Using Natural Gradients to Infer a Potential Response to Climate Change: An Example on the Reproductive Performance of Dactylis Glomerata L.

Abstract
:1. Introduction
2. Results and Discussion
2.1. Results
2.1.1. General Results
2.1.2. Relationships between Resource Investment in Reproduction and Climate Variables

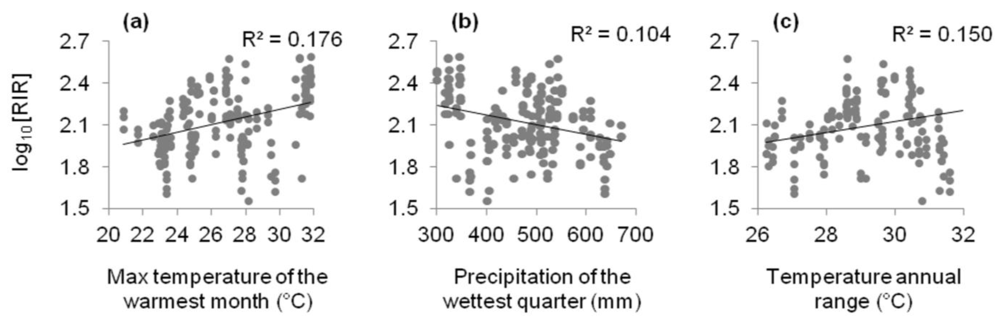{kind=link}
{kind=link}
| Variable | Intercept | β | F | P | R2 |
|---|---|---|---|---|---|
| Environmental energy | |||||
| Max temperature of warmest month | 2.971 | 0.072 | 12.413 | <0.001 | 0.176 |
| Mean temperature of wettest quarter | 3.377 | 0.091 | 11.757 | 0.001 | 0.167 |
| Mean temperature of warmest quarter | 3.377 | 0.091 | 11.757 | 0.001 | 0.167 |
| Annual mean temperature | 3.817 | 0.111 | 12.190 | <0.001 | 0.146 |
| Mean temperature of driest quarter | 4.396 | 0.109 | 11.518 | 0.001 | 0.146 |
| Mean temperature of coldest quarter | 4.396 | 0.109 | 11.518 | 0.001 | 0.146 |
| Min temperature of coldest month | 5.276 | 0.119 | 8.685 | 0.005 | 0.077 |
| Mean diurnal range | 3.819 | 0.096 | 1.273 | 0.264 | 0.020 |
| Water availability | |||||
| Precipitation of wettest quarter | 6.150 | −0.003 | 9.412 | 0.003 | 0.104 |
| Precipitation of wettest month | 6.019 | −0.007 | 6.379 | 0.014 | 0.102 |
| Annual precipitation | 6.252 | −0.001 | 8.642 | 0.005 | 0.093 |
| Precipitation of warmest quarter | 5.154 | −0.001 | 1.012 | 0.319 | 0.018 |
| Precipitation of driest quarter | 5.024 | −0.001 | 0.277 | 0.601 | 0.004 |
| Precipitation of coldest quarter | 5.024 | −0.001 | 0.277 | 0.601 | 0.004 |
| Precipitation of driest month | 4.776 | −0.001 | 0.080 | 0.779 | 0.002 |
| Climatic seasonality | |||||
| Temperature annual range | 1.980 | 0.097 | 9.470 | 0.003 | 0.150 |
| Temperature seasonality | 2.036 | 0.004 | 8.278 | 0.006 | 0.137 |
| Isothermality | 6.519 | −0.044 | 2.891 | 0.095 | 0.052 |
| Precipitation seasonality | 4.430 | 0.820 | 1.181 | 0.282 | 0.021 |

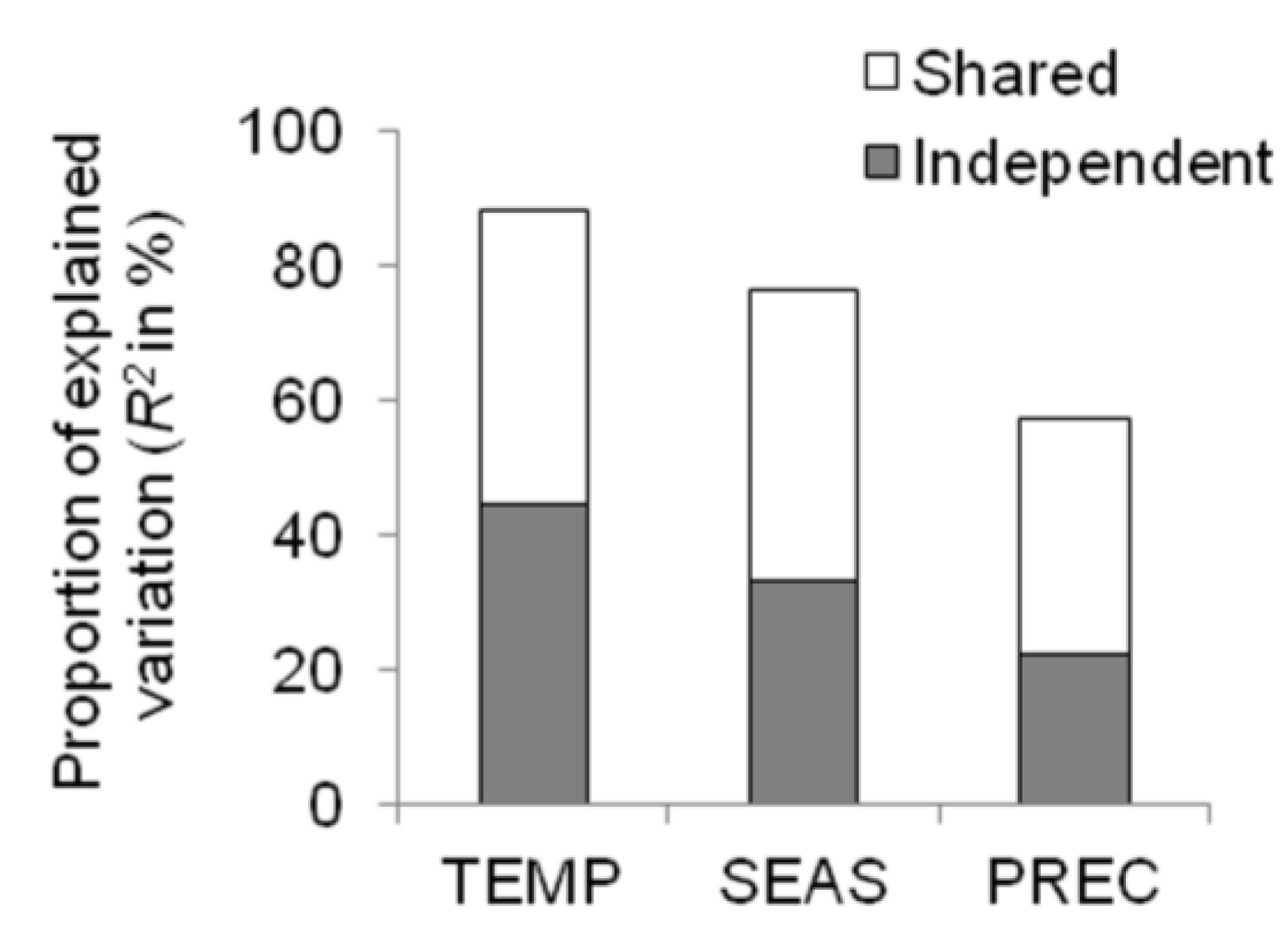
2.1.3. Multi-Model Inference and Hierarchical Partitioning
| Variable importance | Coefficients | 1st mod. | 2nd | 3rd | 4th |
|---|---|---|---|---|---|
| - | R2 | 0.18 | 0.15 | 0.14 | 0.13 |
| - | ΔAICc | 0 | 1.45 | 1.71 | 1.99 |
| Σ wi | Model wi | 0.52 | 0.25 | 0.22 | 0.14 |
| - | Intercept | 3.185 | 2.148 | 2.635 | 2.982 |
| 0.81 | TEMP | 0.060 | - | 0.045 | 0.070 |
| 0.23 | PREC | - | - | - | −0.001 |
| 0.46 | SEAS | - | 0.092 | 0.035 |

2.2. Discussion
3. Experimental Section
3.1. Case Study and Data Collection
3.2. Climate Model
3.3. Data Analysis
4. Conclusions
Acknowledgments
Supplementary Files
References
- Graae, B.J.; de Frenne, P.; Kolb, A.; Brunet, J.; Chabrerie, O.; Verheyen, K.; Pepin, N.; Heinken, T.; Zobel, M.; Shevtsova, A.; et al. On the use of weather data in ecological studies along altitudinal and latitudinal gradients. Oikos 2012, 121, 3–19. [Google Scholar] [CrossRef]
- Wilson, S.D.; Nilsson, C. Arctic alpine vegetation change over 20 years. Glob. Change Biol. 2009, 15, 1676–1684. [Google Scholar] [CrossRef]
- De Frenne, P.; Kolb, A.; Verheyen, K.; Brunet, J.; Chabrerie, O.; Decocq, G.; Diekmann, M.; Eriksson, O.; Heinken, T.; Hermy, M.; et al. Unravelling the effects of temperature, latitude and local environment on the reproduction of forest herbs. Glob. Ecol. Biogeogr. 2009, 18, 641–651. [Google Scholar] [CrossRef]
- Körner, C. Alpine Plant Life, 2nd ed; Springer: Berlin, Germany, 2003. [Google Scholar]
- Fukami, T.; Wardle, D.A. Long-term ecological dynamics: Reciprocal insights from natural and anthropogenic gradients. Proc. Roy. Soc. Lond. B Biol. Sci. 2005, 272, 2105–2115. [Google Scholar] [CrossRef]
- Körner, C. The use of 'altitude' in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Sherry, R.A.; Zhou, X.H.; Gu, S.L.; Arnone, J.A.; Schimel, D.S.; Verburg, P.S.; Wallace, L.L.; Luo, Y.Q. Divergence of reproductive phenology under climate warming. Proc. Natl. Acad. Sci. U. S. A. 2007, 104, 198–202. [Google Scholar]
- Walker, M.D.; Wahren, C.H.; Hollister, R.D.; Henry, G.H.R.; Ahlquist, L.E.; Alatalo, J.M.; Bret-Harte, M.S.; Calef, M.P.; Callaghan, T.V.; Carroll, A.B.; et al. Plant community responses to experimental warming across the tundra biome. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 1342–1346. [Google Scholar]
- Dormann, C.F.; Woodin, S.J. Climate change in the Arctic: Using plant functional types in a meta-analysis of field experiments. Funct. Ecol. 2002, 16, 4–17. [Google Scholar] [CrossRef]
- Rees, M. Trade-offs among dispersal strategies in British plants. Nature 1993, 366, 150–152. [Google Scholar]
- Rees, M. Community structure in sand dune annuals: Is seed weight a key quantity? J. Ecol. 1995, 83, 857–863. [Google Scholar] [CrossRef]
- Guo, Q.F.; Brown, J.H.; Valone, T.J.; Kachman, S.D. Constraints of seed size on plant distribution and abundance. Ecology 2000, 81, 2149–2155. [Google Scholar] [CrossRef]
- Leishman, M.R. Does the seed size/number trade-off model determine plant community structure? An assessment of the model mechanisms and their generality. Oikos 2001, 93, 294–302. [Google Scholar]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Global warming and sexual plant reproduction. Trends Plant Sci. 2009, 14, 30–36. [Google Scholar] [CrossRef]
- De Frenne, P.; Brunet, J.; Shevtsova, A.; Kolb, A.; Graae, B.J.; Chabrerie, O.; Cousins, S.A.; Decocq, G.; de Schrijver, A.; Diekmann, M.; et al. Temperature effects on forest herbs assessed by warming and transplant experiments along a latitudinal gradient. Glob. Change Biol. 2011, 17, 3240–3253. [Google Scholar] [CrossRef]
- De Frenne, P.; Graae, B.J.; Kolb, A.; Brunet, J.; Chabrerie, O.; Cousins, S.A.O.; Decocq, G.; Dhondt, R.; Diekmann, M.; Eriksson, O.; et al. Significant effects of temperature on the reproductive output of the forest herb Anemone nemorosa L. Forest Ecol. Manag. 2010, 259, 809–817. [Google Scholar] [CrossRef]
- Dainese, M. Impact of land use intensity and temperature on the reproductive performance of Dactylis glomerata populations in the southeastern Alps. Plant Ecol. 2011, 212, 651–661. [Google Scholar] [CrossRef]
- Hovenden, M.J.; Wills, K.E.; Chaplin, R.E.; Vander Schoor, J.K.; Williams, A.L.; Osanai, Y.U.I.; Newton, P.C.D. Warming and elevated CO2 affect the relationship between seed mass, germinability and seedling growth in Austrodanthonia caespitosa, a dominant Australian grass. Glob. Change Biol. 2008, 14, 1633–1641. [Google Scholar] [CrossRef]
- Worldclim. Available online: http://www.worldclim.org/ (acceseed on 1 July 2012).
- Borrill, M. Temperate grasses. In Evolution of Crop Plants; Simmonds, N., Ed.; Longman: London, UK, 1976; pp. 137–142. [Google Scholar]
- Beddows, A. Dactylis glomerata L. J. Ecol. 1959, 47, 223–239. [Google Scholar] [CrossRef]
- Äeschimann, D.; Lauber, K.; Moser, D.M.; Theurillat, J.P. Flora Alpina; Zanichelli: Bologna, Italy, 2004. [Google Scholar]
- Moles, A.T.; Warton, D.I.; Warman, L.; Swenson, N.G.; Laffan, S.W.; Zanne, A.E.; Pitman, A.; Hemmings, F.A.; Leishman, M.R. Global patterns in plant height. J. Ecol. 2009, 97, 923–932. [Google Scholar]
- Breza, L.C.; Souza, L.; Sanders, N.J.; Classen, A.T. Within and between population variation in plant traits predicts ecosystem functions associated with a dominant plant species. Evol. Ecol. 2012, 2, 1151–1161. [Google Scholar] [CrossRef]
- HilleRisLambers, J.; Harpole, W.S.; Schnitzer, S.; Tilman, D.; Reich, P.B. CO2, nitrogen, and diversity differentially affect seed production of prairie plants. Ecology 2009, 90, 1810–1820. [Google Scholar] [CrossRef]
- Rajaniemi, T.K. Why does fertilization reduce plant species diversity? Testing three competition-based hypotheses. J. Ecol. 2002, 90, 316–324. [Google Scholar] [CrossRef]
- Benavides, R.; Montes, F.; Rubio, A.; Osoro, K. Geostatistical modelling of air temperature in a mountainous region of Northern Spain. Agr. Forest. Meteorol. 2007, 146, 173–188. [Google Scholar]
- Pinheiro, J.; Bates, D. Mixed Effects Models in S and S-Plus; Springer: New York, NY, USA, 2000. [Google Scholar]
- Pinheiro, J.; Bates, D.; DebRoy, S.; Sarkar, D. R Development Core Team, nlme: Linear and Nonlinear Mixed Effects Models. R Package Version 3.1-102; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- R Development Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.R-project.org/ (acceseed on 16 January 2012).
- Singer, J.; Willett, J. Applied Longitudinal Data Analysis: Modeling Change and Event Occurrence; Oxford University Press: Oxford, UK, 2003. [Google Scholar]
- Burnham, K.; Anderson, D. Model Selection and Multimodel Inference, A Practical Information—Theoretic Approach; Springer: Berlin, Germany, 2002. [Google Scholar]
- Barton, K. MuMIn: Multi-model inference. R package version 1.6.5. R Foundation for Statistical Computing: Vienna, Austria, 2011. Available online: http://CRAN.R-project.org/package=MuMIn (acceseed on 16 January 2012).
- Chevan, A.; Sutherland, M. Hierarchical partitioning. Am. Stat. 1991, 45, 90–96. [Google Scholar]
- Murray, K.; Conner, M. Methods to quantify variable importance: Implications for the analysis of noisy ecological data. Ecology 2009, 90, 348–355. [Google Scholar] [CrossRef]
- Walsh, C.; MacNally, R. hier.part: Hierarchical Partitioning. R Package Version 1.0-3; R Foundation for Statistical Computing: Vienna, Austria, 2008. [Google Scholar]
- Bretagnolle, F.; Thompson, J.D.; Lumaret, R. The influence of seed size variation on seed germination and seedling vigour in diploid and tetraploid dactylis glomerata L. Ann. Bot. 1995, 76, 607–615. [Google Scholar] [CrossRef]
- McKee, J.; Richards, A.J. Variation in seed production and germinability in common reed (Phragmites australis) in Britain and France with respect to climate. New Phytol. 1996, 133, 233–243. [Google Scholar] [CrossRef]
- Vera, M.L. Effects of altitude and seed size on germination and seedling survival of heathland plants in North Spain. Plant Ecol. 1997, 133, 101–106. [Google Scholar] [CrossRef]
- Kidson, R.; Westoby, M. Seed mass and seedling dimensions in relation to seedling establishment. Oecologia 2000, 125, 11–17. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Dainese, M. Using Natural Gradients to Infer a Potential Response to Climate Change: An Example on the Reproductive Performance of Dactylis Glomerata L. Biology 2012, 1, 857-868. https://doi.org/10.3390/biology1030857
Dainese M. Using Natural Gradients to Infer a Potential Response to Climate Change: An Example on the Reproductive Performance of Dactylis Glomerata L. Biology. 2012; 1(3):857-868. https://doi.org/10.3390/biology1030857
Chicago/Turabian StyleDainese, Matteo. 2012. "Using Natural Gradients to Infer a Potential Response to Climate Change: An Example on the Reproductive Performance of Dactylis Glomerata L." Biology 1, no. 3: 857-868. https://doi.org/10.3390/biology1030857