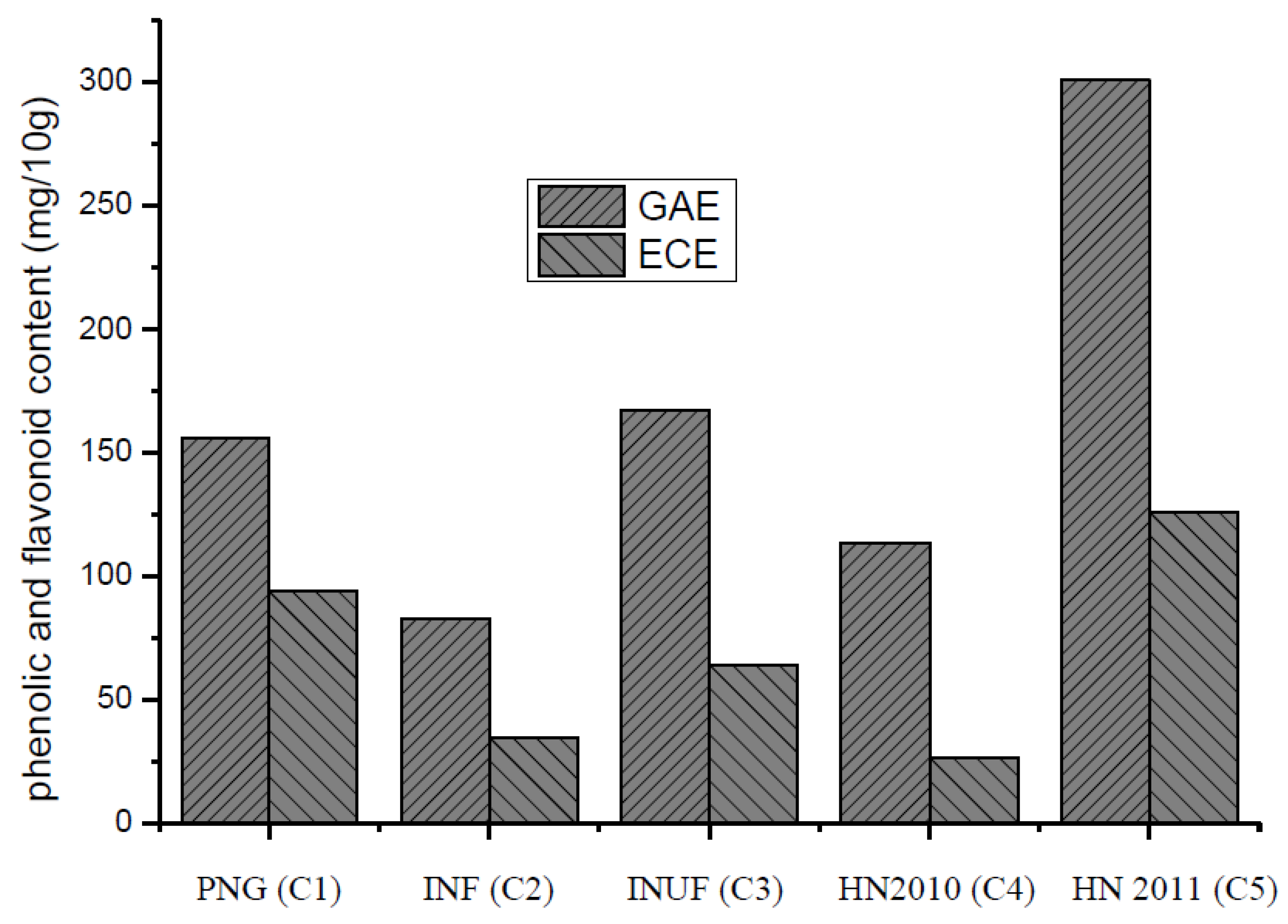
3.3. Total Phenolic Content and Total Flavonoid Content
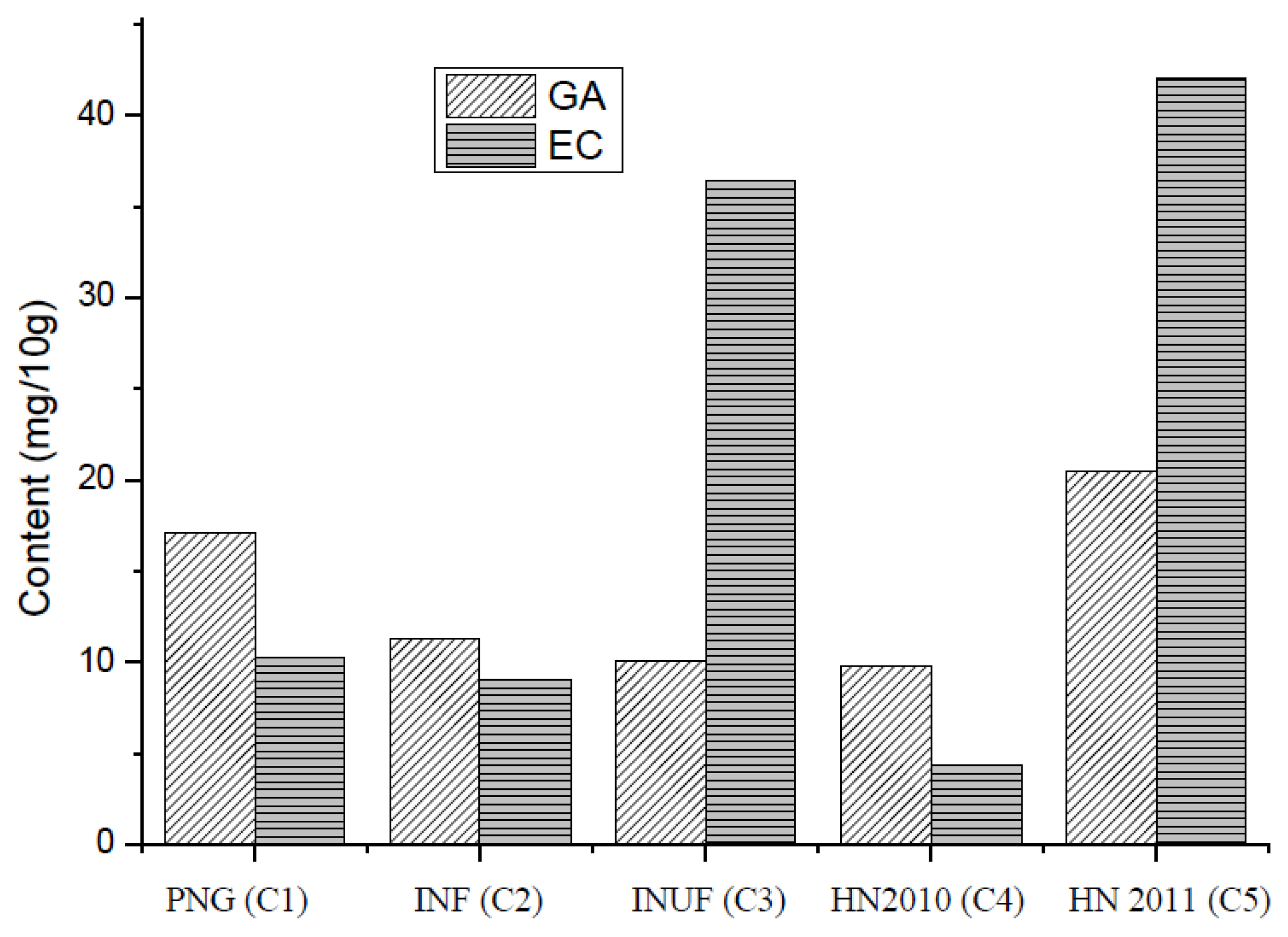
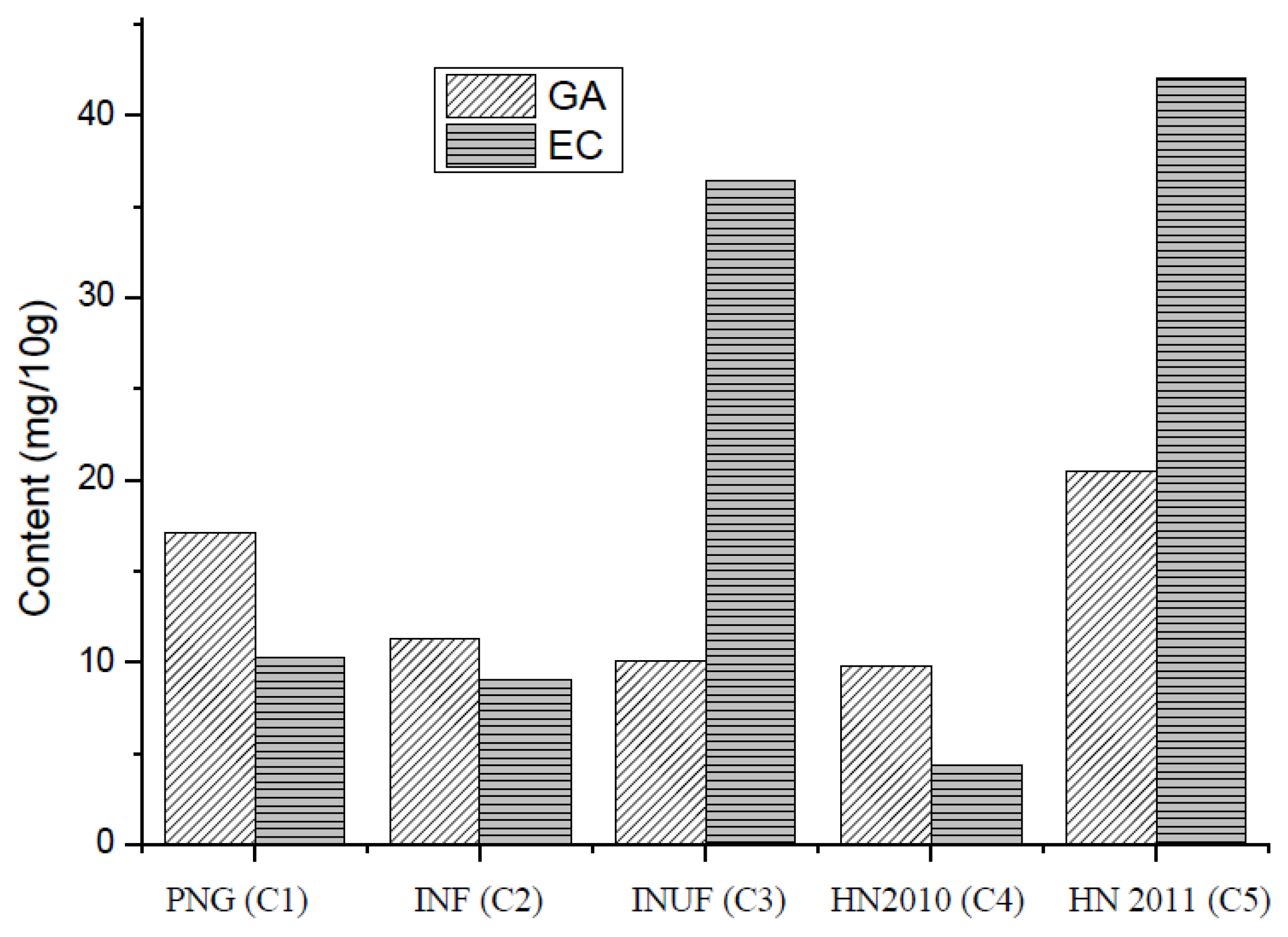
3.4. Gallic Acid (GC) and Epicatechin (EA)
3.5. Total and Free Amino Acids
| Amino acids | C1 (g/100 g) | C2 (g/100 g) | C3 (g/100 g) | C4 (g/100 g) | C5 (g/100 g) |
|---|---|---|---|---|---|
| asp | 1.48234112 | 1.248726 | 2.03329 | 1.638016 | 2.285384 |
| glu | 2.76421738 | 2.33606 | 3.61903 | 2.745753 | 3.024123 |
| ser | 0.66305973 | 0.567914 | 0.928503 | 0.797482 | 0.934248 |
| his | 0.27536076 | 0.231274 | 0.392731 | 0.290148 | 0.342329 |
| gly | 0.67713588 | 0.553377 | 0.85647 | 0.71582 | 0.873638 |
| thr | 0.58388107 | 0.515058 | 0.778265 | 0.684044 | 0.812028 |
| arg | 0.96300469 | 0.884542 | 1.344468 | 1.123294 | 1.302416 |
| ala | 0.72384247 | 0.549737 | 0.911206 | 0.675514 | 0.82077 |
| tyr | 0.40220555 | 0.376532 | 0.569141 | 0.481996 | 0.507722 |
| cys-s | 0.09059641 | 0.103466 | 0.134149 | 0.14175 | 0.144297 |
| val | 0.88030156 | 0.772049 | 1.147787 | 0.929087 | 1.15285 |
| met | 0.16747286 | 0.069999 | 0.204375 | 0.1672 | 0.213946 |
| phe | 0.75009401 | 0.659956 | 1.148313 | 0.837903 | 0.958442 |
| ile | 0.56001189 | 0.503279 | 0.714016 | 0.610186 | 0.740686 |
| leu | 0.91887279 | 0.80002 | 1.254947 | 1.036056 | 1.242061 |
| lys | 0.6899189 | 0.685431 | 1.130527 | 0.802536 | 1.032648 |
| pro | 0.85167766 | 0.723573 | 1.004048 | 0.837604 | 0.858958 |
| Total | 13.4439947 | 11.58099 | 18.17126 | 14.51439 | 17.24655 |
| Amino acids | C1 (g/100 g) | C2 (g/100 g) | C3 (g/100 g) | C4 (g/100 g) | C5 (g/100 g) |
|---|---|---|---|---|---|
| asp | 0.06175624 | 0.052118 | 0.0595 | 0.03942 | 0.080142 |
| glu | 0.0616727 | 0.061372 | 0.076922 | 0.034869 | 0.064857 |
| ser | 0.00145676 | 0.002074 | 0.000417 | 0.000607 | 0.001391 |
| his | 0.00334901 | 0.003545 | 0.006681 | 0.002198 | 0.012053 |
| gly | 0.00981977 | 0.007367 | 0.017332 | 0.006294 | 0.023166 |
| thr | 0.01156708 | 0.011032 | 0.013705 | 0.008447 | 0.022726 |
| arg | 0.03857273 | 0.061132 | 0.017908 | 0.025347 | 0.095447 |
| ala | 0.11614724 | 0.073076 | 0.079882 | 0.047004 | 0.112538 |
| tyr | 0.04184931 | 0.0555 | 0.034151 | 0.029755 | 0.077396 |
| cys-s | 0.00110326 | 0.001001 | 1.25E-07 | 0.001071 | 0.001652 |
| val | 0.07068854 | 0.064691 | 0.054895 | 0.052798 | 0.11922 |
| met | 0.0015651 | 0.002192 | 0.000395 | 0.000734 | 0.003633 |
| phe | 0.06475303 | 0.083083 | 0.020958 | 0.050539 | 0.146415 |
| ile | 0.03503073 | 0.034275 | 0.019892 | 0.029651 | 0.073133 |
| leu | 0.09972465 | 0.117406 | 0.036901 | 0.061232 | 0.168642 |
| lys | 0.02884247 | 0.04466 | 0.018216 | 0.017223 | 0.051872 |
| pro | 0.05347026 | 0.043259 | 0.055476 | 0.048206 | 0.102607 |
| Total | 0.70136889 | 0.717783 | 0.513232 | 0.455394 | 1.15689 |
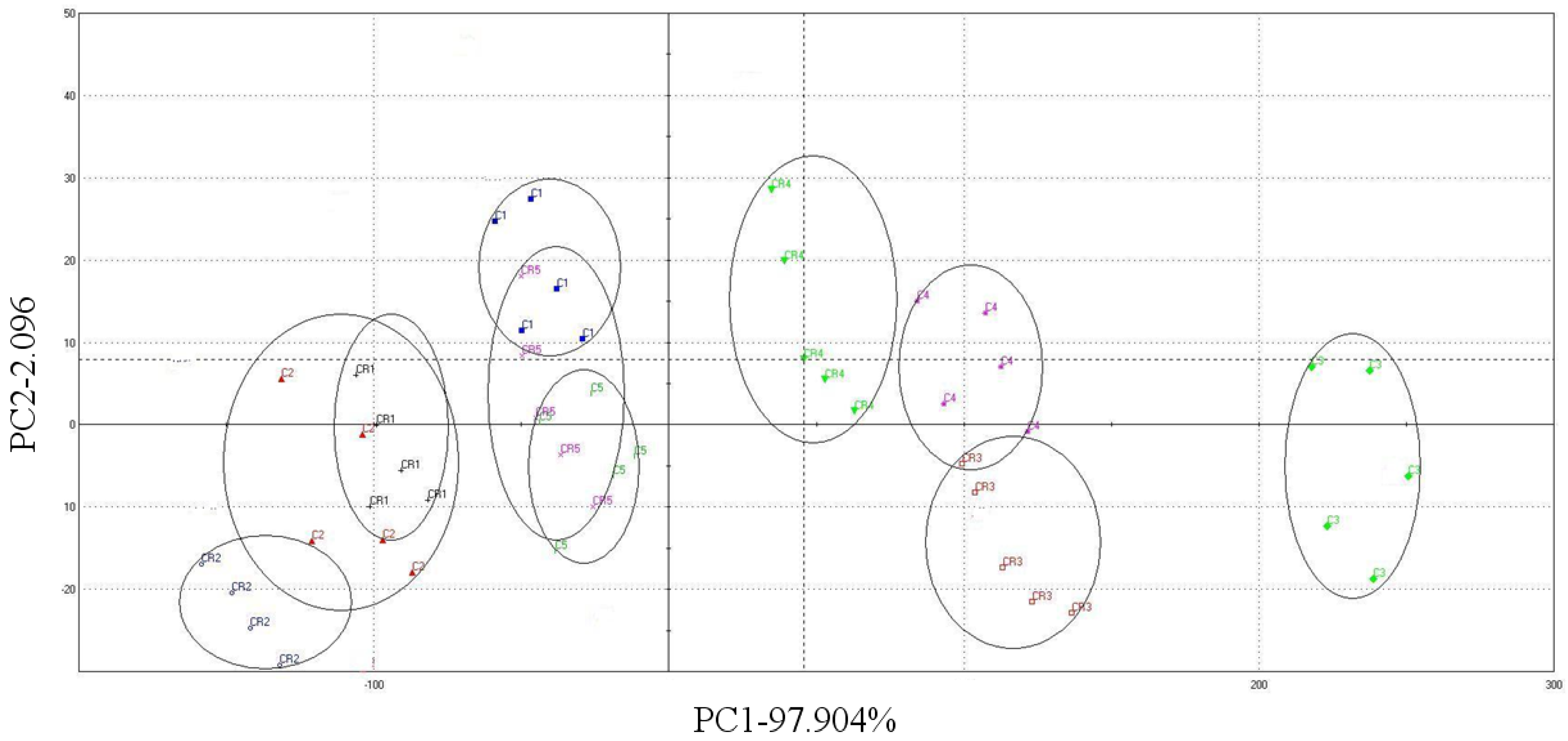
3.6. Flavor with Electronic Nose (E-nose)
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Tafuri, A.; Ferracane, R.; Ritieni, A. Ochratoxin A in Italian marketed cocoa products. Food Chem. 2004, 88, 487–494. [Google Scholar] [CrossRef]
- Thompson, S.S.; Miller, K.B.; Lopez, A.S. Cocoa and Coffee. In Food Microbiology—Fundamentals and Frontiers; Doyle, M.J., Beuchat, L.R., Montville, T.J., Eds.; ASM Press: Washington, DC, USA, 2001; pp. 721–733. [Google Scholar]
- Lopez, A.S.; Dimick, P.S. Cocoa Fermentation. In Enzymes, Biomass, Food and Feed; Reed, G., Nagodawithana, T.W., Eds.; VCH: Weinheim, Germany, 1995; pp. 561–577. [Google Scholar]
- Ardhana, M.M.; Fleet, H.G. The microbial ecology of cocoa bean fermentations in Indonesia. Int. J. Food Microbiol. 2003, 86, 87–99. [Google Scholar] [CrossRef]
- LMC International, Anonymous. In The World Cocoa Market Outlook; LMC International Ltd.: New York, NY, USA, 2001; pp. 1–29.
- Hii, C.L.; Law, C.L.; Cloke, M.; Suzannah, S. Thin layer drying kinetics of cocoa and dried product quality. Biosyst. Eng. 2009, 102, 153–161. [Google Scholar] [CrossRef]
- Biehl, B.; Voigt, J. Biochemistry of Chocolate Flavour Precursors. In Proceedings of the 12th International Cocoa Conference, Salvador, Brazil, 17–23 November 1996.
- Puziah, H.; Jinap, S.; Sharifah, K.S.M.; Asbi, A. Changes in free amino acids, peptide-N, sugar and pyrazine concentration during cocoa fermentation. J. Sci. Food Agric. 1998, 78, 535–542. [Google Scholar] [CrossRef]
- Kratzer, U.; Frank, R.; Kalbacher, H.; Biehl, B.; Wöstemeyer, J.; Voigt, J. Subunit structure of the vicilin-like globular storage protein of cocoa seeds and the origin of cocoa- and chocolate-specific aroma precursors. Food Chem. 2009, 113, 903–913. [Google Scholar] [CrossRef]
- Jinap, M.; Nazamid, S.; Jamilah, B. Activation of remaining key enzymes in dried under-fermented cocoa beans and its effect on aroma precursor formation. Food Chem. 2002, 78, 407–417. [Google Scholar] [CrossRef]
- Hammami, C.; Rene, F.; Marin, M. Processquality optimization of the vacuum freeze-drying of apple slice by the response surface method. Int. J. Food Sci. Technol. 1999, 34, 145–160. [Google Scholar] [CrossRef]
- Lee, W.K.; Kim, J.Y.; Lee, J.H.; Lee, Y.C. Cocoa has more phenolic phytochemicals and a higher antioxidant capacity than teas and red wine. J. Agric. Food Chem. 2003, 51, 7292–7295. [Google Scholar] [CrossRef]
- Jia, Z.S.; Tang, M.C.; Wu, J.M. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Kyi, M.T.; Daud, W.W.R.; Mohammad, B.M.A.; Samsudin, W.; Kadhum, A.A.H.; Talib, M.Z.M. The kinetics of polyphenol degradation during the drying of Malaysian cocoa beans. Int. J. Food Sci. Technol. 2005, 40, 323–331. [Google Scholar] [CrossRef]
- Noor-Soffalina, S.S.; Jinap, S.; Nazamid, S.; Nazimah, S.A.H. Effect of polyphenol and pH on cocoa Maillard-related flavour precursors in a lipidic model system. Int. J. Food Sci. Technol. 2009, 44, 168–180. [Google Scholar] [CrossRef]
- Zak, D.L.; Keeney, P.G. Changes in cocoa proteins during ripening of fruit, fermentation, and further processing of cocoa beans. J. Agric. Food Chem. 1976, 24, 483–486. [Google Scholar] [CrossRef]
- Afoakwa, O.E. Chocolate Science and Technology; Wiley-Blackwell: Oxford, UK, 2010; pp. 110–156. [Google Scholar]
- Beckett, T.S. Industrial Chocolate Manufacture and Use, 4th ed; Wiley-Blackwell: Oxford, UK, 2009; pp. 1–10. [Google Scholar]
- Nazaruddin, R.; Seng, L.K.; Hassan, O.; Said, M. Effect of pulp preconditioning on the content of polyphenols in cocoa beans (Theobroma cacao) during fermentation. Ind. Crops Prod. 2006, 24, 87–94. [Google Scholar] [CrossRef]
- Niemenak, N.; Rohsius, C.; Elwers, S.; Ndoumoua, D.O.; Lieberei, R. Comparative study of different cocoa (Theobroma cacao L.) clones in terms of their phenolics and anthocyanins contents. J. Food Compos. Anal. 2006, 19, 612–619. [Google Scholar]
- Camu, N.; Winter, D.T.; Addo, K.S.; Takrama, S.J.; Bernaert, H.; Vuyst, D.L. Fermentation of cocoa beans: Influence of microbial activities and polyphenol concentrations on the flavour of chocolate. J. Sci. Food Agric. 2008, 88, 2288–2297. [Google Scholar] [CrossRef]
- Redovniković, I.R.; Delonga, K.; Mazor, S.; Dragović-Uzelac, V.; Carić, M.; Vorkapić-Furač, J. Polyphenolic content and composition and antioxidative activity of different cocoa liquors. Czech J. Food Sci. 2009, 27, 330–337. [Google Scholar]
- Ramli, N.; Hassan, O.; Said, M.; Samsudin, W.; Idris, A.N. Influence of roasting conditions on volatile flavor of roasted Malaysian cocoa beans. J. Food Proc. Preserv. 2006, 30, 280–298. [Google Scholar] [CrossRef]
- Tomas-Barberán, F.A.; Cienfuegos-Jovellanos, E.; Marín, A.; Muguerza, B.; Gil-Izquierdo, A.; Cerdá, B.; Zafrilla, P.; Morillas, J.; Mulero, J.; Ibarra, A.; et al. A new process to develop a cocoa powder with higher flavonoid monomer content and enhanced bioavailability in healthy humans. J. Agr. Food Chem. 2007, 55, 3926–3935. [Google Scholar] [CrossRef]
- Miller, B.K.; Hurst, W.J.; Flannigan, N.; Gan, L.N.; Ou, B.X.; Lee, C.Y.; Smith, N.; Stuart, A.D. Survey of commercially available chocolate- and cocoa-containing products in the United States. 2. Comparison of flavan-3-ol content with nonfat cocoa solids, total polyphenols, and percent cacao. J. Agric. Food Chem. 2009, 57, 9169–9180. [Google Scholar]
- Hans-Dieter, W. Discrimination of chocolates and packaging materials by an electronic nose. Eur. Food Res. Technol. 2001, 212, 529–533. [Google Scholar] [CrossRef]
- Schwan, R.F.; Wheals, A.E. The microbiology of cocoa fermentation and its role in chocolate quality. Crit. Rev. Food Sci. Nutr. 2004, 44, 205–221. [Google Scholar] [CrossRef]
- De Brito, E.S.; Pezoa García, N.H.; Gallao, M.I.; Cortelazzo, A.L.; Fevereiro, P.S.; Braga, M.R. Structural and chemical changes in cocoa (Theobroma cacao L) during fermentation, drying and roasting. J. Sci. Food Agric. 2000, 81, 281–288. [Google Scholar]
- Hansen, C.E.; del Olmo, M.; Burri, C. Enzyme activities in cocoa beans during fermentation. J. Sci. Food Agric. 1998, 77, 273–281. [Google Scholar] [CrossRef]
- Jinap, M.; Jamilah, B.; Nazamid, S. Effects of incubation and polyphenol oxidase enrichment on colour, fermentation index, procyanidins and astringency of unfermented and partly fermented cocoa beans. Int. J. Food Sci. Technol. 2003, 38, 285–295. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gu, F.; Tan, L.; Wu, H.; Fang, Y.; Xu, F.; Chu, Z.; Wang, Q. Comparison of Cocoa Beans from China, Indonesia and Papua New Guinea. Foods 2013, 2, 183-197. https://doi.org/10.3390/foods2020183
Gu F, Tan L, Wu H, Fang Y, Xu F, Chu Z, Wang Q. Comparison of Cocoa Beans from China, Indonesia and Papua New Guinea. Foods. 2013; 2(2):183-197. https://doi.org/10.3390/foods2020183
Chicago/Turabian StyleGu, Fenglin, Lehe Tan, Huasong Wu, Yiming Fang, Fei Xu, Zhong Chu, and Qinghuang Wang. 2013. "Comparison of Cocoa Beans from China, Indonesia and Papua New Guinea" Foods 2, no. 2: 183-197. https://doi.org/10.3390/foods2020183