Effects of Light and Wounding on Jasmonates in Rice phyAphyC Mutants


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
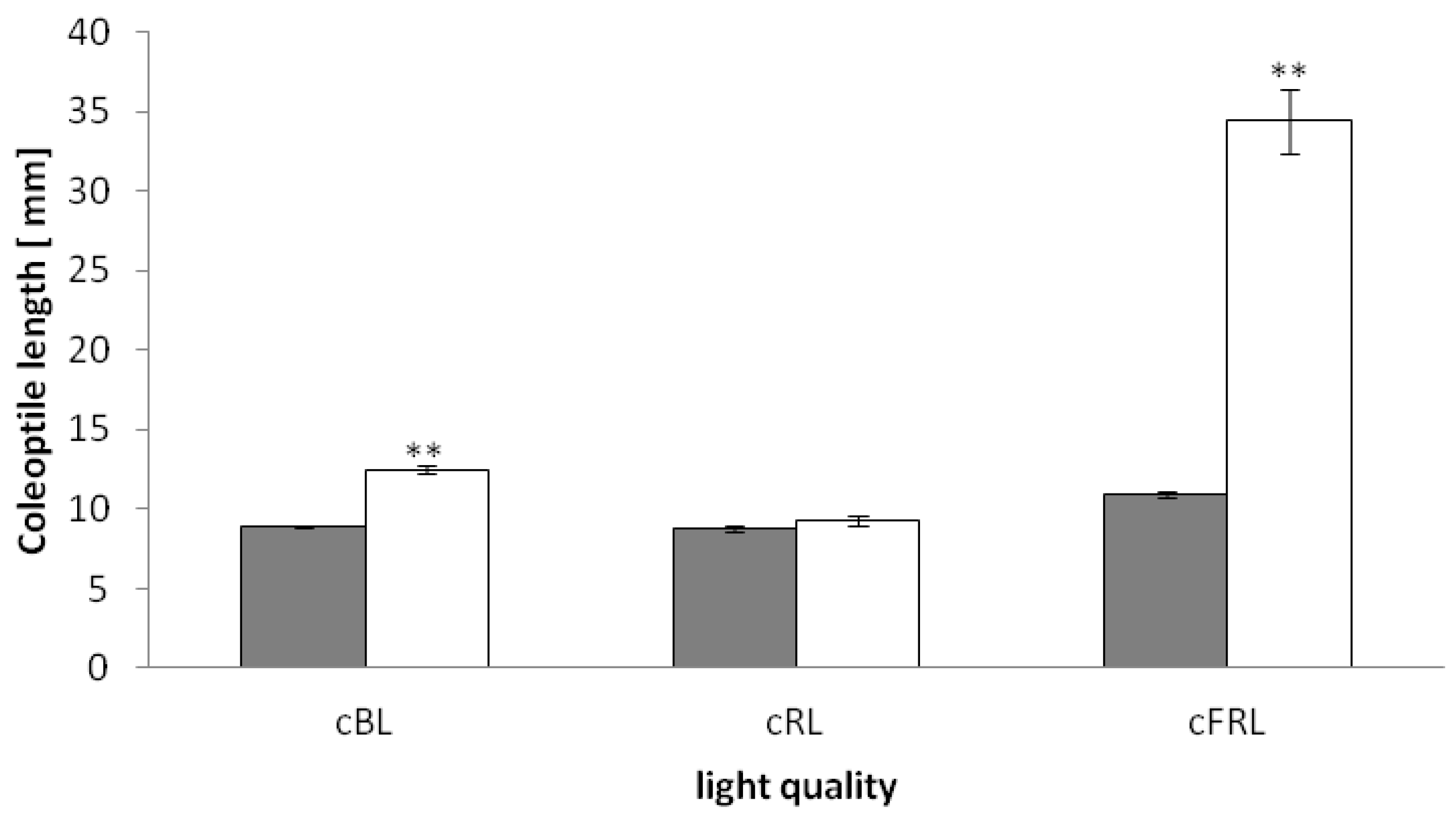
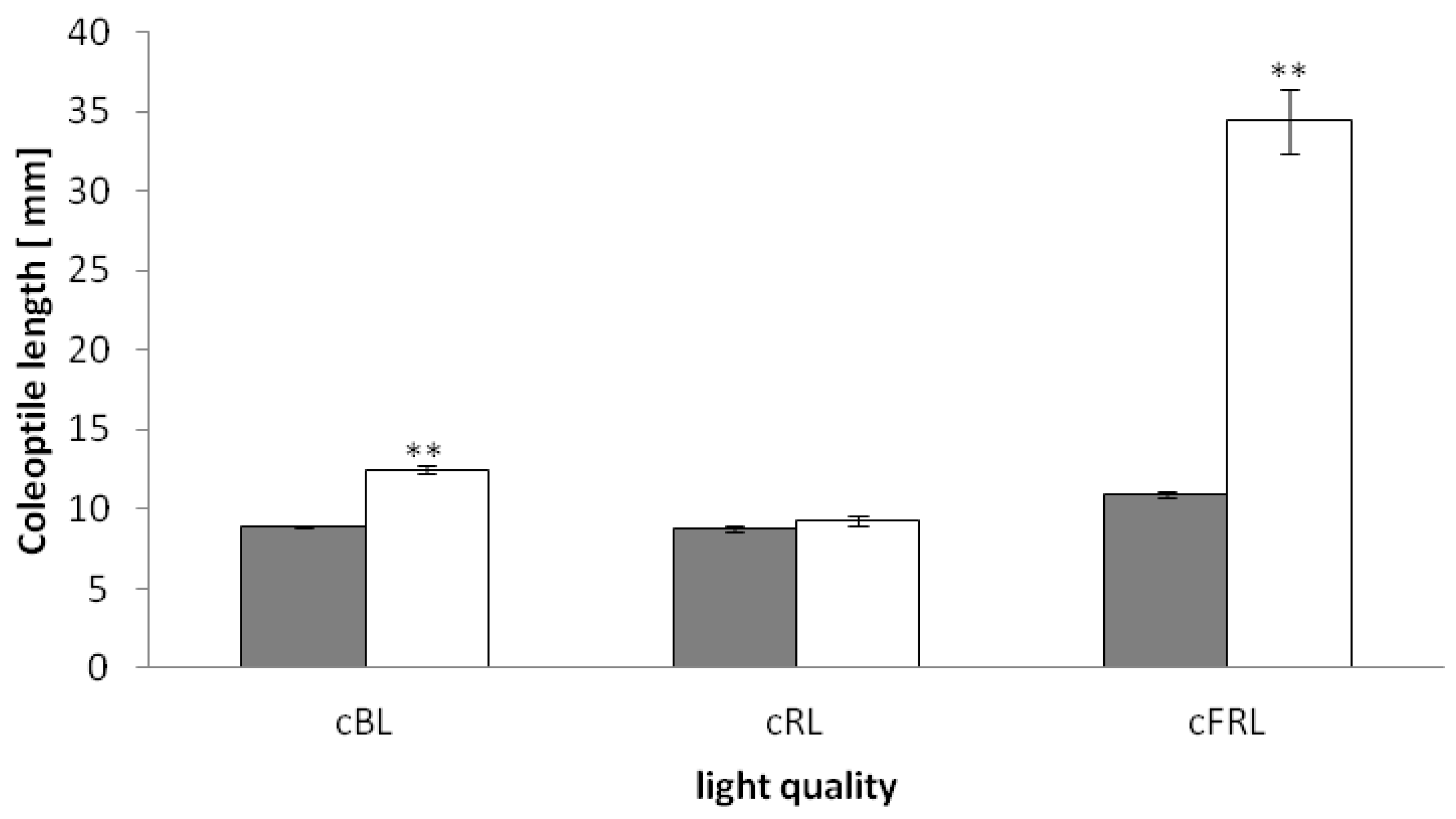
2.1. The phyAphyC Mutant Displays a Phenotype in Response to Continuous Blue and Far-Red Light
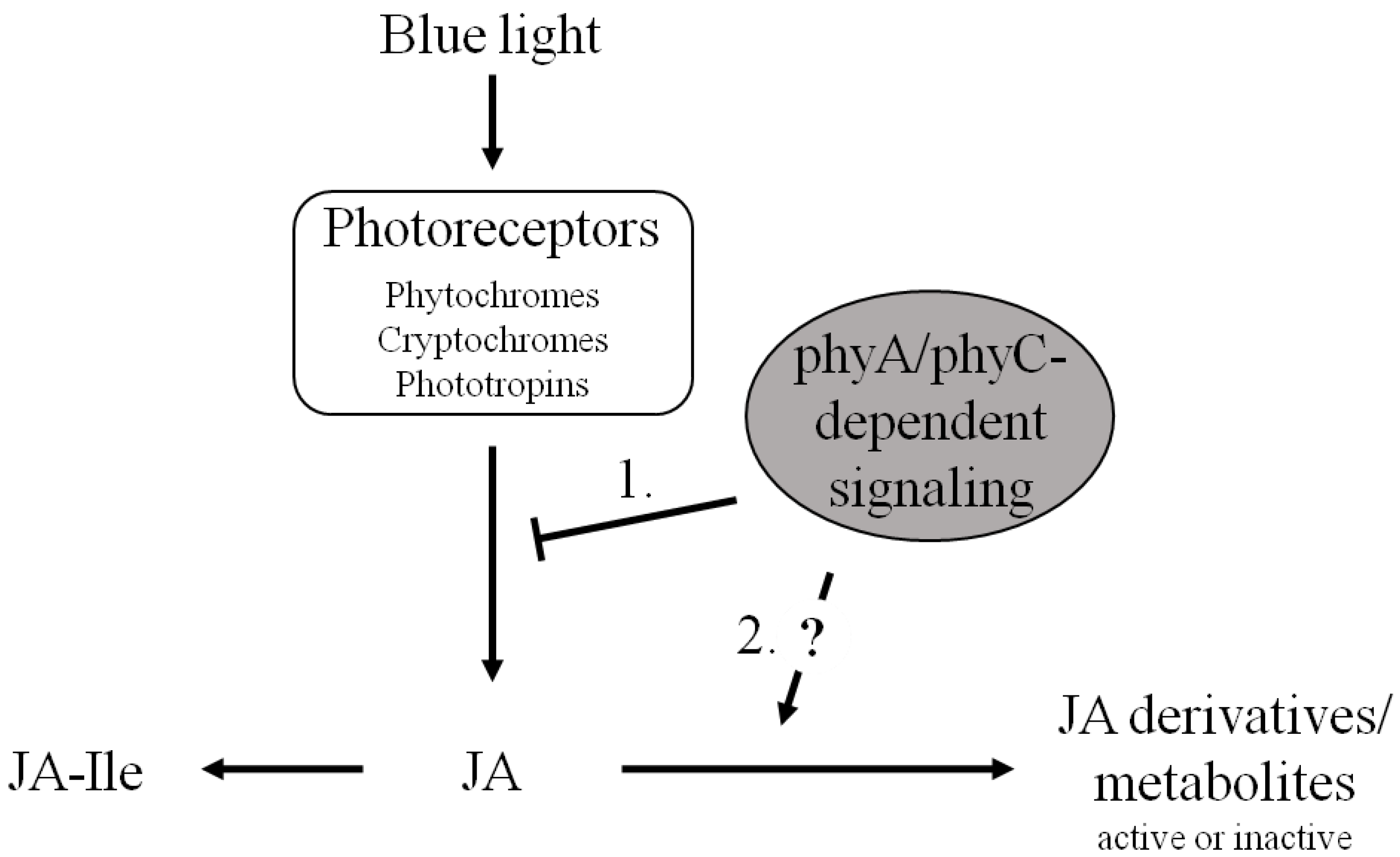
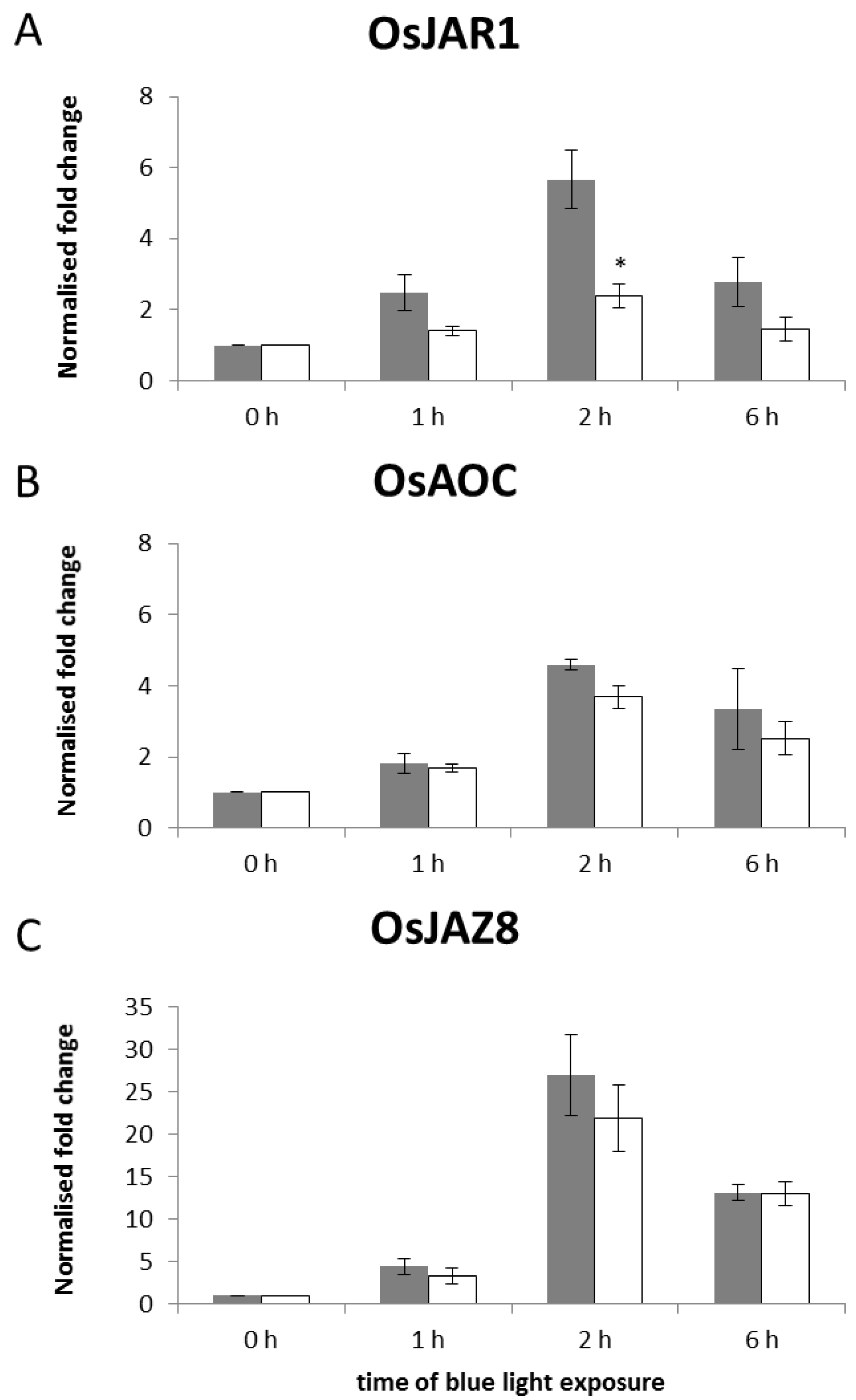
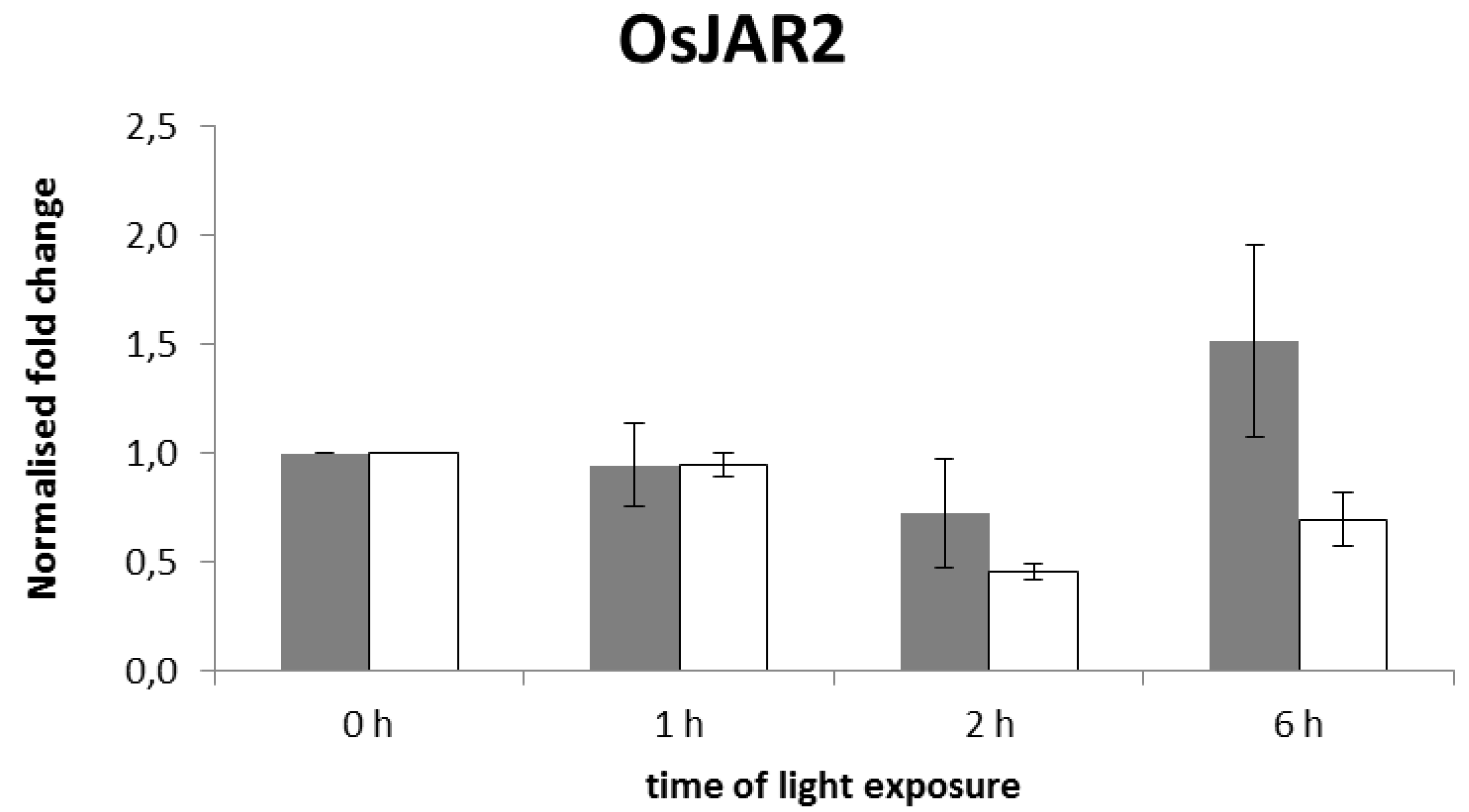
2.2. The Blue-Light Response of OsJAR1 Is Specifically Suppressed in phyAphyC
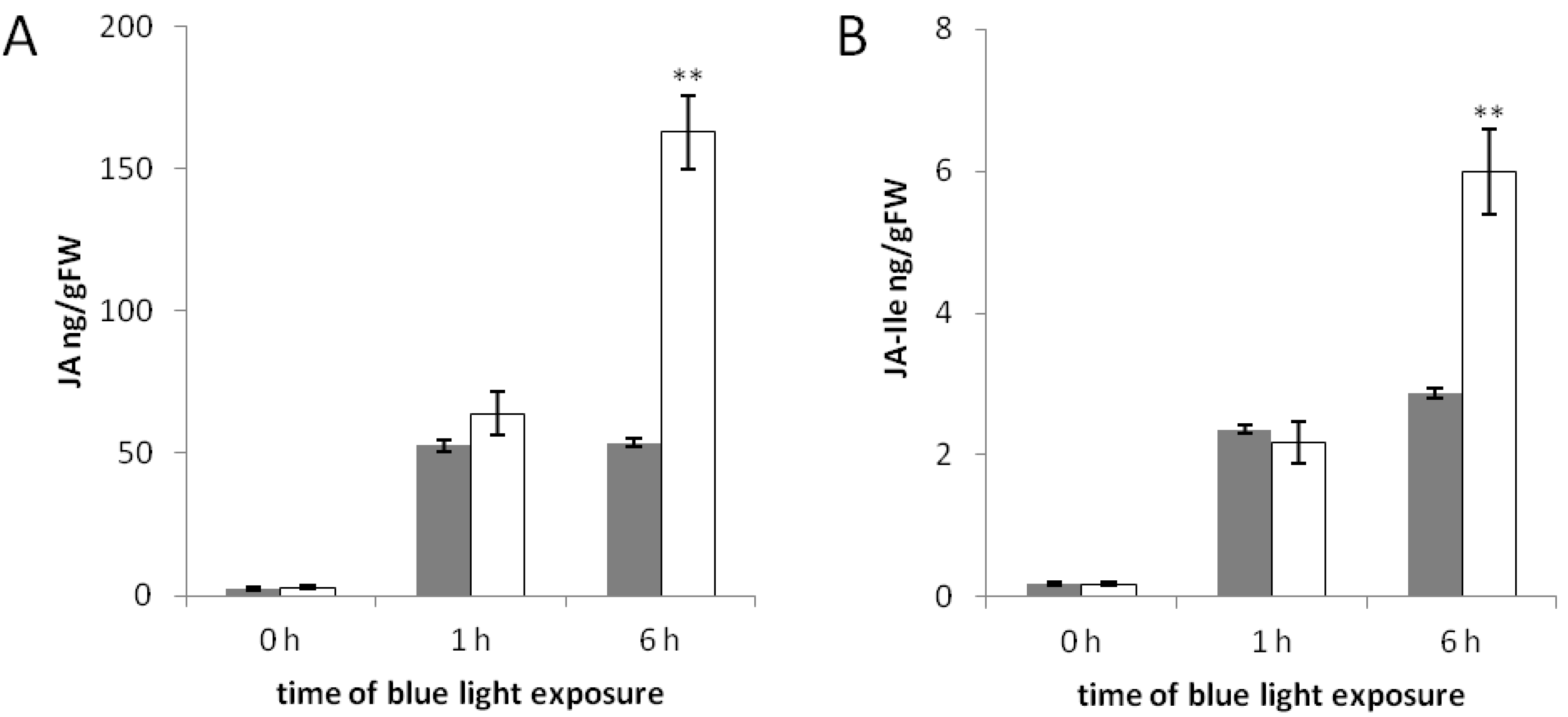
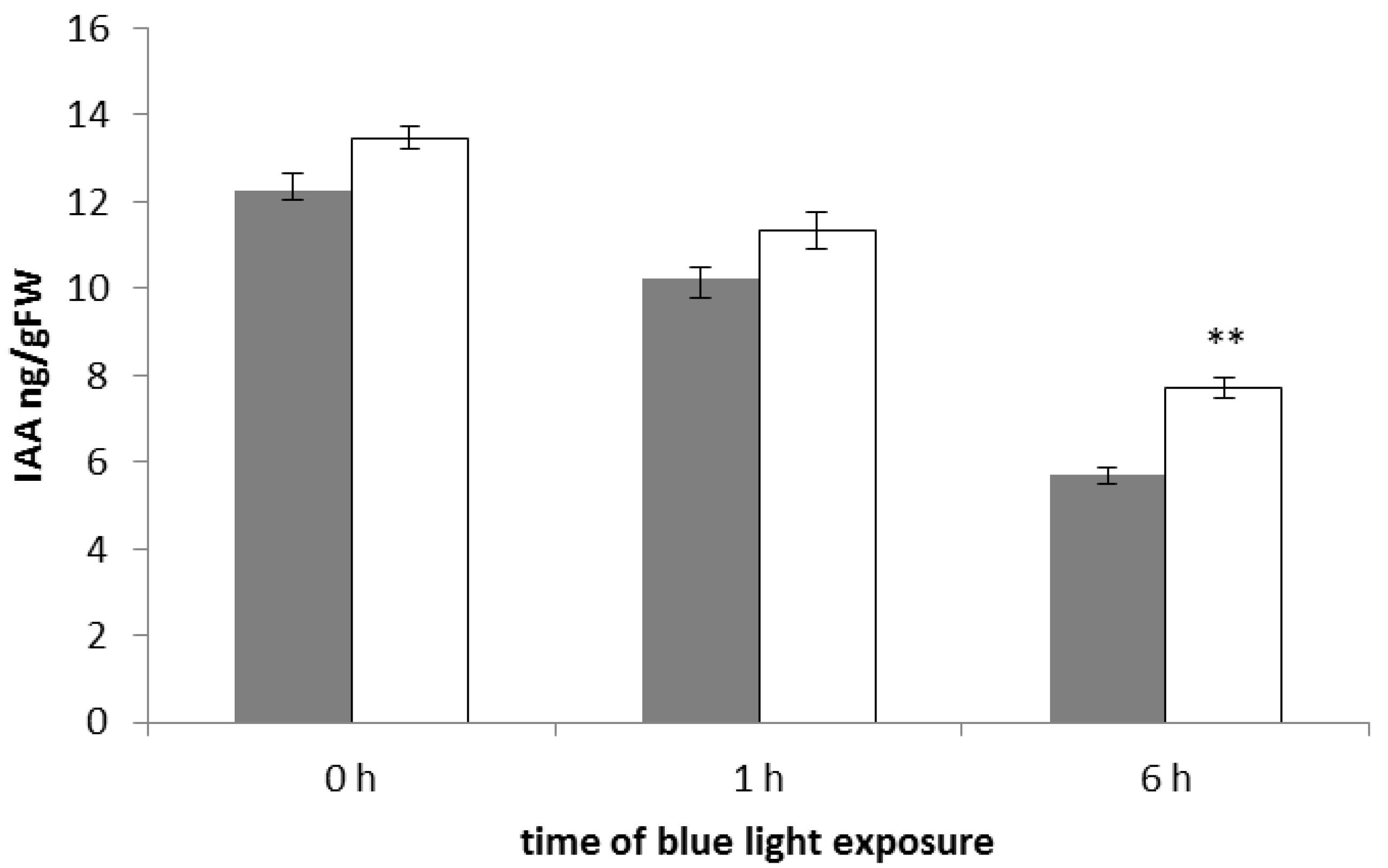
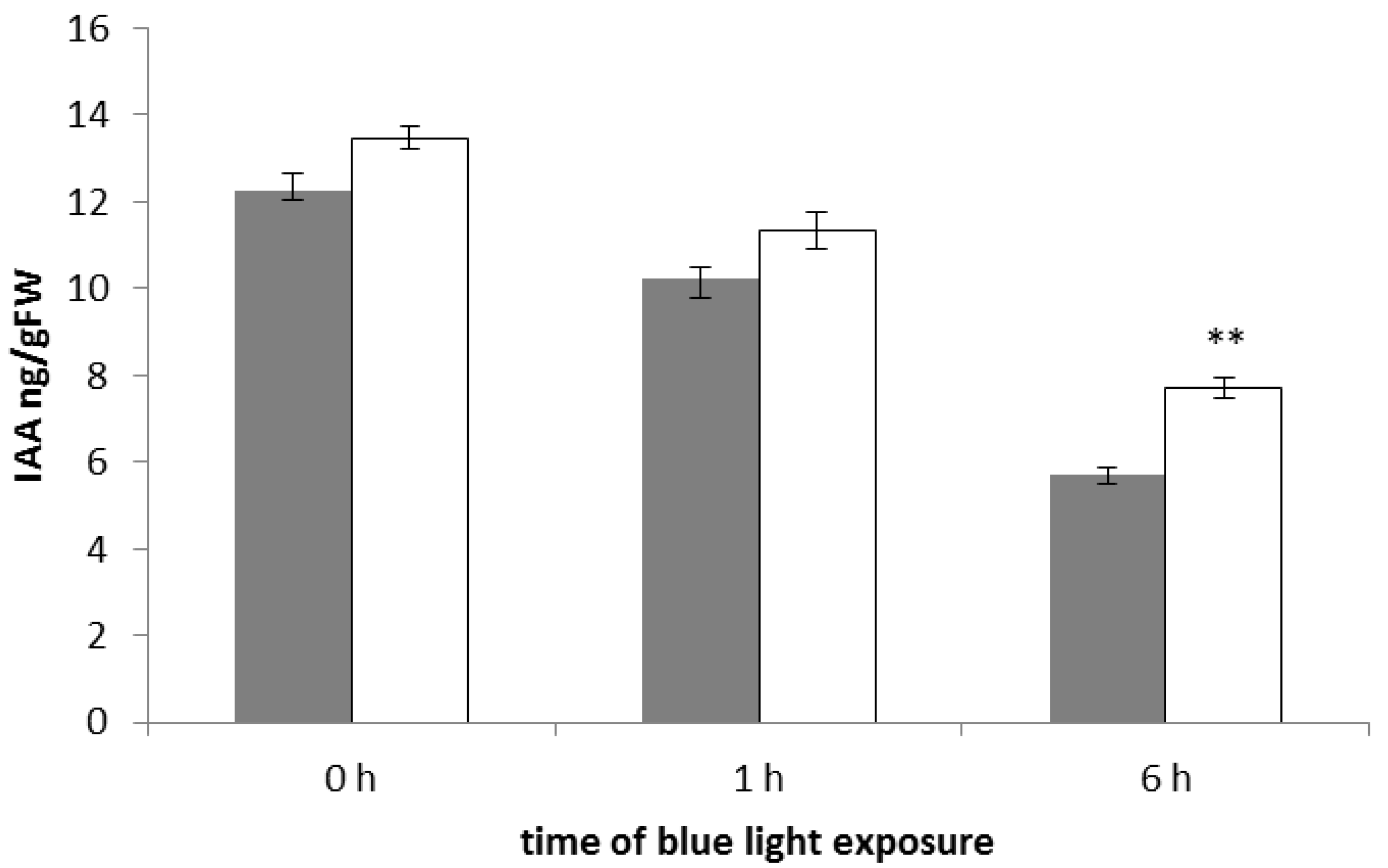
2.3. The phyAphyC Mutant Produces Higher Amounts of Jasmonates in Response to Blue Light
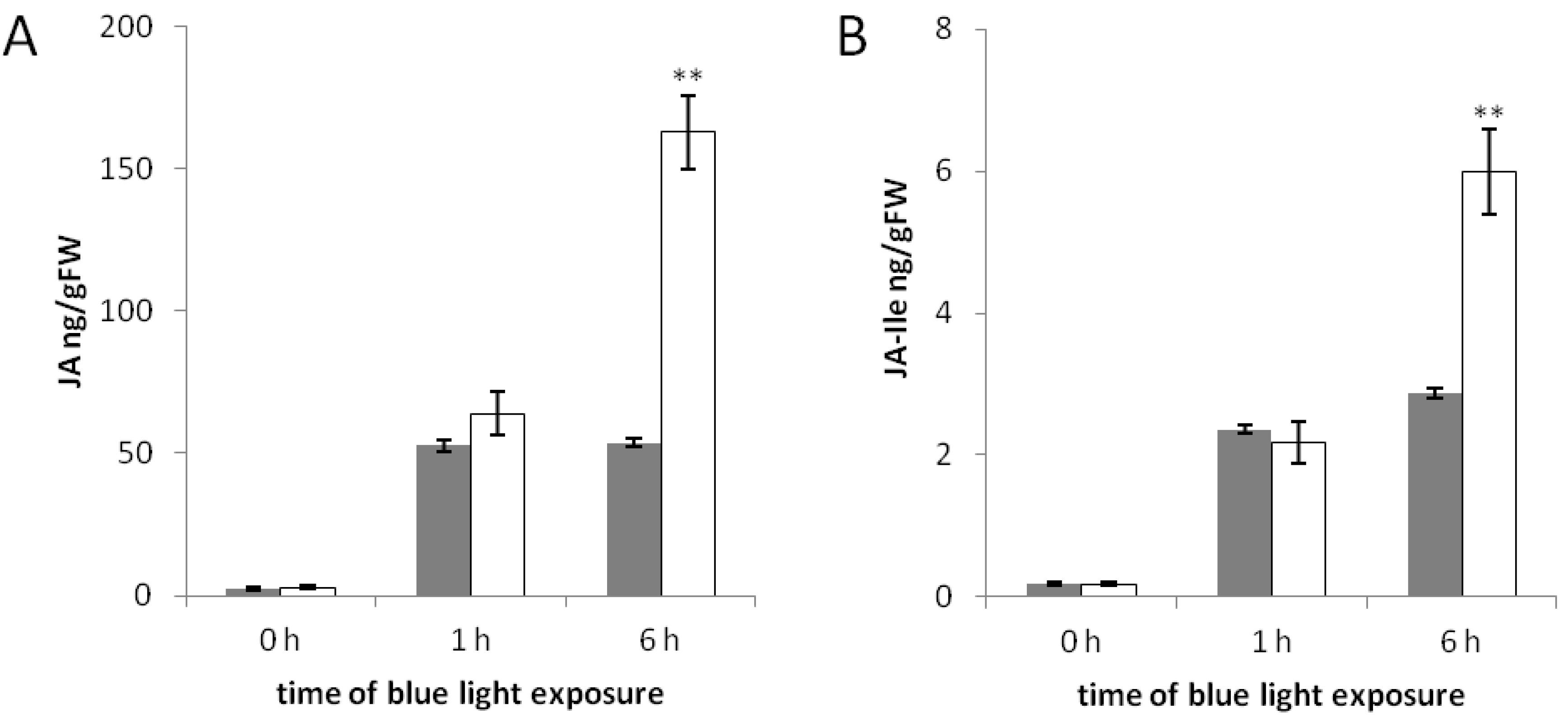
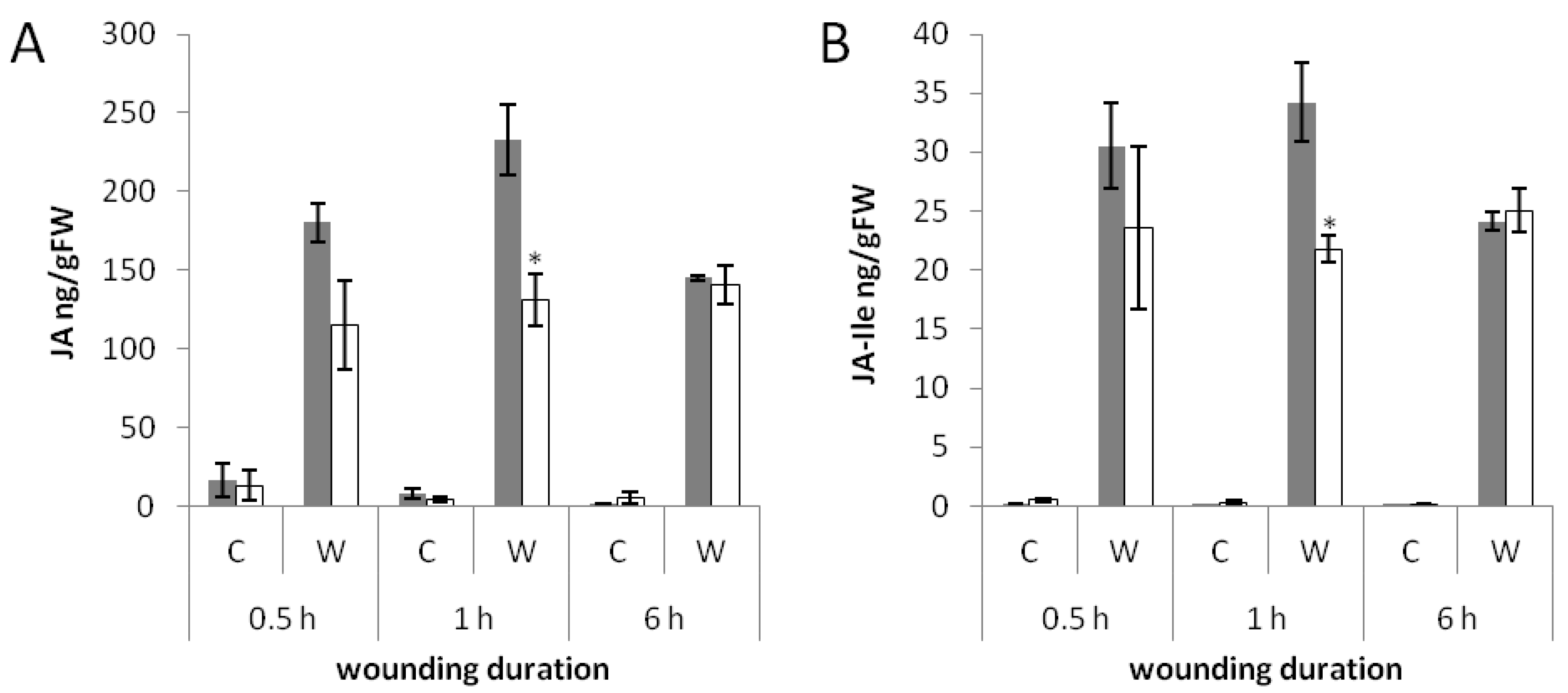
2.4. Less Jasmonates Are Produced in phyAphyC in Response to Mechanical Wounding
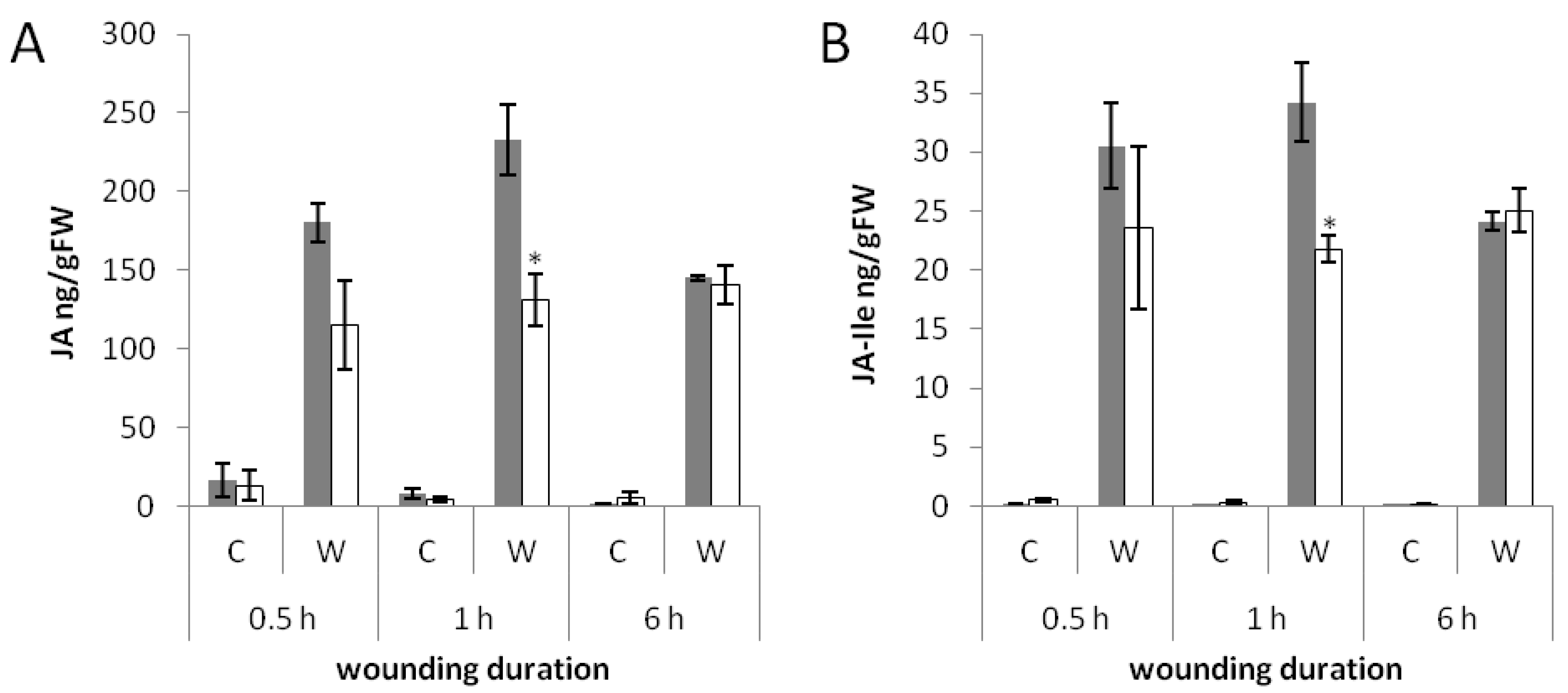
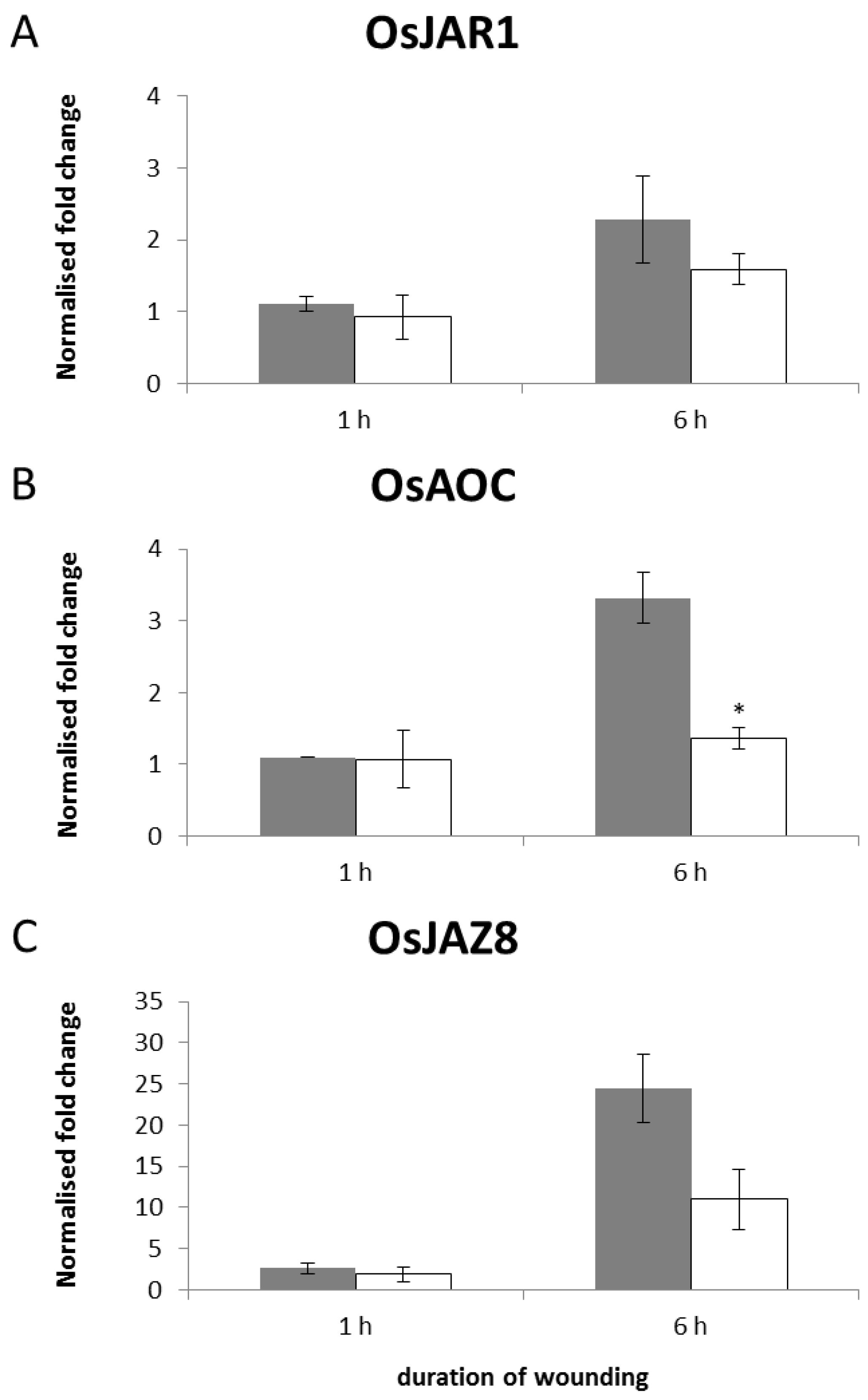
2.5. Transcriptional Regulation of JA-Responsive Genes by Continuous Mechanical Wounding
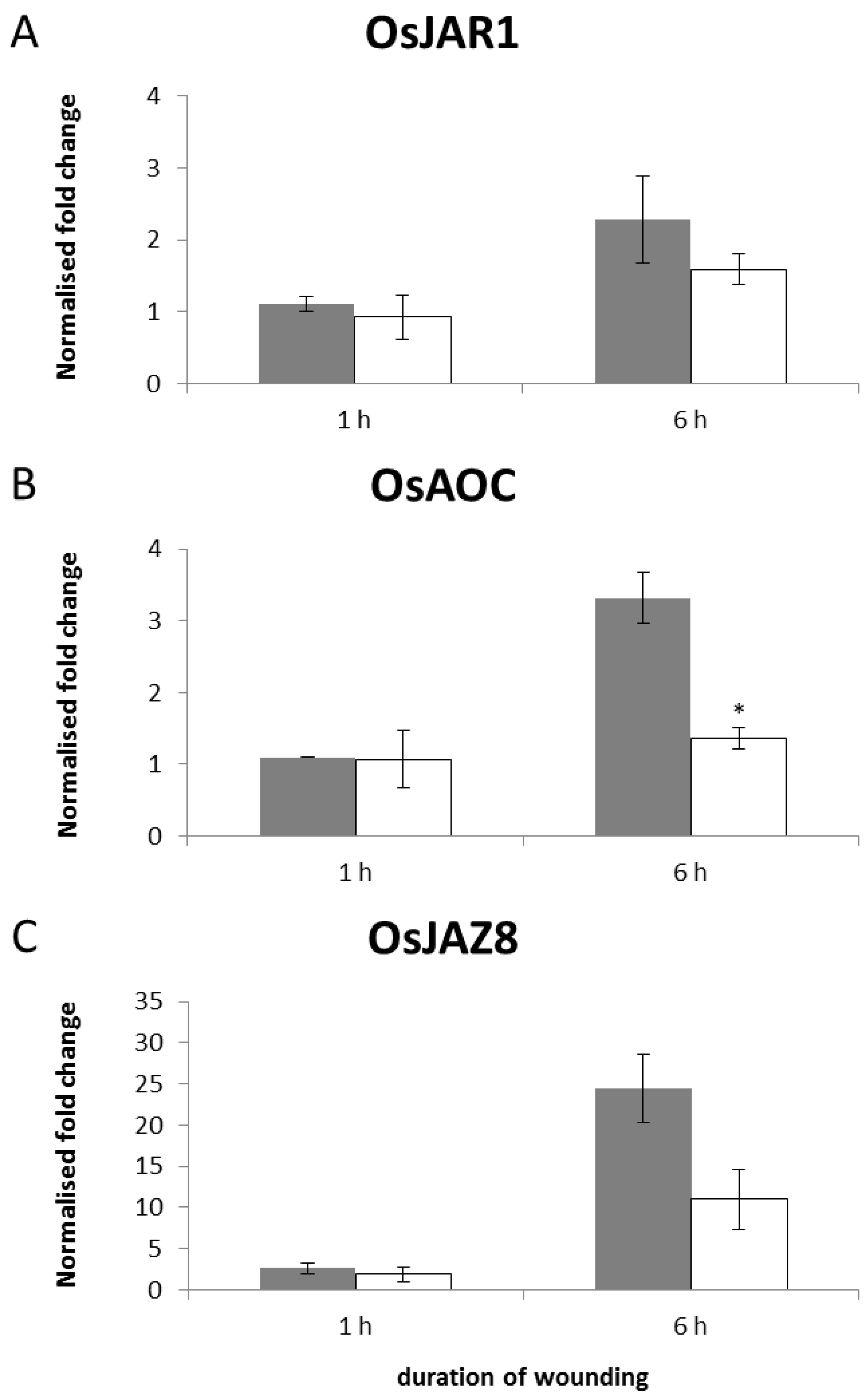
3. Experimental
3.1. Plant Material
3.2. Plant Cultivation and Treatments
3.3. Hormone Analysis
3.4. Gene Expression Analysis
3.5. Statistical Analysis
4. Conclusions
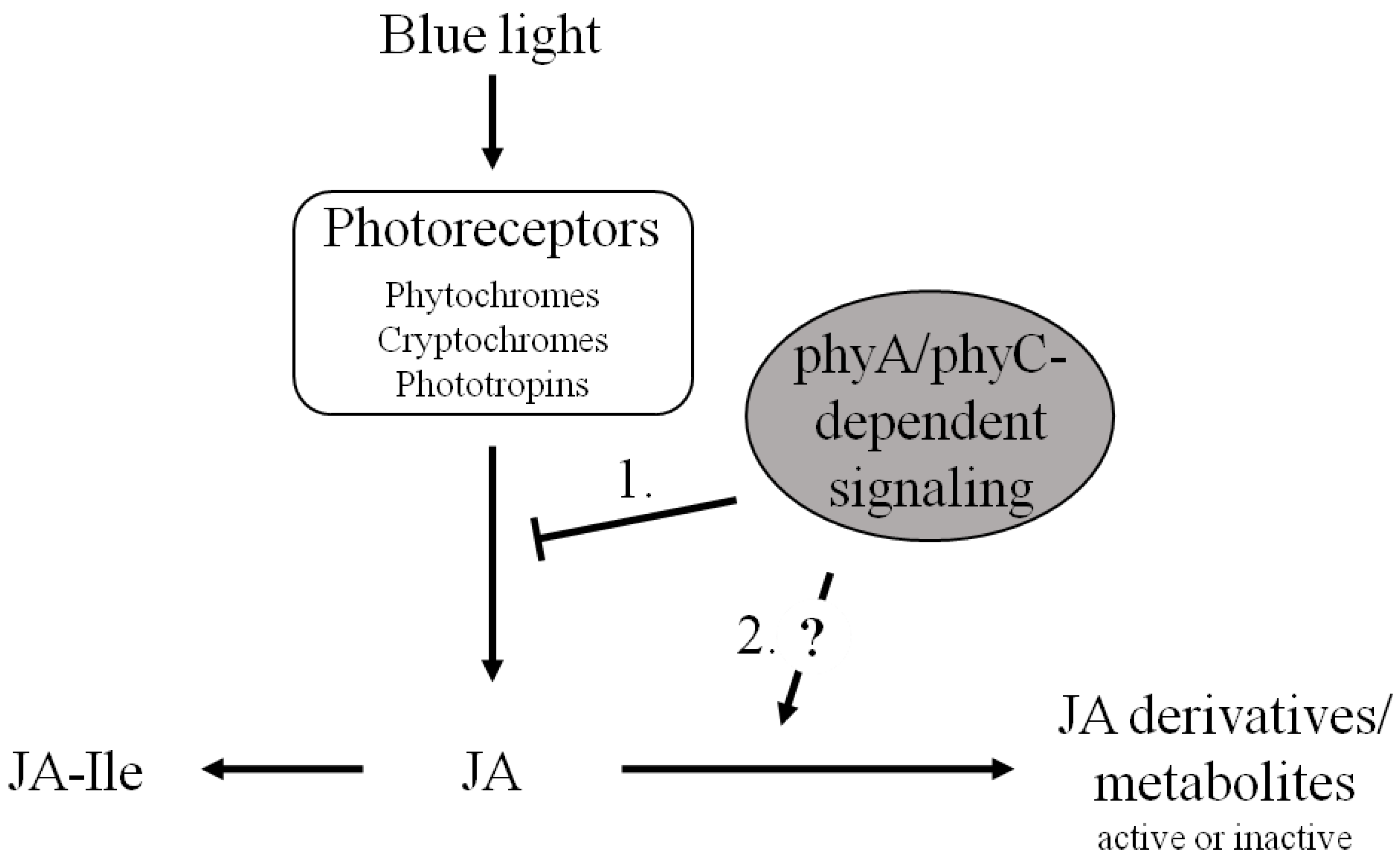
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Briggs, W.R.; Christie, J.M. Phototropins 1 and 2: Versatile plant blue-light receptors. Trends Plant Sci. 2002, 7, 204–210. [Google Scholar]
- Christie, J.M. Phototropin blue-light receptors. Annu. Rev. Plant Biol. 2007, 58, 21–45. [Google Scholar] [CrossRef]
- Lin, C.; Shalitin, D. Cryptochrome structure and signal transduction. Annu. Rev. Plant Biol. 2003, 54, 469–496. [Google Scholar] [CrossRef]
- Sancar, A. Structure and function of DNA photolyase and cryptochrome blue-light photoreceptors. Chem. Rev. 2003, 103, 2203–2237. [Google Scholar] [CrossRef]
- Furuya, M. Phytochromes—Their Molecular Species, Gene Families, and Functions. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 617–645. [Google Scholar] [CrossRef]
- Nagy, F.; Schäfer, E. Phytochromes control photomorphogenesis by differentially regulated, interacting signaling pathways in higher plants. Annu. Rev. Plant Biol. 2002, 53, 329–355. [Google Scholar] [CrossRef]
- Quail, P.H. Phytochrome: A light-activated molecular switch that regulates plant gene expression. Annu. Rev. Genet. 1991, 25, 389–409. [Google Scholar] [CrossRef]
- Smith, H. Physiological and Ecological Function within the Phytochrome Family. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1995, 46, 289–315. [Google Scholar] [CrossRef]
- Takano, M.; Inagaki, N.; Xie, X.; Kiyota, S.; Baba-Kasai, A.; Tanabata, T.; Shinomura, T. Phytochromes are the sole photoreceptors for perceiving red/far-red light in rice. Proc. Natl. Acad. Sci. USA 2009, 106, 14705–14710. [Google Scholar]
- Takano, M.; Inagaki, N.; Xie, X.; Yuzurihara, N.; Hihara, F.; Ishizuka, T.; Yano, M.; Nishimura, M.; Miyao, A.; Hirochika, H.; et al. Distinct and cooperative functions of phytochromes A, B, and C in the control of deetiolation and flowering in rice. Plant Cell 2005, 17, 3311–3325. [Google Scholar] [CrossRef]
- Smith, H.; Holmes, M.G. The function of phytochrome in natural-environment—III. Measurement and calculation of phytochrome photo-equilibria. Photochem. Photobiol. 1977, 25, 547–550. [Google Scholar] [CrossRef]
- Botto, J.F.; Sanchez, R.A.; Whitelam, G.C.; Casal, J.J. Phytochrome a mediates the promotion of seed germination by very low fluences of light and canopy shade light in Arabidopsis. Plant Physiol. 1996, 110, 439–444. [Google Scholar]
- Pjon, C.-J.; Furuya, M. Phytochrome action in Oryza sativa L.: I. Growth responses of etiolated coleoptiles to red, far-red and blue light. Plant Cell Physiol. 1967, 8, 709–718. [Google Scholar]
- Xie, X.; Shinomura, T.; Inagaki, N.; Kiyota, S.; Takano, M. Phytochrome-mediated inhibition of coleoptile growth in rice: Age-dependency and action spectra. Photochem. Photobiol. 2007, 83, 131–138. [Google Scholar] [CrossRef]
- Hirose, F.; Shinomura, T.; Tanabata, T.; Shimada, H.; Takano, M. Involvement of rice cryptochromes in de-etiolation responses and flowering. Plant Cell Physiol. 2006, 47, 915–925. [Google Scholar] [CrossRef]
- Fujioka, S.; Li, J.; Choi, Y.H.; Seto, H.; Takatsuto, S.; Noguchi, T.; Watanabe, T.; Kuriyama, H.; Yokota, T.; Chory, J.; et al. The Arabidopsis deetiolated2 mutant is blocked early in brassinosteroid biosynthesis. Plant Cell 1997, 9, 1951–1962. [Google Scholar]
- Li, J.; Nagpal, P.; Vitart, V.; McMorris, T.C.; Chory, J. A role for brassinosteroids in light-dependent development of Arabidopsis. Science 1996, 272, 398–401. [Google Scholar]
- Haga, K.; Iino, M. Phytochrome-mediated transcriptional up-regulation of ALLENE OXIDE SYNTHASE in rice seedlings. Plant Cell Physiol. 2004, 45, 119–128. [Google Scholar] [CrossRef]
- Riemann, M.; Müller, A.; Korte, A.; Furuya, M.; Weiler, E.W.; Nick, P. Impaired induction of the jasmonate pathway in the rice mutant hebiba. Plant Physiol. 2003, 133, 1820–1830. [Google Scholar] [CrossRef]
- Svyatyna, K.; Riemann, M. Light-dependent regulation of the jasmonate pathway. Protoplasma 2012, 249, S137–S145. [Google Scholar] [CrossRef]
- Wasternack, C.; Hause, B. Jasmonates: Biosynthesis, perception, signal transduction and action in plant stress response, growth and development. An update to the 2007 review in Annals of Botany. Ann. Bot. 2013, 111, 1021–1058. [Google Scholar] [CrossRef]
- Staswick, P.E.; Tiryaki, I. The oxylipin signal jasmonic acid is activated by an enzyme that conjugates it to isoleucine in Arabidopsis. Plant Cell 2004, 16, 2117–2127. [Google Scholar] [CrossRef]
- Sheard, L.B.; Tan, X.; Mao, H.; Withers, J.; Ben-Nissan, G.; Hinds, T.R.; Kobayashi, Y.; Hsu, F.F.; Sharon, M.; Browse, J.; et al. Jasmonate perception by inositol-phosphate-potentiated COI1-JAZ co-receptor. Nature 2010, 468, 400–405. [Google Scholar] [CrossRef]
- Xie, D.X.; Feys, B.F.; James, S.; Nieto-Rostro, M.; Turner, J.G. COI1: An Arabidopsis gene required for jasmonate-regulated defense and fertility. Science 1998, 280, 1091–1094. [Google Scholar]
- Chini, A.; Fonseca, S.; Fernandez, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; Garcia-Casado, G.; Lopez-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCF(COI1) complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- Riemann, M.; Haga, K.; Shimizu, T.; Okada, K.; Ando, S.; Mochizuki, S.; Nishizawa, Y.; Yamanouchi, U.; Nick, P.; Yano, M.; et al. Identification of rice Allene Oxide Cyclase mutants and the function of jasmonate for defence against Magnaporthe oryzae. Plant. J. 2013, 74, 226–238. [Google Scholar] [CrossRef]
- Riemann, M.; Bouyer, D.; Hisada, A.; Müller, A.; Yatou, O.; Weiler, E.W.; Takano, M.; Furuya, M.; Nick, P. Phytochrome A requires jasmonate for photodestruction. Planta 2009, 229, 1035–1045. [Google Scholar] [CrossRef]
- Sineshchekov, V.A.; Loskovich, A.V.; Riemann, M.; Nick, P. The jasmonate-free rice mutant hebiba is affected in the response of phyA'/phyA" pools and protochlorophyllide biosynthesis to far-red light. Photochem. Photobiol. Sci. 2004, 3, 1058–1062. [Google Scholar] [CrossRef]
- Radhika, V.; Kost, C.; Mithöfer, A.; Boland, W. Regulation of extrafloral nectar secretion by jasmonates in lima bean is light dependent. Proc. Natl. Acad. Sci. USA 2010, 107, 17228–17233. [Google Scholar] [CrossRef]
- Svyatyna, K.; Jikumaru, Y.; Brendel, R.; Reichelt, M.; Mithöfer, A.; Takano, M.; Kamiya, Y.; Nick, P.; Riemann, M. Light induces jasmonate-isoleucine conjugation via OsJAR1-dependent and -independent pathways in rice. Plant Cell Environ. 2013. [Google Scholar] [CrossRef]
- Riemann, M.; Takano, M. Rice JASMONATE RESISTANT 1 is involved in phytochrome and jasmonate signalling. Plant Cell Environ. 2008, 31, 783–792. [Google Scholar] [CrossRef]
- Yamada, S.; Kano, A.; Tamaoki, D.; Miyamoto, A.; Shishido, H.; Miyoshi, S.; Taniguchi, S.; Akimitsu, K.; Gomi, K. Involvement of OsJAZ8 in jasmonate-induced resistance to bacterial blight in rice. Plant Cell Physiol. 2012, 53, 2060–2072. [Google Scholar] [CrossRef]
- Terol, J.; Domingo, C.; Talon, M. The GH3 family in plants: Genome wide analysis in rice and evolutionary history based on EST analysis. Gene 2006, 371, 279–290. [Google Scholar] [CrossRef]
- Jain, M.; Kaur, N.; Tyagi, A.K.; Khurana, J.P. The auxin-responsive GH3 gene family in rice (Oryza sativa). Funct. Integr. Genomics 2006, 6, 36–46. [Google Scholar] [CrossRef]
- Ye, H.; Du, H.; Tang, N.; Li, X.; Xiong, L. Identification and expression profiling analysis of TIFY family genes involved in stress and phytohormone responses in rice. Plant Mol. Biol. 2009, 71, 291–305. [Google Scholar] [CrossRef]
- Costigan, S.E.; Warnasooriya, S.N.; Humphries, B.A.; Montgomery, B.L. Root-localized phytochrome chromophore synthesis is required for photoregulation of root elongation and impacts root sensitivity to jasmonic acid in Arabidopsis. Plant Physiol. 2011, 157, 1138–1150. [Google Scholar] [CrossRef]
- Zhai, Q.; Li, C.B.; Zheng, W.; Wu, X.; Zhao, J.; Zhou, G.; Jiang, H.; Sun, J.; Lou, Y.; Li, C. Phytochrome chromophore deficiency leads to overproduction of jasmonic acid and elevated expression of jasmonate-responsive genes in Arabidopsis. Plant Cell Physiol. 2007, 48, 1061–1071. [Google Scholar] [CrossRef]
- Chen, J.; Sonobe, K.; Ogawa, N.; Masuda, S.; Nagatani, A.; Kobayashi, Y.; Ohta, H. Inhibition of arabidopsis hypocotyl elongation by jasmonates is enhanced under red light in phytochrome B dependent manner. J. Plant Res. 2013, 126, 161–168. [Google Scholar] [CrossRef]
- Sun, J.; Xu, Y.; Ye, S.; Jiang, H.; Chen, Q.; Liu, F.; Zhou, W.; Chen, R.; Li, X.; Tietz, O.; et al. Arabidopsis ASA1 is important for jasmonate-mediated regulation of auxin biosynthesis and transport during lateral root formation. Plant Cell 2009, 21, 1495–1511. [Google Scholar] [CrossRef]
- Wakuta, S.; Suzuki, E.; Saburi, W.; Matsuura, H.; Nabeta, K.; Imai, R.; Matsui, H. OsJAR1 and OsJAR2 are jasmonyl-l-isoleucine synthases involved in wound- and pathogen-induced jasmonic acid signalling. Biochem. Biophys. Res. Commun. 2011, 409, 634–639. [Google Scholar] [CrossRef]
- Mithöfer, A.; Wanner, G.; Boland, W. Effects of feeding Spodoptera littoralis on lima bean leaves. II. Continuous mechanical wounding resembling insect feeding is sufficient to elicit herbivory-related volatile emission. Plant Physiol. 2005, 137, 1160–1168. [Google Scholar] [CrossRef]
- Trewavas, A.J. Growth substance sensitivity—The limiting factor in plant development. Physiol. Plant. 1982, 55, 60–72. [Google Scholar] [CrossRef]
- Qiao, F.; Petrášek, J.; Nick, P. Light can rescue auxin-dependent synchrony of cell division in a tobacco cell line. J. Exp. Bot. 2010, 61, 503–510. [Google Scholar] [CrossRef]
Appendix
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brendel, R.; Svyatyna, K.; Jikumaru, Y.; Reichelt, M.; Mithöfer, A.; Takano, M.; Kamiya, Y.; Nick, P.; Riemann, M. Effects of Light and Wounding on Jasmonates in Rice phyAphyC Mutants. Plants 2014, 3, 143-159. https://doi.org/10.3390/plants3010143
Brendel R, Svyatyna K, Jikumaru Y, Reichelt M, Mithöfer A, Takano M, Kamiya Y, Nick P, Riemann M. Effects of Light and Wounding on Jasmonates in Rice phyAphyC Mutants. Plants. 2014; 3(1):143-159. https://doi.org/10.3390/plants3010143
Chicago/Turabian StyleBrendel, Rita, Katharina Svyatyna, Yusuke Jikumaru, Michael Reichelt, Axel Mithöfer, Makoto Takano, Yuji Kamiya, Peter Nick, and Michael Riemann. 2014. "Effects of Light and Wounding on Jasmonates in Rice phyAphyC Mutants" Plants 3, no. 1: 143-159. https://doi.org/10.3390/plants3010143