1. Introduction
The perennially ice covered lakes of the McMurdo Dry Valleys (MDVs), Antarctica, are amongst the most extreme lacustrine environments on Earth. Already set apart from most lakes by their endorheic and meromictic characters, the lakes also have extreme photoperiods, are persistently cold and are almost isolated from all physical forcing by perennial ice cover. In addition, they contain truncated, microbially-dominated food webs within which higher metazoans are largely absent. These conditions combine to result in minimal physical and biological perturbations of lake sediments, a situation that favors the formation of well-developed, complex microbial mat communities [
1,
2]. In response, microbial mats are both widespread and abundant in the lakes of the MDV [
3,
4], where they play a major role in carbon and nutrient cycling on a lake-wide basis [
5,
6,
7].
While it is acknowledged that benthic microbial communities are significant components of the MDV lake systems, to date, quantitative information on their self-organization and growth is sparse, in part because accumulation rates tend to be slow and long duration observations would be required to assess how communities change. However, in recent decades, the MDV lakes have been undergoing gradual increases in lake level, in response to persistently warm summer temperatures [
8,
9]. This rising level provides a natural experiment into the rate and trajectory of microbial mat development on recently inundated soils and on the effects of changing environment on established microbial mat communities as levels increase. In Lake Joyce, for example, we have shown that reduced irradiant flux to deep mat communities has resulted in the extinction of distinctive components of the benthic biota, while inundation of substrates provide opportunities for colonization, albeit at a slow pace [
10]. The relationship between the speed at which lake levels rise and how various communities respond has emerged as an important consideration in understanding ecosystem responses to change [
7].
Lake Vanda, one of the MDV lakes, provides a particularly favorable opportunity to study the temporal characteristics of microbial mat development. In Lake Vanda, a punctuated rise in lake level has been well documented since 1973 [
11], which allows us to construct a precise time-series of inundation. In addition, the upper waters of this lake are well mixed by convective processes [
12] and the water column is very clear [
13]. This means that conditions for microbial development in the zone inundated by lake level rise are not confounded by depth-related changes in water chemistry or by steep light gradients. The mats are internally laminated [
14], and there is good evidence from other nearby lakes to suggest that these laminae record annual growth [
5], allowing cross-validation of estimates of mat age. In this contribution, we present a detailed examination of the microbial mats in Lake Vanda, focusing on those that can be dated by recent changes in lake level. We use measurements made at similar depths and locations in 1998 and 2010 to describe temporal changes in mats and discuss these results in the context of the timescales of environmental change and ecosystem response.
3. Results
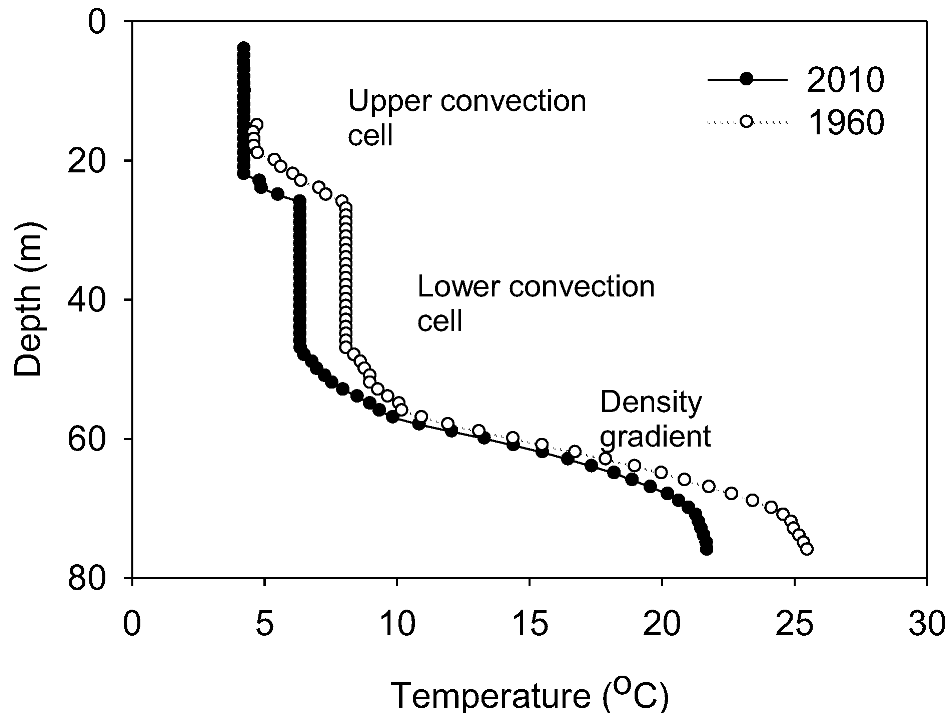
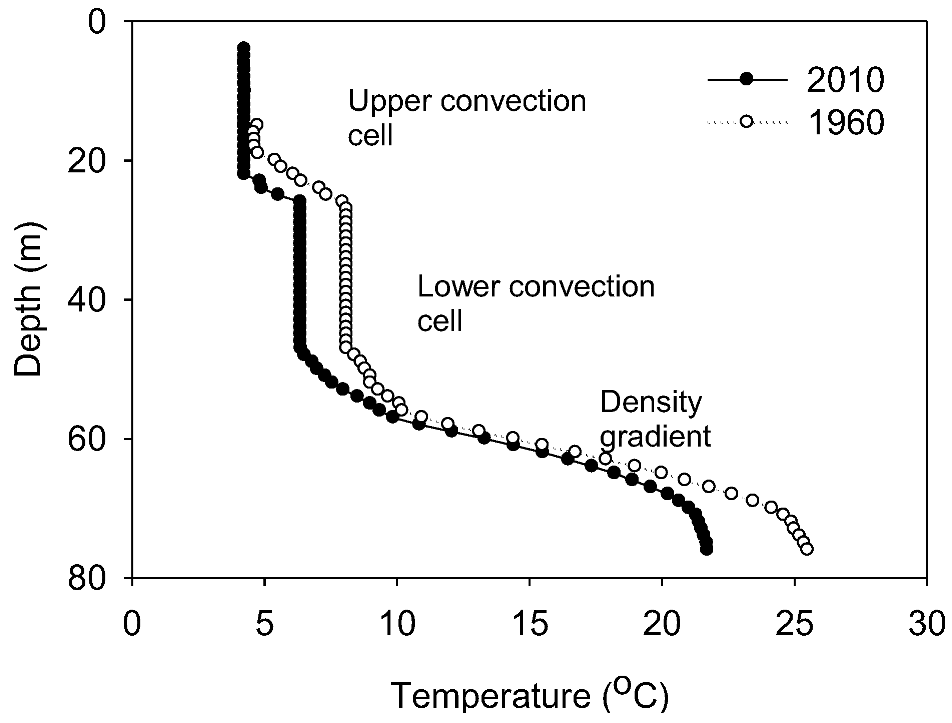
Throughout this section, all depths are referenced to water level in 2010. Our water column profile (
Figure S2) identified upper and lower isohaline and isothermal convection cells overlying a continuous density gradient, and the locations of the discontinuities are consistent with other published profiles of the water column [
12], confirming that lake level rise had been achieved by an increase in the thickness of the upper convection cell.
3.1. Macroscale Morphology
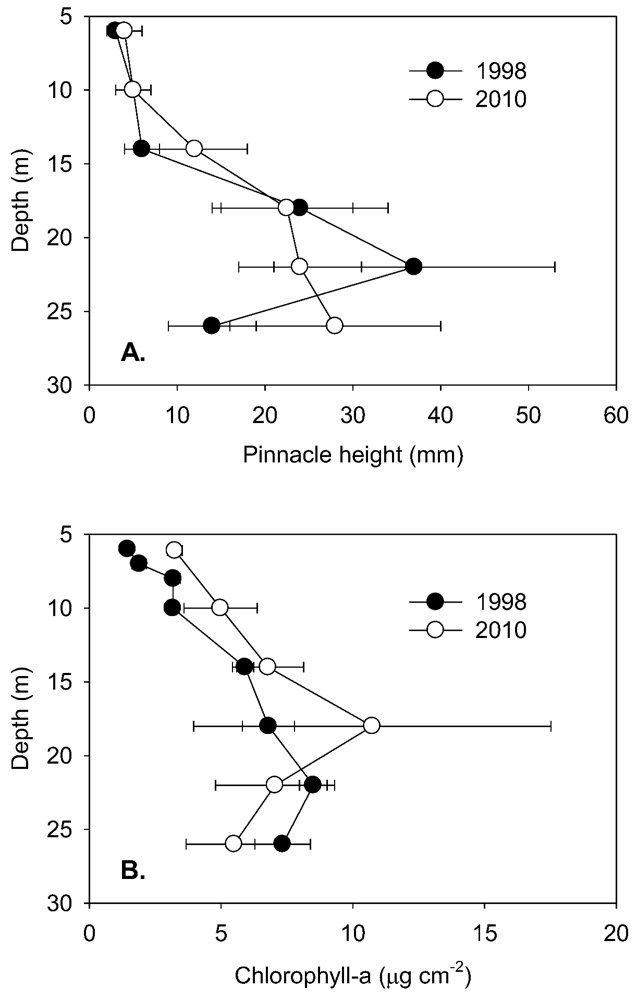
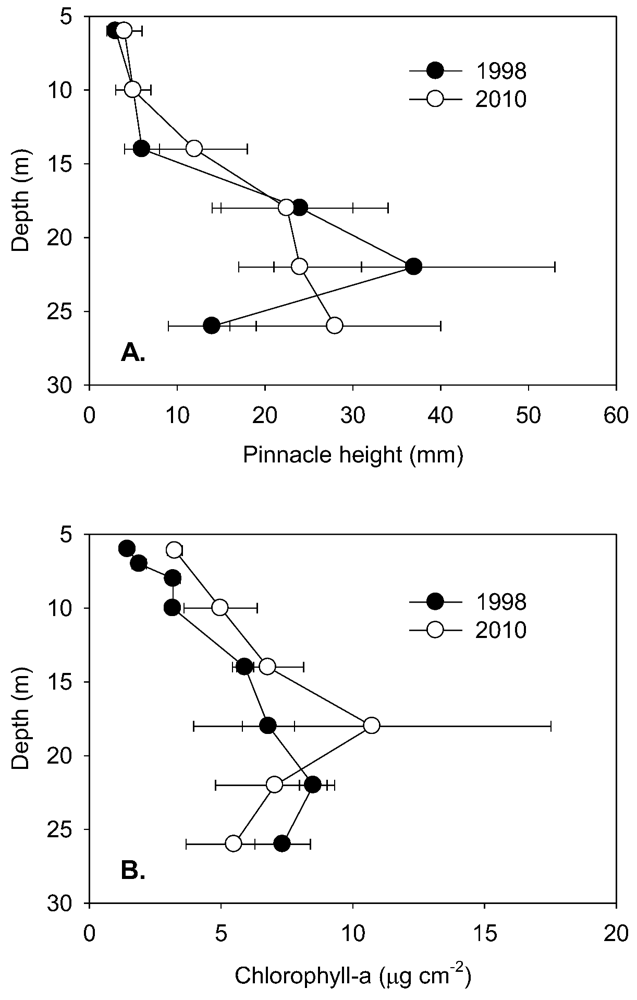
To the maximum depth sampled, the lake floor was covered by a flat to undulating mat from which pinnacles of varying height emerged (
Figure 2). Mat thickness increased with depth on both sampling occasions and, over the 6–18 m depth interval, also increased between samplings (
Table 1). The rate of increase in thickness increased with depth to 18 m, with 12 years of growth producing an increased thickness of 0.3–1.1 mm y
−1 (
Table 1). Pinnacle height also tended to increase with depth into the lake (
Figure 3A), with a discontinuity in the rate of increase at 10–15 m and an apparent maximum height reached at ~22–26 m. Pinnacle height, even after log-transformation, was highly variable and not normally distributed, and a Kruskal-Wallis ANOVA by ranks yielded no significant differences (at
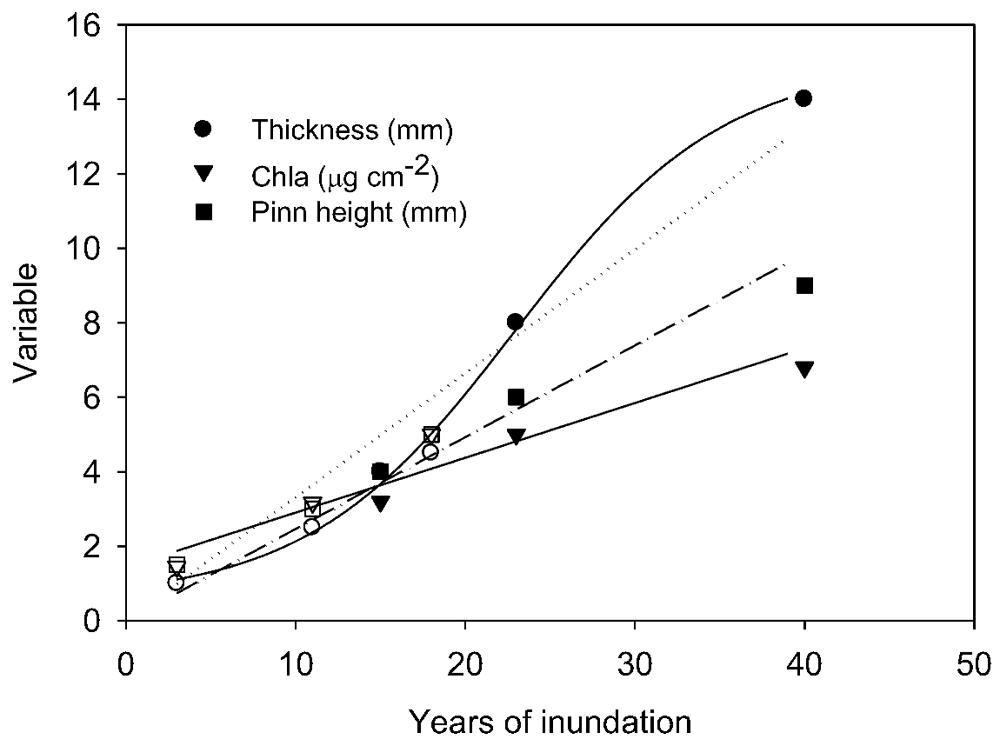
p = 0.05) at specific depths between 1998 and 2010 samplings. In 2010, ANOVA by ranks clustered pinnacle height into three depth-groups, from 6 and 10 m (group median height 5 mm) and from 18, 22 and 26 m depth (group median height 24 mm), while those from 14 m (median height 12 mm) fell significantly between these groups. By plotting thickness and median pinnacle height against time under ice, where this last could be estimated, a consistent trend of a net increase over the early period of inundation was apparent (
Figure 4). A highly significant linear increase in pinnacle height of ~0.3 mm y
−1 (r
2 = 0.97,
p < 0.01) was seen in the known-age part of the water column, that is, from 6–14 m. For thickness, a significant linear relationship was also apparent (r
2 = 0.80, n = 6,
p < 0.05), suggesting a rate of growth of 0.14 mm y
−1. A curve fitting exercise comparing a range of models for these time-dependent data suggested that there was a tendency for rates of increase to decline after several decades (
Table S1), though the estimates of initial rates of increase were very similar to those obtained from linear fits.
Table 1.
Characteristics of microbial mats along a depth profile in Lake Vanda in 1998 and in 2010. Depths are referred to 2010 lake levels.
Table 1.
Characteristics of microbial mats along a depth profile in Lake Vanda in 1998 and in 2010. Depths are referred to 2010 lake levels.
| Depth in 2010 (m) | Approximate age in 1998 (y) | Total thickness of flat mat 1998 (number of laminae) | Total thickness of flat mat 2010 (number of laminae) | Annualized rate of increase in thickness (number of laminae) |
|---|
| 6 | 6 | 1 mm (2–3) | 4 mm (12) | 0.3 (0.85) |
| 10 | 10 | 1–2 mm (7) | 8 mm (18) | 0.5 (0.92) |
| 14 | 28 | 4 mm (>12) | 14 mm (>20) | 0.8 (-) |
| 18 | >50 | 5 mm (>14) | 18 mm (>20) | 1.1 (-) |
| 22 | >50 | >15 mm | 25 mm | - (-) |
| 26 | >50 | >50 mm | >45 mm | - (-) |
Figure 4.
Relationships between median pinnacle height, areal chlorophyll-a content and thickness of flat mat from a range of depths, expressed as time under ice. Solid symbols represent samples taken in 2010; open symbols in 1998. Linear regression models are fitted to each variable for all samples and are significant at p < 0.01. For the mat thickness, a sigmoid curve provided a better fit (r2 = 0.998 cf. r2 = 0.959 for the linear model). Pinn height—pinnacle height.
Figure 4.
Relationships between median pinnacle height, areal chlorophyll-a content and thickness of flat mat from a range of depths, expressed as time under ice. Solid symbols represent samples taken in 2010; open symbols in 1998. Linear regression models are fitted to each variable for all samples and are significant at p < 0.01. For the mat thickness, a sigmoid curve provided a better fit (r2 = 0.998 cf. r2 = 0.959 for the linear model). Pinn height—pinnacle height.
3.2. Internal Morphology
In both 1998 and 2010, mats contained up to four zones distinguishable by pigmentation. An orange-brown zone overlay green and then purple zones, which, in turn overlay a non-pigmented zone. The shallowest mats did not contain green and purple zones in 1998, but at 6 m depth in 2010 and 10 m depth in 1998, all zones were present. On a finer scale, each zone contained multiple laminae, which varied in thickness from several hundred microns to more than a millimeter, with some undulations in thickness within individual laminae. Laminae were defined by bands of fine sediments separating otherwise organic-rich, mucilaginous material. Sediment bands varied from barely discernible to up to 1 mm thick, and a particularly dense band of sediment was seen across depths at the base of the 9th lamina from the surface.
In 1998, at the shallowest depth sampled (equivalent to 6 m in 2010), the mat was 1 mm thick and contained only an orange-brown zone, comprising 2-3 recognizable laminae. At the same location in 2010, the mat had increased in thickness to 4 mm and contained all color zones. Four orange-brown laminae, each approximately 0.2 mm thick, capped 6-7 green-purple laminae of ~0.5 mm thickness, giving a total of ~12 organo-sedimentary laminae of which ten were pigmented.
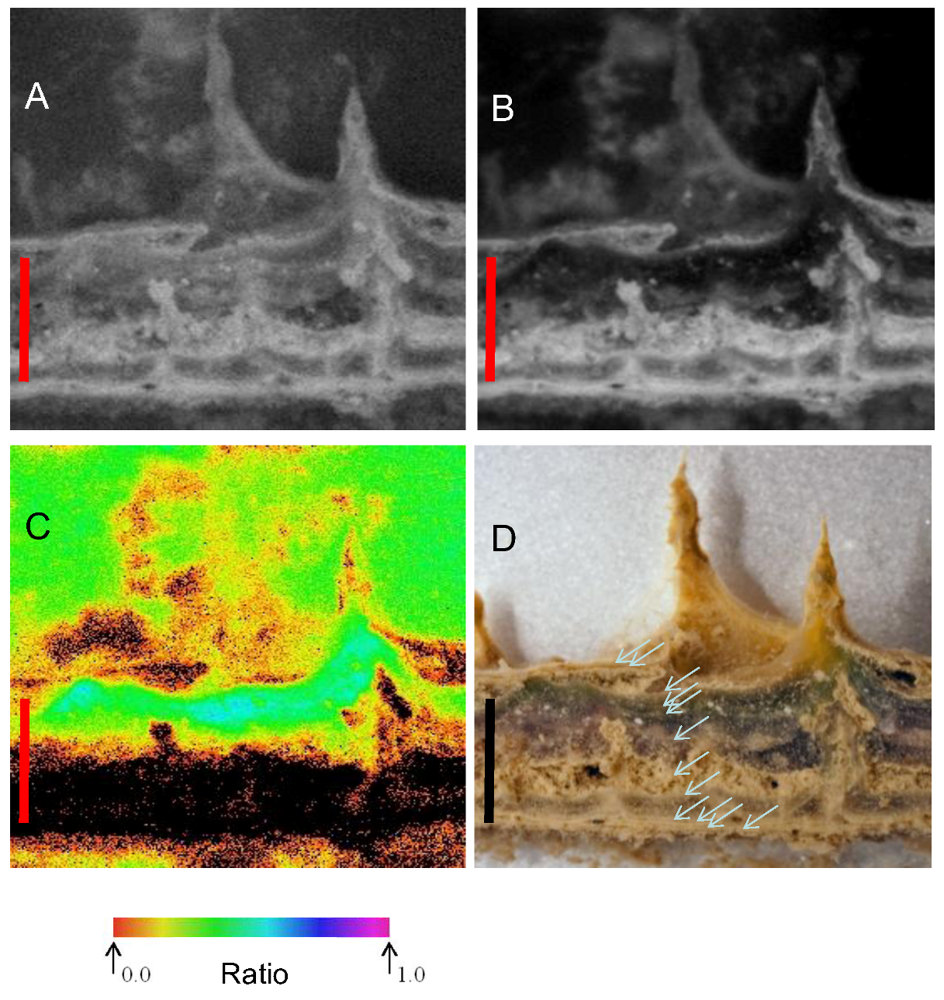
Figure 3.
A. Pinnacle heights (median and quartiles, n > 100) at selected depths in Lake Vanda in 1998 and 2010. B. Concentrations of benthic chlorophyll-a (mean and s.d., n = 5) at various levels in Lake Vanda in 1998 and 2010. Both profiles are referred to depth in 2010.
Figure 3.
A. Pinnacle heights (median and quartiles, n > 100) at selected depths in Lake Vanda in 1998 and 2010. B. Concentrations of benthic chlorophyll-a (mean and s.d., n = 5) at various levels in Lake Vanda in 1998 and 2010. Both profiles are referred to depth in 2010.
At a (2010) lake depth of 10 m, a 2 mm thick mat was present in 1998, with 7 recognizable laminae (3–4 orange, 3–4 green-purple). In 2010 the number of laminae at this location had increased to ~18, with four orange laminae overlying 6–8 green and purple laminae, fading to ~4 colorless laminae and becoming increasingly difficult to resolve. These observations showed that the subsurface laminae and zones were continuing to thicken after new laminae had formed.
At water depths exceeding 14 m, the clearly laminated and pigmented zones overlay an increasingly thick and less distinctly laminated mix of organic material and fine silts. Once the lamina count exceeded 20, counting became difficult. However, wherever counting was feasible, approximately 10–12 more laminae were present at any given depth horizon in 2010 than in 1998, consistent with the accumulation of one lamina per year (
Table 1). The picture that emerges is of mats accumulating by annual lamina accrual, with the characteristics of laminae evolving from thin, orange-brown to thicker, purple or green and, ultimately, to colorless as they are progressively overgrown.
3.3. Species Composition
Cyanobacteria formed the matrix of mats, and the dominant morphotypes on both samplings, across all lake depths and locations within the mats, were identified as belonging to the genus Leptolyngbya. In 1998, Leptolyngbya comprised 40%–50% of the total counts at all depths, with the remainder ascribable to Phormidium (5%–20%) and Oscillatoria (2%–5%). In 1998, the 6 m mat also contained 5% Nostoc, a genus not seen in 2010, but which was also common (36 and 10% relative abundance, declining with depth) in the samples from within the moat of Lake Vanda.
In 2010, four morphotypes of
Leptolyngbya were distinguished on the basis of cell dimensions. Morphotype diversity appeared to decline with depth in the lake, though our data do not allow robust statistical comparisons. At 10 m, three
Leptolyngbya morphotypes were common across all color zones, and one other was present, whereas at 18 m depth, only one morphotype (1–2 µm wide, square cells with no constriction) was dominant in all color zones (
Table S2).
Orange-brown surface zones consisted almost exclusively of
Leptolynbya, usually oriented vertically and taking the form of ropes and columns of intertwined trichomes within laminae and tending to extend vertically out of the surface of the mat as tufts. Trichomes showed a tendency to orient horizontally at lamina interfaces. The green zone, if present, was immediately below the orange layer and comprised two or three laminations.
Leptolyngbya was, again, overwhelmingly dominant.
Oscillatoria cf.
sancta,
Phormidium cf.
autumnale,
P. cf.
murrayi and a
Pseudanabaena sp also occurred in the mats, but always at low abundance. Here, as in the orange-brown zone,
Leptolyngbya trichomes tended to be oriented vertically and were intertwined. The transition from green to purple coincided with an increase in
Phormidium and
Leptolyngbya morphotypes that were visibly full of phycoerythrin. In the purple zone, trichomes were more widely spaced and less well-oriented than higher in the mat (
Table S2).
Diatoms, notably Navicula muticopsis forma murrayi (14%–28% of total diatoms), Diadesmis contenta and D. contenta var. parallela (2%–9%), Hantzschia amphioxys var. maior (7%–34%) and Muelleria peraustralis (7%–28%) were common across depths, though less so close to the ice cover, and a coccoid eukaryote, tentatively identified as a species of Chrysosococcus, was at times frequent. No clear differences were evident in diatom relative abundances between depths, and methodological differences preclude a quantitative comparison between years. Similar species lists and degrees of dominance were, however, seen across years.
3.4. Biomass and Pigments
Quantitative comparisons between 1998 and 2010 are most robust for chlorophyll-a, where analytical methods were similar in the two years. In both years, chlorophyll-a increased with lake depth to a maximum at 18 m (
Figure 3B), and in all but the deepest sample, chlorophyll-a concentrations were higher in 2010 than in 1998. However, only at 6 and 10 m were these increases statistically significant (ANOVA,
p < 0.05). Within the 6–14 m depth band, chlorophyll-a increased steadily with age (
Figure 4) equivalent to an annualized rate of 0.27 µg cm
−2 y
−1 (linear regression, r
2 = 0.90, n = 6,
p < 0.01). The intercept of chlorophyll-a with age below ice is not zero, and this most likely reflects that the under-ice mats are developing not from barren ground, but from mats that were previously in the seasonally frozen marginal zone [
14]. Comparison of other curve fits to this small data set suggested that the rate of increase in chlorophyll-a tended to decrease after some decades (
Table S1), though similar rates of increase during the first 30 years after inundation are indicated. HPLC analysis of pigments in 2010 showed similar pigment profiles, with some pigments showing proportional changes between lake depths (
Table 2). Two cyanobacterial pigments, myxoxanthophyll and nostoxanthin, showed a decline in relative abundance with depth to 14 m, whereas three pigments associated with diatoms, chlorophyll-c, diadinoxanthin and fucoxanthin, increased to 14 m. Two taxonomically widespread carotenoids, β-carotene and canthaxanthin, all tended to decline gradually with depth and reached minima at 26 m. Of the recognized chlorophyll-a derivatives, when expressed as ratio to chlorophyll-a, an allomer, phaeophytin and phaeophorbide increased with depth to 22 m, whereas chlorophyllides showed only a slight increase in relative abundance with depth. No bacteriochlorophylls were observed, though the technique used detects these when they are present.
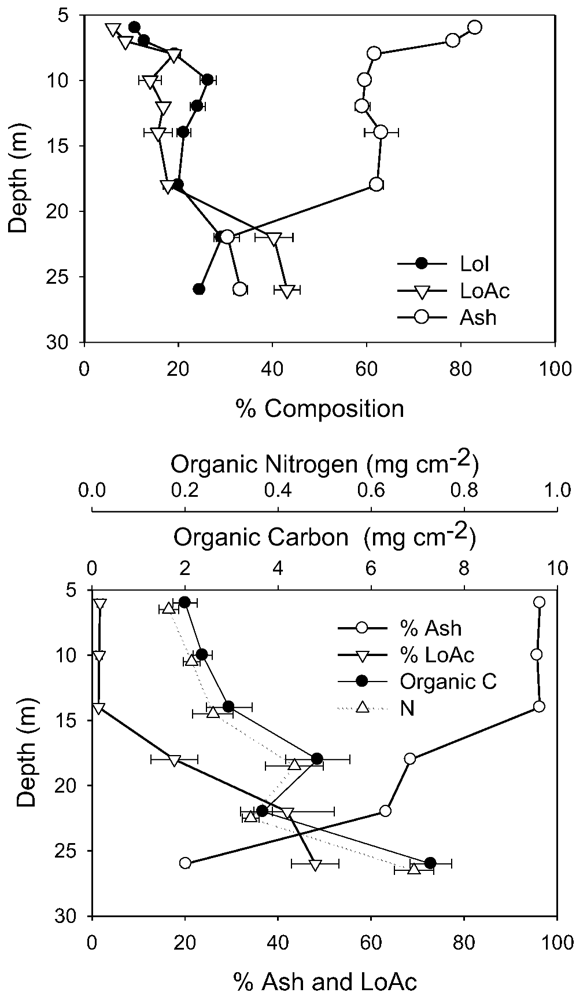
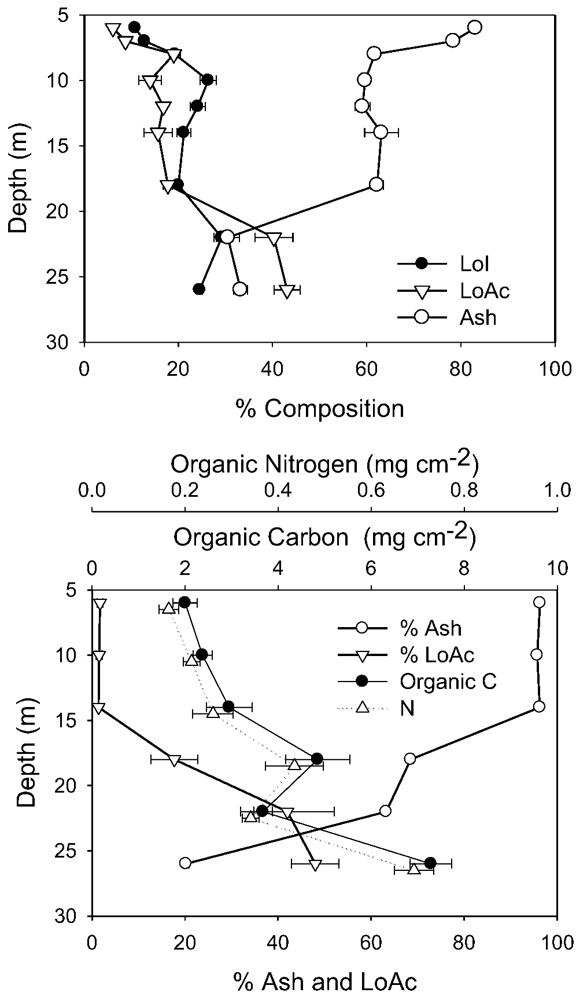
It is not possible to compare directly other biomass determinants between 1998 and 2010 due to slightly different sampling and analytical methods. However, in both years, the compositional changes with depth showed similar trends, and the differences between years were strongly associated with the ash content. In 1998, ash content was very high (90%) immediately below the ice, declining to ~65% from 8 to 18 m, then falling again to 30% at 22 m and below (
Figure 5). In 2010, ash content was also very high immediately under the ice (99%), but on this sampling remained high to 14 m (
Figure 5). However, as in 1998 ash content declined at greater depths, falling to 60% at 18 m and 20% at 26 m. The decline in % ash at 22–26 m depth was balanced by an increase in %LoAc, which in both years, reached 40%–50% here. Microscopy in 2010 supported these composition data, as shallow depth mats contained substantial amounts of both fines and sands, whereas at greater depths, sand was rare, and mineral inclusions were primarily small amounts of fine clay and precipitated calcite.
Figure 5.
Biomass-related variables from 1998 and 2010. In 1998 (top) only % composition was measured, and all depths refer to the “active layer” of mats only. In 2010 (bottom), all depths, except 26 m, comprised cores to the underlying coarse sediments and were normalized to unit area. The 26 m sample represents only the active layer. In each case, the mean and standard deviation are plotted (n = 5).
Figure 5.
Biomass-related variables from 1998 and 2010. In 1998 (top) only % composition was measured, and all depths refer to the “active layer” of mats only. In 2010 (bottom), all depths, except 26 m, comprised cores to the underlying coarse sediments and were normalized to unit area. The 26 m sample represents only the active layer. In each case, the mean and standard deviation are plotted (n = 5).
In 2010, when area-specific concentrations were measured, carbon and nitrogen both increased near exponentially to 18 m depth, closely paralleling the increase in chlorophyll-a (
Figure 3), though at 22 m depth, both C and N were anomalously low. Atomic ratios of C:N in both 1998 and 2010 were consistent across depths at 12–16 (1998) and 12–14 (2010). In 1998, N:P atomic ratio averaged 40:1 (range 33–54:1) across the 6–26 m depth range, with no clear depth-related pattern.
Table 2.
Relative abundance of selected pigments determined by high-performance liquid chromatography (HPLC) at the principal sampling depths in November 2010.
Table 2.
Relative abundance of selected pigments determined by high-performance liquid chromatography (HPLC) at the principal sampling depths in November 2010.
| | 6 m | 10 m | 14 m | 18 m | 22 m | 26 m |
|---|
| Myxoxanthophyll | 5.3 | 2.6 | 1.3 | 1.4 | 1.1 | 1.9 |
| Nostoxanthin | 1.8 | 1.0 | 0.5 | 0.6 | 0.7 | 0.9 |
| Chlorophyll-c | 0.4 | 2.1 | 3.7 | 3.9 | 4.1 | 3.9 |
| Diadinoxanthin | 0.2 | 1.2 | 2.0 | 2.2 | 2.4 | 1.0 |
| Fucoxanthin | 2.7 | 6.4 | 10.9 | 12.0 | 12.5 | 10.0 |
| β-carotene | 1.2 | 1.0 | 0.5 | 0.8 | 0.7 | 0.7 |
| Canthaxanthin | 1.6 | 1.2 | 0.9 | 0.9 | 0.9 | 0.6 |
| Chl-a allomer | 4.8 | 8.9 | 23.5 | 21.3 | 26.1 | 19.7 |
| Phaeophorbide-a | 3.0 | 11.0 | 25.4 | 29.3 | 30.6 | 23.3 |
| Chlorophyllide-a | 13.1 | 18.1 | 19.8 | 17.0 | 28.5 | 18.3 |
| Phaeophytin-a | 1.1 | 6.3 | 18.5 | 15.1 | 14.7 | 4.5 |
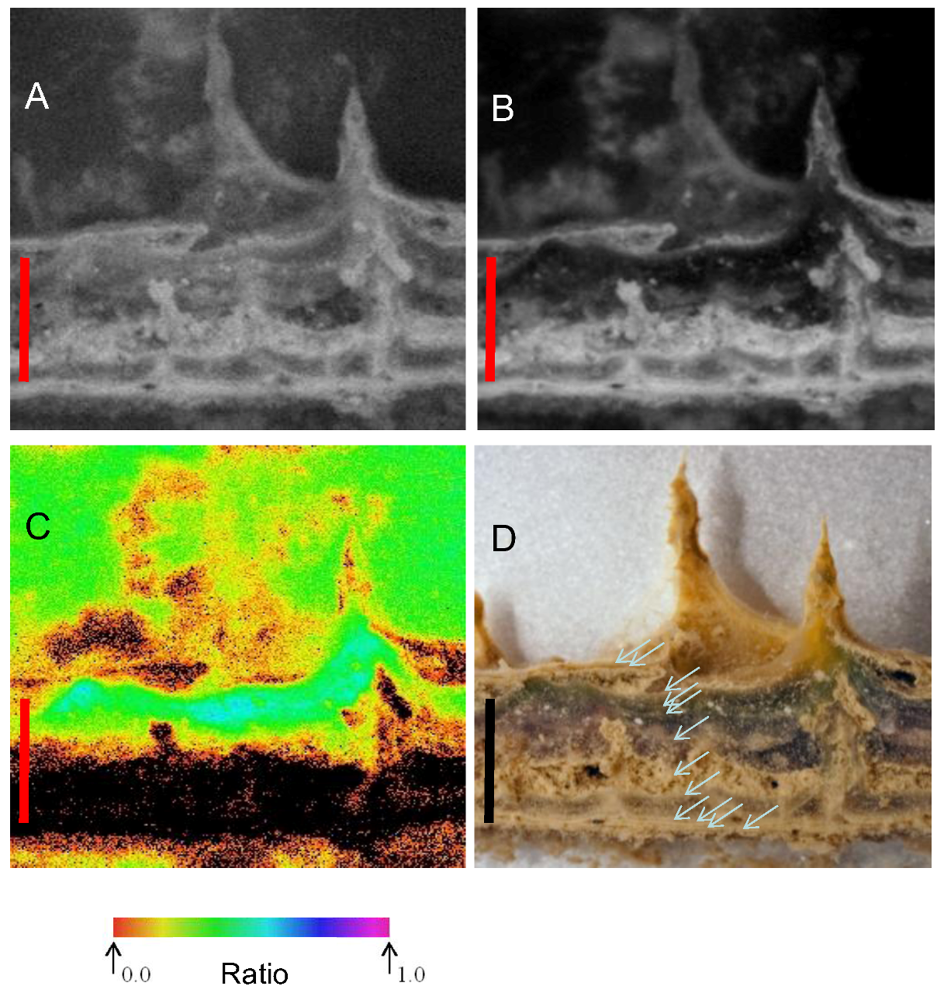
3.5. Variable Chlorophyll-a Fluorescence
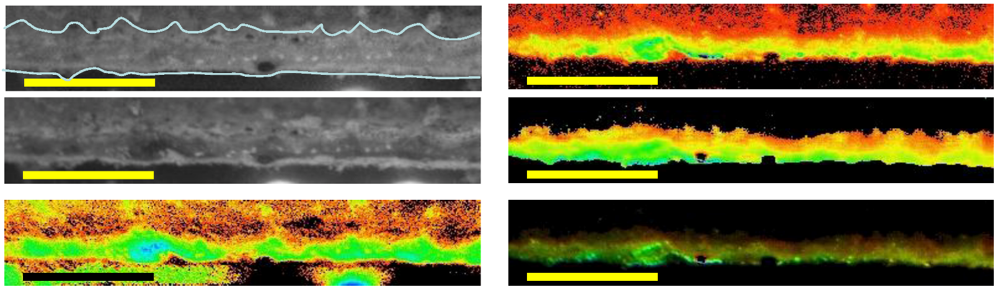
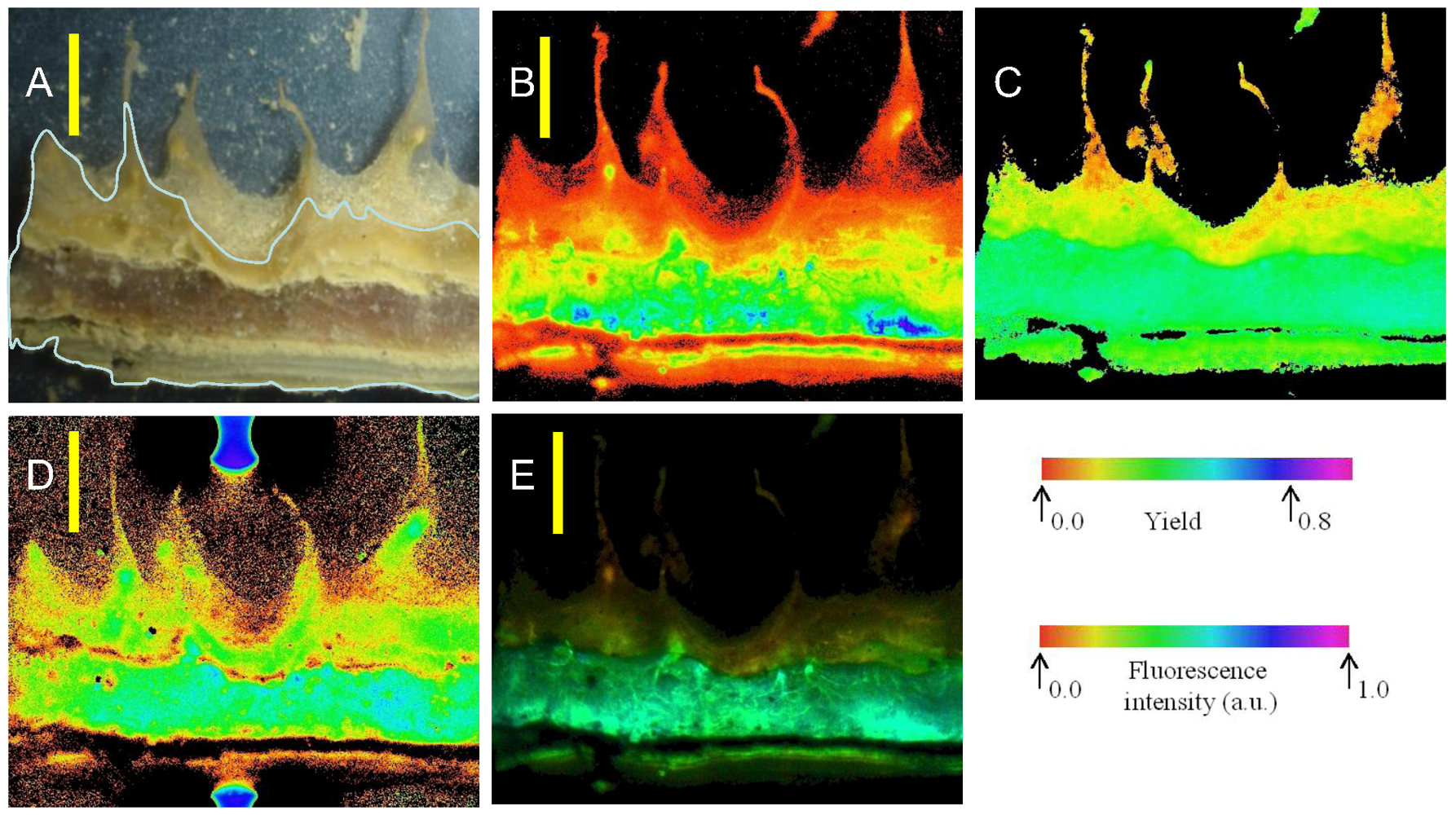
Imaging of variable chlorophyll-a fluorescence provided insights into both the location of areas where pigments were most abundant and of potential electron transport capacity. Four images of a vertical section of microbial mat from 10 m are shown in
Figure 6. At the left, a photograph of the mat shows the orange, green and pink-dominated zones overlying a near-colorless underlayer; the adjacent image shows how
Fm is distributed primarily into the laminae below the surface. Note that the cut surface of the section is indicated in the left hand image, mat components behind the cut surface are further from the camera and fluorescence yield cannot be directly compared. The third image from the left indicates that the yield of PSII under experimental conditions was rather evenly distributed, with
Fv/
Fm of 0.4–0.5, with the possible exception of the orange zone, where
Fv/
Fm values of ~0.25 were observed. The right hand image combines
Fm and
Fv/
Fm, in that the intensity of the image is scaled to
Fm, while the color is scaled to
Fv/
Fm. These images together show that potentially active photosystems are found in all of the pigmented layers and, indeed, that the highest values of
Fv/
Fm occurred within the purple and green zones, rather than close to the mat surfaces. Images also show bands of high pigment concentration at intervals below the mat surface, and comparison with the photograph shows how these are associated with lamina boundaries. Absorption images support the view that chlorophyll-a is distributed well down into the deeper laminae of the mats, indicated by absorption of red light, but not near infra-red light (
Figure S3).
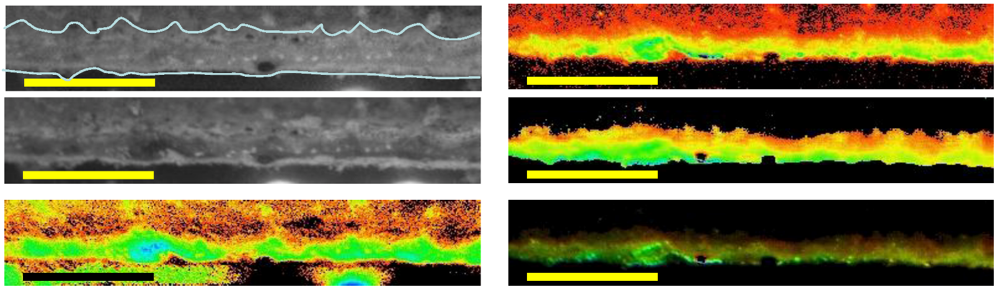
Images of example mat sections from 6 m (
Figure S4) show similar organization to
Figure 6, though with a markedly thinner cross section. The 6 m images are slightly oblique, but when the cut surface is examined once again the distribution of fluorescence and absorptions indicate a concentration of pigments and PSII yield in the sub-surface laminae. Finally, images from 14 m depth samples show further evolution of this arrangement in a thicker mat section (
Figure S5). Images at the left show the arrangement of pigmented zones and the localization of maximal chlorophyll-a absorption in the deeper laminae, while
Fm and
Fv/
Fm images indicate the maximum potential yield occurs in these deeper parts of the mats and that fluorescence is maximal at the lamina boundaries.
Figure 6.
Distribution of fluorescence intensity (minimum fluorescence yield [Fo]—second image from left) and maximum yield of photosystem II (the maximum quantum yield of PSII [Fv/Fm]—third image) in part of a vertical section of a mat from 10 m depth. At far right is a maximum fluorescence yield (Fm)-weighted yield image, where the false color represents the yield and the intensity represents Fm. Color scales below fluorescence images represent imaging pulse amplitude modulated (PAM)-derived false-color values. At far left is a photographic image of the mat section at the same scale on which the annotated line shows the outline of the cut surface. Scale bars are 5 mm.
Figure 6.
Distribution of fluorescence intensity (minimum fluorescence yield [Fo]—second image from left) and maximum yield of photosystem II (the maximum quantum yield of PSII [Fv/Fm]—third image) in part of a vertical section of a mat from 10 m depth. At far right is a maximum fluorescence yield (Fm)-weighted yield image, where the false color represents the yield and the intensity represents Fm. Color scales below fluorescence images represent imaging pulse amplitude modulated (PAM)-derived false-color values. At far left is a photographic image of the mat section at the same scale on which the annotated line shows the outline of the cut surface. Scale bars are 5 mm.
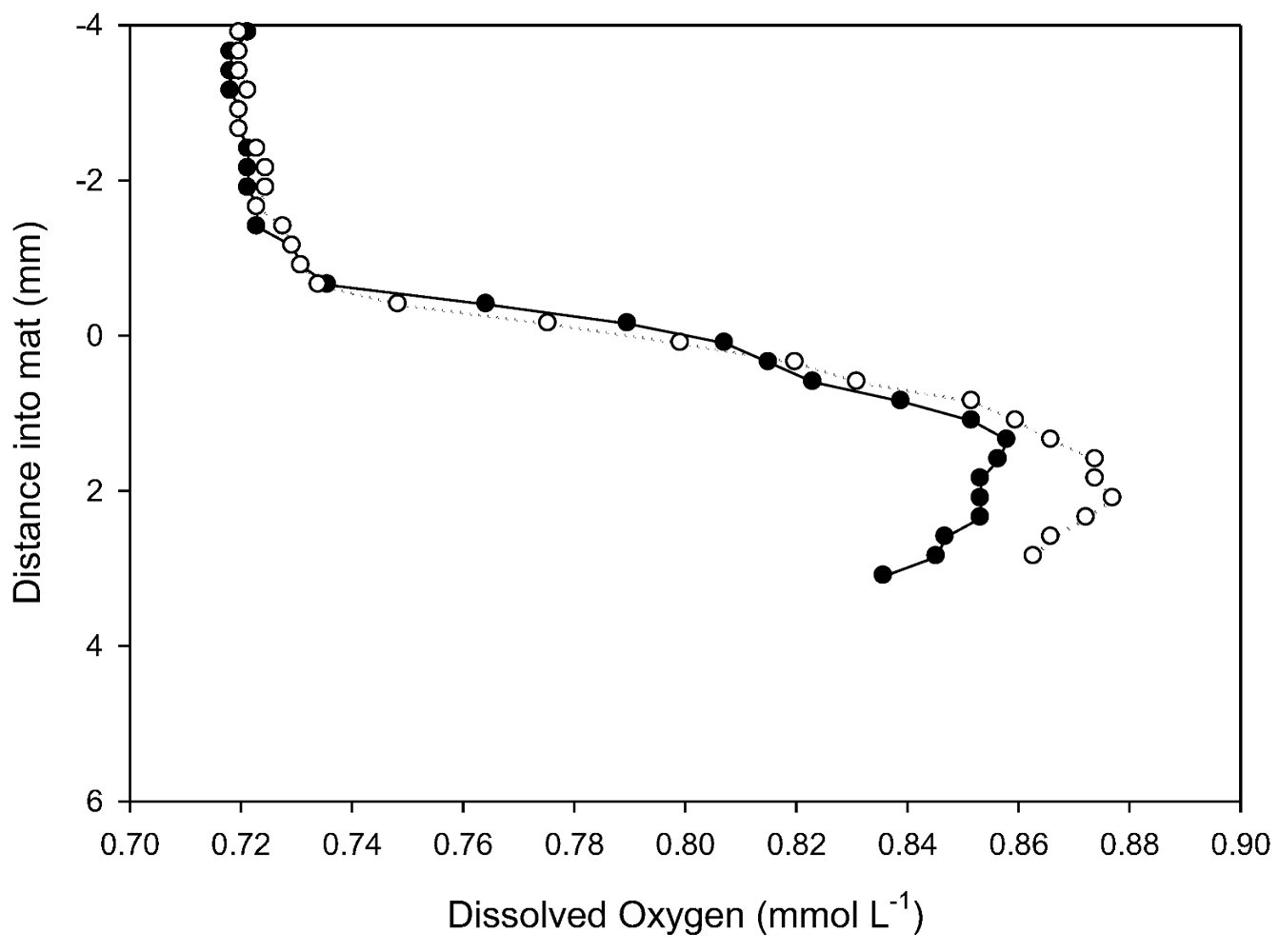
3.6. Oxygen Microprofiles
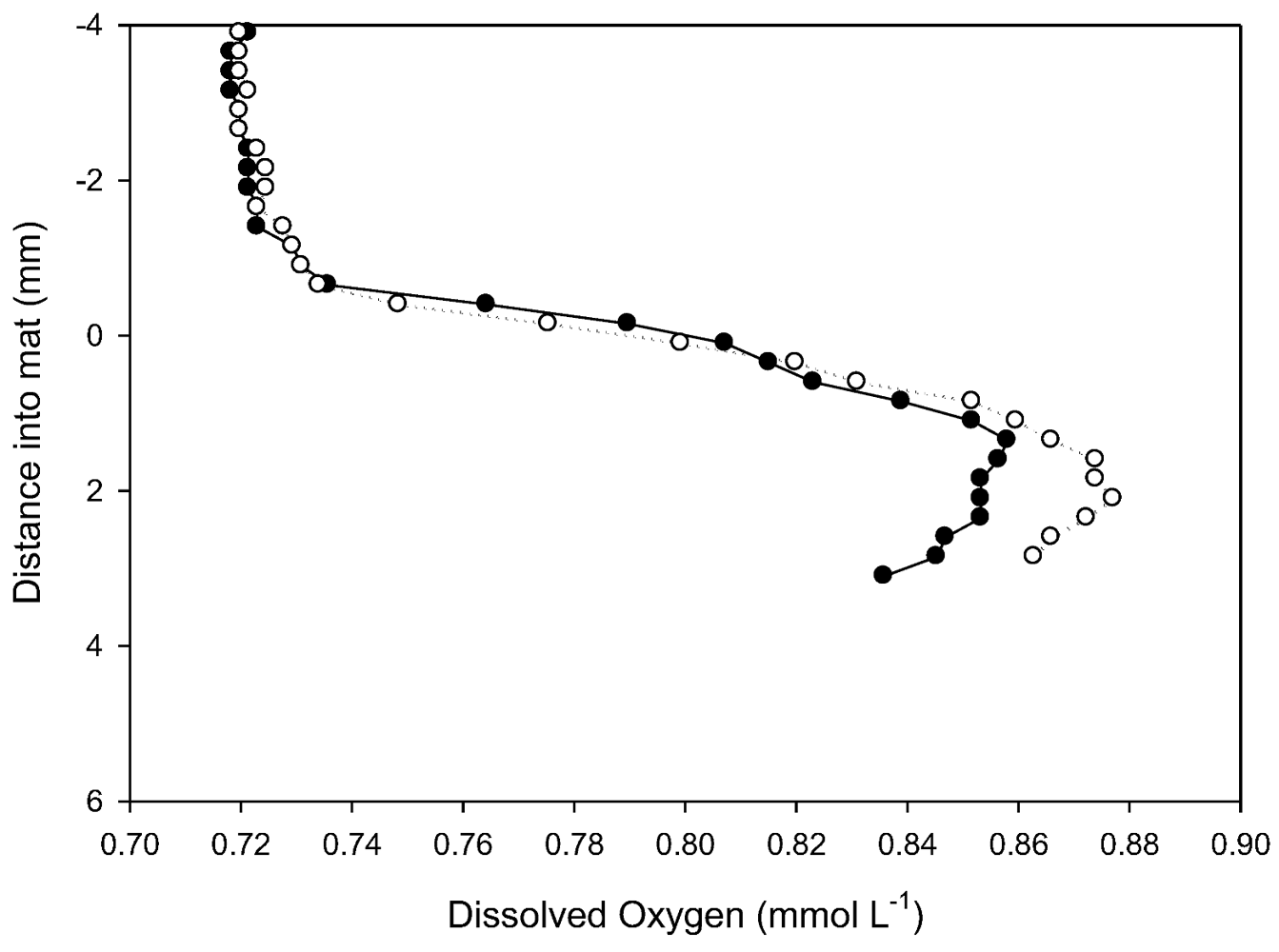
In situ dissolved oxygen microprofiles in prostrate mat at 18 m (
Figure 7) were made at ambient temperature (4.2 °C, 602 µS cm
−1 conductivity, pH 8.52 and a photon flux of 80 µmol m
−2 s
−1). As is common in Antarctic lakes, dissolved oxygen concentration in the water column exceeded atmospheric saturation [
28]). Profiles show a rapid rise in dissolved O
2 concentrations through the diffusive boundary layer and into the top part of the mat and a convex profile indicative of O
2 production via photosynthesis to at least 4 mm. This is consistent with fluorescence analyses that show photosynthetic potential extending into the older laminae. Penetration into the mat was insufficient to document the activity of the full thickness of the microbial mat. Estimation of areal oxygen evolution rate from the concentration gradient in the DBL of
Figure 7 yields a rate of 0.3 to 0.4 µg oxygen cm
−2 h
−1. This is of a similar magnitude to that reported earlier for this lake of −1 to +3 µg oxygen cm
−2 h
−1 from darkness to light saturation using
in vitro incubation methods [
14].
Figure 7.
Two in situ profiles of dissolved oxygen within a prostrate mat at 18 m in Lake Vanda, obtained with microelectrodes. The vertical axis, in mm, locates the mat surface at 0 mm, with the water column as negative and the mat as positive displacements.
Figure 7.
Two in situ profiles of dissolved oxygen within a prostrate mat at 18 m in Lake Vanda, obtained with microelectrodes. The vertical axis, in mm, locates the mat surface at 0 mm, with the water column as negative and the mat as positive displacements.
4. Discussion
Existing models of the growth and lamination of prostrate microbial mats in the MDV lakes are based on the accumulation of a new organic-rich layer during summer photosynthetic growth and the laying down of a sediment-rich layer during winter [
5,
6]. The increase in the number of laminae at specific locations in Lake Vanda between 1998 and 2010 was sufficiently close to 12 to support the view of these laminae as annual alternations between sediment deposition and mat growth. Our data suggest that mats tend to contain 2–3 laminae less than the number of years they have been within the main body of the lake, suggesting that annual banding does not occur within the seasonally frozen moat and becomes evident only 2–3 years after mats have emerged under the ice.
Vertical lamination is a characteristic of accreting microbial mats in clastic sedimentary environments. It is typically viewed as a consequence of filamentous cyanobacteria and diatoms moving through and overgrowing episodic sedimentary events and binding sediments in extracellular materials [
2,
30]. Lamination on an annual timescale, as seen in MDV lakes, is consistent with the seasonal rather than diel photoperiod at high latitude and the short pulse of flowing water and, hence, sediment influx during January [
5,
24]. Variability of the amount of sediment within laminae most likely reflects variations in summer sediment load, and unusually thick deposits are likely related to high sediment influx. In 2001–2002, a MDV-wide high flow event was recorded in response to unusually warm summer temperature [
31], and we interpret the thick sediment layer seen across depths ~9 laminae down to be related to delivery of a large dose of sediment through this climate-driven event. It provides further mechanistic support for near annual nature of lamination, but caution is required in that sediment laminae can be expected to be less distinct during years with low sediment influx and are not completely reliable as annual phenomena. However, our observations show how changing climatic conditions can have complex effects on microbial communities, not just through rising lake levels and associated effects on irradiance, but also through changes to sediment flux.
Gradual accumulation of microbial mat biomass through this process is evident by the increase in thickness, areal C, N and chlorophyll-a content with depth and years since emergence below the ice. Where it is possible to date mat initiation, predictable rates of increase in thickness, chlorophyll-a and pinnacle size were observed, though it is evident that mat dynamics during this early phase of growth, which may last for 3–4 decades, changes once mats reach maturity. At maturity, ongoing growth at the surface is likely to be increasingly offset by decomposition and compression of deeper laminae, though gradual accumulation of organic material and thickness appears to continue. Pigment per unit area can also be expected to saturate, as the effects of increasing biomass on self-shading will be compounded by the effects of reduced irradiance. Even if pigment biomass accumulated to light-limited status, it will still be expected to decline with depth within the mature mat community due to declining irradiance [
24]. Indeed, we would expect that the ultimate, light-limited chlorophyll-a biomass that could be achieved will be declining over time in the deep, mature mats, as irradiance is reduced by increasing water depth. Thus, the response of deep communities to lake level rise can be expected to be opposite to those growing in new habitat, with slower growth and declining photosynthetically active biomass, though our data are insufficient to quantify any such change.
Microbial mats are clearly a sink for C, N and P in Lake Vanda at all depths. The rates of accumulation of C in sediments at depth contours of known age, in the shallow part of the lake where mats are still accruing towards saturation biomass, can be estimated from
Figure 5 as approximately 390 mg m
−2 y
−1. This equates to an average of 104 µg oxygen cm
−2 y
−1 (using a photosynthetic quotient of 1:1) and extrapolating the very limited data on areal oxygen evolution presented here (0.3 µg oxygen cm
−2 h
−1 would generate 104 µg oxygen cm
−2 in 14 days) suggests that such a rate of accrual is quite feasible given that sufficient irradiance is likely to be present for at least 100 days of photosynthesis. Using C:N and N:P mass ratios of approximately 14 and 90:1 measured here, this carbon accrual is equivalent to approximately 24 and 1.4 µg m
−2 y
−1 of N and P. Jones-Lee an Lee [
32] provided data from which an annual external load of P to Lake Vanda can be estimated as 2.3 µg m
−2 y
−1, a value similar to that calculated for mat sequestration and suggesting that mats may be an important sink for nutrients entering the upper part of the water column from outside of the lake.
While annual accretion of thin laminae of microbial mat at the surface is supported by our data, a growth model solely based on accrual of a new layer is only partially supported. Increases in the thickness of subsurface laminae relative to the surface, shifts in pigment contents and species composition and ongoing photosynthetic capacity with higher maximum yield of PSII than surface laminae suggest that the buried laminae remain significantly active for many years and contribute to mat thickening and pigment content
In Lake Vanda, we found a shift in
Fv/
Fm, as well as pigmentation at the orange-pink transition. Shifts in pigment composition can result in alterations in apparent yields of photosystems to a given excitation wavelength [
33], particularly in cyanobacteria, where phycobilins can be associated to PSII rather than PSI [
34]. It is possible that the shifts in
Fv/
Fm at the pink-brown interface are thus methodological and may also involve the relative ability of different pigment complements to respond to the blue measuring light and to connect to the PSII centers.
Internal metabolic zonation is, however, a characteristic of microbial mats. Pearl and Pinckney [
35] summarized how the consortium of microorganisms within a mat typically creates and exploits steep gradients of environmental conditions over mm length scales. Zonation of metabolism within typical temperate microbial mats sees oxygenic photosynthesis dominating surface zones on a diurnal basis, with anoxygenic photosynthesis and sulfide metabolism embedded in steep gradients of light, oxygen, pH, redox potential and sulfide concentrations [
1,
36], as also observed from mats in shallow Antarctic melt ponds [
37,
38]. However, such acute metabolic zonation does not appear to be common in the upper waters of MDV lakes during summer, where steep gradients of dissolved oxygen are not apparent [
28]. The reasons for this are not clear, though Vopel and Hawes [
28] did calculate low rates of oxygen consumption within mats and may also relate to perennially high ambient oxygen concentration in overlying waters that prevent hypoxia within surface sediments.
The shift towards dominance of pigment profiles by phycobilins over carotenoids with depth into mats and the corresponding change in appearance from orange (carotenoids) to green (phycocyanin) and pink (phycoerythrin) is consistent with acclimation to changing spectral characteristics inside the microbial mats. Irradiance becomes increasingly green and orange inside Lake Vanda mats [
14], and such depth-in-mat related spectral adaptation is shown in other types of microbial mat [
1,
39], albeit reflecting a shift to sulfur bacteria rather than phycobilin-rich cyanobacteria. Internal zonation boundaries did not mark a shift in dominant cyanobacterial phototrophs, based on morphotype distribution or in photosynthetic process, since photosystem II was clearly active here. Indeed, profiles of dissolved oxygen in Lake Vanda mats show concentrations well above 100% saturation to extend through the orange layer and into the pink, and no bacteriochlorophylls were detected by HPLC pigment analysis. Chloroflexi 16S rRNA gene sequences have been detected in the Lake Vanda mats (Jungblut, personal observations). Ongoing research is using molecular techniques to determine whether taxonomic shifts undetectable by microscopy are occurring with these zones.
Lake Vanda is, however, highly oligotrophic, and zonation within mats may reflect differential supply of internally recycled nutrients. Quesada
et al. [
7] reported elevated nutrient concentrations within microbial mats relative to bulk lake water in nearby Lake Hoare, and other polar microbial mats from the Arctic can be internally nutrient replete [
40], assisted by potential microbial recycling and scavenging processes [
41]. The paradigm of microbial mats acquiring, then efficiently retaining and recycling, nutrients has been advanced to explain this disparity. A potential “crossed resource gradient” therefore exists in Lake Vanda mats, with irradiance flux decreasing and nutrient concentration increasing inside the mat. The role of nutrients in controlling benthic microbial growth in Lake Vanda has not been addressed, though the plankton in the upper waters are considered strongly P-limited [
16,
18]. Elemental ratios of N:P reported here for microbial mats are high relative to the median of freshwater habitats, while C:N ratio is only slightly above median [
42], supporting the view that P may limit mat growth. P-limited growth may constrain the rate of biomass accrual in the recently flooded parts of the lake, and the availability of recycled nutrients within mats may support internal metabolism.
The picture that emerges from our data, consistent across depths and between sampling occasions, is that benthic biomass begins to accrue as discreet annual laminae within a few years of the emergence of benthic substrate under ice. New laminae are derived by the upwards growth of intertwined bundles of Leptolyngbya trichomes, rich in carotenoids (particularly myxoxanthophyll), giving rise to a thin new orange-brown surface layer, which, during the late summer and winter, receives a variable dose of fine sedimentary material. These tufts occasionally form pinnacles, initially a mm or less tall, which slowly extend over time. Slow, undisturbed accumulation of biomass, layer by layer, perhaps limited by rate of nutrient accrual, provides a simple explanation of the patterns of benthic biomass by depth (and hence by time) in the upper waters of Lake Vanda. It appears that this process may be a quantitatively significant sink for biologically active elements in that lake.
The pattern of accumulation in the part of the lake where inundation history is known is consistent with that history. The multi-variable discontinuity at 14–16 m in the lake (in 2010) relates to a zone that has been submerged by a surge in lake level rise since 1970–1980. The mat characteristics at this discontinuity, thickness of 25 mm accumulating at 0.3 mm y
−1 and pinnacle height of 10 mm accumulating at 0.2 mm y
−1, are consistent with an age of ~50 years. At depths below the discontinuity, mats are substantially thicker with much larger pinnacles and a markedly higher organic carbon and carbonate content. Estimating the age of these mats requires extrapolation of relationships such as those in
Figure 4. Thickness increase rates of 0.3 mm y
−1 and pinnacle net extension of 0.2 mm y
−1 suggest ages for 22 m mats of 60–100 years, though the questionable assumption that rates of change remain constant for such lengths of time renders estimates imprecise.
Rising levels in the MDV lakes are best related to summer degree days above freezing [
8]. In recent decades, the region appears to be undergoing a period when summer climate is sufficiently warm for lake level to have been steadily rising. Microbial mats confirm that this increase has been ongoing since the mid 20
th century and suggest that it may have begun perhaps 100 years ago. At the current rate of increase in lake level, microbial mats in Lake Vanda are able to keep pace with level rise in terms of initiating colonization. However, their slow rate of development means that they will take decades to reach their climax biomass and complexity. What is not addressed here is the fate of microbial mats in deeper parts of the lake where level change is affecting irradiance regime. There, a decline in microbial production can be expected as irradiance becomes increasingly limiting to sustainable biomass. Coring of the microbial mat at the junction between the two convection cells and below will allow the history of mat development to be better constrained, and sampling of mats from significantly below the chemocline will determine qualitative differences between the responses of mat communities there and those in the recently evolved upper cell.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}