NMR-Based Milk Metabolomics



Abstract
:1. Introduction
2. Milk Metabolomics

2.1. Detection of Milk Metabolites and Biomarkers by High-Resolution Proton NMR Spectroscopy

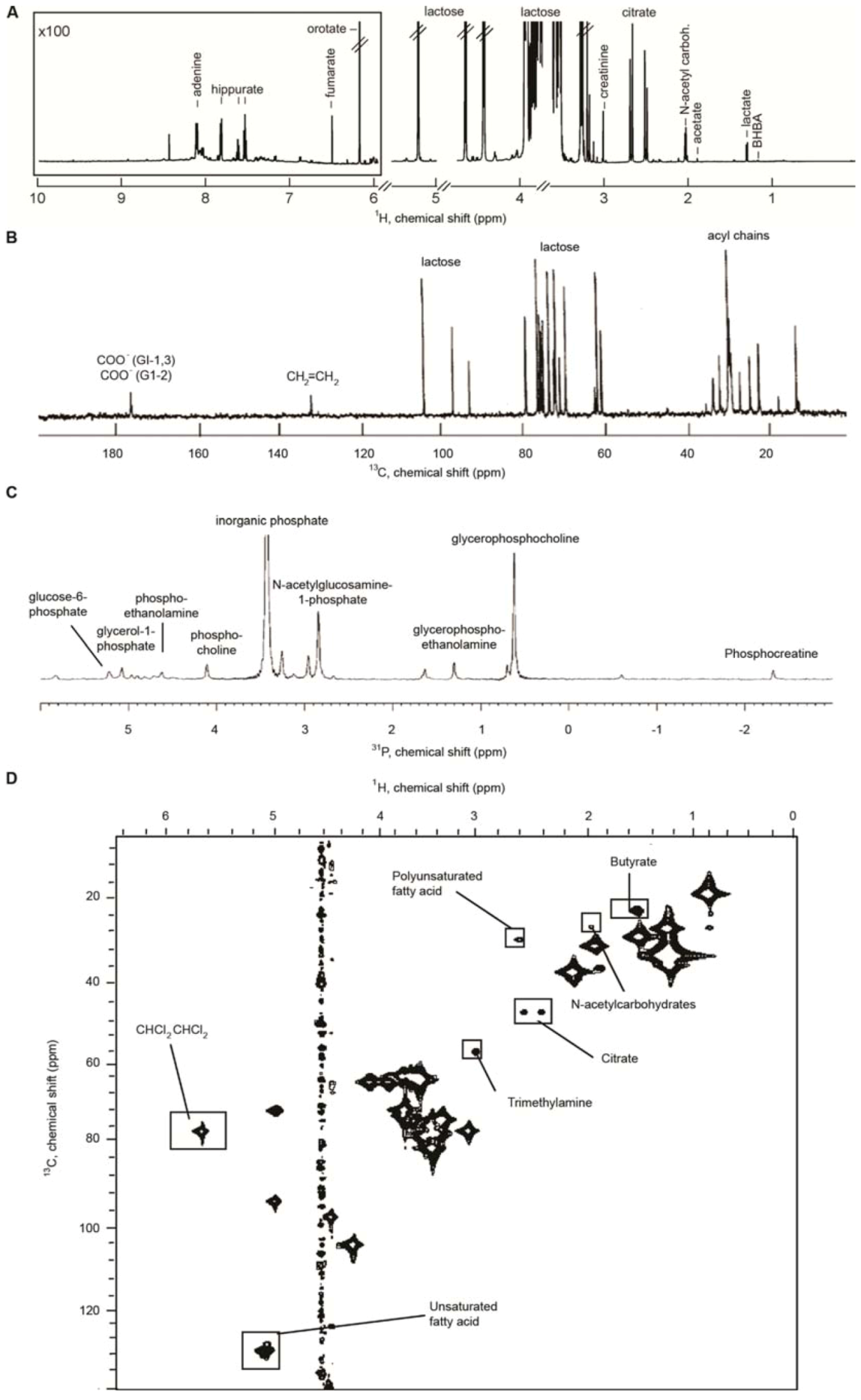{kind=link}
| Metabolite | Assignment | 1H Chemical Shift (ppm) | Reference |
|---|---|---|---|
| Acetate | CH3 | 1.92 | [35] |
| Acetone | CH3 | 2.24 | [35] |
| cis-aconitate | CH2 | 3.15 | [44] |
| Adenine | CH-8 | 8.12 | [37] |
| Adenine | CH-2 | 8.13 | [37] |
| Alanine | CH | 3.79 | [35] |
| Alanine | CH3 | 1.48 | [35] |
| β-hydroxybutyrate | CH3 | 1.20 | [35] |
| Betaine | 3 × CH3 | 3.26 | [35] |
| Butyrate | CH3 | 0.90 | [37] |
| Carnitine | 3 × CH3 | 3.21 | [35] |
| Carnitine | CH2 | 2.44 | [35] |
| Carnitine | N-CH2 | 3.43 | [35] |
| Carnitine | CH | 4.57 | [35] |
| Choline | 3 × CH3 | 3.18 | [35] |
| Choline | O-CH2 | 4.06 | [35] |
| Choline | N-CH2 | 3.51 | [35] |
| Citrate | CH2 | 2.52 | [39] |
| Citrate | CH2 | 2.72 | [39] |
| Creatine | CH3 | 3.79 | [39] |
| Creatine | CH2 | 2.88 | [39] |
| Creatinine | CH2 | 4.06 | [35] |
| Creatinine | CH3 | 3.05 | [35] |
| Ethanolamine | O-CH2 | 3.83 | [35] |
| Ethanolamine | N-CH2 | 3.15 | [35] |
| Formate | CH | 8.45 | [45] |
| Fucose | CH3 | 1.25 | [44] |
| Fumarate | CH | 6.52 | [37] |
| Galactose α | CH | 4.07 | [35] |
| Galactose α | CH | 3.81 | [35] |
| Galactose β | CH | 4.57 | [35] |
| Galactose β | CH | 3.49 | [35] |
| Galactose-1-phosphate | CH-1 | 5.38 | [44] |
| Glucose | CH2 | 5.1 | [44] |
| Glucose-1-phosphate | CH-1 | 5.51 | [44] |
| Glutamate | γ-CH2 | 2.36 | [44] |
| Glycerophosphocholine | O-CH2 | 4.32 | [35] |
| Glycerophosphocholine | N-CH2 | 3.65 | [35] |
| Glycine | CH2 | 3.57 | [35] |
| Hippurate | CH2-2,6 | 7.84 | [37] |
| Hippurate | CH-4 | 7.64 | [37] |
| Hippurate | CH2-3,5 | 7.54 | [37] |
| Isobutyrate | CH3 | 1.16 | [44] |
| Isoleucine | δ-CH3 | 0.93 | [37] |
| Lactate | CH3 | 1.32 | [35] |
| Lactate | CH | 4.11 | [35] |
| Lactose (total) | CH-1’ | 4.45 | [35,39] |
| Lactose (total) | CH-5’ | 3.73 | [35,39] |
| Lactose (total) | CH-2 | 3.94 | [35,39] |
| Lactose (total) | CH-2’ | 3.54 | [35,39] |
| Lactose (total) | CH2-6 | 3.67 | [35,39] |
| Lactose (total) | CH2-6’ | 3.78 | [35,39] |
| Lactose α | CH-1 | 5.23 | [35,39] |
| Lactose α | CH2-6 | 3.88 | [35,39] |
| Lactose α | CH-3 | 3.59 | [35,39] |
| Lactose α | CH-4' | 3.96 | [35,39] |
| Lactose α | CH-4 | 3.66 | [35,39] |
| Lactose α | CH-5' | 3.84 | [35,39] |
| Lactose β | CH-1 | 4.67 | [35,39] |
| Lactose β | CH-4 | 3.66 | [35,39] |
| Lactose β | CH-2 | 3.29 | [35,39] |
| Lactose β | ½ CH2-6 | 3.96 | [35,39] |
| Lactose β | CH-3 | 3.6 | [35,39] |
| Lactose β | ½ CH2-6 | 3.81 | [35,39] |
| Lactose β | CH-5 | 3.84 | [35,39] |
| Lecithin | 3 × CH3 | 3.12 | [39] |
| Lecithin | CH2-4 | 4.22 | [39] |
| Lecithin | CH2-3 | 4.18 | [39] |
| Lecithin | CH2-5 | 3.75 | [39] |
| Lecithin | CH2-5 | 3.83 | [39] |
| Malonic acid | CH2 | 3.11 | [44] |
| 3-Methylhistidine | α-CH | 3.97 | [35] |
| 3-Methylhistidine | β-CH | 3.30 / 2.25 | [35] |
| 3-Methylhistidine | CH3 | 3.74 | [35] |
| 3-Methylhistidine | δ-CH | 7.14 | [35] |
| 3-Methylhistidine | ε-CH | 8.09 | [35] |
| Methionine | γ-CH2, S-CH3 | 2.15 | [44] |
| N-acetylcarbohydrates | CH3 | 2.06 | [39] |
| Ornithine | γ-CH2 | 1.80 | [44] |
| Orotate | CH | 6.20 | [37] |
| Phosphocholine | O-CH2 | 4.16 | [35] |
| Phosphocholine | N-CH2 | 3.58 | [35] |
| Phosphocholine | 3*CH3 | 3.18 | [35] |
| Phosphocreatine | CH3 | 3.03 | [35] |
| Phosphocreatine | CH2 | 3.93 | [35] |
| Taurine | S-CH2 | 3.43 | [35] |
| Taurine | N-CH2 | 3.27 | [35] |
| Triethylamine-N-oxide | CH3 | 3.27 | [35] |
| Urea | NH2 | 5.79 | [44] |
| Valine | CH3 | 1.05 | [44] |
2.2. Metabolomics-Assisted Elucidation of Important Technological Milk Quality Parameters
2.3. Milk Authentication and Control of Geographical Origin
2.4. Metabolomics and Nutritional Quality of Milk
2.5. Impact of Genes on Milk Metabolite Variability
2.6. Summary
| Factor under investigation | Metabolite(s) | Analytical technique | Reference(s) |
|---|---|---|---|
| Coagulation properties | Choline, carnitine, citrate, lactose | 1H NMR | [34] |
| Somatic cell count | BHBA, lactate, lactose, hippurate, acetate, fumarate, butyrate | 1H NMR | [47] |
| Metabolic status of cows | Acetone, BHBA | 1H NMR & GC-MS | [35] |
| Quality control | Citrate, N-acetylcarbohydrates, trimethylamine, lecithin | 1H NMR | [39,41] |
| Quality control | Lactose | 1H NMR | [58] |
| Classification of milk blends from different species | N-acetyllactosamine, citrate, acetamide, phosphocreatine | 1H NMR | [55] |
| Milk authenticity and assessment of adulteration | Melamine | 1H NMR & GC-MS | [56] |
| Milk authenticity | Glycerol 1-phosphate, glucose 6-phosphate, phospholipids | 31P NMR | [40] |
| Genetic influence on milk metabolites | BHBA, orotate, carnitine, malonate | 1H NMR | [44] |
| Infant formula, pre-term, and full-term human milk | Lactose, maltose | 1H NMR | [38] |
| Nutrition | Phospholipids | 31P NMR | [62] |
| Spoilage, storage | Phosphoglycerides | 31P NMR | [52] |
| Nutrition, human milk | Choline, phosphocholines | 1H NMR | [60] |
| Human and rhesus macaque milk, nutrition | Amino acids, oligosaccharides, glycerophosphocholine, hippurate | 1H NMR | [61] |
| Associations of blood-milk metabolites | Trimethylamine, lactose, citrate, dimethylsulphone, orotate, fumarate, valine | 1H NMR | [48] |
3. Experimental Considerations
3.1. NMR Spectroscopy
3.2. Data Handling
3.3. Data Analysis
| Technique | Unsupervised / Supervised | Characteristics |
|---|---|---|
| PCA | Unsupervised | Exploratory clustering technique extremely useful in identification of differences between observations including variable differences and covariances |
| PLS-DA | Supervised | Maximum separation between groups of observations is achieved using rotating PCA components. Useful for obtaining information about which variables are involved in class separation |
| OPLS-DA | Supervised | Systematic variation that is not correlated with classes is removed, which may improve interpretation but not predictivity |
| HCA | Unsupervised | Exploratory tool to visualize groupings of observations and represented as a tree or dendrogram showing observation homology |
| RF | Supervised or unsupervised | A learning algorithm which uses an ensemble of decision trees to assign class relationships to observations |
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Elgersma, A.; Tamminga, S.; Ellen, G. Modifying milk composition through forage. Anim. Feed Sci. Tech. 2006, 131, 207–225. [Google Scholar] [CrossRef]
- Arnould, V.M.R.; Soyeurt, H. Genetic variability of milk fatty acids. J. Appl. Genet. 2009, 50, 29–39. [Google Scholar] [CrossRef]
- Garnsworthy, P.C.; Masson, L.L.; Lock, A.L.; Mottram, T.T. Variation of milk citrate with stage of lactation and de novo fatty acid synthesis in dairy cows. J. Dairy Sci. 2006, 89, 1604–1612. [Google Scholar] [CrossRef]
- Heck, J.M. L.; van Valenberg, H.J.F.; Dijkstra, J.; van Hooijdonk, A.C.M. Seasonal variation in the Dutch bovine raw milk composition. J. Dairy Sci. 2009, 92, 4745–4755. [Google Scholar] [CrossRef]
- Auldist, M.J.; Hubble, I.B. Effects of mastitis on raw milk and dairy products. Aust. J. Dairy Technol. 1998, 53, 28–36. [Google Scholar]
- Walker, G.P.; Dunshea, F.R.; Doyle, P.T. Effects of nutrition and management on the production and composition of milk fat and protein: A review. Aust. J. Agric. Res. 2004, 55, 1009–1028. [Google Scholar] [CrossRef]
- Glantz, M.; Devold, T.G.; Vegarud, G.E.; Lindmark-Månsson, H.; Stålhammer, H.; Paulsson, M. Importance of casein micelle size and milk composition for milk gelation. J. Dairy Sci. 2010, 93, 1444–1451. [Google Scholar] [CrossRef]
- Wedholm, A.; Larsen, L.B.; Lindmark-Månsson, H.; Karlsson, A.H.; Andrén, A. Effect of protein composition on the cheese-making properties of milk from individual dairy cows. J. Dairy Sci. 2006, 89, 3296–3305. [Google Scholar] [CrossRef]
- Bittante, G.; Penasa, M.; Cecchinato, A. Invited review: Genetics and modeling of milk coagulation properties. J. Dairy Sci. 2012, 95, 6843–6870. [Google Scholar] [CrossRef]
- Le Maréchal, C.; Thiéry, R.; Vautor, E.; Le Loir, Y. Mastitis impact on technological properties of milk and quality of milk products—A review. Dairy Sci. Technol. 2011, 91, 247–282. [Google Scholar] [CrossRef]
- Tsioulpas, A.; Lewis, M.J.; Grandison, A.S. Effect of minerals on casein micelle stability of cows’ milk. J. Dairy Res. 2007, 74, 167–173. [Google Scholar] [CrossRef]
- Udabage, P.; McKinnon, I.R.; Augustin, M.A. Effects of mineral salts and calcium chelating agents on the gelation of renneted skim milk. J. Dairy Sci. 2001, 84, 1569–1575. [Google Scholar] [CrossRef]
- McManaman, J.L.; Neville, M.C. Mammary physiology and milk secretion. Adv. Drug. Deliver. Rev. 2003, 55, 629–641. [Google Scholar] [CrossRef]
- Nguyen, D.A.D.; Neville, M.C. Tight junction regulation in the mammary gland. J. Mammary Gland Biol. Neoplasia 1998, 3, 233–246. [Google Scholar] [CrossRef]
- Hettinga, K.; van Valenberg, H.; Lam, T.; van Hooijdonk, A. The origin of the volatile metabolites found in mastitis milk. Vet. Microbiol. 2009, 137, 384–387. [Google Scholar] [CrossRef]
- Hettinga, K.A.; van Valenberg, H.J. F.; Lam, T.J.G.M.; Van Hooijdonk, A.C.M. Detection of mastitis pathogens by analysis of volatile bacterial metabolites. J. Dairy Sci. 2008, 91, 3834–3839. [Google Scholar] [CrossRef]
- Azzara, C.D.; Dimick, P.S. Lipolytic enzyme activity of macrophages in bovine mammary gland secretions. J. Dairy Sci. 1985, 68, 1804–1812. [Google Scholar] [CrossRef]
- German, J.; Hammock, B.D.; Watkins, S.M. Metabolomics: Building on a century of biochemistry to guide human health. Metabolomics 2005, 1, 3–9. [Google Scholar] [CrossRef]
- Lindon, J.C.; Holmes, E.; Nicholson, J.K. Metabonomics techniques and applications to pharmaceutical research & development. Pharm. Res. 2006, 23, 1075–1088. [Google Scholar] [CrossRef]
- Wishart, D.S. Metabolomics: applications to food science and nutrition research. Trends Food Sci. Technol. 2008, 19, 482–493. [Google Scholar] [CrossRef]
- Kalo, P.; Kemppinen, A.; Kilpeläinen, I. Determination of positional distribution of butyryl groups in milkfat triacylglycerols, triacylglycerol mixtures, and isolated positional isomers of triacylglycerols by gas chromatography and 1H nuclear magnetic resonance spectroscopy. Lipids 1996, 31, 331–336. [Google Scholar] [CrossRef]
- Leslie, R.B.; Irons, L.; Chapman, D. High resolution nuclear magnetic resonance studies of alphas1, beta and kappa-caseins. Biochim. Biophys. Acta 1969, 188, 237–246. [Google Scholar]
- Humphrey, R.S.; Jolley, K.W. P-31-NMR Studies of Bovine Beta-Casein. Biochim. Biophys. Acta. 1982, 708, 294–299. [Google Scholar]
- Griffin, M.C.A.; Roberts, G.C.K. A H-1-NMR study of casein micelles. Biochem. J. 1985, 228, 273–276. [Google Scholar]
- Rollema, H.S.; Brinkhuis, J.A. A H-1-NMR study of bovine casein micelles—Influence of Ph, temperature and calcium-ions on micellar structure. J. Dairy Res. 1989, 56, 417–425. [Google Scholar]
- Kakalis, L.T.; Kumosinski, T.F.; Farrell, H.M. A Multinuclear, high-resolution NMR-study of bovine casein micelles and submicelles. Biophys. Chem. 1990, 38, 87–98. [Google Scholar]
- Belloque, J.; Smith, G.M. Thermal Denaturation of β-Lactog lobulin. A 1H NMR Study. J. Agric. Food Chem. 1998, 46, 1805–1813. [Google Scholar]
- Lubke, M.; Guichard, E.; Tromelin, A.; Le Quere, J.L. Nuclear magnetic resonance spectroscopic study of beta-lactoglobulin interactions with two flavor compounds, gamma-decalactone and beta-ionone. J. Agric. Food Chem. 2002, 50, 7094–7099. [Google Scholar]
- Kuwata, K.; Hoshino, M.; Era, S.; Batt, C.A.; Goto, Y. α-β transition of β-lactoglobulin as evidenced by heteronuclear NMR. J. Mol. Biol. 1998, 283, 731–739. [Google Scholar]
- Belloque, J.; De la Fuente, M.A.; Ramos, M. Qualitative and quantitative analysis of phosphorylated compounds in milk by means of P-31-NMR. J. Dairy Res. 2000, 67, 529–539. [Google Scholar]
- Belloque, J.; Ramos, M. Determination of the casein content in bovine milk by P-31-NMR. J. Dairy Res. 2002, 69, 411–418. [Google Scholar]
- Belton, P.S.; Lyster, R.L.J.; Richards, C.P. The 31P nuclear magnetic resonance spectrum of cows’ milk. J. Dairy Res. 1985, 52, 47–54. [Google Scholar]
- Belloque, J.; Ramos, M. Application of NMR spectroscopy to milk and dairy products. Trends Food Sci. Technol. 1999, 10, 313–320. [Google Scholar]
- Sundekilde, U.K.; Frederiksen, P.D.; Clausen, M.R.; Larsen, L.B.; Bertram, H.C. Relationship between the metabolite profile and technological properties of bovine milk from two dairy breeds elucidated by NMR-based metabolomics. J. Agric. Food Chem. 2011, 59, 7360–7367. [Google Scholar]
- Klein, M.S.; Almstetter, M.F.; Schlamberger, G.; Nurnberger, N.; Dettmer, K.; Oefner, P.J.; Meyer, H.H.D.; Wiedemann, S.; Gronwald, W. Nuclear magnetic resonance and mass spectrometry-based milk metabolomics in dairy cows during early and late lactation. J. Dairy Sci. 2010, 93, 1539–1550. [Google Scholar]
- Klein, M.S.; Buttchereit, N.; Miemczyk, S.P.; Immervoll, A.K.; Louis, C.; Wiedemann, S.; Junge, W.; Thaller, G.; Oefner, P.J.; Gronwald, W. NMR metabolomic analysis of dairy cows reveals milk glycerophosphocholine to phosphocholine ratio as prognostic biomarker for risk of ketosis. J. Proteome Res. 2012, 11, 1373–1381. [Google Scholar]
- Sundekilde, U.K.; Poulsen, N.; Larsen, L.B.; Bertram, H.C. NMR metabonomics reveals strong association between milk metabolites and somatic cell count in bovine milk. J. Dairy Sci. 2013, 96, 290–299. [Google Scholar]
- Marincola, F.C.; Noto, A.; Caboni, P.; Reali, A.; Barberini, L.; Lussu, M.; Murgia, F.; Santoru, M.L.; Atzori, L.; Fanos, V. A metabolomic study of preterm human and formula milk by high resolution NMR and GC/MS analysis: preliminary results. J. Matern. Fetal Neonatal. Med. 2012, 25, 62–67. [Google Scholar]
- Hu, F.; Furihata, K.; Ito-Ishida, M.; Kaminogawa, S.; Tanokura, M. Nondestructive observation of bovine milk by NMR spectroscopy: analysis of existing states of compounds and detection of new compounds. J. Agric. Food Chem. 2004, 52, 4969–4974. [Google Scholar]
- Andreotti, G.; Trivellone, E.; Motta, A. Characterization of buffalo milk by 31P-nuclear magnetic resonance spectroscopy. J. Food Comp. Anal. 2006, 19, 843–849. [Google Scholar]
- Hu, F.; Furihata, K.; Kato, Y.; Tanokura, M. Nondestructive quantification of organic compounds in whole milk without pretreatment by two-dimensional NMR spectroscopy. J. Agric. Food Chem. 2007, 55, 4307–4311. [Google Scholar]
- Fox, P.F.; McSweeney, P.L.H. Dairy Chemistry and Biochemistry; Blackie Academic & Professional: London, UK, 1998. [Google Scholar]
- Enjalbert, F.; Nicot, M.C.; Baourthe, C.; Moncoulon, R. Ketone bodies in milk and blood of dairy cows: Relationship between concentrations and utilization for detection of subclinical ketosis. J. Dairy Sci. 2001, 84, 583–589. [Google Scholar]
- Buitenhuis, A.J.; Sundekilde, U.K.; Poulsen, N.; Bertram, H.C.; Larsen, L.B.; Sørensen, P. Estimation of Genetic Parameters and Detection of QTL for Metabolites in Danish Holstein Milk. J. Dairy Sci. 2013. [Google Scholar] [CrossRef]
- Sacco, D.; Brescia, M.A.; Sgaramella, A.; Casiello, G.; Buccolieri, A.; Ogrinc, N.; Sacco, A. Discrimination between Southern Italy and foreign milk samples using spectroscopic and analytical data. Food Chem. 2009, 114, 1559–1563. [Google Scholar]
- Davis, S.R.; Farr, V.C.; Prosser, C.G.; Nicholas, G.D.; Turner, S.A.; Lee, J.; Hart, A.L. Milk L-lactate concentration is increased during mastitis. J. Dairy Res. 2004, 71, 175–181. [Google Scholar]
- Berning, L.M.; Shook, G.E. Prediction of Mastitis Using Milk Somatic Cell Count, N-Acetyl-β-D-Glucosaminidase, and Lactose. J. Dairy Sci. 1992, 75, 1840–1848. [Google Scholar]
- Maher, A.D.; Hayes, B.; Cocks, B.; Marett, L.; Wales, W.J.; Rochfort, S. Latent biochemical relationships in the blood-milk axis of dairy cows revealed by statistical integration of H NMR spectroscopic data. J. Proteome Res. 2013. [Google Scholar] [CrossRef]
- Frederiksen, P.D.; Andersen, K.K.; Hammershøj, M.; Poulsen, H.D.; Sørensen, J.; Bakman, M.; Qvist, K.B.; Larsen, L.B. Composition and effect of blending of noncoagulating, poorly coagulating, and well-coagulating bovine milk from individual Danish Holstein cows. J. Dairy Sci. 2011, 94, 4787–4799. [Google Scholar]
- Feagan, J.T.; Bailey, L.F.; Hehir, A.F.; Mclean, D.M.; Ellis, N.J.S. Coagulation of milk proteins. I. Effect of genetic variants of milk proteins on rennet coagulation and heat stability of normal milk. J. Dairy Technol. 1972, 27, 129–134. [Google Scholar]
- Zabbia, A.; Buys, E.M.; De Kock, H.L. Undesirable Sulphur and Carbonyl Flavor Compounds in UHT Milk: A Review. Crit. Rev. Food Sci. 2011, 52, 21–30. [Google Scholar]
- Belloque, J.; Carrascosa, A.V.; pez, F. Changes in Phosphoglyceride Composition during Storage of Ultrahigh-Temperature Milk, as Assessed by 31P-Nuclear Magnetic Resonance: Possible Involvement of Thermoresistant Microbial Enzymes. J. Food Prot. 2001, 64, 850–855. [Google Scholar]
- Brescia, M.A.; Monfreda, M.; Buccolieri, A.; Carrino, C. Characterisation of the geographical origin of buffalo milk and mozzarella cheese by means of analytical and spectroscopic determinations. Food Chem. 2005, 89, 139–147. [Google Scholar]
- Mazzei, P.; Piccolo, A. 1H HRMAS-NMR metabolomic to assess quality and traceability of mozzarella cheese from Campania buffalo milk. Food Chem. 2012, 132, 1620–1627. [Google Scholar]
- Lamanna, R.; Braca, A.; Di Paolo, E.; Imparato, G. Identification of milk mixtures by 1H NMR profiling. Magn. Reson. Chem. 2011, 49, S22–S26. [Google Scholar]
- Lachenmeier, D.W.; Humpfer, E.; Fang, F.; Schütz, B.; Dvortsak, P.; Sproll, C.; Spraul, M. NMR-spectroscopy for nontargeted screening and simultaneous quantification of health-relevant compounds in foods: The example of melamine. J. Agric. Food Chem. 2009, 57, 7194–7199. [Google Scholar]
- Xin, H.; Stone, R. Tainted milk scandal CHINESE probe unmasks high-tech adulteration with melamine. Science 2008, 322, 1310–1311. [Google Scholar]
- Monakhova, Y.; Kuballa, T.; Leitz, J.; Andlauer, C.; Lachenmeier, D. NMR spectroscopy as a screening tool to validate nutrition labeling of milk, lactose-free milk, and milk substitutes based on soy and grains. Dairy Sci. Technol. 2012, 92, 109–120. [Google Scholar]
- Swallow, D.M. Genetics of lactase persistence and lactose intolerance. Annu. Rev. Genet. 2003, 37, 197–219. [Google Scholar]
- Holmes, H.C.; Snodgrass, I.; Iles, R.A. Changes in the choline content of human breast milk in the first 3 weeks after birth. Eur. J. Pediatr. 2000, 159, 198–204. [Google Scholar]
- O’Sullivan, A.; He, X.; McNiven, E.M.S.; Hinde, K.; Haggarty, N.W.; Lönnerdal, B.; Slupsky, C.M. Metabolomic phenotyping validates the infant rhesus monkey as A model of human infant metabolism. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 355–363. [Google Scholar]
- Garcia, C.; Lutz, N.W.; Confort-Gouny, S.; Cozzone, P.J.; Armand, M.; Bernard, M. Phospholipid fingerprints of milk from different mammalians determined by 31P NMR: Towards specific interest in human health. Food Chem. 2012, 135, 1777–1783. [Google Scholar]
- Jenkins, T.C.; McGuire, M.A. Major advances in nutrition: Impact on milk composition. J. Dairy Sci. 2006, 89, 1302–1310. [Google Scholar]
- Barding, G.A.; Salditos, R.; Larive, C.K. Quantitative NMR for bioanalysis and metabolomics. Anal. Bioanal. Chem. 2012, 404, 1165–1179. [Google Scholar]
- Miyataka, H.; Ozaki, T.; Himeno, S. Effect of pH on H-1-NMR spectroscopy of mouse urine. Biol. Pharm. Bull. 2007, 30, 667–670. [Google Scholar]
- Wishart, D.S.; Knox, C.; Guo, A.C.; Eisner, R.; Young, N.; Gautam, B.; Hau, D.D.; Psychogios, N.; Dong, E.; Bouatra, S.; et al. HMDB: A knowledgebase for the human metabolome. Nucl. Acids Res. 2009, 37, D603–D610. [Google Scholar]
- Ulrich, E.L.; Akutsu, H.; Doreleijers, J.F.; Harano, Y.; Ioannidis, Y.E.; Lin, J.; Livny, M.; Mading, S.; Maziuk, D.; Miller, Z.; et al. BioMagResBank. Nucl. Acids Res. 2008, 36, D402–D408. [Google Scholar]
- Tomasi, G.; van den Berg, F.; Andersson, C. Correlation optimized warping and dynamic time warping as preprocessing methods for chromatographic data. J. Chemometr. 2004, 18, 231–241. [Google Scholar]
- Savorani, F.; Tomasi, G.; Engelsen, S.B. icoshift: A versatile tool for the rapid alignment of 1D NMR spectra. J. Magn. Reson. 2010, 202, 190–202. [Google Scholar]
- Craig, A.; Cloarec, O.; Holmes, E.; Nicholson, J.K.; Lindon, J.C. Scaling and normalization effects in NMR spectroscopic metabonomic data sets. Anal. Chem. 2006, 78, 2262–2267. [Google Scholar]
- De Meyer, T.; Sinnaeve, D.; van Gasse, B.; Tsiporkova, E.; Rietzschel, E.R.; De Buyzere, M.L.; Gillebert, T.C.; Bekaert, S.; Martins, J.C.; van Criekinge, W. NMR-based characterization of metabolic alterations in hypertension using an adaptive, intelligent binning algorithm. Anal. Chem. 2008, 80, 3783–3790. [Google Scholar]
- Anderson, P.E.; Mahle, D.A.; Doom, T.E.; Reo, N.V.; DelRaso, N.J.; Raymer, M.L. Dynamic adaptive binning: an improved quantification technique for NMR spectroscopic data. Metabolomics 2011, 7, 179–190. [Google Scholar]
- Davis, R.A.; Charlton, A.J.; Godward, J.; Jones, S.A.; Harrison, M.; Wilson, J.C. Adaptive binning: An improved binning method for metabolomics data using the undecimated wavelet transform. Chemometr. Intell. Lab. 2007, 85, 144–154. [Google Scholar]
- Anderson, P.E.; Reo, N.V.; DelRaso, N.J.; Doom, T.E.; Raymer, M.L. Gaussian binning: A new kernel-based method for processing NMR spectroscopic data for metabolomics. Metabolomics 2008, 4, 261–272. [Google Scholar]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in H-1 NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar]
- Van den Berg, R.A.; Hoefsloot, H.C.; Westerhuis, J.A.; Smilde, A.K.; van der Werf, M.J. Centering, scaling, and transformations: improving the biological information content of metabolomics data. BMC Genomics 2006, 7. [Google Scholar] [CrossRef]
- Jackson, J.E. A User’s Guide to Principal Components; John Wiley and Sons: New York, NY, USA, 1991. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sundekilde, U.K.; Larsen, L.B.; Bertram, H.C. NMR-Based Milk Metabolomics. Metabolites 2013, 3, 204-222. https://doi.org/10.3390/metabo3020204
Sundekilde UK, Larsen LB, Bertram HC. NMR-Based Milk Metabolomics. Metabolites. 2013; 3(2):204-222. https://doi.org/10.3390/metabo3020204
Chicago/Turabian StyleSundekilde, Ulrik K., Lotte B. Larsen, and Hanne C. Bertram. 2013. "NMR-Based Milk Metabolomics" Metabolites 3, no. 2: 204-222. https://doi.org/10.3390/metabo3020204
APA StyleSundekilde, U. K., Larsen, L. B., & Bertram, H. C. (2013). NMR-Based Milk Metabolomics. Metabolites, 3(2), 204-222. https://doi.org/10.3390/metabo3020204