Abstract
Human coronaviruses (HCoVs) are known respiratory pathogens associated with a range of respiratory outcomes. In the past 14 years, the onset of severe acute respiratory syndrome coronavirus (SARS-CoV) and Middle East respiratory syndrome coronavirus (MERS-CoV) have thrust HCoVs into spotlight of the research community due to their high pathogenicity in humans. The study of HCoV-host interactions has contributed extensively to our understanding of HCoV pathogenesis. In this review, we discuss some of the recent findings of host cell factors that might be exploited by HCoVs to facilitate their own replication cycle. We also discuss various cellular processes, such as apoptosis, innate immunity, ER stress response, mitogen-activated protein kinase (MAPK) pathway and nuclear factor kappa B (NF-κB) pathway that may be modulated by HCoVs.
Keywords:
human coronavirus; virus–host interactions; apoptosis; innate immunity; ER stress; MAPK; NF-κB 1. Introduction
Human coronaviruses (HCoVs) represent a major group of coronaviruses (CoVs) associated with multiple respiratory diseases of varying severity, including common cold, pneumonia and bronchilitis [1]. Today, HCoVs are recognised as one of the most rapidly evolving viruses owing to its high genomic nucleotide substitution rates and recombination [2]. In recent years, evolution of HCoVs has also been expedited by factors such as urbanization and poultry farming. These have permitted the frequent mixing of species and facilitated the crossing of species barrier and genomic recombination of these viruses [3]. To date, six known HCoVs have been identified, namely HCoV-229E, HCoV-NL63, HCoV-OC43, HCoV-HKU1, severe acute respiratory syndrome coronavirus (SARS-CoV) and Middle East respiratory syndrome coronavirus (MERS-CoV); of which, four HCoVs (HCoV-229E, HCoV-NL63, HCoV-OC43 and HCoV-HKU1) are globally circulated in the human population and contribute to approximately one-third of common cold infections in humans [4]. In severe cases, these four HCoVs can cause life-threatening pneumonia and bronchiolitis especially in elderly, children and immunocompromised patients [1,5,6]. Besides respiratory illnesses, they may also cause enteric and neurological diseases [7,8,9,10,11].
SARS-CoV first emerged in 2002–2003 in Guangdong, China as an atypical pneumonia marked by fever, headache and subsequent onset of respiratory symptoms such as cough and pneumonia, which may later develop into life-threatening respiratory failure and acute respiratory distress syndrome [12]. Being highly transmissible among humans, it quickly spread across 29 countries, infecting more than 8000 individuals with a mortality rate of about 10% [13,14]. Originally, palm civets were thought to be the natural reservoir for the virus [15]. However, subsequent phylogenetic studies pointed to the bat origin of SARS-CoV based on sequences of SARS-like virus found in bats [16]. The MERS-CoV epidemic surfaced in Saudi Arabia in 2012 with similar clinical symptoms as SARS-CoV but with a much higher mortality rate of about 35% [17]. Unlike SARS-CoV, which exhibits super-spreader events, transmission of MERS-CoV is geographically limited [12]. In fact, reported cases of MERS-CoV often stem from outbreaks within the Middle Eastern countries or recent travel to the region [18,19].
Taxonomy, Genomic Structure and Morphology
CoVs are a group of large enveloped RNA viruses under the Coronaviridae family. Together with Artierivirdae and Roniviridae, Coronaviridae is classified under the Nidovirale order [20]. As proposed by the International Committee for Taxonomy of Viruses, CoVs are further categorized into four main genera, Alpha-, Beta-, Gamma- and Deltacoronaviruses based on sequence comparisons of entire viral genomes [21,22]. These CoVs can infect a wide variety of hosts, including avian, swine and humans. HCoVs are identified to be either in the Alpha- or Betacoronavirus genera, including Alphacoronaviruses, HCoV-229E and HCoV-NL63, and Betacoronaviruses, HCoV-HKU1, SARS-CoV, MERS-CoV and HCoV-OC43 (Table 1).

Table 1.
Classification of human coronavirus.
Under the electron microscope, the CoV virions appear to be roughly spherical or moderately pleomorphic, with distinct “club-like” projections formed by the spike (S) protein [23,24]. Within the virion interior lies a helically symmetrical nucleocapsid that encloses a single-stranded and positive sense RNA viral genome of an extraordinarily large size of about 26 to 32 kilobases [20]. The positive sense viral genomic RNA acts as a messenger RNA (mRNA), comprising a 5′ terminal cap structure and a 3′ poly A tail. This genomic RNA acts in three capacities during the viral life cycle: (1) as an initial RNA of the infectious cycle; (2) as a template for replication and transcription; and (3) as a substrate for packaging into the progeny virus. The replicase-transcriptase is the only protein translated from the genome, while the viral products of all downstream open reading frames are derived from subgenomic mRNAs. In all CoVs, the replicase gene makes up approximately 5′ two-thirds of the genome and is comprised of two overlapping open reading frames (ORFs), ORF1a and ORF1b, which encodes 16 non-structural proteins. The final one-third of the CoV genomic RNA encodes CoV canonical set of four structural protein genes, in the order of spike (S), envelope (E), membrane (M) and nucleocapsid (N). In addition, several accessory ORFs are also interspersed along the structural protein genes and the number and location varies among CoV species [25] (Figure 1).
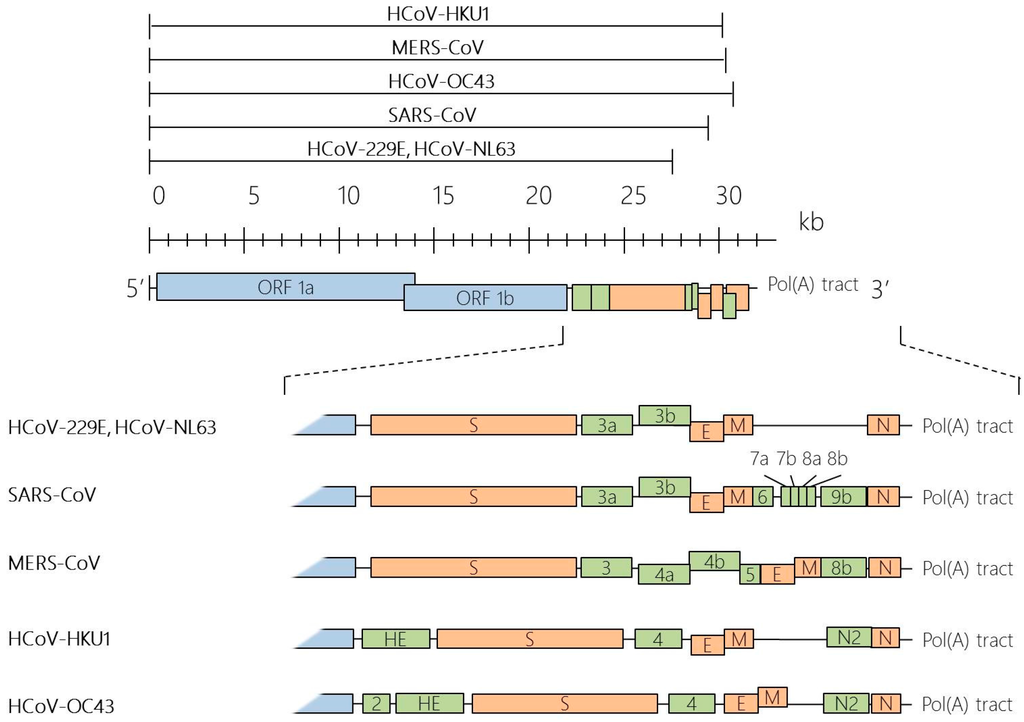
Figure 1.
Genome organisation of human coronaviruses (HCoVs). HCoV genomes range from about 26 to 32 kilobases (kb) in size, as indicated by the black lines above the scale. Coronavirus (CoV) genome is typically arranged in the order of 5′-ORF1a-ORF1b-S-E-M-N-3′. The overlapping open reading frames (ORF) ORF1a and ORF1b comprise two-thirds of the coronavirus genome, which encodes for all the viral components required for viral RNA synthesis. The other one-third of the genome at the 3′ end encodes for a set of structural (orange) and non-structural proteins (green).
2. Involvement of Host Factors in Viral Replication and Pathogenesis
As intracellular obligate parasites, HCoVs exploit the host cell machinery for their own replication and spread. Since virus–host interactions form the basis of diseases, knowledge about their interplay is of great research interest. Here, we describe what is currently known of the cell’s contribution in CoV infection cycle: attachment; entry into the host cell; translation of the replicase-transcriptase; replication of genome and transcription of mRNAs; and assembly and budding of newly packaged virions (Figure 2).
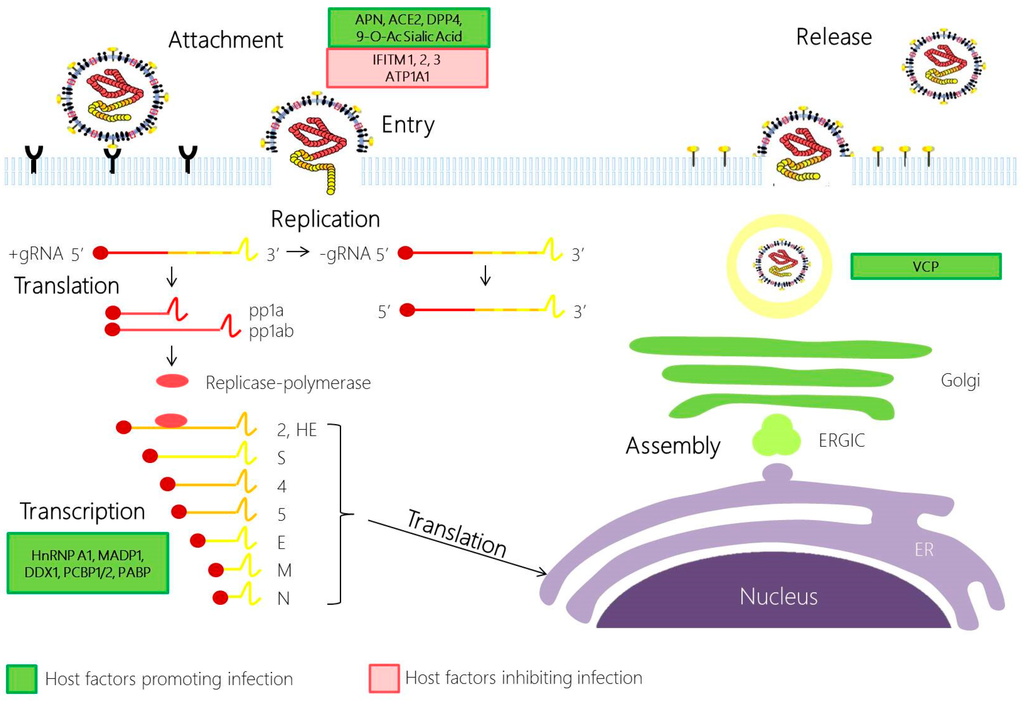
Figure 2.
Coronavirus replication cycle. Coronavirus infection begins with the attachment of the S1 domain of the spike protein (S) with its cognate receptor. This drives the conformational change in the S2 subunit in S, promoting the fusion of the viral and cell plasma membrane. Following the release of the nucleocapsid to the cytoplasm, the viral gRNA is translated through ribosomal frameshifting to produce polyproteins pp1a and pp1ab. pp1a and pp1ab are autoproteolytically processed by host and viral proteases to generate 16 non-structural proteins (NSPs), which will then be assembled to form the replicase-polymerase. The replicase-polymerase is involved in the coronaviral replication, a process in which the genomic RNA are replicated and the subgenomic RNA will be transcribed and translated to form the structural proteins. The viral products produced will be assembled in the ERGIC, and bud out as a smooth-wall vesicle to the plasma membrane to egress via exocytosis. Host factors that promote infection and inhibit infection are highlighted in green and red, respectively. APN, aminopeptidase N; ACE2, Angiotensin converting enzyme 2; DPP4, dipeptidyl peptidase 4; 9-O-Ac Sialic Acid, 9-O-Acetylated Sialic Acid; IFITM, Interferon induced transmembrane protein; ATP1A1, ATPase, Na+/K+ Transporting, Alpha 1 Polypeptide; HnRNP A1, Heterogeneous nuclear ribonucleoprotein A1; MADP1, Zinc Finger CCHC-Type and RNA Binding Motif 1; DDX1, ATP-dependent RNA Helicase; PCBP1/2, Poly r(C) binding protein 1/2; PABP, Poly A binding protein; COPB2, Coatomer protein complex, subunit beta 2 (beta prime); GAPDH, Glyceraldehyde 3-phosphate dehydrogenase; ERGIC, Endoplasmic reticulum Golgi intermediate compartment; ER, endoplasmic reticulum; VCP, Valosin-Containing Protein.
2.1. Coronavirus Attachment and Entry
CoV infection is initiated by the attachment to specific host cellular receptors via the spike (S) protein. The host receptor is a major determinant of pathogenicity, tissue tropism and host range of the virus. The S protein comprises of two domains: S1 and S2. The interaction between the S1 domain and its cognate receptor triggers a conformational change in the S protein, which then promotes membrane fusion between the viral and cell membrane through the S2 domain. Today, the main host cell receptors utilised by all HCoVs are known: aminopeptidase N by HCoV-229E [26], angiotensin-converting enzyme 2 (ACE2) by SARS-CoV [27] and HCoV-NL63 [28,29], dipeptidyl peptidase 4 (DPP4) by MERS-CoV [30] and 9-O-acetylated sialic acid by HCoV-OC43 and HCoV-HKU1 [31,32].
Apart from the conventional endosomal route of entry, some CoVs may also gain entry into the cell via the non-endosomal pathway, or a combination of both. The low pH in the cellular environment and endosomal cysteine protease cathepsins may help to facilitate membrane fusion and endosomal CoV cell entry [33]. Recent evidence has supported the role of cathepsin L in SARS-CoV and MERS-CoV entry [34,35,36]. Other host proteases, such as transmembrane protease serine 2 (TMPRSS2) and airway trypsin-like protease TMPRSS11D, could also perform S1/S2 cleavage to activate the S protein for non-endosomal virus entry at the cell plasma membrane during HCoV-229E and SARS-CoV infection [37,38]. In addition, MERS-CoV is also activated by furin, a serine endopeptidase that has been implicated in the cell entry of other RNA viruses and S1/S2 cleavage during viral egress [39].
Many host cells also utilise its own factors to restrict viral entry. Using cell culture system and pseudotype virus, many groups have identified a family of interferon inducible transmembrane proteins (IFITM), which could inhibit global circulating HCoV-229E and HCoV-NL63 S protein mediated entry, and also the highly pathogenic SARS-CoV and MERS-CoV [12,40]. While the IFITM mode of action remains elusive, cell-to-cell fusion assays performed by some research groups suggest that IFITM3 blocks the enveloped virus entry by preventing fusion of the viral envelope with the plasma membrane or endosomal membranes through modulating the host membrane fluidity [41].
2.2. Coronavirus Replication
Following the release and uncoating of viral nucleocapsid to the cytoplasm, CoV replication begins with the translation of ORF 1a and 1b into polyproteins pp1a (4382 amino acids) and pp1ab (7073 amino acids). Here, the downstream ORF1b is translated through ribosomal frameshifting mechanism, in which a translating ribosome shifts one nucleotide in the −1 direction, from the ORF1a reading frame into ORF1b reading frame. This repositioning is enabled by two RNA elements—a 5′-UUUAAAC-3′ heptanucleotide slippery sequence and RNA pseudoknot structure. Subsequently, polyproteins pp1a and pp1ab are cleaved into at least 15 nsp, which assemble and form the replication-transcription complex. With the assembly of the replicase-polymerase, the full-length positive strand of genomic RNA is transcribed to form a full-length negative-strand template for the synthesis of new genomic RNAs and overlapping subgenomic negative-strand templates. These subgenomic mRNAs are then transcribed and translated to produce the structural and accessory proteins. Several heterologous nuclear ribonucleoprotein (hnRNA) family members (hnRNPA1, PTB, SYN-CRYP) have been found to be essential for efficient RNA replication [42]. Other RNA-binding proteins have also been suggested to play a role in CoV replication, such as m-aconitase and poly-A-binding protein (PABP), DDX1, PCBP1/2 and zinc finger CCHC-type and RNA-binding motif 1 (MADP1) [43,44,45].
2.3. Coronavirus Assembly and Egress
The assembly of virions is quickly ensued with the accumulation of new genomic RNA and structural components. In this phase of the infection cycle, the helical nucleocapsid containing the genomic RNA interacts with other viral structural proteins (S, E and M proteins) to form the assembled virion. The assembly of CoV particles is completed through budding of the helical nucleocapsid through membranes early in the secretory pathway from the endoplasmic reticulum to the Golgi intermediate compartment (ERGIC). The contributions of the host in this phase of the infection cycle have rarely been explored. Currently, it is known that the M protein orchestrates the entire assembly process by selecting and organizing the viral envelope components at the assembly sites and by mediating the interactions with the nucleocapsid to allow the budding of virions [46]. The M protein interacts with different viral structural proteins, such as the E protein, to assemble into a mature virus. This interaction generates the scaffold of the virion envelope and induces the budding and release of the M protein-modified membrane and with the S protein to assemble the spikes into the viral envelope [46,47]. Following assembly and budding, the virions are transported in vesicles and eventually released by exocytosis. In a recent study, an inhibition of a Valosin-containing protein (VCP/p97) resulted in virus accumulation in early endosome in infectious bronchitis virus (IBV), suggesting a role for VCP in the maturation of virus-loaded endosomes [48].
3. Human Coronavirus Infection and Apoptosis
Apoptosis is a process of programmed cell death that is tightly regulated and anti-inflammatory. When cells undergo apoptosis, they demonstrate specific hallmarks such as cell shrinkage, extensive plasma membrane blebbing, nuclear pykosis, DNA fragmentation and asymmetrical distribution of plasma membrane [49,50,51]. To date, two main mechanisms of apoptosis have been established—the extrinsic and intrinsic pathways. The extrinsic pathway is initiated by the binding of extracellular death ligands (such as Fas ligand (FasL) and TNF-receptor-related apoptosis-inducing ligands (TRAIL)) to death receptors from the tumour necrosis factor (TNF) super-family [52]. These death receptors then recruit various death adapter proteins, such as Fas-associated death domain protein (FADD) [53], and initiator procaspases 8 and 10 to form the death-inducing signalling complex (DISC) [54,55]. Consequently, the two initiator procapases are cleaved to their active forms and induce a signalling cascade to eventually activate effector caspases 3 and 7. On the other hand, the intrinsic pathway occurs internally in the cell and involves changes in the mitochondrial outer membrane permeability (MOMP) based on the ratio of pro-apoptotic and anti-apoptotic B-cell lymphoma 2 (Bcl2) family of proteins (Figure 3). Enhanced MOMP causes the release of pro-apoptotic factors such as cytochrome c to activate initiator caspase 9. Like the extrinsic pathway, activation of initiator caspase 9 in the intrinsic pathway results in the proteolytic cleavage effector caspases 3 and 7, which in turn process many key cellular proteins essential for apoptosis [56]. Convergence between the two pathways may also occur even before effector caspase activation, when Bid, a pro-apoptotic Bcl2 family protein, is directly cleaved by caspase 8 [57].
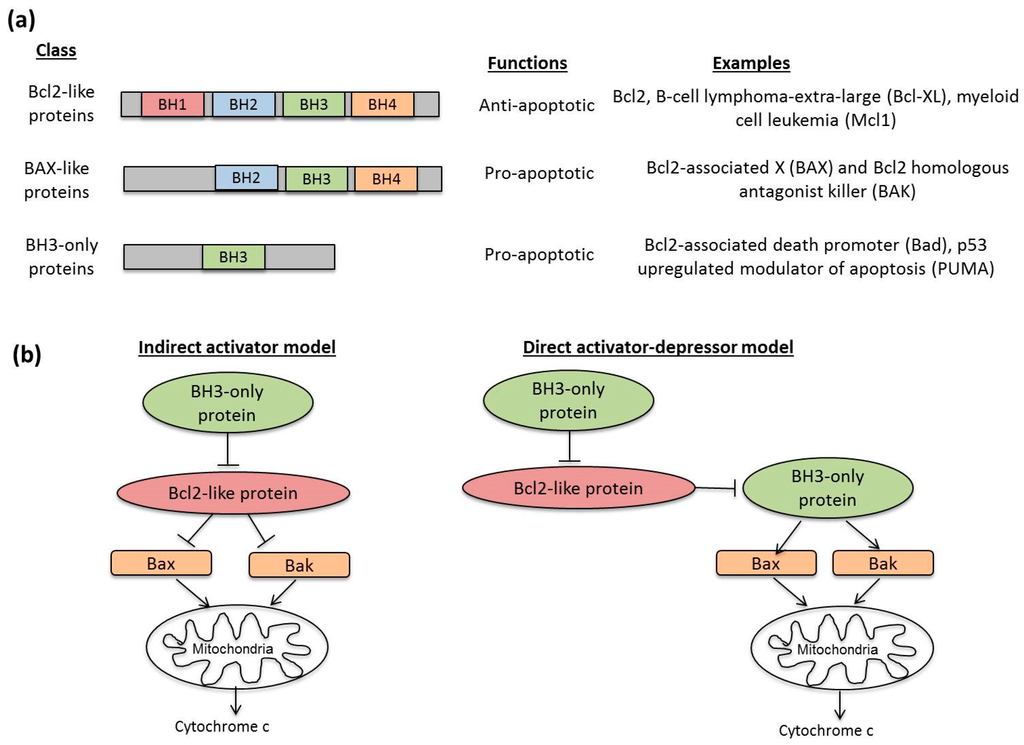
Figure 3.
Regulation of MOMP by Bcl2 family of proteins. (a) The Bcl2 family of proteins is categorized into three main classes according to their functions and number of Bcl2 homology (BH) domains. The pro-survival Bcl2-like family members (Bcl2, B-cell lymphoma-extra-large (Bcl-XL), myeloid cell leukemia (Mcl1)) contain all four BH domains and are anti-apoptotic. A second class, known as Bcl2-associated X (BAX)-like proteins, which includes BAX and Bcl2 homologous antagonist killer (BAK), is pro-apoptotic and lacks the BH4 domain. Finally, the third class, known as BH3-only proteins (Bid, Bcl2-associated death promoter (Bad), and p53-upregulated modulator of apoptosis (PUMA)), contain only BH3 domain and is pro-apoptotic. (b) Two models have been proposed to account for the role of the Bcl2 family proteins in MOMP—the indirect activator model and direct activator–depressor model [11]. In the indirect activator model, the anti-apoptotic Bcl2-like proteins suppress the insertion of Bax-Bak pore complex into the mitochondria to promote MOMP and release of cytochrome c. However, when BH3-only proteins are activated beyond a certain threshold, the inhibitory effects of Bcl2-like proteins can be subverted. In the direct activator–depressor model, BH3-only protein acts as direct activators to induce Bak-Bak insertion into the outer mitochondrial membrane. These BH3-only proteins can be suppressed by Bcl2-like protein, which can in turn be inhibited by another subset of BH3-only proteins. This figure is modified from [12].
During viral infections, apoptosis is induced as one of the host antiviral responses to limit virus replication and production. Many viruses have evolved distinct strategies to subvert apoptosis [58]. For example, some viruses encode for viral proteins that act as Bcl2 family protein homologues [59]. Alternatively, viruses might develop mechanisms to regulate Bcl2 family proteins or caspase activation either directly or indirectly through other molecular pathways such as mitogen-activated protein (MAPK) and nuclear factor kappa B (NF-κB) pathways [60,61,62,63,64,65]. Interestingly, some viruses may engage the apoptotic machineries for efficient viral infection. For instance, alphaviruses and flaviviruses contain phosphatidylserine-rich viral membranes to imitate apoptotic cells to promote viral entry [66].
3.1. Cell Tropism and Apoptosis
As HCoVs are respiratory pathogens known to infect tissue cultures and cell lines derived from the respiratory tract, these viruses may also infect other tissue cultures and cell lines. Infection of these tissues and cells may induce apoptosis [67,68]. However, although HCoVs mainly target the respiratory tract during infection, they have also been associated to apoptosis induction in a wide spectrum of cell types, including intestinal mucosal cells, kidney tubular cells and neuronal cells [69,70,71,72,73,74]. Autopsy studies of SARS-CoV-infected tissues revealed apoptosis induced in lung, spleen and thyroid [75]. HCoVs have also been shown to infect the immune system and induce apoptosis in immune cells such as macrophages, monocytes, T lymphocytes and dendritic cells [69,76,77,78,79]. Because these immune cells are associated with the activation of the innate and acquired immunity, it is reasonable to speculate that the massive elimination of these cells could be a viral strategy to suppress the host innate and adaptive immune responses. In a recent study, it was reported that HCoV-229E infection resulted in massive CPE and cell death in dendritic cells [80]. Since dendritic cells are prevalent throughout our bodies, it is possible that they are used as a vehicle to facilitate viral spread and impair our immune systems [80,81].
3.2. Molecular Mechanisms in Apoptosis
On a molecular level, HCoV infections have been reported to trigger apoptosis through multiple mechanisms. SARS-CoV-induced apoptosis was shown to be caspase-dependent and could be inhibited by caspase inhibitor Z-VAD-FMK or overexpression of Bcl2 [82,83]. Although viral replication was required for apoptosis induction [83], apoptosis did not affect the viral replication kinetics of SARS-CoV [82]. On the other hand, infection of primary T lymphocytes by MERS-CoV was shown to induce DNA fragmentation and caspase 8 and 9 activation, suggesting that both extrinsic and intrinsic pathways were activated. Unlike SARS-CoV infection, MERS-CoV replication was not necessary to induce apoptosis in infected T lymphocytes [79]. Apoptosis can also be induced by the less pathogenic strains of HCoVs, as substantiated by microarray data showing significant changes in pro-apoptotic and anti-apoptotic gene expression of Bcl2 family members during HCoV-229E infection [84]. Infection of HCoV-OC43 was shown to promote BAX translocation to the mitochondria in human neuronal cells [74]. Although caspases 3 and 9 were activated in HCoV-OC43-infected murine and human neuronal cells [9,74], addition of pan-caspase inhibitor Z-VAD-FMK and the caspase-9 inhibitor Z-LEHD-FMK did not affect the viability of these infected neuronal cells, indicating that programmed cell death induced by HCoV-OC43 could be caspase-independent [74]. This highlights the possibility of a non-classical programmed cell death mechanism induced in HCoV infection.
Apoptotic mechanisms during HCoV infection are likely to be manipulated by viral proteins (Figure 4), although this has only mostly been studied in SARS-CoV. Specifically, SARS-CoV S, N, E, M, ORF-6, 7a and 9b proteins have been shown to serve pro-apoptotic functions in their host cells [77,85,86,87,88,89,90,91]. Expression of SARS-CoV E protein and 7a protein promoted mitochondrial-mediated apoptosis by sequestering the anti-apoptotic Bcl-XL protein to the endoplasmic reticulum (ER) membranes [77,92]. SARS-CoV M protein is also highly pro-apoptotic and mediates activation of both caspases 8 and 9 [90]. Additionally, HCoV-OC43 wild type S protein has been shown to induce unfolded protein response (UPR) in human neuronal NT2-N and LA-N-5 cell lines, which may lead to apoptosis [93]. A recombinant HCoV-OC43 harbouring point mutations at its S protein induced stronger caspase 3 activation and nuclear fragmentation than the wild-type virus [93]. It is interesting to note that the localisation of SARS-CoV N and 9b protein is associated with induction of caspase-dependent apoptosis [89,94]. This finding opens up to novel perspectives of the link between subcellular localisation of viral proteins and caspase activation as a mode of apoptosis regulation by HCoVs.
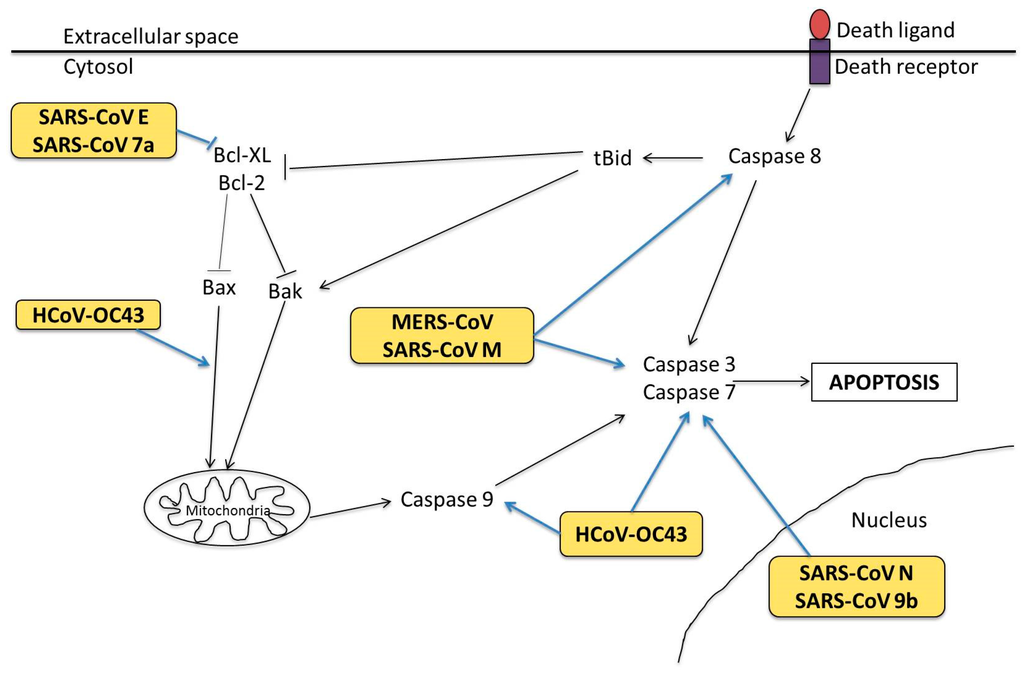
Figure 4.
Activation of apoptosis by HCoVs. Binding of death ligands to death receptor induces caspase 8 activation, which in turn activates effector caspases 3 and 7 to stimulate apoptosis. On the other hand, intrinsic pathway is regulated by pro-apoptotic and anti-apoptotic Bcl2 family proteins, such as Bcl-XL, Bcl2, Bax and Bak to induce MOMP. Subsequent caspase 9 activation caused by enhanced MOMP stimulates caspases 3 and 7 activation. During HCoV infection, the virus or specific viral proteins (yellow-orange boxes) target at multiple stages of both the extrinsic and intrinsic apoptosis signalling pathways.
4. Human Coronavirus Infection and Innate Immunity
When the cells are exposed to pathogens such as viruses, immune responses are induced as a form of host defence. The immune response is modulated during pathogen exposure in a cell-type dependent fashion. Innate immunity is the first line of defence mounted against the virus before the adaptive immune system is generated. Both the host and virus can manipulate innate immune mechanisms as a form of defence or evasion strategy [95,96].
4.1. Pattern Recognition Receptors
Cells in the immune system detect the viral pathogens via several recognition strategies. Of which, the most well characterized is the pattern recognition receptors (PRR), which engage various microbial pathogens via evolutionarily conserved structures known as pathogen-associated molecular patterns (PAMPs). PRRs are mainly categorized into three classes, namely Toll-like receptors (TLRs), retinoic acid-inducible gene I (RIG-I)-like receptors (RLRs) and nucleotide oligomerisation domain (NOD)-like receptors (NLRs).
TLR is a type I transmembrane protein localized to either the cell surface or endosomal vesicles. Their leucine-rich repeats (LRR) domain mediates the recognition of PAMPs and damage-associated molecular patterns (DAMPs) from various sources including bacteria, fungi and viruses [97]. Activation of TLRs occurs mainly in antigen-presenting cells such as dendritic cells (DCs), macrophages, monocytes and B cells. Of the 10 known TLRs in human, TLR2, 3, 4, 7 and 9 are found to be involved in viral detection [98,99]. TLR3 recognizes double-stranded RNA (dsRNA), a replicative intermediate produced during viral RNA replication [100]. TLR7 and 8 detect single-stranded RNA (ssRNA) and TLR9 recognizes unmethylated CpG DNA present in DNA viruses [101,102,103]. Besides nucleic acids, other TLRs, such as TLR2 and 4, sense viral proteins as exemplified in respiratory syncytial virus (RSV), hepatitis virus, measles virus and human immunodeficiency virus [104,105,106,107]. Upon recognition of viral components, TLRs recruit Toll/interleukin-1 receptor (TIR)-containing signalling adaptor molecules, such as MyD88 (myeloid differentiation primary response protein 88) and TIR-domain-containing adapter-inducing interferon-β (TRIF) [108,109,110]. MyD88 and TRIF then stimulate the MAPK and NF-κB pathways to boost IFN and pro-inflammatory cytokine production [111].
Unlike TLRs, RLRs and NLRs are expressed ubiquitously. RLRs are a family of cytoplasmic receptors that comprise of three members: retinoic acid-inducible gene I (RIG-I), melanoma differentiation associated factor 5 (MDA5), and laboratory of genetics and physiology 2 (LGP2). RIG-I and MDA5 possess two caspase-recruitment domains (CARDs) at their N terminal, a DExD/H-box RNA helicase domain (where x can be any amino acid) and a repressor domain (RD) at the C terminal. On the other hand, LGP2 lacks the CARD domain [112], and it regulates RIG-I and MDA5 either positively or negatively [113,114,115]. RIG-I recognizes 5′-triphosphate moieties present in viral genomic RNA, as well as double-stranded “panhandle” structure formed by self-annealing of complementary ends of the viral genome [116,117]. In contrast, MDA5 usually detects dsRNA sequences of longer length. Binding of RIG-I and MDA5 to the viral RNA causes a conformational change to expose the CARD domain. An adaptor protein known as mitochondrial antiviral signalling adaptor, MAVS, which is localized at the mitochondria and peroxisomes, is recruited. MAVS then activates transcription factors such as interferon regulatory factor 1 (IRF1), IRF3 and NF-κB to trigger the expression of interferons (IFNs) and pro-inflammatory cytokines [117] (Figure 5).
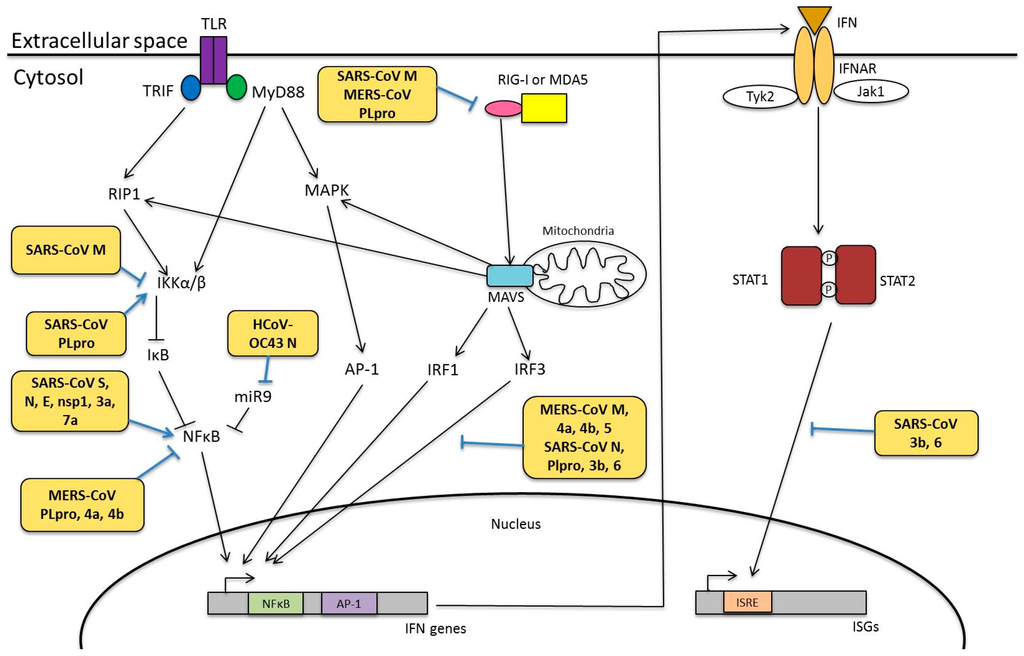
Figure 5.
HCoV viral proteins on innate immunity. During HCoV infection, PRRs such as TLRs, RIG-I and MDA5 are activated to trigger a series of signalling pathway, including MAPK and NF-κB, for IFN production. These IFNs then act on IFNAR and activate the JAK-STAT signalling pathway to induce ISGs. The yellow-orange boxes show the viral proteins that have been reported to modulate host innate immunity at multiple stages.
NLRs are another large family of cytosolic proteins that are organized into three main domains: a CARD domain, pyrin domain (PYD), or baculovirus inhibitor repeat domain at the N terminal, a conserved NOD motif at the intermediate region and LRR motifs at the C terminal. The LRR motifs detect viral PAMPs to induce a structural rearrangement. Subsequently, a diverse range of signalling pathways, including MAPK and NF-κB pathways, is activated [118], Furthermore, assembly of multimeric protein complexes known as inflammasomes is mediated by some NLR family members such as NLR family PYD-containing 1 (NLRP1), NLRP3 and NLR family CARD-containing 4 (NLRC4). These inflammasomes activate inflammatory caspase, capase-1, that induces the cleavage of pro-IL-1β and pro-IL-18 into their active forms [119].
4.2. Interferon Responses
IFNs are classified into type I, II and III based on their preference for specific IFN receptors. In particular, type I IFN is best known for its antiviral actions. Binding of type I IFNs to the IFN-α/β receptor (IFNAR) induces the oligomerisation of its receptor subunits, IFN-αR1 and IFN-αR2, and consequently conveys downstream signalling via the Janus kinase-Signal Transducer and Activator of Transcription (JAK-STAT) pathway. Autophosphorylation of JAK domains in the IFNAR results in subsequent phosphorylation of STAT1 and STAT2 proteins at their tyrosine residues. This is followed by dimerisation and nuclear translocation of the activated STAT proteins, which recruit IFN regulatory factor 9 (IRF9) to form the IFN-stimulated gene factor 3 (ISGF3). ISFG3 is a transcription factor that binds to its cognate DNA sequence known as IFN-stimulated response elements (ISREs) to activate transcription of IFN-stimulated genes (ISGs) [120] (Figure 5). Many of these ISGs such as 2′–5′ oligoadenylatesynthetase and protein kinase R (PKR) confer resistance against virus invasions [121]. Additionally, type I IFNs facilitates the maturation of dendritic cells (DCs), cytotoxicity of natural killer (NK) cells, and differentiation of T lymphocytes [98].
4.3. Modulation of Innate Immunity
Infection by HCoVs, especially the highly pathogenic SARS-CoV and MERS-CoV, is associated with suppression of IFN synthesis [122,123,124,125,126]. The capability of the virus to regulate type I IFN signalling is an important hallmark for virulence [127]. As compared to SARS-CoV and MERS-CoV, a huge rise in type I IFNs were detected in cells infected with HCoV strain 229E [80,124,128].
Based on studies from SARS-CoV and Mouse Hepatitis Virus (MHV)-infected cells, two mechanisms have been proposed to explain the HCoV-mediated inhibition of type I IFN production [13,126]. Firstly, CoV genomic and subgenomic RNA replication takes place in double membrane vesicles to prevent detection by PRRs [13,129]. Secondly, proteins encoded by the virus could interfere with innate immune pathways [13,130]. The structural proteins, nonstructural proteins and accessory proteins of HCoVs have been shown to modify innate immune responses (Figure 5).
4.3.1. Viral Proteins Involved in Innate Immunity
Structural Proteins of HCoVs
Expression of SARS-CoV M protein could suppress type I IFN production mediated by RIG-I, but not MDA5, in infected HEK293 cells [131], likely through its first transmembrane domain. However, this inhibition was not observed when expressing the M protein of HCoV-HKU1, suggesting that this activity is not conserved among all HCoV strains [132]. In another study, it was shown that the MERS-CoV M protein could also suppress type I IFN by inhibiting the translocation of IRF3 into the nucleus, although the exact mechanism has not yet been elucidated [133]. Additionally, SARS-CoV N protein was also shown to interfere with the function of IRF3 [134]. The N protein of SARS-CoV likely act at the initial recognition stage of viral RNA via its RNA binding activity, although it neither forms a complex with RIG-I nor MDA5 [135]. This implies that the N protein possibly acts on other viral RNA recognition strategies of the host.
Non-Structural and Accessory Proteins of HCoVs
Besides the structural proteins, other nonstructural proteins (nsp) and accessory proteins of HCoVs have also been implicated in the modulation of innate immunity. For instance, nsp1 of both SARS-CoV and MERS-CoV has been demonstrated to modify capped non-viral RNAs to facilitate endonucleolytic cleavage of host messenger RNA (mRNA) [136,137]. Additionally, SARS-CoV nsp1 interacted with the 40S subunit of ribosome to prevent host mRNA translation [136]. This induces the host shutoff mechanism, as transcription and translation of viral RNA are more favoured over that of host mRNA. In a recent study, several residues of SARS-CoV nsp1 were identified to affect IFN-dependent signalling [138]. In addition to nsp1, SARS-CoV and MERS-CoV nsp3 proteins, which possess papain-like protease (PLpro) domain and a PLP2 domain, also antagonize IFN production. Both SARS-CoV and MERS-CoV PLpro domains are deISGylating enzymes and they downregulated mRNA levels of pro-inflammatory cytokines including CCL5, IFNβ, and CXCL10 [139]. Suppression of IFN responses by SARS-CoV PLpro is not mediated by its protease activity. Rather, SARS-CoV PLpro inhibited the phosphorylation of interferon-regulatory factor 3 (IRF3) and its translocation to the nucleus to enhance IFN gene transcription [140]. Expression of MERS-CoV PLpro also antagonizes IFN production and is required for suppression on RIG-I and MDA5 [139,141]. Furthermore, it has been identified that the ADP-ribose-1-monophosphatase macrodomain encoded within nsp3 in HCoV-229E and SARS-CoV is responsible for suppressing IFN induction [142].
Despite being dispensable in viral replication, HCoV accessory proteins are essential in diverse cellular signalling, such as cell proliferation, apoptosis and interferon signalling [25]. In SARS-CoV, ORF3b and -6 are shown to interfere with IFNβ synthesis by inhibiting the phosphorylation and nuclear translocation of IRF3. Furthermore, these accessory proteins also disrupt IFN signalling by preventing IFNβ-induced activation of interferon-stimulated response element (ISRE) found in the promoter region of ISG [134]. The accessory proteins of MERS-CoV, ORF4a, -4b and -5, could similarly suppress IRF3 nuclear translocation, hence significantly reducing IFN-β promoter-driven luciferase activity in cells transfected with these accessory proteins [133].
5. Human Coronavirus and ER Stress Response
The endoplasmic reticulum (ER) is a cellular organelle important for protein synthesis, folding, processing and post-translational modifications. In normal circumstances, the ER can be loaded with a very high concentration of proteins without perturbing its unique luminal environment [143]. However, when the protein load exceeds the ER folding and processing capacity, rapid accumulation of misfolded or unfolded proteins occurs within the ER lumen. Various signalling pathways, collectively known as ER stress response or UPR, are activated. These pathways are initiated by three ER transmembrane sensors-protein-kinase-R (PKR)-like endoplasmic reticulum kinase (PERK), inositol-requiring protein 1 (IRE1) and activating transcriptional factor 6 (ATF6) to orchestrate the restoration of ER homeostasis by enhancing protein folding, attenuating protein translation and upregulating genes related to protein folding, chaperoning and ER-assisted degradation (ERAD) (Figure 6). In cases of prolonged and irreversible ER stress, apoptosis mechanisms are triggered [144]. During viral infections, ER stress response is induced. This massive utilisation of the ER elicit immense burden, causing the host to mount UPR as its antiviral response [145].
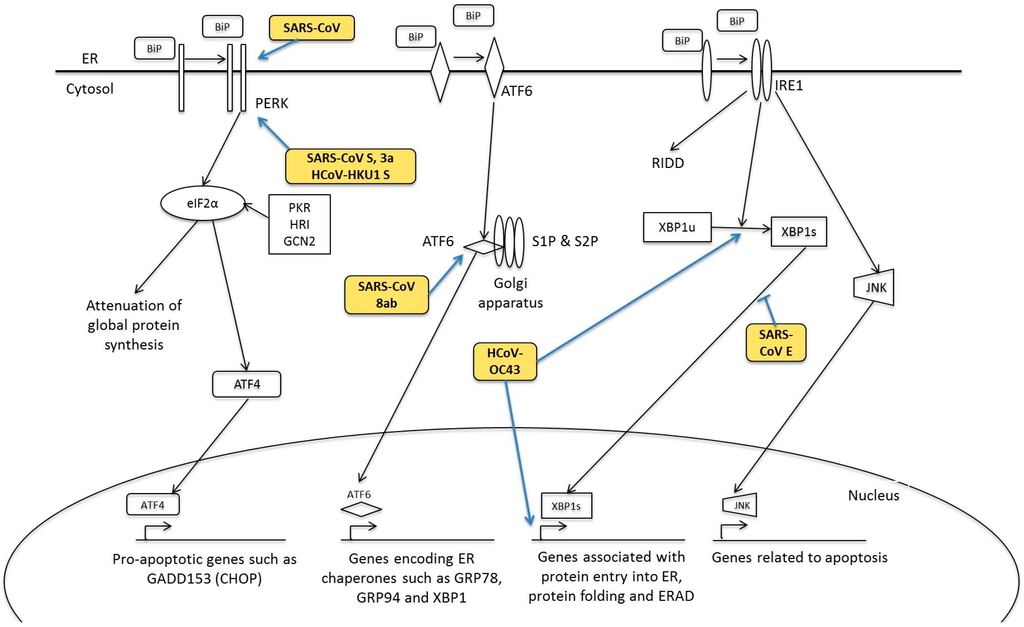
Figure 6.
HCoVs on ER stress response. During HCoV infections, the ER stress response, which comprises of three signalling pathways, PERK, ATF6 and IRE1, is activated. HCoVs encode many viral proteins (yellow-orange boxes) that target the various signalling pathway of ER stress during viral infections.
5.1. PERK Signalling Pathway
Activation of PERK is initiated by its dissociation of the luminal domain from the ER chaperone, binding immunoglobulin protein (BiP). This is followed by the oligomerisation and autophosphorylation of PERK. In its active form, PERK phosphorylates Ser51 at the α-subunit of eukaryotic initiation factor 2 (eIF2α) to attenuate protein translation [146]. Activation of PERK plays a pro-survival role in cells, as clearly demonstrated by PERK−/− mouse embryonic fibroblasts, which exhibited higher cell death when treated with cycloheximide, an ER stress-inducing agent [147]. Phosphorylated eIF2α not only triggers a shutdown of global protein synthesis, but also enhances the translation of activating transcription factor ATF4 [148]. ATF4 stimulates target gene expression such as GADD153 (also known as CHOP or C/EBP-homologous protein), to enhance transcription of pro-apoptotic genes [149]. Additionally, eIF2α can be phosphorylated by other kinases such as PKR, heme-regulated inhibitor kinase (HRI), and general control non-derepressible-2 (GCN2) [144]. These kinases activate various downstream signalling pathways, which together form the integrated stress response [146,150].
PKR and eIF2α phosphorylation was detected in SARS-CoV-infected cells and inhibition of PKR using antisense peptide-conjugated phosphorodiamidate morpholino oligomers did not affect eIF2α phosphorylation but significantly reduced SARS-CoV-induced apoptosis. SARS-CoV protein replication and virus production were not affected by PKR knockdown. Therefore, it is likely that SARS-CoV adopts a strategy to counteract against the antiviral effects of PKR, thus enabling viral mRNA translation to proceed regardless of eIF2α phosphorylation. PERK was also found to be activated during SARS-CoV infection [75], possibly through its S and 3a proteins [151,152]. In another study, it was demonstrated that expression of a dominant-negative PERK mutant, that inhibited PERK kinase activity, suppressed the transcriptional activation of Grp78 and Grp94 promoters mediated by S proteins of SARS-CoV and HCoV-HKU1 [153]. However, PERK activation is unlikely to occur in all HCoV strains. In neuronal cells lines infected with HCoV-OC43, it was shown that eIF2α was only transiently phosphorylated at the early stage of infection, but was subsequently suppressed and returned back to its basal level of phosphorylation, similar to the mock-infected cells [93]. On the other hand, previous studies from this group showed that PKR, PERK and eIF2α were moderately induced at the early stages of IBV infection, but were subsequently suppressed at late infection stages [154,155]. Nevertheless, the moderate and transient increase in eIF2α phosphorylation was sufficient to activate ATF4 protein translation and upregulate the downstream targets of ATF4, ATF3 and GADD153. Knockdown of PKR and PERK in IBV-infected cells attenuated IBV-induced GADD153 upregulation and IBV-induced apoptosis, although the viral protein replication was unaffected [155]. Upregulation of GADD153 is postulated to induce pro-apoptotic protein TRIB3 and inhibit pro-survival ERK protein [154], as well as provide a negative feedback to rapidly dephosphorylate eIF2α at late stages of IBV infection [155]. Based on these findings, we speculate the HCoVs might use similar mechanism to modulate the PKR/PERK/eIF2α pathway in infected cells. More studies could be done on HCoV infection to analyze the activation of the PKR/PERK/eIF2α pathway at various stages of infection.
5.2. ATF6 Signalling Pathway
Like PERK, activation of ATF6 is initiated by dissociation from the ER chaperone, BiP, although alternative mechanisms such as deglycosylation and reduction of disulphide bonds could occur [156,157]. ATF6 then translocates into the Golgi apparatus, where it is proteolyzed by site-1 and site-2 proteases (S1P and S2P). The processed ATF6 then migrates to the nucleus where it turns on expression of genes containing an ER stress response element (ERSE) in their promoters [158]. Like ATF4, ATF6 also induces expression of ER chaperone proteins such as GRP78, GRP94 and transcription factors CHOP and X box-binding protein 1 (XBP1) [150]. XBP1 is essential for IRE signalling [159].
As compared to the two other UPR branches, PERK and IRE1, the ATF6 branch is less well studied. As GRP94/78 are also target genes of ATF6, and their promoter activities were enhanced by SARS-CoV S protein, one could hypothesize that ATF6 pathway could also be induced by SARS-CoV S. Surprisingly, overexpression of SARS-CoV S protein did not affect ATF6 promoter luciferase activity [152]. Deletion of E protein in recombinant SARS-CoV also did not significantly activate ATF6 [91]. Intriguingly, 8ab protein, an accessory protein of SARS-CoV, was shown to reside in the luminal surface of the ER surface and activate ATF6 via facilitating its proteolysis and translocation of the processed ATF6 into the nucleus [160]. 8ab protein from SARS-CoV was found in civet cats and early human isolates, but was subsequently split into two accessory proteins, 8a and 8b, with a characteristic 29-nucleotide deletion [161].
5.3. IRE1 Signalling Pathway
IRE1 is believed to be the last UPR branch to be activated in cells undergoing ER stress [162]. It is also the most conserved among all UPR arms [163]. Although IRE1 was initially proposed to be activated in the same mechanism as PERK [162], later studies suggested that the N-terminal luminal domain (NLD) of IRE1 can directly bind unfolded proteins [164,165]. Activation of its RNase domain results in unconventional splicing of a 252-nucleotide intron from homologous to Atf/Creb1 (HAC1) mRNA in yeasts and a 26-nucleotide intron from X-box binding protein 1 (XBP1) mRNA in humans [166]. Splicing of XBP1 generates a potent transcription factor, XBP1s, that induces expression of genes related to protein entry into the ER, folding and ERAD [159]. In a negative feedback mechanism, XBP1s also promotes the transcription of E3 ubiquitin ligase synoviolin to enhance IRE1 ubiquitination [167]. The unspliced variant XBP1u contained a nuclear exclusion signal to sequester XBP1s from the nucleus, thus making XBP1u another negative feedback regulator of XBP1s. [168]. In a separate mechanism, IRE1 can cleave ER-associated mRNA species through regulated IRE1-dependent decay (RIDD) during late stages of ER stress [145]. It is believed that initial XBP1/HAC1 splicing by IRE1 promotes survival but subsequent activation of RIDD upon prolonged ER stress leads to cell death, thus allowing IRE1 to play dual role in apoptosis [169,170]. Another important enzymatic activation of IRE1 is its kinase activity. The kinase domain of phosphorylated IRE1 recruits the TNF receptor-associated factor 2 (TRAF2), which then activates other kinases to eventually activate the c-Jun N-terminal kinase (JNK) and regulates ER stress-dependent apoptosis [171].
Previous studies have investigated the role of IRE1-XBP1 pathway during SARS-CoV infection. Although no increase in XBP1 splicing was observed in SARS-CoV-infected cells [172], deletion of E protein in recombinant SARS-CoV resulted in significant XBP1 splicing and higher rate of apoptosis [91]. On the other hand, infection of HCoV-OC43 caused an induction in XBP1 splicing and enhanced expression of genes regulated by XBP1s, namely Edem, Herp, Grp94 and P58-ipk. However, introduction of two point mutations (H183R and Y241H) in the S protein of HCoV-OC43 led to a higher degree of XBP1 cleavage, followed by a strong activation of caspase-3 and nuclear fragmentation [93]. Since IRE1 pathway is closely associated to JNK activation, it is possible that the JNK pathway is also implicated during HCoV-OC43 infection.
Similar to HCoV infections, it has been shown that the IRE1-XBP1 pathway is activated during IBV infection. Knockdown of IRE1 using specific siRNA in IBV-infected cells augmented IBV-induced apoptosis; however, an opposite effect was observed by XBP1 knockdown in IBV-infected cells. Consistent with the knockdown experiments, transient overexpression of the full-length IRE1α attenuated IBV-induced apoptosis. When both spliced and unspliced forms of XBP1 were overexpressed in IBV-infected cells, the spliced form of XBP1 was shown to be anti-apoptotic and the unspliced form was pro-apoptotic. Overexpression of a dominant-negative XBP1 enhanced IBV-induced apoptosis. Therefore, our findings showed that the anti-apoptotic function of IRE1 during IBV infection could be mediated by its splicing of XBP1, hence converting XBP1 from a pro-apoptotic to anti-apoptotic form. Finally, IRE1 induction during IBV infection was shown to mediate JNK hyperphosphorylation and Akt hypophosphorylation to potentiate the IBV-infected cells to apoptosis [173].
6. Human Coronavirus and MAPK Pathways
The MAPKs are a group of evolutionally conserved serine/theronine kinases that are activated in response to environmental stresses including oxidative stress, DNA damage, cancer development and viral infections [174,175,176]. To date, multiple MAPK pathways have been identified in mammals, and they can be broadly classified into three major categories—the extracellular signal-regulated kinase (ERK), p38 MAPK and stress-activated protein kinase/c-Jun N-terminal kinase (SAPK/JNK) [177]. In all MAPK pathways, the signals are transduced downstream via a three-tier protein kinase cascade. In each tier, the kinases are activated by upstream kinases by dual phosphorylation at the Thr-X-Tyr motif (X represents any amino acid). Presence of extracellular stimuli triggers the activation of MAP kinase kinase kinases (MAPKKKs), which then activate the MAP kinase kinases (MAPKKs). These sequential phosphorylation events eventually activate MAPKs which in turn regulate a variety of fundamental cellular processes such as cell proliferation, survival, motility, differentiation, autophagy, apoptosis and regulation of cytokine production (Figure 7) [178].
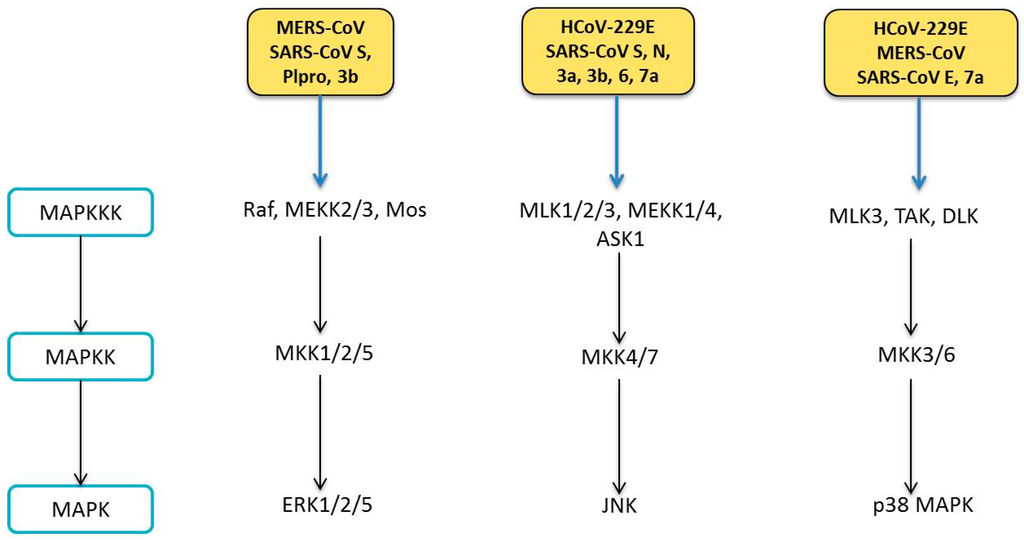
Figure 7.
HCoVs on MAPK signalling pathways. MAPK pathways comprises of the ERK, JNK and p38 MAPK pathways. During HCoV infections, the signals are transduced by the MAPK pathways by a three-tier protein kinase cascade, with the kinase of each tier being phosphorylated by upstream kinases at the Thr and Tyr residues. HCoVs and their viral proteins (yellow-orange boxes) have been shown to induce these MAPK pathways as shown in the figure.
6.1. Modulation of MAPK Pathways
Phosphorylation of all three MAPK members has been detected in cells infected with SARS-CoV [179,180]. Additionally, the MAPK pathways are also activated during infection by other HCoVs, as discussed below (Figure 7).
6.1.1. ERK Pathway
Activation of ERK and its upstream kinases, MEK1/2, was detected in cells infected with SARS-CoV or overexpressed with SARS-CoV S protein [179,181]. However, ERK activation did not contribute to phosphorylation of its downstream target, p90 ribosomal S6 kinase (p90RSK) in SARS-CoV-infected Vero E6 cells [182]. This could be attributed to the higher phosphorylation levels of ERK1 compared to ERK2 in the SARS-CoV-infected cells, as ERK1 has been shown to suppress p90RSK phosphorylation and functional activity [183]. In another recent study, vimentin, a type III intermediate filament protein, was shown to be critical for entry of SARS-CoV, via direct interaction with the viral S protein [184]. Since vimentin could associate with β-adrenergic receptor to regulate ERK activation, one could speculate that induction of ERK pathway by SARS-CoV S protein might be through the interaction between vimentin and the ACE2 receptor, which is required for entry of SARS-CoV [185]. Additionally, binding of SARS-CoV S protein to ACE2 receptor stimulates upregulation of chemokine (C–C motif) ligand 2 (CCL2) mediated by ERK/AP-1 activation. CCL2 is believed to be responsible for respiratory inflammatory symptoms in SARS patients [181]. SARS-CoV S protein-induced ERK phosphorylation was shown to enhance IL8 release [186]. Other viral proteins of SARS-CoV have also been shown to induce ERK activation. Expression of SARS-CoV PLpro was shown to increase ERK1 ubiquitin-mediated degradation to suppress IFN-induced responses [187]. SARS-CoV 3b protein was also involved in ERK phosphorylation to potentiate AP-1 dependent activity of pro-inflammatory cytokine monocyte chemoattractant protein-1 (MCP-1) [188]. Besides SARS-CoV, MERS-CoV infection is also associated with enhanced ERK phosphorylation profile. The use of ER pathway inhibitor inhibited MERS-CoV infection by approximately 50% [189]. Although ERK phosphorylation was not significantly affected by HCoV-229E infection, the level of phosphorylated ERK can be enhanced by the use of chloroquine, a known antiviral agent against viruses [190]. Therefore, targeting the ERK pathway might have significant antiviral potential during HCoV infection.
6.1.2. JNK Pathway
Phosphorylation of JNK and its upstream kinases, MKK4/7, was detected in SARS-CoV-infected cells [191]. Overexpression of SARS-CoV 3a and 7a protein increased JNK activation and augmented IL8 promoter activity [192]. SARS-CoV 3b protein was shown to induce JNK/c-Jun/AP-1 activation to mediate transcription of MCP-1 [188]. Concurrently, the S protein of SARS-CoV was also shown to induce the activation of protein kinase epsilon via JNK activation [193]. Expression of SARS-CoV N protein is associated with the downregulation of prosurvival factors and apoptosis induction in COS-1 cells, possibly mediated by JNK activation [86]. Apoptosis induced by SARS-CoV 6 and 7a protein in Vero E6 and COS-7 cells were blocked by a JNK inhibitor [88]. Therefore, these findings suggest that JNK could act as a pro-apoptotic protein during SARS-CoV infection. However, in another study, phosphorylation of JNK was required for the maintenance of Vero E6 cells persistently infected with SARS-CoV. As persistent infection is only established after apoptotic events, it was proposed that JNK might act as a pro-apoptotic during acute phase of infection, but subsequently switched to become anti-apoptotic during prolonged infection [191]. It is uncertain if these observations noted were dependent on cell-type specificity. In our recent study, our group has found out that JNK was also activated during HCoV-229E infection and serves an anti-apoptotic role via modulation of Bcl2 family proteins. Furthermore, JNK contributes to the production of IFNβ and IL8 in HCoV-229E-infected cells (unpublished). The anti-apoptotic role of JNK during HCoV-229E infection contradicts our observations in H1299 cells infected with animal coronaviruses such as IBV, in which JNK has been shown to be pro-apoptotic [194]. The discrepancy of the involvement of JNK in CoV infections could be attributed to the different virus strains used in the experiments.
6.1.3. p38 MAPK Pathway
Activation of p38 MAPK has been reported in cells infected with SARS-CoV, MERS-CoV and HCoV-229E [180,189,190]. The upstream kinases of p38 MAPK, MKK3/6 were also shown to be phosphorylated [179]. On the other hand, downstream effectors of p38, MAPK-activated protein kinase-2 (MAPKAPK-2), cAMP response element-binding protein (CREB) and activation transcription factor-1 (ATF-1), were also activated in SARS-CoV-infected cells. SARS-CoV-induced phosphorylation of eukaryotic initiation factor 4E (eIF4a) was attenuated by p38 MAPK inhibitor. Although eIF4a is involved in protein translation, SARS-CoV viral protein translation and viral replication was not attenuated by p38 MAPK inhibitor, suggesting that SARS-CoV did not ultilise p38 for viral protein synthesis [180]. Furthermore, SARS-CoV infection induced p90RSK Ser380 phosphorylation, which could be partially negated by p38 MAPK inhibitor. Analysis of individual SARS-CoV proteins demonstrated that transfection of its 7a protein induced p38 activation when fused with GFP [195]. Since SARS-CoV 7a and 3a protein co-immunoprecipitated, they are likely to co-interact in virus-infected cells [196]. Therefore, it is possible that simultaneous co-expression of 7a and 3a proteins might further enhance p38 phosphorylation. In a recent study, the PBZ-binding motif (PBM) of SARS-CoV E protein was demonstrated to bind to syntenin and redistributes it to the cytoplasm, where it acts as a major scaffolding protein for p38 signalling cascade. Inhibition of syntenin using specific siRNA in cells transfected with functional E protein led to decreased p38 MAPK signalling [197]. Consistent with cell culture studies, phosphorylated p38 MAPK level was found to be increased in leukocytes of SARS patients. Enhanced p38, but not ERK, activation likely contributes to elevated IL8 levels and abnormal cytokine profile in SARS patients [198]. Additionally, p38 might also be involved in HCoV replication, as exemplified by HCoV-229E. Inhibition of p38 by its inhibitor SB203580 suppressed the HCoV-229E-induced cytopathic effects and significantly reduced viral titres in a dose dependent manner [190]. The use of chloroquine was also shown to exert antiviral effects against HCoV-229E infection, possibly via its attenuation on p38 activation [190].
8. Conclusions
The relationship between a virus and its host is a complicated affair: a myriad of factors from the virus and the host are involved in viral infection and consequential pathogenesis. During viral infections, the host must respond to the virus by putting multiple lines of defence mechanisms in place. As intracellular obligate parasites, viruses have also evolved various strategies to hijack the host machineries. In this review, we first showed how viral factors could manipulate the host cell to expedite its own replication cycle and pathogenesis. We also highlighted how multiple cellular and viral factors come into play in their long-standing battle against one another.
For years, HCoVs have been identified as mild respiratory pathogens that affect the human population. However, it was the emergence of SARS-CoV that thrust these human viruses into the spotlight of the research field. Therefore, most of the HCoV research today is pertained towards SARS-CoV. While the recent MERS-CoV outbreak has been mostly limited to the Middle East region, it is likely that more emerging or re-emerging HCoVs might surface to threaten the global public health, as seen from the high mortality rates in the past two outbreaks: SARS-CoV (10%) and MERS-CoV (35%). Therefore, study of the pathogenesis of all HCoVs would gain more insights for the development of antiviral therapeutics and vaccines.
Acknowledgments
This work was partially supported by a Competitive Research Programme (CRP) grant (NRF-CRP8-2011-05), the National Research Foundation, Singapore, an Academic Research Fund (AcRF) Tier 1 grant (RGT17/13), Nanyang Technological University and Ministry of Education, Singapore, and an AcRF Tier 2 grant (ACR47/14), Ministry of Education, Singapore.
Author Contributions
Yvonne Xinyi Lim and Yan Ling Ng wrote the paper; and James P. Tam and Ding Xiang Liu revised the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pene, F.; Merlat, A.; Vabret, A.; Rozenberg, F.; Buzyn, A.; Dreyfus, F.; Cariou, A.; Freymuth, F.; Lebon, P. Coronavirus 229E-Related Pneumonia in Immunocompromised Patients. Clin. Infect. Dis. 2003, 37, 929–932. [Google Scholar] [CrossRef] [PubMed]
- Vijgen, L.; Keyaerts, E.; Moës, E.; Maes, P.; Duson, G.; van Ranst, M. Development of One-Step, Real-Time, Quantitative Reverse Transcriptase PCR Assays for Absolute Quantitation of Human Coronaviruses OC43 and 229E. J. Clin. Microbiol. 2005, 43, 5452–5456. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis emergence linked to agricultural intensification and environmental change. Proc. Natl. Acad. Sci. USA 2013, 21, 8399–8340. [Google Scholar] [CrossRef] [PubMed]
- Van der Hoek, L. Human coronaviruses: What do they cause? Antivir. Ther. 2007, 12, 651–658. [Google Scholar] [PubMed]
- Walsh 2007, E.E.; Shin, J.H.; Falsey, A.R. Clinical Impact of Human Coronaviruses 229E and OC43 Infection in Diverse Adult Populations. J. Infect. Dis. 2013, 208, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Gorse, G.J.; O’Connor, T.Z.; Hall, S.L.; Vitale, J.N.; Nichol, K.L. Human Coronavirus and Acute Respiratory Illness in Older Adults with Chronic Obstructive Pulmonary Disease. J. Infect. Dis. 2009, 199, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Arbour, N.; Day, R.; Newcombe, J.; Talbot, P.J. Neuroinvasion by Human Respiratory Coronaviruses. J. Virol. 2000, 74, 8913–8921. [Google Scholar] [CrossRef] [PubMed]
- Arbour, N.; Ekandé, S.; Côté, G.; Lachance, C.; Chagnon, F.; Tardieu, M.; Cashman, N.R.; Talbot, P.J. Persistent Infection of Human Oligodendrocytic and Neuroglial Cell Lines by Human Coronavirus 229E. J. Virol. 1999, 73, 3326–3337. [Google Scholar] [PubMed]
- Jacomy, H.; Fragoso, G.; Almazan, G.; Mushynski, W.E.; Talbot, P.J. Human coronavirus OC43 infection induces chronic encephalitis leading to disabilities in BALB/C mice. Virology 2006, 349, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Vabret, A.; Mourez, T.; Gouarin, S.; Petitjean, J.; Freymuth, F. An Outbreak of Coronavirus OC43 Respiratory Infection in Normandy, France. Clin. Infect. Dis. 2003, 36, 985–989. [Google Scholar] [CrossRef] [PubMed]
- Smuts, H. Human coronavirus NL63 infections in infants hospitalised with acute respiratory tract infections in South Africa. Influenza Other Respir. Viruses 2008, 2, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.L.; Donaldson, E.F.; Baric, R.S. A decade after SARS: Strategies for controlling emerging coronaviruses. Nat. Rev. Microbiol. 2013, 11, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Frieman, M.; Baric, R. Mechanisms of Severe Acute Respiratory Syndrome Pathogenesis and Innate Immunomodulation. Microbiol. Mol. Biol. Rev. MMBR 2008, 72, 672–685. [Google Scholar] [CrossRef] [PubMed]
- Peiris, J.S.M.; Guan, Y.; Yuen, K.Y. Severe acute respiratory syndrome. Nat. Med. 2004, 10, S88–S97. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yan, M.; Xu, H.; Liang, W.; Kan, B.; Zheng, B.; Chen, H.; Zheng, H.; Xu, Y.; Zhang, E.; et al. SARS-CoV Infection in a Restaurant from Palm Civet. Emerg. Infect. Dis. 2005, 11, 1860–1865. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Ge, X.; Wang, L.-F.; Shi, Z. Bat origin of human coronaviruses. Virol. J. 2015, 12, 221. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Cheon, S.; Min, C.-K.; Sohn, K.M.; Kang, Y.J.; Cha, Y.-J.; Kang, J.I.; Han, S.K.; Ha, N.Y.; Kim, G.; et al. Spread of Mutant Middle East Respiratory Syndrome Coronavirus with Reduced Affinity to Human CD26 during the South Korean Outbreak. mBio 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Oboho, I.K.; Tomczyk, S.M.; Al-Asmari, A.M.; Banjar, A.A.; Al-Mugti, H.; Aloraini, M.S.; Alkhaldi, K.Z.; Almohammadi, E.L.; Alraddadi, B.M.; Gerber, S.I.; et al. 2014 MERS-CoV Outbreak in Jeddah—A Link to Health Care Facilities. N. Engl. J. Med. 2015, 372, 846–854. [Google Scholar] [CrossRef] [PubMed]
- The Korean Society of Infectious Diseases; Korean Society for Healthcare-associated Infection Control and Prevention. An Unexpected Outbreak of Middle East Respiratory Syndrome Coronavirus Infection in the Republic of Korea, 2015. Infect. Chemother. 2015, 47, 120–122. [Google Scholar]
- Masters, P.S. The Molecular Biology of Coronaviruses Advances in Virus Research; Academic Press: Massachusetts, MA, USA, 2006; Volume 66, pp. 193–292. [Google Scholar]
- McBride, R.; Fielding, B.C. The Role of Severe Acute Respiratory Syndrome (SARS)-Coronavirus Accessory Proteins in Virus Pathogenesis. Viruses 2012, 4, 2902–2923. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Snijder, E.J.; Spaan, W.J.M. Severe Acute Respiratory Syndrome Coronavirus Phylogeny: Toward Consensus. J. Virol. 2004, 78, 7863–7866. [Google Scholar] [CrossRef] [PubMed]
- Kolesnikova, L.; Slenczka, W.; Brodt, H.; Klenk, H.; Becker, S. Electron microscopy in diagnostics of SARS case. Microsc. Microanal. 2003, 9, 438–439. [Google Scholar]
- Marsolais, G.; Berthiaume, L.; DiFranco, E.; Marois, P. Rapid Diagnosis by Electron Microscopy of Avian Coronavirus Infection. Can. J. Comp. Med. 1971, 35, 285–288. [Google Scholar] [PubMed]
- Liu, D.X.; Fung, T.S.; Chong, K.K.-L.; Shukla, A.; Hilgenfeld, R. Accessory proteins of SARS-CoV and other coronaviruses. Antivir. Res. 2014, 109, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Yeager, C.L.; Ashmun, R.A.; Williams, R.K.; Cardellichio, C.B.; Shapiro, L.H.; Look, A.T.; Holmes, K.V. Human aminopeptidase N is a receptor for human coronavirus 229E. Nature 1992, 357, 420–422. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Sui, J.; Huang, I.C.; Kuhn, J.H.; Radoshitzky, S.R.; Marasco, W.A.; Choe, H.; Farzan, M. The S proteins of human coronavirus NL63 and severe acute respiratory syndrome coronavirus bind overlapping regions of ACE2. Virology 2007, 367, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Li, W.; Peng, G.; Li, F. Crystal structure of NL63 respiratory coronavirus receptor-binding domain complexed with its human receptor. Proc. Natl. Acad. Sci. USA 2009, 106, 19970–19974. [Google Scholar] [CrossRef] [PubMed]
- Van Doremalen, N.; Miazgowicz, K.L.; Milne-Price, S.; Bushmaker, T.; Robertson, S.; Scott, D.; Kinne, J.; McLellan, J.S.; Zhu, J.; Munster, V.J. Host Species Restriction of Middle East Respiratory Syndrome Coronavirus through Its Receptor, Dipeptidyl Peptidase 4. J. Virol. 2014, 88, 9220–9232. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Dong, W.; Milewska, A.; Golda, A.; Qi, Y.; Zhu, Q.K.; Marasco, W.A.; Baric, R.S.; Sims, A.C.; Pyrc, K.; et al. Human Coronavirus HKU1 Spike Protein Uses O-Acetylated Sialic Acid as an Attachment Receptor Determinant and Employs Hemagglutinin-Esterase Protein as a Receptor-Destroying Enzyme. J. Virol. 2015, 89, 7202–7213. [Google Scholar] [CrossRef] [PubMed]
- Butler, N.; Pewe, L.; Trandem, K.; Perlman, S. Murine encephalitis caused by HCoV-OC43, a human coronavirus with broad species specificity, is partly immune-mediated. Virology 2006, 347, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.; Chan, J.W.; Azhar, E.I.; Hui, D.C.; Yuen, K. Coronaviruses—Drug discovery and therapeutic options. Nat. Rev. Drug Discov. 2016, 15, 327–347. [Google Scholar] [CrossRef] [PubMed]
- Bosch, B.J.; Bartelink, W.; Rottier, P.J.M. Cathepsin L Functionally Cleaves the Severe Acute Respiratory Syndrome Coronavirus Class I Fusion Protein Upstream of Rather than Adjacent to the Fusion Peptide. J. Virol. 2008, 82, 8887–8890. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.; Dominguez, S.R.; Holmes, K.V. Role of the Spike Glycoprotein of Human Middle East Respiratory Syndrome Coronavirus (MERS-CoV) in Virus Entry and Syncytia Formation. PLoS ONE 2013, 8, e76469. [Google Scholar] [CrossRef] [PubMed]
- Simmons, G.; Gosalia, D.N.; Rennekamp, A.J.; Reeves, J.D.; Diamond, S.L.; Bates, P. Inhibitors of cathepsin L prevent severe acute respiratory syndrome coronavirus entry. Proc. Natl. Acad. Sci. USA 2005, 102, 11876–11881. [Google Scholar] [CrossRef] [PubMed]
- Bertram, S.; Dijkman, R.; Habjan, M.; Heurich, A.; Gierer, S.; Glowacka, I.; Welsch, K.; Winkler, M.; Schneider, H.; Hofmann-Winkler, H.; et al. TMPRSS2 Activates the Human Coronavirus 229E for Cathepsin-Independent Host Cell Entry and Is Expressed in Viral Target Cells in the Respiratory Epithelium. J. Virol. 2013, 87, 6150–6160. [Google Scholar] [CrossRef] [PubMed]
- Bertram, S.; Glowacka, I.; Müller, M.A.; Lavender, H.; Gnirss, K.; Nehlmeier, I.; Niemeyer, D.; He, Y.; Simmons, G.; Drosten, C.; et al. Cleavage and Activation of the Severe Acute Respiratory Syndrome Coronavirus Spike Protein by Human Airway Trypsin-Like Protease. J. Virol. 2011, 85, 13363–13372. [Google Scholar] [CrossRef] [PubMed]
- Millet, J.K.; Whittaker, G.R. Host cell entry of Middle East respiratory syndrome coronavirus after two-step, furin-mediated activation of the spike protein. Proc. Natl. Acad. Sci. USA 2014, 111, 15214–15219. [Google Scholar] [CrossRef] [PubMed]
- Huang, I.C.; Bailey, C.C.; Weyer, J.L.; Radoshitzky, S.R.; Becker, M.M.; Chiang, J.J.; Brass, A.L.; Ahmed, A.A.; Chi, X.; Dong, L.; et al. Distinct Patterns of IFITM-Mediated Restriction of Filoviruses, SARS Coronavirus, and Influenza A Virus. PLoS Pathog. 2011, 7, e1001258. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Markosyan, R.M.; Zheng, Y.-M.; Golfetto, O.; Bungart, B.; Li, M.; Ding, S.; He, Y.; Liang, C.; Lee, J.C.; et al. IFITM Proteins Restrict Viral Membrane Hemifusion. PLoS Pathog. 2013, 9, e1003124. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Chen, Q.; Chen, J.; Chen, K.; Shen, X.; Jiang, H. The nucleocapsid protein of SARS coronavirus has a high binding affinity to the human cellular heterogeneous nuclear ribonucleoprotein A1. FEBS Lett. 2005, 579, 2623–2628. [Google Scholar] [CrossRef] [PubMed]
- Nanda, S.K.; Leibowitz, J.L. Mitochondrial Aconitase Binds to the 3′ Untranslated Region of the Mouse Hepatitis Virus Genome. J. Virol. 2001, 75, 3352–3362. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.-H.; Chen, P.-J.; Yeh, S.-H. Nucleocapsid Phosphorylation and RNA Helicase DDX1 Recruitment Enables Coronavirus Transition from Discontinuous to Continuous Transcription. Cell Host Microb. 2014, 16, 462–472. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.W.; Hong, W.; Liu, D.X. Binding of the 5′-untranslated region of coronavirus RNA to zinc finger CCHC-type and RNA-binding motif 1 enhances viral replication and transcription. Nucleic Acids Res. 2012, 40, 5065–5077. [Google Scholar] [CrossRef] [PubMed]
- Neuman, B.W.; Kiss, G.; Kunding, A.H.; Bhella, D.; Baksh, M.F.; Connelly, S.; Droese, B.; Klaus, J.P.; Makino, S.; Sawicki, S.G.; et al. A structural analysis of M protein in coronavirus assembly and morphology. J. Struct. Biol. 2011, 174, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Wu, D.; Shen, C.; Chen, K.; Shen, X.; Jiang, H. Severe acute respiratory syndrome coronavirus membrane protein interacts with nucleocapsid protein mostly through their carboxyl termini by electrostatic attraction. Int. J. Biochem. Cell Biol. 2006, 38, 589–599. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.H.; Kumar, P.; Tay, F.P.L.; Moreau, D.; Liu, D.X.; Bard, F. Genome-Wide Screen Reveals Valosin-Containing Protein Requirement for Coronavirus Exit from Endosomes. J. Virol. 2015, 89, 11116–11128. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.R.; Wyllie, A.H.; Currie, A.R. Apoptosis: A Basic Biological Phenomenon with Wide-ranging Implications in Tissue Kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of Poly(ADP-ribose) Polymerase (PARP) Cleavage in Apoptosis: Caspase 3-resistant parp mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar] [CrossRef] [PubMed]
- Segawa, K.; Kurata, S.; Yanagihashi, Y.; Brummelkamp, T.R.; Matsuda, F.; Nagata, S. Caspase-mediated cleavage of phospholipid flippase for apoptotic phosphatidylserine exposure. Science 2014, 344, 1164–1168. [Google Scholar] [CrossRef] [PubMed]
- Walczak, H.; Krammer, P.H. The CD95 (APO-1/Fas) and the TRAIL (APO-2L) Apoptosis Systems. Exp. Cell Res. 2000, 256, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Bender, L.M.; Morgan, M.J.; Thomas, L.R.; Liu, Z.G.; Thorburn, A. The adaptor protein TRADD activates distinct mechanisms of apoptosis from the nucleus and the cytoplasm. Cell Death Differ. 2005, 12, 473–481. [Google Scholar] [CrossRef] [PubMed]
- Stennicke, H.R.; Jürgensmeier, J.M.; Shin, H.; Deveraux, Q.; Wolf, B.B.; Yang, X.; Zhou, Q.; Ellerby, H.M.; Ellerby, L.M.; Bredesen, D.; et al. Pro-caspase-3 Is a Major Physiologic Target of Caspase-8. J. Biol. Chem. 1998, 273, 27084–27090. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Sebbagh, M.; Renvoize, C.; Hamelin, J.; Riche, N.; Bertoglio, J.; Breard, J. Caspase-3-mediated cleavage of ROCK I induces MLC phosphorylation and apoptotic membrane blebbing. Nat. Cell Biol. 2001, 3, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhu, H.; Xu, C.-J.; Yuan, J. Cleavage of BID by Caspase 8 Mediates the Mitochondrial Damage in the Fas Pathway of Apoptosis. Cell 1998, 94, 491–501. [Google Scholar] [CrossRef]
- Benedict, C.A.; Norris, P.S.; Ware, C.F. To kill or be killed: Viral evasion of apoptosis. Nat. Immunol. 2002, 3, 1013–1018. [Google Scholar] [CrossRef] [PubMed]
- Kvansakul, M.; Hinds, M.G. Structural biology of the Bcl-2 family and its mimicry by viral proteins. Cell Death Dis. 2013, 4, e909. [Google Scholar] [CrossRef] [PubMed]
- Pearce, A.F.; Lyles, D.S. Vesicular Stomatitis Virus Induces Apoptosis Primarily through Bak Rather than Bax by Inactivating Mcl-1 and Bcl-XL. J. Virol. 2009, 83, 9102–9112. [Google Scholar] [CrossRef] [PubMed]
- Aillet, F.; Masutani, H.; Elbim, C.; Raoul, H.; Chêne, L.; Nugeyre, M.T.; Paya, C.; Barré-Sinoussi, F.; Gougerot-Pocidalo, M.A.; Israël, N. Human immunodeficiency virus induces a dual regulation of Bcl-2, resulting in persistent infection of CD4(+) T- or monocytic cell lines. J. Virol. 1998, 72, 9698–9705. [Google Scholar] [PubMed]
- Tamura, R.; Kanda, T.; Imazeki, F.; Wu, S.; Nakamoto, S.; Tanaka, T.; Arai, M.; Fujiwara, K.; Saito, K.; Roger, T.; et al. Hepatitis C Virus Nonstructural 5A Protein Inhibits Lipopolysaccharide-Mediated Apoptosis of Hepatocytes by Decreasing Expression of Toll-Like Receptor 4. J. Infect. Dis. 2011, 204, 793–801. [Google Scholar] [CrossRef] [PubMed]
- Aweya, J.J.; Sze, C.W.; Bayega, A.; Mohd-Ismail, N.K.; Deng, L.; Hotta, H.; Tan, Y.J. NS5B induces up-regulation of the BH3-only protein, BIK, essential for the hepatitis C virus RNA replication and viral release. Virology 2015, 474, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Nakamura-Lopez, Y.; Villegas-Sepúlveda, N.; Gómez, B. RSV P-protein impairs extrinsic apoptosis pathway in a macrophage-like cell line persistently infected with respiratory syncytial virus. Virus Res. 2015, 204, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Mocarski, E.S.; Upton, J.W.; Kaiser, W.J. Viral infection and the evolution of caspase 8-regulated apoptotic and necrotic death pathways. Nat. Rev. Immunol. 2012, 12, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Amara, A.; Mercer, J. Viral apoptotic mimicry. Nat. Rev. Microbiol. 2015, 13, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. Induction of Apoptosis in MRC-5, Diploid Human Fetal Lung Cells after Infection with Human Coronavirus OC43. In The Nidoviruses: Coronaviruses and Arteriviruses; Lavi, E., Weiss, S.R., Hingley, S.T., Eds.; Springer US: Boston, MA, USA, 2001; pp. 677–682. [Google Scholar]
- Pyrc, K.; Berkhout, B.; van der Hoek, L. The Novel Human Coronaviruses NL63 and HKU1. J. Virol. 2007, 81, 3051–3057. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Korteweg, C. Pathology and Pathogenesis of Severe Acute Respiratory Syndrome. Am. J. Pathol. 2007, 170, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Hill, T.E.; Morimoto, C.; Peters, C.J.; Ksiazek, T.G.; Tseng, C.-T.K. Bilateral Entry and Release of Middle East Respiratory Syndrome Coronavirus Induces Profound Apoptosis of Human Bronchial Epithelial Cells. J. Virol. 2013, 87, 9953–9958. [Google Scholar] [CrossRef] [PubMed]
- Yeung, M.-L.; Yao, Y.; Jia, L.; Chan, J.F.W.; Chan, K.-H.; Cheung, K.-F.; Chen, H.; Poon, V.K.M.; Tsang, A.K.L.; To, K.K.W.; et al. MERS coronavirus induces apoptosis in kidney and lung by upregulating Smad7 and FGF2. Nat. Microbiol. 2016, 1, 16004. [Google Scholar] [CrossRef]
- Desforges, M.; Coupanec, A.; Brison, É.; Meessen-Pinard, M.; Talbot, P.J. Neuroinvasive and Neurotropic Human Respiratory Coronaviruses: Potential Neurovirulent Agents in Humans. In Proceedings of the Infectious Diseases and Nanomedicine I: First International Conference (ICIDN-2012), Kathmandu, Nepal, 15–18 December 2012; Adhikari, R., Thapa, S., Eds.; Springer: New Delhi, India, 2014; pp. 75–96. [Google Scholar]
- Desforges, M.; Le Coupanec, A.; Stodola, J.K.; Meessen-Pinard, M.; Talbot, P.J. Human coronaviruses: Viral and cellular factors involved in neuroinvasiveness and neuropathogenesis. Virus Res. 2014, 194, 145–158. [Google Scholar] [CrossRef] [PubMed]
- Favreau, D.J.; Meessen-Pinard, M.; Desforges, M.; Talbot, P.J. Human Coronavirus-Induced Neuronal Programmed Cell Death Is Cyclophilin D Dependent and Potentially Caspase Dispensable. J. Virol. 2012, 86, 81–93. [Google Scholar] [CrossRef] [PubMed]
- Krähling, V.; Stein, D.A.; Spiegel, M.; Weber, F.; Mühlberger, E. Severe Acute Respiratory Syndrome Coronavirus Triggers Apoptosis via Protein Kinase R but Is Resistant to Its Antiviral Activity. J. Virol. 2009, 83, 2298–2309. [Google Scholar] [CrossRef] [PubMed]
- Collins, A.R. In Vitro Detection of Apoptosis in Monocytes/Macrophages Infected with Human Coronavirus. Clin. Diagn. Lab. Immunol. 2002, 9, 1392–1395. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xiong, Z.; Zhang, S.; Yan, Y.; Nguyen, J.; Ng, B.; Lu, H.; Brendese, J.; Yang, F.; Wang, H.; et al. Bcl-xL inhibits T-cell apoptosis induced by expression of SARS coronavirus E protein in the absence of growth factors. Biochem. J. 2005, 392, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Ying, T.; Li, W.; Dimitrov, D.S. Discovery of T-Cell Infection and Apoptosis by Middle East Respiratory Syndrome Coronavirus. J. Infect. Dis. 2015. [Google Scholar] [CrossRef] [PubMed]
- Chu, H.; Zhou, J.; Wong, B.H.-Y.; Li, C.; Chan, J.F.-W.; Cheng, Z.-S.; Yang, D.; Wang, D.; Lee, A.C.; Li, C.; et al. Middle East Respiratory Syndrome Coronavirus Efficiently Infects Human Primary T Lymphocytes and Activates the Extrinsic and Intrinsic Apoptosis Pathways. J. Infect. Dis. 2016, 213, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Mesel-Lemoine, M.; Millet, J.; Vidalain, P.-O.; Law, H.; Vabret, A.; Lorin, V.; Escriou, N.; Albert, M.L.; Nal, B.; Tangy, F. A Human Coronavirus Responsible for the Common Cold Massively Kills Dendritic Cells but Not Monocytes. J. Virol. 2012, 86, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, M.; Schneider, K.; Weber, F.; Weidmann, M.; Hufert, F. Interaction of severe acute respiratory syndrome-associated coronavirus with dendritic cells. J. Gen. Virol. 2006, 87, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Bordi, L.; Castilletti, C.; Falasca, L.; Ciccosanti, F.; Calcaterra, S.; Rozera, G.; Di Caro, A.; Zaniratti, S.; Rinaldi, A.; Ippolito, G.; et al. Bcl-2 inhibits the caspase-dependent apoptosis induced by SARS-CoV without affecting virus replication kinetics. Arch. Virol. 2005, 151, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Yang, R.; Guo, L.; Qu, J.; Wang, J.; Hung, T. Apoptosis Induced by the SARS-Associated Coronavirus in Vero Cells Is Replication-Dependent and Involves Caspase. DNA Cell Biol. 2005, 24, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.S.F.; Chan, K.-h.; Cheng, V.C.C.; Woo, P.C.Y.; Lau, S.K.P.; Lam, C.C.K.; Chan, T.L.; Wu, A.K.; Hung, I.F.; Leung, S.Y.; et al. Comparative Host Gene Transcription by Microarray Analysis Early after Infection of the Huh7 Cell Line by Severe Acute Respiratory Syndrome Coronavirus and Human Coronavirus 229E. J. Virol. 2005, 79, 6180–6193. [Google Scholar] [CrossRef] [PubMed]
- Chow, K.Y.C.; Yeung, Y.S.; Hon, C.C.; Zeng, F.; Law, K.M.; Leung, F.C.C. Adenovirus-mediated expression of the C-terminal domain of SARS-CoV spike protein is sufficient to induce apoptosis in Vero E6 cells. FEBS Lett. 2005, 579, 6699–6704. [Google Scholar] [CrossRef] [PubMed]
- Surjit, M.; Liu, B.; Jameel, S.; Chow, V.T.K.; Lal, S.K. The SARS coronavirus nucleocapsid protein induces actin reorganization and apoptosis in COS-1 cells in the absence of growth factors. Biochem. J. 2004, 383, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.-J.; Fielding, B.C.; Goh, P.-Y.; Shen, S.; Tan, T.H.P.; Lim, S.G.; Hong, W. Overexpression of 7a, a Protein Specifically Encoded by the Severe Acute Respiratory Syndrome Coronavirus, Induces Apoptosis via a Caspase-Dependent Pathway. J. Virol. 2004, 78, 14043–14047. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Wong, C.K.; Li, P.; Xie, Y. A SARS-CoV protein, ORF-6, induces caspase-3 mediated, ER stress and JNK-dependent apoptosis. Biochim. Biophys. Acta 2008, 1780, 1383–1387. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Åkerström, S.; Sharma, A.K.; Chow, V.T.K.; Teow, S.; Abrenica, B.; Booth, S.A.; Booth, T.F.; Mirazimi, A.; Lal, S.K. SARS-CoV 9b Protein Diffuses into Nucleus, Undergoes Active Crm1 Mediated Nucleocytoplasmic Export and Triggers Apoptosis When Retained in the Nucleus. PLoS ONE 2011, 6, e19436. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, H.; Li, L.; Chen, Zhefan S.; Lau, K.-F.; Tsui, S.K.W.; Chan, H.Y.E. The SARS-coronavirus membrane protein induces apoptosis via interfering with PDK1-PKB/Akt signalling. Biochem. J. 2014, 464, 439–447. [Google Scholar] [CrossRef] [PubMed]
- De Diego, M.L.; Nieto-Torres, J.L.; JimÈnez-GuardeÒo, J.M.; Regla-Nava, J.A.; Álvarez, E.; Oliveros, J.C.; Zhao, J.; Fett, C.; Perlman, S.; Enjuanes, L. Severe Acute Respiratory Syndrome Coronavirus Envelope Protein Regulates Cell Stress Response and Apoptosis. PLoS Pathog. 2011, 7, e1002315. [Google Scholar]
- Tan, Y.-X.; Tan, T.H.P.; Lee, M.J.R.; Tham, P.-Y.; Gunalan, V.; Druce, J.; Birch, C.; Catton, M.; Fu, N.Y.; Yu, V.C.; et al. Induction of Apoptosis by the Severe Acute Respiratory Syndrome Coronavirus 7a Protein Is Dependent on Its Interaction with the Bcl-X(L) Protein. J. Virol. 2007, 81, 6346–6355. [Google Scholar] [CrossRef] [PubMed]
- Favreau, D.J.; Desforges, M.; St-Jean, J.R.; Talbot, P.J. A human coronavirus OC43 variant harboring persistence-associated mutations in the S glycoprotein differentially induces the unfolded protein response in human neurons as compared to wild-type virus. Virology 2009, 395, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Diemer, C.; Schneider, M.; Seebach, J.; Quaas, J.; Frösner, G.; Schätzl, H.M.; Gilch, S. Cell Type-Specific Cleavage of Nucleocapsid Protein by Effector Caspases during SARS Coronavirus Infection. J. Mol. Biol. 2008, 376, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Alcami, A.; Koszinowski, U.H. Viral mechanisms of immune evasion. Immunol. Today 2000, 21, 447–455. [Google Scholar] [CrossRef]
- Bowie, A.G.; Unterholzner, L. Viral evasion and subversion of pattern-recognition receptor signalling. Nat. Rev. Immunol. 2008, 8, 911–922. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. Toll-like Receptors and Their Crosstalk with Other Innate Receptors in Infection and Immunity. Immunity 2011, 34, 637–650. [Google Scholar] [CrossRef] [PubMed]
- Jensen, S.; Thomsen, A.R. Sensing of RNA Viruses: A Review of Innate Immune Receptors Involved in Recognizing RNA virus Invasion. J. Virol. 2012, 86, 2900–2910. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.Y.; Kok, K.-H.; Jaume, M.; Cheung, T.K.W.; Yip, T.-F.; Lai, J.C.C.; Guan, Y.; Webster, R.G.; Jin, D.Y.; Peiris, J.S. Toll-like receptor 10 is involved in induction of innate immune responses to influenza virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 3793–3798. [Google Scholar] [CrossRef] [PubMed]
- Weber, F.; Wagner, V.; Rasmussen, S.B.; Hartmann, R.; Paludan, S.R. Double-Stranded RNA Is Produced by Positive-Strand RNA Viruses and DNA Viruses but Not in Detectable Amounts by Negative-Strand RNA Viruses. J. Virol. 2006, 80, 5059–5064. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Takeuchi, O.; Kawai, T.; Kaisho, T.; Sato, S.; Sanjo, H.; Matsumoto, M.; Hoshino, K.; Wagner, H.; Takeda, K.; et al. A Toll-like receptor recognizes bacterial DNA. Nature 2000, 408, 740–745. [Google Scholar] [PubMed]
- Krug, A.; Luker, G.D.; Barchet, W.; Leib, D.A.; Akira, S.; Colonna, M. Herpes simplex virus type 1 activates murine natural interferon-producing cells through toll-like receptor 9. Blood 2003, 103, 1433–1437. [Google Scholar] [CrossRef] [PubMed]
- Krug, A.; French, A.R.; Barchet, W.; Fischer, J.A.A.; Dzionek, A.; Pingel, J.T.; Orihuela, M.M.; Akira, S.; Yokoyama, W.M.; Colonna, M. TLR9-Dependent Recognition of MCMV by IPC and DC Generates Coordinated Cytokine Responses that Activate Antiviral NK Cell Function. Immunity 2004, 21, 107–119. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Zeisel, M.B.; Jilg, N.; Paranhos-Baccalà, G.; Stoll-Keller, F.; Wakita, T.; Hafkemeyer, P.; Blum, H.E.; Barth, H.; Henneke, P.; et al. Toll-like receptor 2 senses hepatitis C virus core protein but not infectious viral particles. J. Innate Immunity 2009, 1, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Bieback, K.; Lien, E.; Klagge, I.M.; Avota, E.; Schneider-Schaulies, J.; Duprex, W.P.; Wagner, H.; Kirschning, C.J.; Ter Meulen, V.; Schneider-Schaulies, S. Hemagglutinin Protein of Wild-Type Measles Virus Activates Toll-Like Receptor 2 Signaling. J. Virol. 2002, 76, 8729–8736. [Google Scholar] [CrossRef] [PubMed]
- Rallabhandi, P.; Phillips, R.L.; Boukhvalova, M.S.; Pletneva, L.M.; Shirey, K.A.; Gioannini, T.L.; Weiss, J.P.; Chow, J.C.; Hawkins, L.D.; Vogel, S.N.; et al. Respiratory Syncytial Virus Fusion Protein-Induced Toll-Like Receptor 4 (TLR4) Signaling Is Inhibited by the TLR4 Antagonists Rhodobacter sphaeroides Lipopolysaccharide and Eritoran (E5564) and Requires Direct Interaction with MD-2. mBio 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Ben Haij, N.; Leghmari, K.; Planès, R.; Thieblemont, N.; Bahraoui, E. HIV-1 Tat protein binds to TLR4-MD2 and signals to induce TNF-α and IL-10. Retrovirology 2013, 10, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Kawai, T.; Akira, S. Toll-like receptors and innate immunity. Biochem. Biophys. Res. Commun. 2009, 388, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Taniguchi, T. IRFs: Master regulators of signalling by Toll-like receptors and cytosolic pattern-recognition receptors. Nat. Rev. Immunol. 2006, 6, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Wang, H.; Hajishengallis, G.N.; Martin, M. TLR-signaling Networks: An Integration of Adaptor Molecules, Kinases, and Cross-talk. J. Dent. Res. 2011, 90, 417–427. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors [mdash] redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Yoneyama, M.; Kikuchi, M.; Matsumoto, K.; Imaizumi, T.; Miyagishi, M.; Taira, K.; Foy, E.; Loo, Y.M.; Gale, M., Jr.; Akira, S.; et al. Shared and Unique Functions of the DExD/H-Box Helicases RIG-I, MDA5, and LGP2 in Antiviral Innate Immunity. J. Immunol. 2005, 175, 2851–2858. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, K.R.; Bruns, A.M.; Horvath, C.M. MDA5 and LGP2: Accomplices and Antagonists of Antiviral Signal Transduction. J. Virol. 2014, 88, 8194–8200. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Kato, H.; Kumagai, Y.; Yoneyama, M.; Sato, S.; Matsushita, K.; Tsujimura, T.; Fujita, T.; Akira, S.; Takeuchi, O. LGP2 is a positive regulator of RIG-I- and MDA5-mediated antiviral responses. Proc. Natl. Acad. Sci. USA 2010, 107, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, X.; Wang, G.; Zheng, H. The Laboratory of Genetics and Physiology 2: Emerging Insights into the Controversial Functions of This RIG-I-Like Receptor. BioMed Res. Int. 2014, 2014, 7. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Park, H.-S.; Pyo, H.-M.; Liu, Q.; Zhou, Y. Influenza A Virus Panhandle Structure Is Directly Involved in RIG-I Activation and Interferon Induction. J. Virol. 2015, 89, 6067–6079. [Google Scholar] [CrossRef] [PubMed]
- Gack, M.U. Mechanisms of RIG-I-like Receptor Activation and Manipulation by Viral Pathogens. J. Virol. 2014. [Google Scholar] [CrossRef] [PubMed]
- Kanneganti, T.-D. Central roles of NLRs and inflammasomes in viral infection. Nat. Rev. Immunol. 2010, 10, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Callaway, J.B.; Ting, J.P.Y. Inflammasomes: Mechanism of action, role in disease, and therapeutics. Nat. Med. 2015, 21, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B.; Donlin, L.T. Regulation of type I interferon responses. Nat. Rev. Immunol. 2014, 14, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Kindler, E.; Jónsdóttir, H.R.; Muth, D.; Hamming, O.J.; Hartmann, R.; Rodriguez, R.; Geffers, R.; Fouchier, R.A.; Drosten, C.; Müller, M.A.; et al. Efficient Replication of the Novel Human Betacoronavirus EMC on Primary Human Epithelium Highlights Its Zoonotic Potential. mBio 2013, 4, e00611–e00612. [Google Scholar] [CrossRef] [PubMed]
- Frieman, M.; Heise, M.; Baric, R. SARS coronavirus and innate immunity. Virus Res. 2008, 133, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Clementz, M.A.; Chen, Z.; Banach, B.S.; Wang, Y.; Sun, L.; Ratia, K.; Baez-Santos, Y.M.; Wang, J.; Takayama, J.; Ghosh, A.K.; et al. Deubiquitinating and Interferon Antagonism Activities of Coronavirus Papain-Like Proteases. J. Virol. 2010, 84, 4619–4629. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Lau, C.C.Y.; Chan, K.-H.; Li, C.P.Y.; Chen, H.; Jin, D.-Y.; Chan, J.F.; Woo, P.C.; Yuen, K.Y. Delayed induction of proinflammatory cytokines and suppression of innate antiviral response by the novel Middle East respiratory syndrome coronavirus: Implications for pathogenesis and treatment. J. Gen. Virol. 2013, 94, 2679–2690. [Google Scholar] [CrossRef] [PubMed]
- Cheung, C.Y.; Poon, L.L.M.; Ng, I.H.Y.; Luk, W.; Sia, S.-F.; Wu, M.H.S.; Chan, K.H.; Yuen, K.Y.; Gordon, S.; Guan, Y.; et al. Cytokine Responses in Severe Acute Respiratory Syndrome Coronavirus-Infected Macrophages In Vitro: Possible Relevance to Pathogenesis. J. Virol. 2005, 79, 7819–7826. [Google Scholar] [CrossRef] [PubMed]
- Randall, R.E.; Goodbourn, S. Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 2008, 89, 1–47. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.J.; Wang, J.; Ito, Y.; Travanty, E.A.; Voelker, D.R.; Holmes, K.V.; Mason, R.J. Infection of human alveolar macrophages by human coronavirus strain 229E. J. Gen. Virol. 2012, 93, 494–503. [Google Scholar] [CrossRef] [PubMed]
- Gosert, R.; Kanjanahaluethai, A.; Egger, D.; Bienz, K.; Baker, S.C. RNA Replication of Mouse Hepatitis Virus Takes Place at Double-Membrane Vesicles. J. Virol. 2002, 76, 3697–3708. [Google Scholar] [CrossRef] [PubMed]
- Perlman, S.; Netland, J. Coronaviruses post-SARS: Update on replication and pathogenesis. Nature reviews. Microbiology 2009, 7, 439–450. [Google Scholar] [PubMed]
- Siu, K.L.; Kok, K.H.; Ng, M.H.; Poon, V.K.; Yuen, K.Y.; Zheng, B.J.; Jin, D.Y. Severe Acute Respiratory Syndrome Coronavirus M Protein Inhibits Type I Interferon Production by Impeding the Formation of TRAF3·TANK·TBK1/IKKε Complex. J. Biol. Chem. 2009, 284, 16202–16209. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.-L.; Chan, C.-P.; Kok, K.-H.; Chiu-Yat Woo, P.; Jin, D.-Y. Suppression of innate antiviral response by severe acute respiratory syndrome coronavirus M protein is mediated through the first transmembrane domain. Cell. Mol. Immunol. 2014, 11, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, L.; Geng, H.; Deng, Y.; Huang, B.; Guo, Y.; Zhao, Z.; Tan, W. The structural and accessory proteins M, ORF 4a, ORF 4b, and ORF 5 of Middle East respiratory syndrome coronavirus (MERS-CoV) are potent interferon antagonists. Protein Cell 2013, 4, 951–961. [Google Scholar] [CrossRef] [PubMed]
- Kopecky-Bromberg, S.A.; Martínez-Sobrido, L.; Frieman, M.; Baric, R.A.; Palese, P. Severe Acute Respiratory Syndrome Coronavirus Open Reading Frame (ORF) 3b, ORF 6, and Nucleocapsid Proteins Function as Interferon Antagonists. J. Virol. 2007, 81, 548–557. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Pan, J.A.; Tao, J.; Guo, D. SARS-CoV nucleocapsid protein antagonizes IFN-β response by targeting initial step of IFN-β induction pathway, and its C-terminal region is critical for the antagonism. Virus Genes 2010, 42, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Lokugamage, K.G.; Rozovics, J.M.; Narayanan, K.; Semler, B.L.; Makino, S. SARS Coronavirus nsp1 Protein Induces Template-Dependent Endonucleolytic Cleavage of mRNAs: Viral mRNAs Are Resistant to nsp1-Induced RNA Cleavage. PLoS Pathog. 2011, 7, e1002433. [Google Scholar] [CrossRef] [PubMed]
- Lokugamage, K.G.; Narayanan, K.; Nakagawa, K.; Terasaki, K.; Ramirez, S.I.; Tseng, C.-T.K.; Ramirez, S.I.; Tseng, C.T.; Makino, S. Middle East respiratory syndrome coronavirus nsp1 inhibits host gene expression by selectively targeting nuclear-transcribed mRNAs but spares mRNAs of cytoplasmic origin. J. Virol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Jauregui, A.R.; Savalia, D.; Lowry, V.K.; Farrell, C.M.; Wathelet, M.G. Identification of Residues of SARS-CoV nsp1 That Differentially Affect Inhibition of Gene Expression and Antiviral Signaling. PLoS ONE 2013, 8, e62416. [Google Scholar] [CrossRef] [PubMed]
- Mielech, A.M.; Kilianski, A.; Baez-Santos, Y.M.; Mesecar, A.D.; Baker, S.C. MERS-CoV papain-like protease has deISGylating and deubiquitinating activities. Virology 2014, 450–451, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Devaraj, S.G.; Wang, N.; Chen, Z.; Chen, Z.; Tseng, M.; Barretto, N.; Lin, R.; Peters, C.J.; Tseng, C.T.; Baker, S.C.; et al. Regulation of IRF-3 dependent innate immunity by the papain-like protease domain of the sars coronavirus. J. Biol. Chem. 2007, 282, 32208–32221. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Chen, X.; Bian, G.; Tu, J.; Xing, Y.; Wang, Y.; Chen, Z. Proteolytic processing, deubiquitinase and interferon antagonist activities of Middle East respiratory syndrome coronavirus papain-like protease. J. Gen. Virol. 2014, 95, 614–626. [Google Scholar] [CrossRef] [PubMed]
- Kuri, T.; Eriksson, K.K.; Putics, A.; Züst, R.; Snijder, E.J.; Davidson, A.D.; Siddell, S.G.; Thiel, V.; Ziebuhr, J.; Weber, F. The ADP-ribose-1′′-monophosphatase domains of severe acute respiratory syndrome coronavirus and human coronavirus 229E mediate resistance to antiviral interferon responses. J. Gen. Virol. 2011, 92, 1899–1905. [Google Scholar] [CrossRef] [PubMed]
- Stevens, F.J.; Argon, Y. Protein folding in the ER. Semin. Cell Dev. Biol. 1999, 10, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Liu, D.X. Coronavirus infection, ER stress, apoptosis and innate immunity. Front. Microbiol. 2014, 5, 296. [Google Scholar] [CrossRef] [PubMed]
- Teske, B.F.; Wek, S.A.; Bunpo, P.; Cundiff, J.K.; McClintick, J.N.; Anthony, T.G.; Wek, R.C. The eIF2 kinase PERK and the integrated stress response facilitate activation of ATF6 during endoplasmic reticulum stress. Mol. Biol. Cell 2011, 22, 4390–4405. [Google Scholar] [CrossRef] [PubMed]
- Harding, H.P.; Zhang, Y.; Bertolotti, A.; Zeng, H.; Ron, D. Perk Is Essential for Translational Regulation and Cell Survival during the Unfolded Protein Response. Mol. Cell 2000, 5, 897–904. [Google Scholar] [CrossRef]
- Harding, H.P.; Novoa, I.; Zhang, Y.; Zeng, H.; Wek, R.; Schapira, M.; Ron, D. Regulated Translation Initiation Controls Stress-Induced Gene Expression in Mammalian Cells. Mol. Cell 2000, 6, 1099–1108. [Google Scholar] [CrossRef]
- Oyadomari, S.; Mori, M. Roles of CHOP//GADD153 in endoplasmic reticulum stress. Cell Death Differ. 2003, 11, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Huang, M.; Liu, D.X. Coronavirus-induced ER stress response and its involvement in regulation of coronavirus-host interactions. Virus Res. 2014, 194, 110–123. [Google Scholar] [CrossRef] [PubMed]
- Minakshi, R.; Padhan, K.; Rani, M.; Khan, N.; Ahmad, F.; Jameel, S. The SARS Coronavirus 3a Protein Causes Endoplasmic Reticulum Stress and Induces Ligand-Independent Downregulation of the Type 1 Interferon Receptor. PLoS ONE 2009, 4, e8342. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.-P.; Siu, K.-L.; Chin, K.-T.; Yuen, K.-Y.; Zheng, B.; Jin, D.-Y. Modulation of the Unfolded Protein Response by the Severe Acute Respiratory Syndrome Coronavirus Spike Protein. J. Virol. 2006, 80, 9279–9287. [Google Scholar] [CrossRef] [PubMed]
- Siu, K.-L.; Chan, C.-P.; Kok, K.-H.; C-Y Woo, P.; Jin, D.-Y. Comparative analysis of the activation of unfolded protein response by spike proteins of severe acute respiratory syndrome coronavirus and human coronavirus HKU1. Cell Biosci. 2014, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Fung, T.S.; Huang, M.; Fang, S.G.; Zhong, Y.; Liu, D.X. Upregulation of CHOP/GADD153 during Coronavirus Infectious Bronchitis Virus Infection Modulates Apoptosis by Restricting Activation of the Extracellular Signal-Regulated Kinase Pathway. J. Virol. 2013, 87, 8124–8134. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liao, Y.; Yap, P.L.; Png, K.J.; Tam, J.P.; Liu, D.X. Inhibition of Protein Kinase R Activation and Upregulation of GADD34 Expression Play a Synergistic Role in Facilitating Coronavirus Replication by Maintaining De Novo Protein Synthesis in Virus-Infected Cells. J. Virol. 2009, 83, 12462–12472. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Luo, S.; Baumeister, P.; Huang, J.-M.; Gogia, R.K.; Li, M.; Lee, A.S. Underglycosylation of ATF6 as a Novel Sensing Mechanism for Activation of the Unfolded Protein Response. J. Biol. Chem. 2004, 279, 11354–11363. [Google Scholar] [CrossRef] [PubMed]
- Nadanaka, S.; Okada, T.; Yoshida, H.; Mori, K. Role of Disulfide Bridges Formed in the Luminal Domain of ATF6 in Sensing Endoplasmic Reticulum Stress. Mol. Cell. Biol. 2007, 27, 1027–1043. [Google Scholar] [CrossRef] [PubMed]
- Schröder, M.; Kaufman, R.J. The mammalian unfolded protein response. Annu. Rev. Biochem. 2005, 74, 739–789. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Matsui, T.; Yamamoto, A.; Okada, T.; Mori, K. XBP1 mRNA Is Induced by ATF6 and Spliced by IRE1 in Response to ER Stress to Produce a Highly Active Transcription Factor. Cell 2001, 107, 881–891. [Google Scholar] [CrossRef]
- Sung, S.-C.; Chao, C.-Y.; Jeng, K.-S.; Yang, J.-Y.; Lai, M.M.C. The 8ab protein of SARS-CoV is a luminal ER membrane-associated protein and induces the activation of ATF6. Virology 2009, 387, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Oostra, M.; de Haan, C.A.M.; Rottier, P.J.M. The 29-Nucleotide Deletion Present in Human but Not in Animal Severe Acute Respiratory Syndrome Coronaviruses Disrupts the Functional Expression of Open Reading Frame 8. J. Virol. 2007, 81, 13876–13888. [Google Scholar] [CrossRef] [PubMed]
- Szegezdi, E.; Logue, S.E.; Gorman, A.M.; Samali, A. Mediators of endoplasmic reticulum stress-induced apoptosis. EMBO Rep. 2006, 7, 880–885. [Google Scholar] [CrossRef] [PubMed]
- Mori, K. Signalling Pathways in the Unfolded Protein Response: Development from Yeast to Mammals. J. Biochem. 2009, 146, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Credle, J.J.; Finer-Moore, J.S.; Papa, F.R.; Stroud, R.M.; Walter, P. On the mechanism of sensing unfolded protein in the endoplasmic reticulum. Proc. Natl. Acad. Sci. USA 2005, 102, 18773–18784. [Google Scholar] [CrossRef] [PubMed]
- Gardner, B.M.; Walter, P. Unfolded Proteins Are Ire1-Activating Ligands That Directly Induce the Unfolded Protein Response. Science 2011, 333, 1891–1894. [Google Scholar] [CrossRef] [PubMed]
- Prischi, F.; Nowak, P.R.; Carrara, M.; Ali, M.M.U. Phosphoregulation of Ire1 RNase splicing activity. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Gao, B.; Lee, S.-M.; Chen, A.; Zhang, J.; Zhang, D.D.; Kannan, K.; Ortmann, R.A.; Fang, D. Synoviolin promotes IRE1 ubiquitination and degradation in synovial fibroblasts from mice with collagen-induced arthritis. EMBO Rep. 2008, 9, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Oku, M.; Suzuki, M.; Mori, K. pXBP1(U) encoded in XBP1 pre-mRNA negatively regulates unfolded protein response activator pXBP1(S) in mammalian ER stress response. J. Cell Biol. 2006, 172, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Tam, A.B.; Koong, A.C.; Niwa, M. Ire1 Has Distinct Catalytic Mechanisms for XBP1/HAC1 Splicing and RIDD. Cell Rep. 2014, 9, 850–858. [Google Scholar]
- Walter, F.; Schmid, J.; Dussmann, H.; Concannon, C.G.; Prehn, J.H.M. Imaging of single cell responses to ER stress indicates that the relative dynamics of IRE1/XBP1 and PERK/ATF4 signalling rather than a switch between signalling branches determine cell survival. Cell Death Differ. 2015, 22, 1502–1516. [Google Scholar] [CrossRef] [PubMed]
- Urano, F.; Wang, X.; Bertolotti, A.; Zhang, Y.; Chung, P.; Harding, H.P.; Ron, D. Coupling of Stress in the ER to Activation of JNK Protein Kinases by Transmembrane Protein Kinase IRE1. Science 2000, 287, 664–666. [Google Scholar] [CrossRef] [PubMed]
- Versteeg, G.A.; van de Nes, P.S.; Bredenbeek, P.J.; Spaan, W.J.M. The Coronavirus Spike Protein Induces Endoplasmic Reticulum Stress and Upregulation of Intracellular Chemokine mRNA Concentrations. J. Virol. 2007, 81, 10981–10990. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S.; Liao, Y.; Liu, D.X. The Endoplasmic Reticulum Stress Sensor IRE1α Protects Cells from Apoptosis Induced by the Coronavirus Infectious Bronchitis Virus. J. Virol. 2014, 88, 12752–12764. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 0000, 26, 3279–3290. [Google Scholar] [CrossRef] [PubMed]
- Davis, R.J. Signal Transduction to the Nucleus by MAP Kinase. In Signaling Networks and Cell Cycle Control: The Molecular Basis of Cancer and Other Diseases; Gutkind, J.S., Ed.; Humana Press: Totowa, NJ, USA, 2000; pp. 153–164. [Google Scholar]
- Panteva, M.; Korkaya, H.; Jameel, S. Hepatitis viruses and the MAPK pathway: Is this a survival strategy? Virus Res. 2003, 92, 131–140. [Google Scholar] [CrossRef]
- Arthur, J.S.C.; Ley, S.C. Mitogen-activated protein kinases in innate immunity. Nat. Rev. Immunol. 2013, 13, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Keshet, Y.; Seger, R. The MAP Kinase Signaling Cascades: A System of Hundreds of Components Regulates a Diverse Array of Physiological Functions. In MAP Kinase Signaling Protocols, 2nd ed.; Peger, R., Ed.; Humana Press: Totowa, NJ, USA, 2010; pp. 3–38. [Google Scholar]
- Mizutani, T.; Fukushi, S.; Murakami, M.; Hirano, T.; Saijo, M.; Kurane, I.; Morikawa, S. Tyrosine dephosphorylation of STAT3 in SARS coronavirus-infected Vero E6 cells. FEBS Lett. 2004, 577, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Fukushi, S.; Saijo, M.; Kurane, I.; Morikawa, S. Phosphorylation of p38 MAPK and its downstream targets in SARS coronavirus-infected cells. Biochem. Biophys. Res. Commun. 2004, 319, 1228–1234. [Google Scholar] [CrossRef] [PubMed]
- Chen, I.Y.; Chang, S.C.; Wu, H.-Y.; Yu, T.-C.; Wei, W.-C.; Lin, S.; Chien, C.L.; Chang, M.F. Upregulation of the Chemokine (C-C Motif) Ligand 2 via a Severe Acute Respiratory Syndrome Coronavirus Spike-ACE2 Signaling Pathway. J. Virol. 2010, 84, 7703–7712. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Fukushi, S.; Saijo, M.; Kurane, I.; Morikawa, S. Regulation of p90RSK phosphorylation by SARS-CoV infection in Vero E6 cells. FEBS Lett. 2006, 580, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Li, W.; Wu, J.; Germann, U.A.; Su, M.S.; Kuida, K.; Boucher, D.M. Extracellular Signal-Regulated Kinase 2 Is Necessary for Mesoderm Differentiation. Proc. Natl. Acad. Sci. USA 2003, 100, 12759–12764. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.T.-C.; Chien, S.-C.; Chen, I.Y.; Lai, C.-T.; Tsay, Y.-G.; Chang, S.C.; Chang, M.F. Surface vimentin is critical for the cell entry of SARS-CoV. J. Biomed.Sci. 2016, 23, 14. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Robidoux, J.; Daniel, K.W.; Guzman, G.; Floering, L.M.; Collins, S. Requirement of Vimentin Filament Assembly for β3-Adrenergic Receptor Activation of ERK MAP Kinase and Lipolysis. J. Biol. Chem. 2007, 282, 9244–9250. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-J.; Liu, C.Y.-Y.; Chiang, B.-L.; Chao, Y.-C.; Chen, C.-C. Induction of IL-8 Release in Lung Cells via Activator Protein-1 by Recombinant Baculovirus Displaying Severe Acute Respiratory Syndrome-Coronavirus Spike Proteins: Identification of Two Functional Regions. J. Immunol. 2004, 173, 7602–7614. [Google Scholar] [CrossRef] [PubMed]
- Li, S.-W.; Lai, C.-C.; Ping, J.-F.; Tsai, F.-J.; Wan, L.; Lin, Y.-J.; Kung, S.H.; Lin, C.W. Severe acute respiratory syndrome coronavirus papain-like protease suppressed alpha interferon-induced responses through downregulation of extracellular signal-regulated kinase 1-mediated signalling pathways. J. Gen. Virol. 2011, 92, 1127–1140. [Google Scholar] [CrossRef] [PubMed]
- Varshney, B.; Lal, S. SARS-CoV Accessory Protein 3b Induces AP-1 Transcriptional Activity through Activation of JNK and ERK Pathways. Biochemistry 2011, 50, 5419–5425. [Google Scholar] [CrossRef] [PubMed]
- Kindrachuk, J.; Ork, B.; Hart, B.J.; Mazur, S.; Holbrook, M.R.; Frieman, M.B.; Traynor, D.; Johnson, R.F.; Dyall, J.; Kuhn, J.H.; et al. Antiviral Potential of ERK/MAPK and PI3K/AKT/mTOR Signaling Modulation for Middle East Respiratory Syndrome Coronavirus Infection as Identified by Temporal Kinome Analysis. Antimicrob. Agents Chemother. 2015, 59, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Kono, M.; Tatsumi, K.; Imai, A.M.; Saito, K.; Kuriyama, T.; Shirasawa, H. Inhibition of human coronavirus 229E infection in human epithelial lung cells (L132) by chloroquine: Involvement of p38 MAPK and ERK. Antivir. Res. 2008, 77, 150–152. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, T.; Fukushi, S.; Saijo, M.; Kurane, I.; Morikawa, S. JNK and PI3k/Akt signaling pathways are required for establishing persistent SARS-CoV infection in Vero E6 cells. Biochim. Biophys. Acta 2005, 1741, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Kanzawa, N.; Nishigaki, K.; Hayashi, T.; Ishii, Y.; Furukawa, S.; Niiro, A.; Yasui, F.; Kohara, M.; Morita, K.; Matsushima, K.; et al. Augmentation of chemokine production by severe acute respiratory syndrome coronavirus 3a/X1 and 7a/X4 proteins through NF-κB activation. FEBS Lett. 2006, 580, 6807–6812. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Yang, Y.; Gu, C.; Yue, Y.; Wu, K.K.; Wu, J.; Zhu, Y. Spike protein of SARS-CoV stimulates cyclooxygenase-2 expression via both calcium-dependent and calcium-independent protein kinase C pathways. FASEB J. 2007, 21, 1586–1596. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.S. Molecular Characterization of Cellular Stress Responses during Coronavirus Infection. Ph.D. Thesis, Nanyang Technological University, Singapore, Singapore, 2015. [Google Scholar]
- Kopecky-Bromberg, S.A.; Martinez-Sobrido, L.; Palese, P. 7a Protein of Severe Acute Respiratory Syndrome Coronavirus Inhibits Cellular Protein Synthesis and Activates p38 Mitogen-Activated Protein Kinase. J. Virol. 2006, 80, 785–793. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.-J.; Teng, E.; Shen, S.; Tan, T.H.P.; Goh, P.-Y.; Fielding, B.C.; Ooi, E.E.; Tan, H.C.; Lim, S.G.; Hong, W. A Novel Severe Acute Respiratory Syndrome Coronavirus Protein, U274, Is Transported to the Cell Surface and Undergoes Endocytosis. J. Virol. 2004, 78, 6723–6734. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Guardeño, J.M.; Nieto-Torres, J.L.; DeDiego, M.L.; Regla-Nava, J.A.; Fernandez-Delgado, R.; Castaño-Rodriguez, C.; Enjuanes, L. The PDZ-Binding Motif of Severe Acute Respiratory Syndrome Coronavirus Envelope Protein Is a Determinant of Viral Pathogenesis. PLoS Pathog. 2014, 10, e1004320. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Chen, R.-F.; Liu, J.-W.; Yeh, W.-T.; Chang, J.-C.; Liu, P.-M.; Eng, H.L.; Lin, M.C.; Yang, K.D. Altered p38 Mitogen-Activated Protein Kinase Expression in Different Leukocytes with Increment of Immunosuppressive Mediators in Patients with Severe Acute Respiratory Syndrome. J. Immunol. 2004, 172, 7841–7847. [Google Scholar] [CrossRef] [PubMed]
- Santoro, M.; Rossi, A.; Amici, C. NEW EMBO MEMBER’S REVIEW: NF-κB and virus infection: Who controls whom. EMBO J. 2003, 22, 2552–2560. [Google Scholar] [CrossRef] [PubMed]
- Hoesel, B.; Schmid, J.A. The complexity of NF-κB signaling in inflammation and cancer. Mol. Cancer 2013, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hiscott, J.; Kwon, H.; Génin, P. Hostile takeovers: Viral appropriation of the NF-κB pathway. J. Clin. Investig. 2001, 107, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.-C. Non-canonical NF-κB signaling pathway. Cell Res. 2011, 21, 71–85. [Google Scholar] [CrossRef] [PubMed]
- De Diego, M.L.; Nieto-Torres, J.L.; Regla-Nava, J.A.; Jimenez-Guardeño, J.M.; Fernandez-Delgado, R.; Fett, C.; Castaño-Rodriguez, C.; Perlman, S.; Enjuanes, L. Inhibition of NF-κB-Mediated Inflammation in Severe Acute Respiratory Syndrome Coronavirus-Infected Mice Increases Survival. J. Virol. 2014, 88, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Dosch, S.F.; Mahajan, S.D.; Collins, A.R. SARS Coronavirus Spike Protein-Induced Innate Immune Response occurs via Activation of the NF-κB pathway in Human Monocyte Macrophages in vitro. Virus Res. 2009, 142, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.-J.; Ye, L.-B.; Timani, K.A.; Zeng, Y.-C.; She, Y.-L.; Ye, L.; Wu, Z.H. Activation of NF-κB by the Full-length Nucleocapsid Protein of the SARS Coronavirus. Acta Biochim. Biophys. Sin. 2005, 37, 607–612. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Leeson, A.; Andonov, A.; Li, Y.; Bastien, N.; Cao, J.; Osiowy, C.; Dobie, F.; Cutts, T.; Ballantine, M.; et al. Activation of AP-1 signal transduction pathway by SARS coronavirus nucleocapsid protein. Biochem. Biophys. Res. Commun. 2003, 311, 870–876. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Gao, J.; Zheng, H.; Li, B.; Kong, L.; Zhang, Y.; Wang, W.; Zeng, Y.; Ye, L. The membrane protein of SARS-CoV suppresses NF-κB activation. J. Med. Virol. 2007, 79, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Lai, F.W.; Stephenson, K.B.; Mahony, J.; Lichty, B.D. Human Coronavirus OC43 Nucleocapsid Protein Binds MicroRNA 9 and Potentiates NF-κB Activation. J. Virol. 2014, 88, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Law, A.H.Y.; Lee, D.C.W.; Cheung, B.K.W.; Yim, H.C.H.; Lau, A.S.Y. Role for Nonstructural Protein 1 of Severe Acute Respiratory Syndrome Coronavirus in Chemokine Dysregulation. J. Virol. 2007, 81, 416–422. [Google Scholar] [CrossRef] [PubMed]
- Frieman, M.; Ratia, K.; Johnston, R.E.; Mesecar, A.D.; Baric, R.S. Severe Acute Respiratory Syndrome Coronavirus Papain-Like Protease Ubiquitin-Like Domain and Catalytic Domain Regulate Antagonism of IRF3 and NF-κB Signaling. J. Virol. 2009, 83, 6689–6705. [Google Scholar] [CrossRef] [PubMed]
- Ratia, K.; Kilianski, A.; Baez-Santos, Y.M.; Baker, S.C.; Mesecar, A. Structural Basis for the Ubiquitin-Linkage Specificity and deISGylating Activity of SARS-CoV Papain-Like Protease. PLoS Pathog. 2014, 10, e1004113. [Google Scholar] [CrossRef] [PubMed]
- Matthews, K.L.; Coleman, C.M.; van der Meer, Y.; Snijder, E.J.; Frieman, M.B. The ORF4b-encoded accessory proteins of Middle East respiratory syndrome coronavirus and two related bat coronaviruses localize to the nucleus and inhibit innate immune signalling. J. Gen. Virol. 2014, 95, 874–882. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).