
Fungal Metabolites for the Control of Biofilm Infections



Helmholtz Centre for Infection Research, Chemical Microbiology, Inhoffenstrasse 7, 38124 Braunschweig, Germany
*
Author to whom correspondence should be addressed.
Agriculture 2016, 6(3), 37; https://doi.org/10.3390/agriculture6030037
Submission received: 24 June 2016
/
Revised: 1 August 2016
/
Accepted: 9 August 2016
/
Published: 12 August 2016
(This article belongs to the Special Issue Fungal Metabolites)
Abstract
:Many microbes attach to surfaces and produce a complex matrix of polymers surrounding their cells, forming a biofilm. In biofilms, microbes are much better protected against hostile environments, impairing the action of most antibiotics. A pressing demand exists for novel therapeutic strategies against biofilm infections, which are a grave health wise on mucosal surfaces and medical devices. From fungi, a large number of secondary metabolites with antimicrobial activity have been characterized. This review discusses natural compounds from fungi which are effective against fungal and bacterial biofilms. Some molecules are able to block the cell communication process essential for biofilm formation (known as quorum sensing), others can penetrate and kill cells within the structure. Several targets have been identified, ranging from the inhibition of quorum sensing receptors and virulence factors, to cell wall synthesizing enzymes. Only one group of these fungal metabolites has been optimized and made it to the market, but more preclinical studies are ongoing to expand the biofilm-fighting arsenal. The broad diversity of bioactive compounds from fungi, their activities against various pathogens, and the multi-target trait of some molecules are promising aspects of fungal secondary metabolites. Future screenings for biofilm-controlling compounds will contribute to several novel clinical applications.
1. Biofilm Infections Are a Therapeutic Challenge
In biofilms microbial cells are embedded in a complex matrix of macromolecules that adhere to various surfaces. While biofilms have several roles in the environment, like cleaning waste water in sewage plants, they can also be harmful, e.g., causing damage to buildings and monuments or clogging tubes and pipelines in industrial plants. In humans, they can play a protective role in the skin and mucosa, but often can represent an infectious threat. In this review we focus on infectious biofilms formed by pathogenic microorganisms, and we review approaches for their control using compounds produced by fungi. Biofilm infections are caused not only by bacteria, e.g., Pseudomonas aeruginosa, Escherichia coli, or Staphylococcus aureus, but also by fungi, e.g., Candida albicans or Aspergillus fumigatus. Biofilms can form on any implant and also on mucosal surfaces, e.g., in the cystic fibrosis lung [1], the middle ear [2], the urinary tract [3], or gastric mucosa [4]. Biofilm formation is a key factor for microbial survival in hostile environments, from where cells can disperse to colonize new habitats [5]. Infections caused by biofilms often result in severe complications and 12% to 25% of bloodstream infections are attributable to patient mortality [6].
To form a biofilm, microbes have to determine their cell density and to coordinate their population behavior. This is mediated by small molecules released into the environment and acting as autoinducers to start genetic programs [7]. If the autoinducer concentration reaches a certain threshold, the genetic program of biofilm formation starts, followed by the production of virulence factors. This intercellular communication principle is used by many microbes and called quorum-sensing (QS) [8]. To date more than 100 different autoinducers are known for bacteria, archaea, and fungi. The best studied autoinducers are N-acyl-l-homoserine lactones, only known from Gram-negative bacteria. The acyl side chains are 4–16 carbons long, usually saturated, and can be oxidized at C-3 1–3 [9] (Figure 1). Contrary to the acyl-homoserine lactones, autoinducer-2 (AI-2) is known both from Gram-positive and -negative bacteria, having the rather unusual cyclic boronic ester 4 [10]. There is also some species-specific autoinducers, e.g., the Pseudomonas quinolone signal (PQS) 5. Gram-positive bacteria use small peptides as autoinducers which are derived by post-translational processing of larger precursor peptides [11].
Quorum sensing is also known from fungi [12,13] and best studied in Candida albicans [14]. The known quorum sensing molecules from fungi are structurally very different from those found in bacteria. Tyrosol 6 [15], farnesol 7 [16,17] and 3R-hydroxy-tetradecanoic acid 8 [18], have been reported. These compounds are probably not very specific for C. albicans, since one third of all Ascomyotina strains tested could produce farnesol [19].
Most investigations on biofilm control have been done with pure strains; however, in nature, microbes seldom live alone and often form polymicrobial communities [20]. The individual species within these communities do not exist independently from each other, having complex and multifold interactions [21]. Any future drug for the control of biofilm infections has to deal with these interactions in order to avoid suppressing one pathogen while fostering the development of another one in the same biofilm community.
The eradication of biofilm infections is complicated because of the high protection of pathogens against host defenses (e.g., macrophages) and antibiotics [22]. For instance, 220-times higher antibiotic concentrations were required for killing E. coli in a biofilm than for the same strain living in planktonic form in the serum [23]. Similar high resistances have been reported for fungal biofilms [24]. Because of these complications and under the background of increasing antibiotic resistances [25] novel antibiotics and novel targets are needed to control biofilm infections [26]. Several approaches are being developed in this context. One is the search for novel, biofilm-penetrating antibiotics [27]. Another one aims to disrupt communication of pathogens by blocking quorum sensing [28,29]. Most of these efforts are directed against bacteria, but also a number of techniques have been described for the screening of compounds against pathogenic fungi biofilms [30]. The search for biofilm-modulating secondary metabolites has been undertaken by several groups. The rationale behind this approach is that all organisms are confronted with the formation of pathogenic biofilms on their bodies, meaning they are driven to develop strategies to fight pathogenic biofilms. Compounds emerging from these searches are therefore already tailored for biofilm-control, and can serve as lead compounds for drug development. Bacteria and plants seem to be currently the main sources for such bioactive compounds; however fungi, mainly those of marine origin, have also been included in these screenings [31].
In this review we discuss the control of bacterial and fungal biofilm infections by novel antibiotics and biofilm-modulating compounds of fungal origin. The advantages and short-comings of the different classes of compounds are evaluated and an outlook for future developments is given.
2. Fungal Metabolites Reported to Modulate Biofilms of Pathogens
In the attempts to find novel natural products for the control of biofilm infections, fungal extracts have been the subject of several screenings. One approach is to look for sporocarps (fruiting bodies) of fungi which are exposed to a wet climate, favorable conditions for biofilm formation. In a small study of sporocarps from Basidiomycotina it was found that ectomycorhizal fungi had a higher diversity of bacterial biofilms on their fruiting bodies than saprophytic ones. Consequently, saprophytic Basidiomycotina had a higher percentage of biofilm-inhibiting extracts than their ectomycorrhizal cousins [32]. For obtaining the required novel isolates of fungi, sometimes unusual methods have been applied, for instance crowdsourcing, where samples from citizen scientists were included in the screening procedure [33]. Often these extracts showed activity, however sometimes the investigations ended in the screening, and no results on the nature of the bioactive compounds or their modes of action were reported. This leaves us only with the knowledge that there is some activity, but not what causes it. Many of these studies were directed against specific pathogenic biofilms, e.g., Lentinula edodes extract against the bacterial human pathogens Actinomyces naeslundii, Prevotella intermedia, Neisseria subflava, and Streptococcus sanguinis [34,35], sometimes using highly elaborated and specific methods [36]. Also lichens, the association between fungi and algae, have been investigated over decades for their spectrum of bioactive secondary metabolites; however, only relatively few reports appeared on biofilm-modulating compounds from this group of organisms [37].
2.1. Biofilm-Modulating Terpenes from Fungi
The finding that high concentrations of farnesol 7, a quorum sensing compound in C. albicans, inhibits filamentation led to the application of this sesquiterpene for preventing biofilm formation by pathogenic C. albicans. When added at an early stage of biofilm formation, 300 μM farnesol could completely prevent the formation of a fungal biofilm [38]. Similar to the observation for many quorum sensing quenchers active against bacteria, the effect was less pronounced for mature biofilms. To find out whether the effect of farnesol is specific for C. albicans, Weber et al. investigated the production of eight different Candida species under different culture conditions. They found that only C. albicans and C. dubliniensis produced farnesol and from these two species, C. albicans produced more farnesol and stronger biofilms. For C. kefyr, C. glabrata, C. krusei, C. parapsilosis, C. tropicalis, and C. guilliermondii only low concentrations of farnesol were found in the media. The amount of farnesol production was not only species- but also strain-dependent [39].
When analyzing biofilm formation of a Pneumocystis species, Cushion et al. found that the ability of farnesol to prevent biofilm formation is not limited to Candida species. Species of the fungal genus Pneumocystis can cause severe pneumonia in immune-compromised patients, but lack of experimental models still prevents a better understanding of these pathogens. The addition of farnesol to cultures of P. carinii from rat or P. murina from mouse could completely prevent the formation of biofilms, which is a probable virulence mechanism in the lung [40]. Farnesol has inhibiting effects not only on fungal but also on bacterial biofilms. Here it seems that the results vary a lot between the different species tested. When methicillin-resistant Staphylococcus aureus biofilms were treated with 150 μM farnesol the resistance against methicillin could be reversed. More importantly, the sensitivity against several other antibiotics, e.g., gentamycin, could be enhanced 100-fold for established biofilms [41]. In the closely related bacteria S. epidermidis, 200 μM farnesol were required to have an effect both on planctonic and biofilm-embedded cells, a result which was comparable to that of vancomycin [42]. Not surprisingly, a reduction in biofilm volume was also observed [43]. Such a reduction in biofilm volume was also found for Streptococcus mutans biofilms; however, here farnesol caused dispersion of the biofilm rather than significant killing of the cells [44].
Biofilms of C. albicans often develop resistance against the antifungal fluconazole. When these resistant biofilms were treated with farnesol at an early stage of biofilm development, the resistance could be reduced by more than three orders. Treatment at a later stage of biofilm development had a somewhat weaker but still very significant effect [45]. A similar observation was made also for the yeast C. dubliniensis [46], and it is noteworthy that these are the two farnesol-producing Candida species.
Downregulation of genes involved in hyphal formation, GTPase activation, mitosis and DNA replication in C. albicans when treated with 40 μM farnesol 7 shed some light on the mode of action of this compound. Interestingly, this could be reversed by the addition of 1-oleoyl-2-acetyl-sn-glycerol, a diacylglycerol analogue. In mammalian cells this diacylglycerol is an activator of protein kinase C; however, in C. albicans a PKC1 deletion mutant still showed the same response indicating that protein kinase C is not the target of 1-oleoyl-2-acetyl-sn-glycerol [47]. While farnesol showed no effect at all on the Gram-negative bacteria Burkholderia multivorans and B. cenocepacia [48] it affected the virulence of another Gram-negative pathogen, Pseudomonas aeruginosa. Here treatment with farnesol led to a decreased production of the Pseudomonas quinolone signal (PQS), and the virulence factor pyocyanin, which is controlled by PQS. Transcriptome analysis revealed that the transcription of the genes involved in PQS-synthesis was reduced [49], adding another mode of action of farnesol in biofilms. The authors also tested related compounds and found that farnesyl acetate 9 and geranyllinalool 10 were active too, but not the saturated and unbranched alcohol n-dodecanol 11 (Figure 2). Farnesol has also been shown to induce apoptosis in C. albicans, causing up-regulation of the caspase gene MCA1 and the intracellular presence of caspases [50]. Apoptosis is triggered by farnesol not only in C. albicans, but also in Aspergillus nidulans [51] and Saccharomyces cerevisiae, where reactive oxygen species participate in programmed cell death [52]. Finally, apoptosis caused by farnesol was also reported in human carcinoma cell lines [53].
Besides farnesol only a few other fungal terpenes have been reported possessing biofilm-modulating activities. One of the simplest ones, mevalonolactone 12, is produced by an unidentified marine fungus isolated from the sponge Mycale magnirhaphidifera, and was found to prevent biofilm formation by S. epidermidis [54]. From a culture of a marine-derived strain of Emericella variecolor, the sesterterpenes ophiobolin K 13, 6-epi-ophiobolin K 14, and 6-epi-ophiobolin G 15 were isolated. All three ophiobolins inhibited biofilm formation of Mycobacterium smegmatis with biofilm inhibitory concentrations (BICs) of 4.1–65 μM, showing no antimicrobial activity at these concentrations. The most active compound was ophiobolin K, which was also active against M. bovis BCG [55]. The fungus Neosatorya fischeri KUTC 6344 was isolated from soil and shown to produce several secondary metabolites, including sartorypyrone A 16 and aszonapyrone A 17 and B 18 [56]. When applied at minimal inhibitory concentration (MIC)-levels, aszonapyrone A or sartorypyrone A almost completely prevented biofilm formation by S. aureus, Bacillus subtilis or Enterococcus faecalis, but were much less active in dispersing mature biofilms. Aszonapyrone B 18, only lacking the acetoxy group of aszonapyrone A 17, was inactive. Similar to most antibiotics, these compounds cause stress to planctonic cells since at subtoxic concentrations they were not only unable to prevent biofilm formation but, on the contrary, fostered it [57]. An endophytic Guignardia species isolated from the plant Euphorbia sieboldiana produced several meroterpenes when grown as solid culture on rice grains. Two of these terpenes, guignardone N 19 and guignardic acid 20, were synergistic with fluconazole in biofilm inhibition and killing of drug-resistant C. albicans, lowering the MIC from >1000 μg·mL−1 for fluconazole alone to 63 μg·mL−1 for the combination fluconazole–meroterpene [58].
2.2. Fungal Metabolites of Polyketide Origin for Biofilm-Control
For more than a century lichens have been used against bacterial infections. One of the best known compounds in this respect is usnic acid 21, produced by the fungal part in several lichens (Figure 3) [59]. It is commercially available and has been studied for its several bioactivities and its toxicity [60]. Usnic acid acts as antibiotic on group A Streptococcus [61] and C. albicans biofilms [62]. As a consequence it has been applied as surface coating against bacterial biofilm formation. Here it does not prevent biofilm formation but kills the cells when they settle at the surface [63]. Another lichen metabolite, protolichesterinic acid 22, has been reported to inhibit biofilm formation by interfering with the quorum sensing systems of P. aeruginosa and B. cenocepacia [64]. Screening a number of lichen extracts further identified thamnolic acid 23 as active against C. albicans biofilms [65].
A number of rather diverse secondary metabolites from fungi have been characterized which are formed via polyketide precursors and are able to suppress biofilm development. Patulin 24 and penicillic acid 25 from Penicillium species were found displaying quorum quenching activity, i.e., suppression of communication by quorum sensing. As already observed for most quorum-quenchers, patulin enhanced P. aeruginosa biofilm susceptibility to tobramycin. Furthermore, patulin and penicillic acid activate polymorphonuclear neutrophils and break the blockage of the oxidative burst [66]. Coprinus comatus is an edible fungus which is also used for various medical applications. From this fungus the small gamma-lactone 26 has been isolated and shown to cause dispersion of P. aeruginosa biofilms by interfering with its quorum sensing. It also reduces the production of pyocyanin and rhamnolipids, two virulence factors of P. aeruginosa [67]. From Leucostoma persoonii, isolated from red mangrove, a number of cytosporones have been isolated. One of them, cytosporone E 27, could inhibit biofilm formation of methicillin-resistant S. aureus at 39 μM, about half of its MIC, and was also active against Plasmodium flaciparum [68]. Terreic acid 28, long known from Aspergillus terreus, has been demonstrated to suppress biofilm growth of E. coli by inhibiting the bifunctional GlmU uridyltransferase, which catalyzes the acetylation and uridylation of glucosamine-1-phosphate [69]. Natural products often have several rather than only one target in the cell, and terreic acid has also been shown to covalently inhibit the enzyme MurA, which catalyzes the reaction of this substrate with phosphoenolpyruvate [70]. The soil derived fungal strain Metarhizium anisopliae produced a number of isocoumarin glycosides when growing in solid culture on rice. One of these metabolites, 29, strongly inhibited the formation of biofilms by P. aeruginosa. 6-Acetylation of the sugar seems to be essential for activity because metabolite 30, lacking the acetoxy group, did not show any activity [71]. Myriocin 31, a strong antifungal agent, was first isolated from Myriococcum albomyces [72] and later also from Isaria sinclairii and Mycelia sterilia. Myriocin acts by strong inhibition of serine palmitoyltransferase, a key enzyme in the synthesis of sphingolipids [73]. It has been demonstrated that C. albicans biofilms possess a higher ratio of sphingolipids than planktonic cells, and accordingly, treatment with myriocin completely prevented the formation of C. albicans biofilms [74], as well as biofilms of Aspergillus fumigatus clinical isolates [75]. Although these reports are rather optimistic, one should keep in mind that myriocin is toxic for eukaryotes and has a small therapeutic window.
Besides these relatively simple compounds, a number of much more complex fungal secondary metabolites have been demonstrated to possess biofilm-modulating activities. From a Penicillium sp. several closely related alkaloids were isolated and two of them, shearinines D 33 and E 34, inhibited the formation of C. albicans biofilms (Figure 4); however, they were unable to disperse already established ones. Interestingly, their parent compound, shearinine A 32, not oxidized at C-22, did not show any activity [76]. Bionectria ochroleuca produced the polyketide glycosides bionectriol B-D 35–37 and TMC-151 C-F 38–41, which could all prevent biofilm formation by C. albicans [77]. Waikialoid A 43 and waikialide A 42 from Aspergillus sp. could do the same; remarkably, waikialoid A had an IC50 of 1.4 μM, one of the lowest so far reported for the inhibition of C. albicans biofilms [78].
2.3. Amino Acids and Derivatives Controlling Biofilm Formation
Tyrosol 6 is derived from tyrosine and acts in C. albicans, together with farnesol, as a quorum sensing compound. At 200 mM, tyrosol reduced biofilm formation and metabolic activity of C. albicans, C. glabrata, S. mutans and their mixed biofilms, at a degree comparable to the effect of chlorhexidine gluconate [79].
Bioactive diketopiperazines have been reported from fungi including cyclo(l-Phe-l-Pro) 44 from Rosellinia necatrix [80] and cyclo(l-Tyr-l-Pro) 45 from Alternaria alternata [81]. These compounds interfere with the quorum sensing system of some bacteria by inhibiting the action of acyl-homoserine lactones [82], but there are contradicting reports. Cyclo(l-Tyr-l-Pro) 45 reduced colony expansion in Serratia liquefaciens and was able to inhibit the swarming behavior of a S. liquefaciens swarming motility mutant, when added together with autoinducer butyl-homoserine lactone. Diketopiperazines not only interfere with quorum sensing by competing with acyl-homoserine lactones in Gram-negative bacteria, but can also modulate biofilms of Gram-positive bacteria. Cyclo(l-Phe-l-Pro) 44 and cyclo(l-Tyr-l-Pro) 45 strongly inhibit the Ptst promoter of the accessory gene regulator (agr) quorum sensing system of S. aureus. This interaction seems to be not very specific since inhibition has been observed for all four known agr subgroups of S. aureus, despite differences in their autoinducer structures [83]. A marine Penicillium sp. isolated from the sponge Axinella corrugata produced cyclo(l-Tyr-l-Leu) 46 which inhibited biofilm formation by Staphylococcus epidermidis [84]. Cyclo(l-Leu-l-Pro) 47 from an isolate of Penicillium commune was active against S. aureus biofilms [85]. From the marine fungus Cladosporium sp., F14 cyclo-(Phe-Pro) 44 and cyclo-(Val-Pro) 48 were isolated. When tested against biofilm-forming bacteria species, both compounds were active against Loktanella hongkongenis, but only cyclo-(Phe-Pro) showed antimicrobial activity against Micrococcus luteus and Ruegeria sp. [86]. Flavipesin A 49 is an antibiotic from a marine isolate of Aspergillus flavipes which can penetrate mature biofilms of S. aureus and B. subtilis [87]. The activity of several diketopiperazines against biofilms is in line with the many activities displayed by this class of natural compounds [88] (Figure 5).
Campbell et al. synthesized several diketopiperazines and tested their library against a Vibrio fischeri reporter strain. The authors could not confirm that certain diketopiperazines can promote quorum sensing in bioreporter strains. They found that the diketopiperazines reported to be active showed no quorum sensing activity. In their screening they identified a few diketopiperazines which prevented biofilm formation of bacteria, but none of these compounds competed with acyl-homoserine lactones for their receptor. During the study a diketopiperazine macroarray, consisting of 400 different compounds, was constructed [89] which can form a basis for systematic structure-activity studies on quorum sensing control by diketopiperazines [90].
2.4. From Papulacandins and Echinocandins to Drugs against Fungal Biofilms
Infections caused by fungal pathogens are difficult to treat and fungal biofilm infections pose an even greater challenge. This dilemma caused a continuous search for novel secondary metabolites able to destroy fungal biofilms. In 1977, the first compounds from a series of double esterified disaccharides were isolated from the fungus Papularia sphaerosperma. They were named papulacandins 50–53 because of their high efficiency in inhibiting Candida albicans [91]. The mechanism of action interferes with the production of 1,3-β-glucan via inhibition of the enzyme 1,3-β-d-glucan synthase, which is required for fungal cell wall synthesis. Although the anti-biofilm activity of this class of compounds is still largely unexplored, the pivotal role played by 1,3-β-glucan in the extracellular matrix of Candida albicans biofilms [92] offers strong indication of the therapeutic potential of such inhibitors.
The discovery of the first papulacandins was followed by the findings of the closely related compounds L-687,781 54 from Dactyochaeta simplex [93,94], Mer-WF3010 54 from Phialophora cyclaminis [95,96], F-10748 A1-D1 56–59 from a Lophodermium species [97], BU-4794F 60 from a Gilmaniella species [98], and PF-1042 61 and saricandin 62 from a Fusarium species [99] (Figure 6).
The skeleton of these papulacandins bears a spiroketal moiety and its two esters are required for the activity against Candida sp. The long fatty acid at the spirocarbohydrate is essential for the bioactivity, and it has been assumed that it is required for the interaction with the fungal cell wall.
In a slightly smaller group of papulacandins the spiroketal moiety has been reduced, leading to a hydroxybenzyl-moiety. To this group of secondary metabolites belong F-10748 A2 63, B2 64, C2 65, and D2 66, also from Papularia sphaerosperma, chaetiacandin 67 from Monochaetia dimorphospora [100,101], and corynecandin 68 from Coryneum modonium [102]. Fusacandin A 69 and B 70 have also such a reduced spiroketal, but they are unique due to the attachment of an additional galactose to the galactose unit [103,104] (Figure 7). Chaetiacandin 67 and corynecandin 68 are the only papulacandins with a shortened fatty acid at the glucose unit.
Several papulacandins show high activities against several strains of Candida sp., but their pathogen spectrum is narrow and very strain-dependent. In order to improve the performance of papulacandins several synthetic derivatives were synthesized and tested [105]. Systematic variations of papulacandins revealed that the second sugar is not required for activity. This has been demonstrated by papulacandin D 50, which lacks this sugar. Hydrogenation of the two fatty acids, or even removal of the shorter fatty acids in the galactose unit have only small effects on the inhibition of glucan biosynthesis, although it completely abolishes growth inhibition. The presence of the long fatty acid is essential for activity [106]. It seems that the configuration of the fatty acid is important for penetration into the cells, thus reaching the target site to block 1,3-glucan synthase.
In 1974 the first member of another group of antifungals also active against fungal biofilms was found [107,108]. This group of cyclopetides esterified with a long-chain fatty acid is named echinocandins after its first representative echinocandin B 78, from Aspergillus echinulatus [109,110]. Later, a number of similar compounds were discovered and their biological activities were characterized [111]. These compounds include: echinocandins C 79 and D 80 from Aspergillus rugulosus [112]; aculeacin Aα 74, Aγ 75, Dα 76, and Dγ 77 from Aspergillus aculeatus [113], in addition to aculeacins B, C, D, E, F and G of which the structures have not been reported [114]; FR190293 81 and FR227673 85 from a Chalara species and from Tolypocladium parasiticum (now identified as Phialophora sp.) [115]; FR209602 82 from Coleophoma crateriformis [116]; FR901379 86 from Coleophoma empetri; mulundocandin 89 and desoxy-mulundocandin 90 from Aspergillus mulundensis [117]; pneumocandins A0-A4 91–95, B0 96, B2 97 and C0 98 from Zalerion arboricola, Glarea lozoyensis, and some Cryptosporiopsis sp. [118]; and sporiofungins A-C 99–101 from a Cryptosporiopsis species [119] (Figure 8).
Echinocandins are efficient inhibitors of 1,3- and 1,6-d-glucan synthases which are required for fungal cell wall synthesis. They are active against Candida spp., especially C. parapsilosis, C. lusitaniae, and C. guilliermondii, Aspergillus spp., and Pneumocystis carinii, but not against Zygomycetes. However as for most antibiotics, their activity is also strain-dependent, indicating differences in cell permeability. As more and more genome sequences of echinocandin-producing fungi become available, a much clearer picture of the gene organization and evolution of echinocandins emerges. Echinocandins are found in Eurotiomycetes (Aspergillus spp.) and in Leotiomycetes (Glarea, Coleophoma, Phialophora species), which diverged about 350 million years ago. Gene organizations point to aculeacin 74 as having the most ancient biosynthesis pathway, whereas FR190293 81 has the most evolved one. Since no sign of horizontal gene transfer from bacteria could be observed, an origin of echinocandin biosynthesis in the genus Aspergillus can be assumed [120,121].
In animal experiments it became evident that one complication of treatments with natural echinocandins was hemolysis. As further studies identified the amide side chain as the main factor for this problem, semi-synthetic derivatives were generated and tested [122]. To produce the cyclic peptide needed for derivatizations, the fatty acid side chain has to be selectively removed. In order to achieve this for FR901379 86, 3800 microbial isolates were tested for cyclic lipopeptide acylase activity. As a result, five strains (three Streptomyces spp. and two fungi) were identified with high levels of the enzyme [123]. Similar approaches were taken for a number of echinocandins, yielding the primary amine which was further on esterified with a number of different long chain fatty acids. A rigid linear aromatic chain and a flexible aliphatic tail were found to be favorable for these fatty acids, leading to highly active semi-synthetic echinocandins [124].
Starting from the sulfate bearing echinocandin FR901379 86 several derivatives were synthesized, all varying in the ester side chain [125,126]. Their biological activity was evaluated [127]. From these compounds, caspofungin 71, micafungin 72 [128], and anidulafungin 73 [129] finally made it to the market [130,131]. The long and challenging way from the discovery of the bioactive natural compound, over optimization of the producing strain, fermentation scale-up, optimization of the enzymatic cleavage of the side chain, formulation of the drug and, finally, to the introduction into the market has been described by Balkove et al. in the case of caspofungin 71 [132]. Comparisons of the characteristics, specificities, and pharmacokinetics of commercial echinocandins were given in a detailed review [133]. The latest addition to the semi-synthetic echinocandins is aminocandin, derived from desoxy-mulundocandin 90, which is currently being assessed in clinical trials. Results so far demonstrated good efficacy of aminocandin against fluconazole-resistant C. albicans [134] and itraconazole-resistant Aspergillus fumigatus strains [135].
One of the most important characteristics of these semisynthetic echinocadins is their activity against biofilms. Micafungin showed antifungal activity against Candida albicans and C. parapsilosis biofilms, but has a better activity against biofilms with high metabolic activity [136,137]. Micafungin showed a considerably higher acitivity against C. tropicalis biofilms when compared to amphotericin [138]. Caspofungin could eradicate mature C. lusitaniae or C. guilliermondii biofilms at concentrations feasible for catheters [139]. Nevertheless, some biofilms of C. albicans are difficult to clear by echinocandins as an animal model of foreign-body infection demonstrated. Here, anidulafungin or caspofungin reached only a cure rate of 25%, with caspofungin being the most effective [140]. In an in vitro model, andulafungin was effective against nine Candida strains, four of them were C. albicans. However, when combined with the nonsteroidal anti-inflammatory drugs aspirin, diclofenac or ibuprofen, a clear synergism in biofilm clearance was observed [141]. The effect of caspofungin or micafungin alone or in combination with farnesol 7 against C. parapsilosis biofilms has been tested. Both echinocandins could clear the biofilm, but were up to 64-fold more effective when combined with farnesol. Interestingly, micafungin could not clear biofilms older than 24 h [142].
3. Conclusions
The present review brings an extensive, although not exhaustive, list of small metabolites from fungi which have anti-biofilm activity. Figure 9 summarizes examples of biofilm formation mechanisms which are targeted by fungal bioactive compounds. The pathway from finding interesting activities in fungal extracts to actual drug discovery, however, is a long and winding one. Among the hundred compounds discussed here, only three echinocandins (caspofungin, micafungin. and anidulafungin) were fully developed and are currently approved as commercial drugs. They are indicated mainly for the treatment of invasive or disseminated fungal infections, and despite being notorious for their ability to disrupt fungal biofilms, their clinical application against biofilm-related diseases and colonization of medical devices is still being continuously developed [107,133].
Two important issues are confronted when applying echinocandins in the clinics. One, which is inherent to virtually all antibiotics, is the increasing number of clinical strains displaying resistance [143]. Another concern, which is rather distinctive of echinocandins, is known as the paradoxical effect; it is characterized by an ability of the target organism to reverse growth inhibition at high concentrations, above the MIC drug concentrations [144]. Both phenomena jeopardize the efficacy of such drugs in the clinics, and deserve researchers’ attention. Importantly, echinocandin resistance, as well as the paradoxical effect, is shown to be divergent between planktonic cells and biofilms of given Candida isolates [145]. The mechanism of action of echinocandins has the cell wall synthesis as a major target, and mutations in the beta-glucan synthase enzyme (fks1 gene) are implicated in resistance-development [146]. The paradoxical effect, in turn, is also correlated to cell wall stress and general stress responses, including Hsp90. Additionally, the influence of paradoxical effect-related stress responses in the induction of resistance development has also been demonstrated [147]. Altogether, it highlights the need for a continuous effort from the research community, to maintain or enhance the effectiveness of currently available anti-fungal agents, as well as to open up new avenues from natural compounds with divergent modes of action.
Apart from glucan-synthesis inhibition by antifungal compounds, a number of the metabolites reviewed here act through different mechanisms, some even combining more than one target. This is the case for terreic acid, which has been shown to inhibit two different bacterial biosynthetic enzymes [69,70]. The multi-target trait is a highly desired one when trying to avoid resistance development. Also, some diketopiperazines are reported to be multi-functional in that they are able to inhibit Gram-negative biofilm formation by competing with acyl-homoserine lactone quorum-sensing signaling, as well as to interfere with Gram-positive agr quorum-sensing system [83]. Focusing on quorum-quenching compounds will likely expand the potential use of fungal metabolites in the control of bacterial biofilms. Indeed, several compounds of different classes can be highlighted here for displaying both anti-fungal and anti-bacterial activities (e.g., farnesol, tyrosol, usnic acid, cytosporone E and some echinocandins), and these should be considered of particular interest for drug development.
Noticeably, most studies on anti-biofilm compounds of fungal origin to date have demonstrated their ability to kill and/or disrupt fungal biofilms, and in particular the model organism and medically important pathogen genus Candida. Accordingly, biofilm-related fungal infections are the targets for a number of fungal metabolites tested in preclinical setups, i.e., using in vivo models, medical surfaces or isolates from patients. Also bacterial biofilm infections and mixed-species biofilms (as is often the reality in the clinics) are being addressed in some such studies. Table 1 gives a summary of preclinical results on selected fungal compounds, with focus on substantial advancements in their prospective use as anti-biofilm drugs, indicating they are in continuous progress towards clinical application. Further reading on in vivo models for anti-biofilm drug discovery, and translational research of echinocandins can be found in recent reviews [107,148,149].
4. Outlook
Additional alternative strategies to benefit from anti-biofilm compounds of fungal origin in the treatment of infectious diseases are under study, which may in the future offer promising results in the clinics. One such strategy is the combination of fungal metabolites with other classes of anti-fungal drugs, to enhance their performance, especially towards biofilms (some examples are presented in Table 1, and a recent review is available [186]). Another approach consists in immobilizing the drugs on medical surfaces. This has been described for caspofungin on propanal biosurfaces [187] and on titanium substrata which were therefore rendered less susceptible to C. albicans biofilm colonization, at the same time supporting osseointegration (integration of the implant into the bone) [188]. Osteocompatibility is an important trait to be taken into account, the lack of which can limit the clinical use of cytotoxic compounds (e.g., farnesol, [189]). Also, usnic acid has been successfully incorporated onto biodegradable polymer thin films [190], or cellulose-silica membranes [191] for the production of biocompatible, S. aureus-resistant surfaces. A third possibility still to be explored is the production of nanostructures for a more efficient delivery of anti-biofilm drugs. This has recently been tested with cefotaxime-impregnated chitosan particles [192] and farnesol-loaded cationic nanoparticles [193]. Polymer microparticles and magnetic nanoparticles also yielded positive results as carriers for usnic acid in vitro antimicrobial and anti-biofilm assays [194,195].
An astounding biodiversity found within the kingdom Fungi, which is estimated to include not less than five million species [196], associated to an unparalleled metabolic diversity inherent to a broad range of ecological strategies [197], draws attention to the biotechnological potential contained among fungal secondary metabolites. The number of different compounds summarized in this review highlights the usefulness of such sources in the much-needed search for novel antibiotic drugs which are effective against biofilm infections [198]. This urging issue is not a novelty in the context of the so-called “post-antibiotic era”, neither is the fact surprising that natural compounds’ research still consists of a promising approach. Not only the constant exposure of fungal tissues to other microorganisms in favorable environmental conditions, posing a natural demand for the production of anti-biofilm metabolites to avoid potentially harmful inter-species interactions, but also the biochemical machinery employed in intra-specific cell communication and self-regulation seem to offer us valuable leads for anti-biofilm drug development. We believe that a thorough exploration of natural compounds from fungi will broaden the possibilities for their use in the treatment of biofilm infections, offering novel ways to close the gaps left by current therapies.
Acknowledgments
The many valuable discussions by colleagues within the European Graduate School “Pseudomonas: biotechnology and pathogenicity” and the Collaborative Research Centre “Sustainable bioresorbable and permanent implants of metallic and ceramic materials” (SFB 599), both supported by the German Research Foundation, are sincerely acknowledged. A.B.E. is thankful for the support by a stipend of the “Science without border (Ciência sem Fronteiras)” program of Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) from Brazil and the German Academic Exchange Service (DAAD).
Author Contributions
Both authors were involved in searching and evaluating the literature and writing the review. W.R.A. did the drawings of chemical formulae.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| MIC | Minimal inhibitory concentration |
| IC50 | Half maximal inhibitory concentration |
References
- Singh, P.K.; Schaefer, A.L.; Parsek, M.R.; Moninger, T.O.; Welsh, M.J.; Greenberg, E.P. Quorum-sensing signals indicate that cystic fibrosis lungs are infected with bacterial biofilms. Nature 2000, 407, 762–764. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Hu, F.Z.; Gieseke, A.; Nistico, L.; Nguyen, D.; Hayes, J.; Forbes, M.; Greenberg, D.P.; Dice, B.; Burrows, A.; et al. Direct detection of bacterial biofilms on the middle-ear mucosa of children with chronic otitis media. JAMA 2006, 296, 202–211. [Google Scholar] [CrossRef] [PubMed]
- Rosen Da, H.T.; Stamm, W.E.; Humphrey, P.A.; Hultgren, S.J. Detection of intracellular bacterial communities in human urinary tract infection. PLoS Med. 2007, 4, e329. [Google Scholar] [CrossRef] [PubMed]
- Carron, M.A.; Tran, V.R.; Sugawa, C.; Coticchia, J.M. Identification of Helicobacter pylori biofilms in human gastric mucosa. J. Gastrointest. Surg. 2006, 10, 712–717. [Google Scholar] [CrossRef] [PubMed]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Leone, M.; Dillon, L.R. Catheter outcomes in home infusion. J. Infus. Nurs. 2008, 31, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Camilli, A.; Bassler, B.L. Bacterial small-molecule signaling pathways. Science 2006, 311, 1113–1116. [Google Scholar] [CrossRef] [PubMed]
- Schauder, S.; Bassler, B.L. The languages of bacteria. Genes Dev. 2001, 15, 1468–1480. [Google Scholar] [CrossRef] [PubMed]
- Abraham, W.-R. Controlling Gram-negative pathogenic bacteria by interfering with their biofilm formation. Drug Des. Rev. Online 2005, 2, 13–33. [Google Scholar] [CrossRef]
- Chen, X.; Schauder, S.; Potier, N.; Van Dorsselaer, A.; Pelczer, I.; Bassler, B.L.; Hughson, F.M. Structural identification of a bacterial quorum-sensing signal containing boron. Nature 2002, 415, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Lyon, G.J.; Novick, R.P. Peptide signaling in Staphylococcus aureus and other Gram-positive bacteria. Peptides 2004, 25, 1389–1403. [Google Scholar] [CrossRef] [PubMed]
- Cottier, F.; Mühlschlegel, F.A. Communication in Fungi. Int. J. Microbiol. 2012, 351832. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque, P.; Casadevall, A. Quorum sensing in fungi—A review. Med. Mycol. 2012, 50, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Nobile, C.J.; Johnson, A.D. Candida albicans biofilms and human disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef] [PubMed]
- Alem, M.A.S.; Oteef, M.D.Y.; Flowers, T.H.; Douglas, L.J. Production of tyrosol by Candida albicans biofilms and its role in quorum sensing and biofilm development. Eukaryot. Cell 2006, 5, 1770–1779. [Google Scholar] [CrossRef] [PubMed]
- Hogan, D.A. Talking to themselves: Autoregulation and quorum sensing in fungi. Eukaryot. Cell 2006, 5, 613–619. [Google Scholar] [CrossRef] [PubMed]
- Nickerson, K.W.; Atkin, A.L.; Hornby, J.M. Quorum sensing in dimorphic fungi: Farnesol and beyond. Appl. Environ. Microbiol. 2006, 72, 3805–3813. [Google Scholar] [CrossRef] [PubMed]
- Nigam, S.; Ciccoli, R.; Ivanov, I.; Sczepanski, M.; Deva, R. On mechanism of quorum sensing in Candida albicans by 3(R)-hydroxy-tetradecaenoic acid. Curr. Microbiol. 2011, 62, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, M.; Obata, S.; Shimizu, S. Microorganisms for Production of Geranylgeraniol and Analogous Compounds. European Patent EP 1,219,714, 2 July 2002. [Google Scholar]
- Harriott, M.M.; Noverr, M.C. Importance of Candida-bacterial polymicrobial biofilms in disease. Trends Microbiol. 2011, 19, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Wargo, M.J.; Hogan, D.A. Fungal—Bacterial interactions: A mixed bag of mingling microbes. Curr. Opin. Microbiol. 2006, 9, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Costerton, J.W. Antibiotic resistance of bacteria in biofilms. Lancet 2001, 358, 135–138. [Google Scholar] [CrossRef]
- Joly, V.; Pangon, B.; Vallois, J.M.; Abel, L.; Brion, N.; Bure, A.; Chau, N.P.; Contrepois, A.; Carbon, C. Value of antibiotic levels in serum and cardiac vegetations for predicting antibacterial effect of ceftriaxone in experimental Escherichia coli endocarditis. Antimicrob. Agents Chemother. 1987, 31, 1632–1639. [Google Scholar] [CrossRef] [PubMed]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal biofilm resistance. Int. J. Microbiol. 2012, 528521. [Google Scholar] [CrossRef] [PubMed]
- Austin, D.J.; Kristinsson, K.G.; Anderson, R.M. The relationship between the volume of antimicrobial consumption in human communities and the frequency of resistance. Proc. Natl. Acad. Sci. USA 1999, 96, 1152–1156. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.J. Microbiology. Desperately seeking new antibiotics. Science 2008, 321, 1644–1645. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Something old, something new: Revisiting natural products in antibiotic drug discovery. Can. J. Microbiol. 2014, 60, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Worthington, R.J.; Richards, J.J.; Melander, C. Non-microbicidal control of bacterial biofilms with small molecules. Anti-Inf. Agents 2014, 12, 120–138. [Google Scholar] [CrossRef]
- Zhu, J.; Kaufmann, G.F. Quo vadis quorum quenching? Curr. Opin. Pharmacol. 2013, 13, 688–898. [Google Scholar] [CrossRef] [PubMed]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L., Jr.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Meng, W.; Cao, C.; Wang, J.; Shan, W.; Wang, Q. Antibacterial and antifungal compounds from marine fungi. Mar. Drugs 2015, 13, 3479–3513. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, M.P.; Türck, P.; Abraham, W.-R. Secondary metabolites control the associated bacterial communities of saprophytic Basidiomycotina fungi. Microbes Environ. 2015, 30, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Robles, A.J.; King, J.B.; Powell, D.R.; Miller, A.N.; Mooberry, S.L.; Cichewicz, R.H. Crowdsourcing natural products discovery to access uncharted dimensions of fungal metabolite diversity. Angew. Chem. Int. Ed. Engl. 2014, 53, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Spratt, D.A.; Daglia, M.; Papetti, A.; Stauder, M.; O’Donnell, D.; Ciric, L.; Tymon, A.; Repetto, B.; Signoretto, C.; Houri-Haddad, Y.; et al. Evaluation of plant and fungal extracts for their potential antigingivitis and anticaries activity. J. Biomed. Biotechnol. 2012, 510198. [Google Scholar] [CrossRef] [PubMed]
- Signoretto, C.; Marchi, A.; Bertoncelli, A.; Burlacchini, G.; Papetti, A.; Pruzzo, C.; Zaura, E.; Lingström, P.; Ofek, I.; Pratten, J.; et al. The anti-adhesive mode of action of a purified mushroom (Lentinus edodes) extract with anticaries and antigingivitis properties in two oral bacterial pathogens. BMC Compl. Altern. Med. 2014, 14, 75. [Google Scholar] [CrossRef] [PubMed]
- Signoretto, C.; Marchi, A.; Bertoncelli, A.; Burlacchini, G.; Milli, A.; Tessarolo, F.; Caola, I.; Papetti, A.; Pruzzo, C.; Zaura, E.; et al. Effects of mushroom and chicory extracts on the shape, physiology and proteome of the cariogenic bacterium Streptococcus mutans. BMC Compl. Altern. Med. 2013, 13, 117. [Google Scholar] [CrossRef] [PubMed]
- Boustie, J.; Tomasi, S.; Grube, M. Bioactive lichen metabolites: Alpine habitats as an untapped source. Phytochem. Rev. 2011, 10, 287–307. [Google Scholar] [CrossRef]
- Ramage, G.; Saville, S.P.; Wickes, B.L.; Lopez-Ribot, J.L. Inhibition of Candida albicans biofilm formation by farnesol, a quorum-sensing molecule. Appl. Environ. Microbiol. 2002, 68, 5459–5463. [Google Scholar] [CrossRef] [PubMed]
- Weber, K.; Sohr, R.; Schulz, B.; Fleischhacker, M.; Ruhnke, M. Secretion of E,E-farnesol and biofilm formation in eight different Candida species. Antimicrob. Agents Chemother. 2008, 52, 1859–1861. [Google Scholar] [CrossRef]
- Cushion, M.T.; Collins, M.S.; Linke, M.J. Biofilm formation by Pneumocystis spp. Eukaryot. Cell 2009, 8, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Jabra-Rizk, M.A.; Meiller, T.F.; James, C.E.; Shirtliff, M.E. Effect of farnesol on Staphylococcus aureus biofilm formation and antimicrobial susceptibility. Antimicrob. Agents Chemother. 2006, 50, 1463–1469. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.I.A.; Teixeira, P.; Azeredo, J.; Oliveira, R. Effect of farnesol on planktonic and biofilm cells of Staphylococcus epidermidis. Curr. Microbiol. 2009, 59, 118–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, F.; Teixeira, P.; Cerca, N.; Azeredo, J.; Oliveira, R. Effect of farnesol on structure and composition of Staphylococcus epidermidis biofilm matrix. Curr. Microbiol. 2011, 63, 354–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koo, H.; Hayacibara, M.F.; Schobel, B.D.; Cury, J.A.; Rosalen, P.L.; Park, Y.K.; Vacca-Smith, A.M.; Bowen, W.H. Inhibition of Streptococcus mutans biofilm accumulation and polysaccharide production by apigenin and tt-farnesol. J. Antimicrob. Chemother. 2003, 52, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.-H.; Wei, X.; Ma, M.; Chen, X.-J.; Xu, S.-B. Possible inhibitory molecular mechanism of farnesol on the development of fluconazole resistance in Candida albicans biofilm. Antimicrob. Agents Chemother. 2012, 56, 770–775. [Google Scholar] [CrossRef] [PubMed]
- Jabra-Rizk, M.A.; Shirtliff, M.; James, C.; Meiller, T. Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res. 2006, 6, 1063–1073. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Mekala, S.; Chaffin, W.L. Farnesol-mediated inhibition of Candida albicans yeast growth and rescue by a diacylglycerol analogue. Yeast 2007, 24, 681–693. [Google Scholar] [CrossRef] [PubMed]
- Brackman, G.; Hillaert, U.; Van Calenbergh, S.; Nelis, H.J.; Coenye, T. Use of quorum sensing inhibitors to interfere with biofilm formation and development in Burkholderia multivorans and Burkholderia cenocepacia. Res. Microbiol. 2009, 160, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cugini, C.; Calfee, M.W.; Farrow, J.M., III; Morales, D.K.; Pesci, E.C.; Hogan, D.A. Farnesol, a common sesquiterpene, inhibits PQS production in Pseudomonas aeruginosa. Mol. Microbiol. 2007, 65, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Shirtliff, M.E.; Krom, B.P.; Meijering, R.A.M.; Peters, B.M.; Zhu, J.; Scheper, M.A.; Harris, M.L.; Jabra-Rizk, M.A. Farnesol-induced apoptosis in Candida albicans. Antimocrob. Agents Chemother. 2009, 53, 2392–2401. [Google Scholar] [CrossRef] [PubMed]
- Semighini, C.P.; Hornby, J.M.; Dumitru, R.; Nickerson, K.W.; Harris, S.D. Farnesol-induced apoptosis in Aspergillus nidulans reveals a possible mechanism for antagonistic interactions between fungi. Mol. Microbiol. 2006, 59, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Fairn, G.D.; MacDonald, K.; McMaster, C.R. A chemogenomic screen in Saccharomyces cerevisiae uncovers a primary role for the mitochondria in farnesol toxicity and its regulation by the Pkc1 pathway. J. Biol. Chem. 2007, 282, 4868–4874. [Google Scholar] [CrossRef] [PubMed]
- Scheper, M.A.; Shirtliff, M.E.; Meiller, T.F.; Peters, B.; Jabra-Rizk, M.A. Farnesol a fungal quorum sensing molecule triggers apoptosis in human oral squamous carcinoma cells. Neoplasia 2008, 10, 954–963. [Google Scholar] [CrossRef] [PubMed]
- Scopel, M.; Abraham, W.-R.; Antunes, A.L.; Barth, A.; Ribeiro, V.B.; Henriques, A.T.; Macedo, A.J. Mevalonolactone: An inhibitor of Staphylococcus epidermidis adherence and biofilm formation. Med. Chem. 2014, 10, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Arai, M.; Niikawa, H.; Kobayashi, M. Marine-derived fungal sesterterpenes, ophiobolins, inhibit biofilm formation of Mycobacterium species. J. Nat. Med. 2013, 67, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Eamvijarn, A.; Gomes, N.M.; Dethoup, T.; Buaruang, J.; Manoch, L.; Silva, A.; Pedro, M.; Marini, I.; Roussis, V.; Kijjoa, A. Bioactive meroditerpenes and indole alkaloids from the soil fungus Neosartorya fischeri (KUFC 6344), and the marine-derived fungi Neosartorya laciniosa (KUFC 7896) and Neosartorya tsunodae (KUFC 9213). Tetrahedron 2013, 69, 8583–8591. [Google Scholar] [CrossRef]
- Gomes, N.M.; Bessa, L.J.; Buttachon, S.; Costa, P.M.; Buaruang, J.D.; Tida, S.A.M.S.; Kijjoa, A. Antibacterial and antibiofilm activities of tryptoquivalines and meroditerpenes isolated from the marine-derived fungi Neosartorya paulistensis, N. laciniosa, N. tsunodae, and the soil fungi N. fischeri and N. siamensis. Mar. Drugs 2014, 12, 822–839. [Google Scholar] [CrossRef] [PubMed]
- Li, T.X.; Yang, M.H.; Wang, X.B.; Wang, Y.; Kong, L.Y. Synergistic antifungal meroterpenes and dioxolanone derivatives from the endophytic fungus Guignardia sp. J. Nat. Prod. 2015, 78, 2511–2520. [Google Scholar] [CrossRef] [PubMed]
- Cocchietto, M.; Skert, N.; Nimis, P.L.; Sava, G. A review on usnic acid, an interesting natural compound. Naturwissenschaften 2002, 89, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Araújo, A.A.; de Melo, M.G.; Rabelo, T.K.; Nunes, P.S.; Santos, S.L.; Serafini, M.R.; Santos, M.R.; Quintans-Júnior, L.J.; Gelain, D.P. Review of the biological properties and toxicity of usnic acid. Nat. Prod. Res. 2015, 29, 2167–2180. [Google Scholar] [CrossRef] [PubMed]
- Nithyanand, P.; Shafreen, R.M.B.; Muthamil, S.; Pandian, S.K. Usnic acid, a lichen secondary metabolite inhibits group A Streptococcus biofilms. Antonie Leeuwenhoek 2015, 107, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Nithyanand, P.; Beema Shafreen, R.M.; Muthamil, S.; Karutha Pandian, S. Usnic acid inhibits biofilm formation and virulent morphological traits of Candida albicans. Microbiol. Res. 2015, 179, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Francolini, I.; Norris, P.; Piozzi, A.; Donelli, G.; Stoodley, P. Usnic acid, a natural antimicrobial agent able to inhibit bacterial biofilm formation on polymer surfaces. Antimic. Agents Chemother. 2004, 48, 4360–4365. [Google Scholar] [CrossRef] [PubMed]
- Riedel, K.; Boustie, J.; Eberl, L.; Berg, G.; Grube, M. Effects of lichen secondary metabolites on bacterial functions and biofilm formation. Planta Med. 2008, 74, PA85. [Google Scholar] [CrossRef]
- Millot, M.; Girardot, M.; Dutreix, L.; Imbert, C.; Mambu, L. Lichen biodiversity: A source of secondary metabolites active against Candida biofilms. Planta Med. 2014, 80, P1N24. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Skindersoe, M.E.; Bjarnsholt, T.; Phipps, R.K.; Christensen, K.B.; Jensen, P.O.; Andersen, J.B.; Koch, B.; Larsen, T.O.; Hentzer, M. Identity and effects of quorum-sensing inhibitors produced by Penicillium species. Microbiology 2005, 151, 1325–1340. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, M.P. Use of Comatuslactone to Modulate Microbial Biofilms. Ph.D. Thesis, Technical University Braunschweig, Braunschweig, Germany, 1 April 2014. [Google Scholar]
- Beau, J.; Mahid, N.; Burda, W.N.; Harrington, L.; Shaw, L.N.; Mutka, T.; Kyle, D.E.; Barisic, B.; van Olphen, A.; Baker, B.J. Epigenetic tailoring for the production of anti-infective cytosporones from the marine fungus Leucostoma persoonii. Mar. Drugs 2012, 10, 762–774. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Lambu, M.R.; Jamwal, U.; Rani, C.; Chib, R.; Wazir, P.; Mukherjee, D.; Chaubey, A.; Khan, I.A. Escherichia coli N-acetylglucosamine-1-phosphate-uridyltransferase/glucosamine-1-phosphate-acetyltransferase (GlmU) inhibitory activity of terreic acid isolated from Aspergillus terreus. J. Biomol. Screen. 2016, 21, 342–353. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yang, Y.; Olesen, S.H.; Becker, A.; Betzi, S.; Schönbrunn, E. The fungal product terreic acid is a covalent inhibitor of the bacterial cell wall biosynthetic enzyme UDP-N-acetylglucosamine 1-carboxyvinyltransferase (MurA). Biochemistry 2010, 49, 4276–4282. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.F.; Li, P.J.; Li, X.X.; Sun, P.H.; Gao, H.; Liu, X.Z.; Huang, P.; Tang, J.S.; Yao, X.S. New antibacterial isocoumarin glycosides from a wetland soil derived fungal strain Metarhizium anisopliae. Bioorg. Med. Chem. Lett. 2016, 26, 1391–1396. [Google Scholar] [CrossRef] [PubMed]
- Kluepfel, D.; Bagli, J.; Baker, H.; Charest, M.P.; Kudelski, A. Myriocin, a new antifungal antibiotic from Myriococcum albomyces. J. Antibiot. 1972, 25, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Kozutsumi, Y.; Nakamura, S.; Fujita, T.; Kawasaki, T. Serine palmitoyltransferase is the primary target of a sphingosine-like immunosuppressant, ISP-1/myriocin. Biochem. Biophys. Res. Commun. 1995, 211, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Lattif, A.A.; Mukherjee, P.K.; Chandra, J.; Roth, M.R.; Welti, R.; Rouabhia, M.; Ghannoum, M.A. Lipidomics of Candida albicans biofilms reveals phase-dependent production of phospholipid molecular classes and role for lipid rafts in biofilm formation. Microbiology 2011, 157, 3232–3242. [Google Scholar] [CrossRef] [PubMed]
- Perdoni, F.; Signorelli, P.; Cirasola, D.; Caretti, A.; Galimberti, V.; Biggiogera, M.; Gasco, P.; Musicanti, C.; Morace, G.; Borghi, E. Antifungal activity of Myriocin on clinically relevant Aspergillus fumigatus strains producing biofilm. BMC Microbiol. 2015, 15, 248. [Google Scholar] [CrossRef] [PubMed]
- You, J.; Du, L.; King, J.B.; Hall, B.E.; Cichewicz, R.H. Small-molecule suppressors of Candida albicans biofilm formation synergistically enhance the antifungal activity of amphotericin B against clinical Candida isolates. ACS Chem. Biol. 2013, 8, 840–848. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; You, J.; King, J.B.; Cai, S.; Park, E.; Powell, D.R.; Cichewicz, R.H. Polyketide glycosides from Bionectria ochroleuca inhibit Candida albicans biofilm formation. J. Nat. Prod. 2014, 77, 2273–2279. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; You, J.; King, J.B.; Powell, D.R.; Cichewicz, R.H. Waikialoid A suppresses hyphal morphogenesis and inhibits biofilm development in pathogenic Candida albicans. J. Nat. Prod. 2012, 75, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Arias, L.S.; Delbem, A.C.; Fernandes, R.A.; Barbosa, D.B.; Monteiro, D.R. Activity of tyrosol against single and mixed-species oral biofilms. J. Appl. Microbiol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y. Studies on the metabolic products of Rosellinia necatrix. I. Isolation and characterization of several physiologically active neutral substances. Bull. Agric. Chem. Soc. Jpn. 1960, 24, 372–381. [Google Scholar]
- Stierle, A.C.; Cardellina, J.H., II; Strobel, G.A. Maculosin, a host-specific phytotoxin for spotted knapweed from Alternaria alternata. Proc. Natl. Acad. Sci. USA 1988, 85, 8008–8011. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.P.; Dow, J.M. Diffusible signals and interspecies communication in bacteria. Microbiology 2008, 154, 1845–1858. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, W.; Xu, S.X.; Magarvey, N.A.; McCormick, J.K. Lactobacillus reuteri-produced cyclic dipeptides quench agr-mediated expression of toxic shock syndrome toxin-1 in staphylococci. Proc. Natl. Acad. Sci. USA 2011, 108, 3360–3365. [Google Scholar] [CrossRef] [PubMed]
- Scopel, M.; Abraham, W.-R.; Henriques, A.T.; Macedo, A.J. Dipeptide cis-cyclo(leucyl-tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorg. Med. Chem. Lett. 2013, 23, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Diblasi, L.; Arrighi, F.; Silva, J.; Bardón, A.; Cartagena, E. Penicillium commune metabolic profile as a promising source of antipathogenic natural products. Nat. Prod. Res. 2015, 29, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Qi, S.-H.; Xu, Y.; Xiong, H.-R.; Qian, P.-Y.; Zhang, S. Antifouling and antibacterial compounds from a marine fungus Cladosporium sp. F14. World J. Microbiol. Biotechnol. 2009, 25, 399–406. [Google Scholar] [CrossRef]
- Bai, Z.Q.; Lin, X.; Wang, Y.; Wang, J.; Zhou, X.; Yang, B.; Liu, J.; Yang, X.; Wang, Y.; Liu, Y. New phenyl derivatives from endophytic fungus Aspergillus flavipes AIL8 derived of mangrove plant Acanthus ilicifolius. Fitoterapia 2014, 95, 194–202. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, M.P.; Abraham, W.-R. Antimicrobial and biofilm inhibiting diketopiperazines. Curr. Med. Chem. 2012, 19, 3564–3577. [Google Scholar] [CrossRef] [PubMed]
- Campbell, J.; Lin, Q.; Geske, G.D.; Blackwell, H.E. New and unexpected insights into the modulation of LuxR-type quorum sensing by cyclic dipeptides. ACS Chem. Biol. 2009, 4, 1051–1059. [Google Scholar] [CrossRef] [PubMed]
- Estrela, A.B.; Heck, M.G.; Abraham, W.-R. Novel approaches to control biofilm infections. Curr. Med. Chem. 2009, 16, 1512–1530. [Google Scholar] [CrossRef] [PubMed]
- Traxler, P.; Fritz, H.; Fuhrer, H.; Richter, W.J. Papulacandins, a new family of antibiotics with antifungal activity. Structures of papulacandins A, B, C and D. J. Antibiot. 1980, 33, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Nett, J.; Lincoln, L.; Marchillo, K.; Massey, R.; Holoyda, K.; Hoff, B.; VanHandel, M.; Andes, D. Putative role of β-1,3 glucans in Candida albicans biofilm resistance. Antimicrob. Agents Chemother. 2007, 51, 510–520. [Google Scholar] [CrossRef] [PubMed]
- VanMiddlesworth, F.; Omstead, M.N.; Schmatz, D.; Bartizal, K.; Fromtling, R.; Bills, G.; Nollstadt, K.; Honeycutt, S.; Zweerink, M.; Garrity, G.; et al. L-687,781, a new member of the papulacandin family of beta-1,3-d-glucan synthesis inhibitors. I. Fermentation, isolation, and biological activity. J. Antibiot. 1991, 44, 45–51. [Google Scholar] [PubMed]
- VanMiddlesworth, F.; Dufresne, C.; Smith, J.; Wilson, K.E. Structure elucidation of L-687,781, a new β-1,3-d-glucan synthesis inhibitor. Tetrahedron 1991, 47, 7563–7568. [Google Scholar] [CrossRef]
- Kaneto, R.; Chiba, H.; Agematu, H.; Shibamoto, N.; Yoshioka, T.; Nishida, H.; Okamoto, R. Mer-WF3010, a new member of the papulacandin family. I. Fermentation, isolation and characterization. J. Antibiot. 1993, 46, 247–250. [Google Scholar] [CrossRef] [PubMed]
- Chiba, H.; Kaneto, R.; Agematu, H.; Shibamoto, N.; Yoshioka, T.; Nishida, H.; Okamoto, R. Mer-WF3010, a new member of the papulacandin family. II. Structure determination. J. Antibiot. 1993, 46, 356–358. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, T.; Iwadate-Kurihara, Y.; Hosoya, T.; Ishikawa, T.; Miyakoshi, S.; Hamano, K.; Inukai, M. F-10748 A1, A2, B1, B2, C1, C2, D1 and D2, novel papulacandins. J. Antibiot. 2002, 55, 758–763. [Google Scholar] [CrossRef] [PubMed]
- Aoki, M.; Andoh, T.; Ueki, T.; Masuyoshi, S.; Sugawara, K.; Oki, T. BU-4794F, a new beta-1,3-glucan synthase inhibitor. J. Antibiot. 1993, 46, 952–960. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.H.; Tennant, S.; Frost, D.; O’Beirne, M.J.; Karwowski, J.P.; Humphrey, P.E.; Malmberg, L.-H.; Choi, W.; Brandt, K.D.; West, P.; et al. Discovery of saricandin, a novel papulacandin, from a Fusarium species. J. Antibiot. 1996, 49, 596–598. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Yamashita, M.; Tsurumi, Y.; Kohsaka, M. Chaetiacandin, a novel papulacandin. I. Fermentation, isolation and characterization. J. Antibiot. 1985, 38, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Komori, T.; Itoh, Y. Chaetiacandin, a novel papulacandin. II. Structure determination. J. Antibiot. 1985, 38, 544–546. [Google Scholar] [CrossRef] [PubMed]
- Gunawardana, G.; Rasmussen, R.R.; Scherr, M.; Frost, D.; Brandt, K.D.; Choi, W.; Jackson, M.; Karwowski, J.P.; Sunga, G.; Malmberg, L.-H.; et al. Corynecandin: A novel antifungal glycolipid from Coryneum modonium. J. Antibiot. 1997, 50, 884–886. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.; Frost, D.J.; Karwowski, J.P.; Humphrey, P.E.; Dahod, S.K.; Choi, W.S.; Brandt, K.; Malmberg, L.-H.; Rasmussen, R.R.; Scherr, M.H.; et al. Fusacandins A and B; novel antifungal antibiotics of the papulacandin class from Fusarium sambucinum. I. Identity of the producing organism, fermentation and biological activity. J. Antibiot. 1995, 48, 608–613. [Google Scholar] [CrossRef] [PubMed]
- Hochlowski, J.E.; Whittern, D.N.; Buko, A.; Alder, L.; McAlpine, J.B. Fusacandins A and B; novel antifungal antibiotics of the papulacandin class from Fusarium sambucinum. II. Isolation and structural elucidation. J. Antibiot. 1995, 48, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Van der Kaaden, M.; Breukink, E.; Pieters, R.J. Synthesis and antifungal properties of papulacandin derivatives. Beilstein J. Org. Chem. 2012, 8, 732–737. [Google Scholar] [CrossRef] [PubMed]
- Römmele, G.; Traxler, P.; Wehrli, W. Papulacandins—The relationship between chemical structure and effect on glucan synthesis in yeast. J. Antibiot. 1983, 36, 1539–1542. [Google Scholar] [CrossRef] [PubMed]
- Katragkou, A.; Roilides, E.; Walsh, T.J. Role of echinocandins in fungal biofilm-related disease: Vascular catheter-related infections, immunomodulation, and mucosal surfaces. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S622–S629. [Google Scholar] [CrossRef] [PubMed]
- Arvanitis, M.; Mylonakis, E. Characteristics, clinical relevance, and the role of echinocandins in fungal-bacterial interactions. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S630–S634. [Google Scholar] [CrossRef] [PubMed]
- Nyfeler, R.; Keller-Schierlein, W. Metabolites of microorganisms. 143. Echinocandin B, a novel polypeptide-antibiotic from Aspergillus nidulans var. echinulatus: Isolation and structural components. Helv. Chim. Acta 1974, 57, 2459–2477. [Google Scholar] [PubMed]
- Keller-Juslén, C.; Kuhn, M.; Loosli, H.R.; Petcher, T.J.; Weber, H.P.; von Wartburg, A. Struktur des Cyclopeptid-Antibiotikums sl 7810 (= Echinocandin B). Helv. Chim. Acta 1976, 17, 4147–4150. [Google Scholar]
- Emri, T.; Majoros, L.; Tóth, V.; Pócsi, I. Echinocandins: Production and applications. Appl. Microbiol. Biotechnol. 2013, 97, 3267–3284. [Google Scholar] [CrossRef] [PubMed]
- Traber, R.; Keller-Juslén, C.; Loosli, H.-R.; Kuhn, M.; Von Wartburg, A. Cyclopeptid-Antibiotika aus Aspergillus-Arten. Struktur der Echinocandine C und D. Helv. Chim. Acta 1979, 62, 1252–1267. [Google Scholar] [CrossRef]
- Mizuno, K.; Yagi, A.; Satoi, S.; Takada, M.; Hayashi, M.; Asano, K.; Matsuda, T. Studies on aculeacin. I. Isolation and characterization of aculeacin A. J. Antibiot. 1977, 30, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Satoi, S.; Yagi, A.; Asano, K.; Mizuno, K.; Watanabe, T. Studies of aculeacin. II. Isolation and characterization of aculeacins B, C, D, E, F, and G. J. Antibiot. 1977, 30, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, R.; Kobayashi, M.; Fujine, K.; Sato, I.; Hashimoto, M.; Takase, S.; Tsurumi, Y.; Fujie, A.; Hino, M.; Hashimoto, S.; et al. FR227673 and FR190293, novel antifungal lipopeptides from Chalara sp. No. 22210 and Tolypocladium parasiticum No. 16616. J. Antibiot. 2006, 59, 158–167. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, R.; Sakamoto, K.; Hashimoto, M.; Takase, S.; Tsurumi, Y.; Fujie, A.; Hino, M.; Hashimoto, S.; Hori, Y. FR209602 and related compounds, novel antifungal lipopeptides from Coleophoma crateriformis no.738. I. Taxonomy, fermentation, isolation and physico-chemical properties. J. Antibiot. 2006, 59, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Bills, G.F.; Yue, Q.; Chen, L.; Li, Y.; An, Z.; Frisvad, J.C. Aspergillus mulundensis sp. nov., a new species for the fungus producing the antifungal echinocandin lipopeptides, mulundocandins. J. Antibiot. 2016, 69, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Hensens, O.D.; Liesch, J.M.; Zink, D.L.; Smith, J.L.; Wichmann, C.F.; Schwartz, R.E. Pneumocandins from Zalerion arboricola. III. Structure elucidation. J. Antibiot. 1992, 45, 1875–1885. [Google Scholar] [CrossRef] [PubMed]
- Tscherter, H.; Dreyfuss, M.M. Antibiotics from a Cryptosporiopsis Species and Their Therapeutic Use. Sandoz S.A, 1982. Belgian Patent 889,955, 15 February 1982. [Google Scholar]
- Bills, G.; Li, Y.; Chen, L.; Yue, Q.; Niu, X.M.; An, Z. New insights into the echinocandins and other fungal non-ribosomal peptides and peptaibiotics. Nat. Prod. Rep. 2014, 31, 1348–1375. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Chen, L.; Zhang, X.; Li, K.; Sun, J.; Liu, X.; An, Z.; Bills, G.F. Evolution of chemical diversity in echinocandin lipopeptide antifungal metabolites. Eukaryot. Cell 2015, 14, 698–718. [Google Scholar] [CrossRef] [PubMed]
- Morikawa, H.; Tomishima, M.; Kayakiri, N.; Araki, T.; Barrett, D.; Akamatsu, S.; Matsumoto, S.; Uchida, S.; Nakai, T.; Takeda, S.; et al. Synthesis and antifungal activity of ASP9726, a novel echinocandin with potent Aspergillus hyphal growth inhibition. Bioorg. Med. Chem. Lett. 2014, 24, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Ueda, S.; Sakamoto, K.; Oohata, N.; Tsuboi, M.; Yamashita, M.; Hino, M.; Yamada, M.; Hashimoto, S. Screening and characterization of microorganisms with FR901379 acylase activity. J. Antibiot. 2010, 63, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Balkovec, J.M. Section review: Anti-infectives: Lipopeptide antifungal agents. Expert Opin. Investig. Drugs 1994, 3, 65–82. [Google Scholar] [CrossRef]
- Tomishima, M.; Ohki, H.; Yamada, A.; Maki, K.; Ikeda, F. Novel echinocandin antifungals. Part 1: Novel side-chain analogs of the natural product FR901379. Bioorg. Med. Chem. Lett. 2008, 18, 1474–1477. [Google Scholar] [CrossRef] [PubMed]
- Tomishima, M.; Ohki, H.; Yamada, A.; Maki, K.; Ikeda, F. Novel echinocandin antifungals. Part 2: Optimization of the side chain of the natural product FR901379. Discovery of micafungin. Bioorg. Med. Chem. Lett. 2008, 18, 2886–2890. [Google Scholar] [CrossRef] [PubMed]
- Hino, M.; Fujie, A.; Iwamoto, T.; Hori, Y.; Hashimoto, M.; Tsurumi, Y.; Sakamoto, K.; Takase, S.; Hashimoto, S. Chemical diversity in lipopeptide antifungal antibiotics. J. Ind. Microbiol. Biotechnol. 2001, 27, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, S. Micafungin: A sulfated echinocandin. J. Antibiot. 2009, 62, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Hof, H.; Dietz, A. Antifungal activity of anidulafungin, a product of Aspergillus nidulans, against Aspergillus nidulans. Int. J. Antimicrob. Agents 2009, 33, 285–286. [Google Scholar] [CrossRef] [PubMed]
- Barrett, D. From natural products to clinically useful antifungals. Biochim. Biophys. Acta 2002, 1587, 224–233. [Google Scholar] [CrossRef]
- Turner, M.S.; Drew, R.H.; Perfect, J.R. Emerging echinocandins for treatment of invasive fungal infections. Expert Opin. Emerg. Drugs 2006, 11, 231–250. [Google Scholar] [CrossRef]
- Balkovec, J.M.; Hughes, D.L.; Masurekar, P.S.; Sable, C.A.; Schwartz, R.E.; Singh, S.B. Discovery and development of first in class antifungal caspofungin (CANCIDAS®)—A case study. Nat. Prod. Rep. 2014, 31, 15–34. [Google Scholar] [CrossRef] [PubMed]
- Eschenauer, G.; Depestel, D.D.; Carver, P.L. Comparison of echinocandin antifungals. Ther. Clin. Risk Manag. 2007, 3, 71–97. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; Kim, H.G.; Long, L. Efficacy of aminocandin in the treatment of immunocompetent mice with haematogenously disseminated fluconazole-resistant candidiasis. J. Antimicrob. Chemother. 2007, 59, 556–559. [Google Scholar] [PubMed]
- Warn, P.A.; Sharp, A.; Morrissey, G.; Denning, D.W. Activity of aminocandin (IP960; HMR3270) compared with amphotericin B, itraconazole, caspofungin and micafungin in neutropenic murine models of disseminated infection caused by itraconazole-susceptible and -resistant strains of Aspergillus fumigatus. Int. J. Antimicrob. Agents 2009, 35, 146–151. [Google Scholar] [PubMed]
- Guembe, M.; Guinea, J.; Marcos-Zambrano, L.J.; Fernández-Cruz, A.; Peláez, T.; Muñoz, P.; Bouza, E. Micafungin at physiological serum concentrations shows antifungal activity against Candida albicans and Candida parapsilosis biofilms. Antimicrob. Agents Chemother. 2014, 58, 5581–5584. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Zambrano, L.J.; Escribano, P.; González del Vecchio, M.; Bouza, E.; Guinea, J. Micafungin is more active against Candida albicans biofilms with high metabolic activity. J. Antimicrob. Chemother. 2014, 69, 2984–2987. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Zambrano, L.J.; Escribano, P.; Bouza, E.; Guinea, J. Comparison of the antifungal activity of micafungin and amphotericin B against Candida tropicalis biofilms. J. Antimicrob. Chemother. 2016. [Google Scholar] [CrossRef] [PubMed]
- Simitsopoulou, M.; Kyrpitzi, D.; Velegraki, A.; Walsh, T.J.; Roilides, E. Caspofungin at catheter lock concentrations eradicates mature biofilms of Candida lusitaniae and Candida guilliermondii. Antimicrob. Agents Chemother. 2014, 58, 4953–4956. [Google Scholar] [CrossRef] [PubMed]
- Maiolo, E.M.; Oliva, A.; Furustrand Tafin, U.; Perrotet, N.; Borens, O.; Trampuz, A. Antifungal activity against planktonic and biofilm Candida albicans in an experimental model of foreign-body infection. J. Infect. 2016, 72, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Rosato, A.; Catalano, A.; Carocci, A.; Carrieri, A.; Carone, A.; Caggiano, G.; Franchini, C.; Corbo, F.; Montagna, M.T. In vitro interactions between anidulafungin and nonsteroidal anti-inflammatory drugs on biofilms of Candida spp. Bioorg. Med. Chem. 2016, 24, 1002–1005. [Google Scholar] [CrossRef] [PubMed]
- Kovács, R.; Bozó, A.; Gesztelyi, R.; Domán, M.; Kardos, G.; Nagy, F.; Tóth, Z.; Majoros, L. Effect of caspofungin and micafungin in combination with farnesol against Candida parapsilosis biofilms. Int. J. Antimicrob. Agents 2016, 47, 304–310. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Perlin, D.S. Echinocandin resistance: An emerging clinical problem? Curr. Opin. Infect. Dis. 2014, 27, 484–492. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Lamoth, F.; Juvvadi, P.R. Potential microbiological effects of higher dosing of echinocandins. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S669–S677. [Google Scholar] [CrossRef] [PubMed]
- Walraven, C.J.; Bernardo, S.M.; Wiederhold, N.P.; Lee, S.A. Paradoxical antifungal activity and structural observations in biofilms formed by echinocandin-resistant Candida albicans clinical isolates. Med. Mycol. 2014, 52, 131–139. [Google Scholar] [CrossRef] [PubMed]
- Alexander, B.D.; Johnson, M.D.; Pfeiffer, C.D.; Jimenez-Ortigosa, C.; Catania, J.; Booker, R.; Castanheira, M.; Messer, S.A.; Perlin, D.S.; Pfaller, M.A. Increasing echinocandin resistance in Candida glabrata: Clinical failure correlates with presence of FKS mutations and elevated minimum inhibitory concentrations. Clin. Infect. Dis. 2013, 56, 1724–1732. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Ohno, H.; Imamura, Y.; Kohno, S.; Miyazaki, Y. The effects of an Hsp90 inhibitor on the paradoxical effect. Jpn. J. Infect. Dis. 2009, 62, 392–393. [Google Scholar] [PubMed]
- Nett, J.E.; Andes, D. Fungal biofilms: In vivo models for discovery of anti-biofilm drugs. Microbiol. Spectr. 2015, 3, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.J.; Azie, N.; Andes, D.R. Development of new strategies for echinocandins: Progress in translational research. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S601–S603. [Google Scholar] [CrossRef] [PubMed]
- Fiori, B.; Posteraro, B.; Torelli, R.; Tumbarello, M.; Perlin, D.S.; Fadda, G.; Sanguinetti, M. In vitro activities of anidulafungin and other antifungal agents against biofilms formed by clinical isolates of different Candida and Aspergillus species. Antimicrob. Agents Chemother. 2011, 55, 3031–3035. [Google Scholar] [CrossRef] [PubMed]
- Rajendran, R.; Williams, C.; Lappin, D.F.; Millington, O.; Martins, M.; Ramage, G. Extracellular DNA release acts as an antifungal resistance mechanism in mature Aspergillus fumigatus biofilms. Eukaryot. Cell 2013, 12, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, R.A.; Teixeira, C.E.C.; Brilhante, R.S.N.; Castelo-Branco, D.S.C.M.; Paiva, M.A.N.; Giffoni Leite, J.J.; Lima, D.T.; Monteiro, A.J.; Sidrim, J.J.C.; Rocha, M.F.G. Minimum inhibitory concentrations of amphotericin B, azoles and caspofungin against Candida species are reduced by farnesol. Med. Mycol. 2013, 51, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, J.A.G.; Carr, J.H.; Starling, C.E.F.; De Resende, M.A.; Donlan, R.M. Biofilm formation and effect of caspofungin on biofilm structure of Candida species bloodstream isolates. Antimicrob. Agents Chemother. 2009, 53, 4377–4384. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.J.; Piper, K.E.; Nguyen, G.; Steckelberg, J.M.; Patel, R. In vitro activity of anidulafungin against Candida albicans biofilms. Antimicrob. Agents Chemother. 2008, 52, 2242–2243. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.J.; Steckelberg, K.E.; Piper, K.E.; Steckelberg, J.M.; Patel, R. In vitro activity of micafungin against planktonic and sessile Candida albicans isolates. Antimicrob. Agents Chemother. 2009, 53, 2638–2639. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Miyagawa, S.; Takeda, O.; Hakariya, M.; Matsumoto, S.; Ohno, H.; Miyazaki, Y. Real-time microscopic observation of Candida biofilm development and effects due to micafungin and fluconazole. Antimicrob. Agents Chemother. 2013, 57, 2226–2230. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, D.M.; George, T.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Antifungal susceptibility of Candida biofilms: Unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrob. Agents Chemother. 2002, 46, 1773–1780. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Zambrano, L.J.; Escribano, P.; Bouza, E.; Guinea, J. Susceptibility of Candida albicans biofilms to caspofungin and anidulafungin is not affected by metabolic activity or biomass production. Med. Mycol. 2016, 54, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.S.; Colombo, A.L.; Arthington-Skaggs, B.A. Paradoxical growth effect of caspofungin observed on biofilms and planktonic cells of five different Candida species. Antimicrob. Agents Chemother. 2007, 51, 3081–3088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajendran, R.; Sherry, L.; Nile, C.J.; Sherriff, A.; Johnson, E.M.; Hanson, M.F.; Williams, C.; Munro, C.A.; Jones, B.J.; Ramage, G. Biofilm formation is a risk factor for mortality in patients with Candida albicans bloodstream infection-Scotland, 2012–2013. Clin. Microbiol. Infect. 2016, 22, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirazi, F.; Kontoyiannis, D.P. Micafungin triggers caspase-dependent apoptosis in Candida albicans and Candida parapsilosis biofilms, including caspofungin non-susceptible isolates. Virulence 2015, 6, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Shuford, J.A.; Piper, K.E.; Steckelberg, J.M.; Patel, R. In vitro biofilm characterization and activity of antifungal agents alone and in combination against sessile and planktonic clinical Candida albicans isolates. Diagn. Microbiol. Infect. Dis. 2007, 57, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Simitsopoulou, M.; Peshkova, P.; Tasina, E.; Katragkou, A.; Kyrpitzi, D.; Velegraki, A.; Walsh, T.J.; Roilides, E. Species-specific and drug-specific differences in susceptibility of Candida biofilms to echinocandins: Characterization of less common bloodstream isolates. Antimicrob. Agents Chemother. 2013, 57, 2562–2570. [Google Scholar] [CrossRef] [PubMed]
- Di Bonaventura, G.; Spedicato, I.; Picciani, C.; D’Antonio, D.; Piccolomini, R. In vitro pharmacodynamic characteristics of amphotericin B, caspofungin, fluconazole, and voriconazole against bloodstream isolates of infrequent Candida species from patients with hematologic malignancies. Antimicrob. Agents Chemother. 2004, 48, 4453–4456. [Google Scholar] [CrossRef] [PubMed]
- Shanmughapriya, S.; Sornakumari, H.; Lency, A.; Kavitha, S.; Natarajaseenivasan, K. Synergistic effect of amphotericin B and tyrosol on biofilm formed by Candida krusei and Candida tropicalis from intrauterine device users. Med. Mycol. 2014, 52, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Valentin, A.; Canton, E.; Peman, J.; Quindos, G. In vitro activity of amphotericin B and anidulafungin against Candida spp. biofilms. Rev. Iberoam. Micol. 2007, 24, 272–277. [Google Scholar] [PubMed]
- Liao, Y.; Yang, S.; Cong, L.; Lu, X.; Ao, J.; Yang, R. In vitro activities of antifungal combinations against biofilms and planktonic forms of clinical Trichosporon asahii isolates. Antimicrob. Agents Chemother. 2014, 58, 7615–7616. [Google Scholar] [CrossRef] [PubMed]
- Bazzi, W.; Sabra, A.; Zahreddine, L.; Khairallah, M.T.; Baroud, M.; Hadi, U.; Matar, G.M. The inhibitory effect of micafungin on biofilm formation by Pseudomonas aeruginosa. Biofouling 2013, 29, 909–915. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.; Roilides, E.; Katragkou, A.; Petraitis, V.; Walsh, T.J. The role of echinocandins in Candida biofilm-related vascular catheter infections: In vitro and in vivo model systems. Clin. Infect. Dis. 2015, 61 (Suppl. S6), S618–S621. [Google Scholar] [CrossRef] [PubMed]
- Shuford, J.A.; Rouse, M.S.; Piper, K.E.; Steckelberg, J.M.; Patel, R. Evaluation of caspofungin and amphotericin B deoxycholate against Candida albicans biofilms in an experimental intravascular catheter infection model. J. Infect. Dis. 2006, 194, 710–713. [Google Scholar] [CrossRef] [PubMed]
- Bink, A.; Kucharíková, S.; Neirinck, B.; Vleugels, J.; Van Dijck, P.; Cammue, B.P.A.; Thevissen, K. The nonsteroidal antiinflammatory drug diclofenac potentiates the in vivo activity of caspofungin against Candida albicans biofilms. J. Infect. Dis. 2012, 206, 1790–1797. [Google Scholar] [CrossRef] [PubMed]
- Kucharikova, S.; Tournu, H.; Holtappels, M.; Van Dijck, P.; Lagrou, K. In vivo efficacy of anidulafungin against mature Candida albicans biofilms in a novel rat model of catheter-associated candidiasis. Antimicrob. Agents Chemother. 2010, 54, 4474–4475. [Google Scholar] [CrossRef] [PubMed]
- Kucharikova, S.; Sharma, N.; Spriet, I.; Maertens, J.; Van Dijck, P.; Lagrou, K. Activities of systemically administered echinocandins against in vivo mature Candida albicans biofilms developed in a rat subcutaneous model. Antimicrob. Agents Chemother. 2013, 57, 2365–2368. [Google Scholar] [CrossRef] [PubMed]
- Kucharikova, S.; Neirinck, B.; Sharma, N.; Vleugels, J.; Lagrou, K.; Van Dijck, P. In vivo Candida glabrata biofilm development on foreign bodies in a rat subcutaneous model. J. Antimicrob. Chemother. 2015, 70, 846856. [Google Scholar] [CrossRef] [PubMed]
- Jeon, J.-G.; Pandit, S.; Xiao, J.; Gregoire, S.; Falsetta, M.L.; Klein, M.I.; Koo, H. Influences of trans-trans farnesol, a membrane-targeting sesquiterpenoid, on Streptococcus mutans physiology and survival within mixed-species oral biofilms. Int. J. Oral Sci. 2011, 3, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Chifiriuc, M.C.; Ditu, L.M.; Oprea, E.; Litescu, S.; Bucur, M.; Marutescu, L.; Enache, G.; Saviuc, C.; Burlibasa, M.; Traistaru, T.; et al. In vitro study of the inhibitory activity of usnic acid on dental plaque biofilm. Roum. Arch. Microbiol. Immunol. 2009, 68, 215–222. [Google Scholar] [PubMed]
- Cirasola, D.; Sciota, R.; Vizzini, L.; Ricucci, V.; Morace, G.; Borghi, E. Experimental biofilm-related Candida infections. Future Microbiol. 2013, 8, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Cateau, E.; Rodier, M.H.; Imbert, C. In vitro efficacies of caspofungin or micafungin catheter lock solutions on Candida albicans biofilm growth. J. Antimicrob. Chemother. 2008, 62, 153–155. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.P.; Samples, M.L.; Mercier, R.C.; Spilde, M.N. Activities and ultrastructural effects of antifungal combinations against simulated Candida endocardial vegetations. Antimicrob. Agents Chemother. 2008, 52, 2367–2376. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.P. Antifungal combinations against simulated Candida albicans endocardial vegetations. Antimicrob. Agents Chemother. 2009, 53, 2629–2631. [Google Scholar] [CrossRef] [PubMed]
- Serefko, A.; Chudzik, B.; Malm, A. In vitro activity of caspofungin against planktonic and sessile Candida sp. cells. Pol. J. Microbiol. 2006, 55, 133–137. [Google Scholar] [PubMed]
- Lazzell, A.L.; Chaturvedi, A.K.; Pierce, C.G.; Prasad, D.; Uppuluri, P.; Lopez-Ribot, J.L. Treatment and prevention of Candida albicans biofilms with caspofungin in a novel central venous catheter murine model of candidiasis. J. Antimicrob. Chemother. 2009, 64, 567–570. [Google Scholar] [CrossRef] [PubMed]
- Pammi, M.; Liang, R.; Hicks, J.M.; Barrish, J.; Versalovic, J. Farnesol decreases biofilms of Staphylococcus epidermidis and exhibits synergy with nafcillin and vancomycin. Pediatr. Res. 2011, 70, 578–583. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.W.; Chen, X.; Lim, J.; Kim, S.H.; Kim, S.T.; Cho, Y.H.; Yoon, J.; Park, S. In vivo fluorescence imaging of bacteriogenic cyanide in the lungs of live mice infected with cystic fibrosis pathogens. PLoS ONE 2011, 6, e21387. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Dinakaran, H.; Thomas, D.P.; Chaturvedi, A.K.; Lopez-Ribot, J.L. Characteristics of Candida albicans biofilms grown in a synthetic urine medium. J. Clin. Microbiol. 2009, 47, 4078–4083. [Google Scholar] [CrossRef] [PubMed]
- De Cremer, K.; Staes, I.; Delattin, N.; Cammue, B.P.A.; Thevissen, K.; De Brucker, K. Combinatorial drug approaches to tackle Candida albicans biofilms. Expert Rev. Anti-Infect. Ther. 2015, 13, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Griesser, S.S.; Jasieniak, M.; Coad, B.R.; Griesser, H.J. Antifungal coatings by caspofungin immobilization onto biomaterials surfaces via a plasma polymer interlayer. Biointerphases 2015, 10, 04A307. [Google Scholar] [CrossRef] [PubMed]
- Kucharikova, S.; Gerits, E.; De Brucker, K.; Braem, A.; Ceh, K.; Majdic, G.; Spanic, T.; Pogorevc, E.; Verstraeten, N.; Tournu, H.; et al. Covalent immobilization of antimicrobial agents on titanium prevents Staphylococcus aureus and Candida albicans colonization and biofilm formation. J. Antimicrob. Chemother. 2016, 71, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Rawson, M.; Haggard, W.; Jennings, J.A. Osteocompatibility of biofilm inhibitors. Open Orthop. J. 2014, 8, 442–449. [Google Scholar] [PubMed]
- Grumezescu, V.; Holban, A.M.; Grumezescu, A.M.; Socol, G.; Ficai, A.; Vasile, B.S.; Trusca, R.; Bleotu, C.; Lazar, V.; Chifiriuc, C.M.; et al. Usnic acid-loaded biocompatible magnetic PLGA-PVA microsphere thin films fabricated by MAPLE with increased resistance to Staphylococcal colonization. Biofabrication 2014, 6, 35002. [Google Scholar] [CrossRef] [PubMed]
- Guzun, A.S.; Stroescu, M.; Jinga, S.I.; Voicu, G.; Grumezescu, A.M.; Holban, A.M. Plackett-Burman experimental design for bacterial cellulose-silica composites synthesis. Mater. Sci. Eng. C. Mater. Biol. Appl. 2014, 42, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Jamil, B.; Habib, H.; Abbasi, S.A.; Ihsan, A.; Nasir, H.; Imran, M. Development of cefotaxime impregnated chitosan as nano-antibiotics: De novo strategy to combat biofilm forming multi-drug resistant pathogens. Front. Microbiol. 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Horev, B.; Klein, M.I.; Hwang, G.; Li, Y.; Kim, D.; Koo, H.; Benoit, D.S.W. pH-Activated nanoparticles for controlled topical delivery of farnesol to disrupt oral biofilm virulence. ACS Nano 2015, 9, 2390–2404. [Google Scholar] [CrossRef] [PubMed]
- Martinelli, A.; Bakry, A.; D’Ilario, L.; Francolini, I.; Piozzi, A.; Taresco, V. Release behavior and antibiofilm activity of usnic acid-loaded carboxylated poly(l-lactide) microparticles. Eur. J. Pharm. Biopharm. 2014, 88, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Taresco, V.; Francolini, I.; Padella, F.; Bellusci, M.; Boni, A.; Innocenti, C.; Martinelli, A.; D’Ilario, L.; Piozzi, A. Design and characterization of antimicrobial usnic acid loaded-core/shell magnetic nanoparticles. Mater. Sci. Eng. C. Mater. Biol. Appl. 2015, 52, 72–81. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, M. The fungi: 1, 2, 3 ... 5.1 million species? Am. J. Bot. 2011, 98, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Wisecaver, J.H.; Slot, J.C.; Rokas, A. The evolution of fungal metabolic pathways. PLoS Genet. 2014, 10, e1004816. [Google Scholar] [CrossRef] [PubMed]
- Borghi, E.; Morace, G.; Borgo, F.; Rajendran, R.; Sherry, L.; Nile, C.; Ramage, G. New strategic insights into managing fungal biofilms. Front. Microbiol. 2015, 6, 1077. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Molecules involved in quorum sensing in bacteria (1–5) and in fungi (6–8).
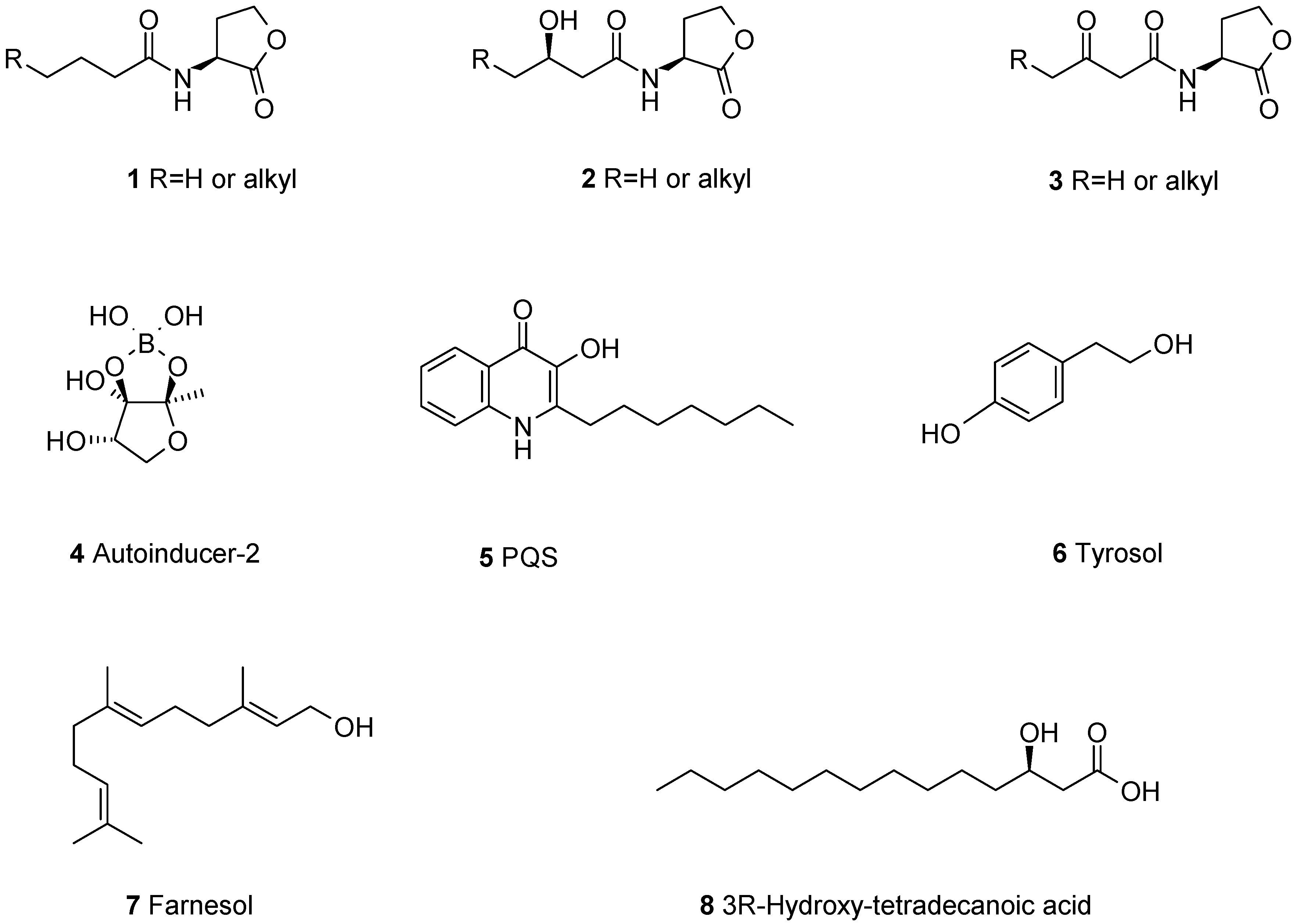
Figure 2.
Terpene-derived secondary metabolites from fungi reported to inhibit biofilms of bacteria or fungi, and related compounds.
Figure 2.
Terpene-derived secondary metabolites from fungi reported to inhibit biofilms of bacteria or fungi, and related compounds.

Figure 3.
Biofilm-controlling metabolites from fungi synthesized via a polyketide biosynthetic pathway.
Figure 3.
Biofilm-controlling metabolites from fungi synthesized via a polyketide biosynthetic pathway.

Figure 4.
Relatively complex secondary metabolites from fungi directed against biofilms of C. albicans.
Figure 4.
Relatively complex secondary metabolites from fungi directed against biofilms of C. albicans.
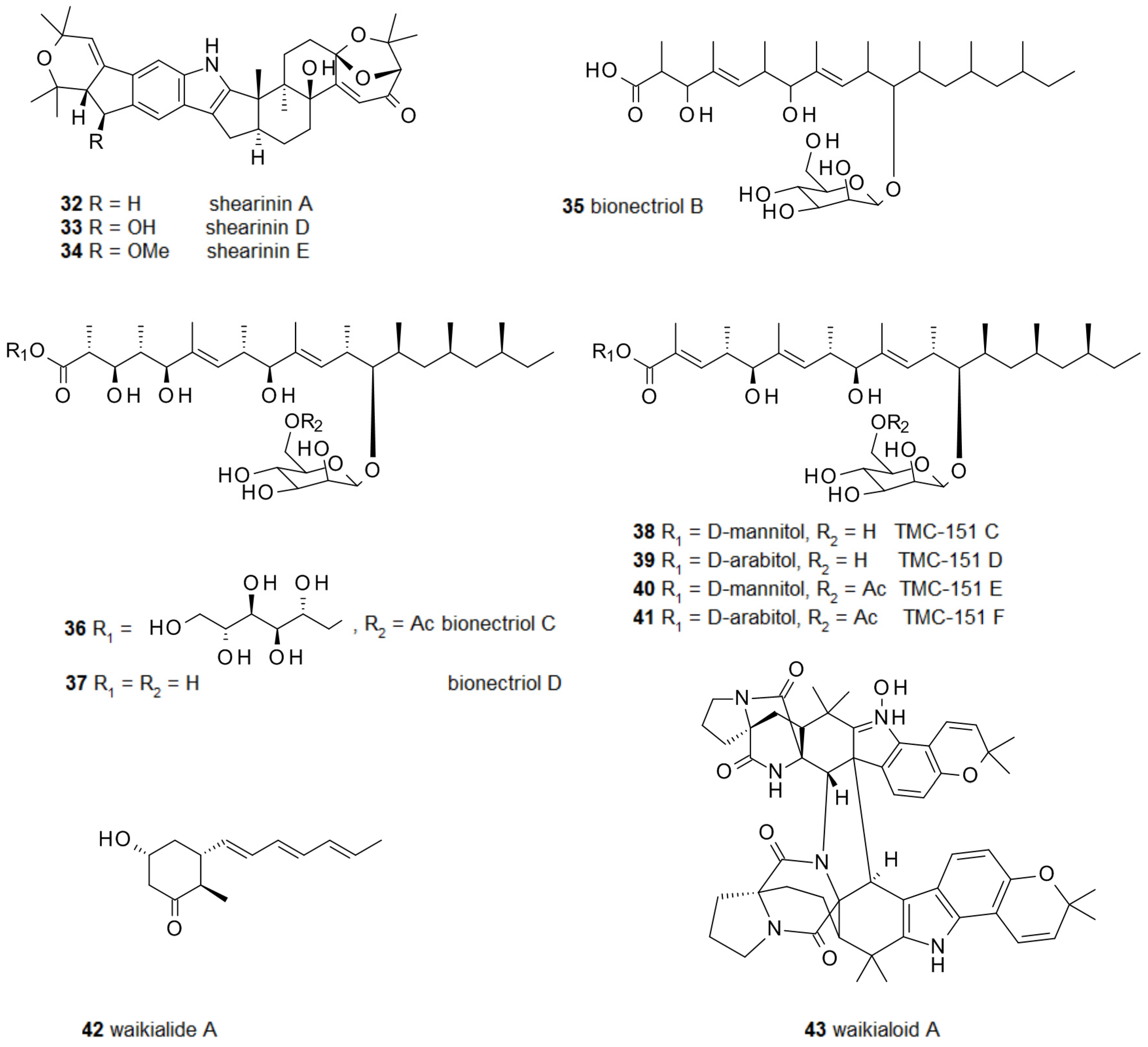
Figure 5.
Biofilm-controlling metabolites from fungi derived from amino acids.
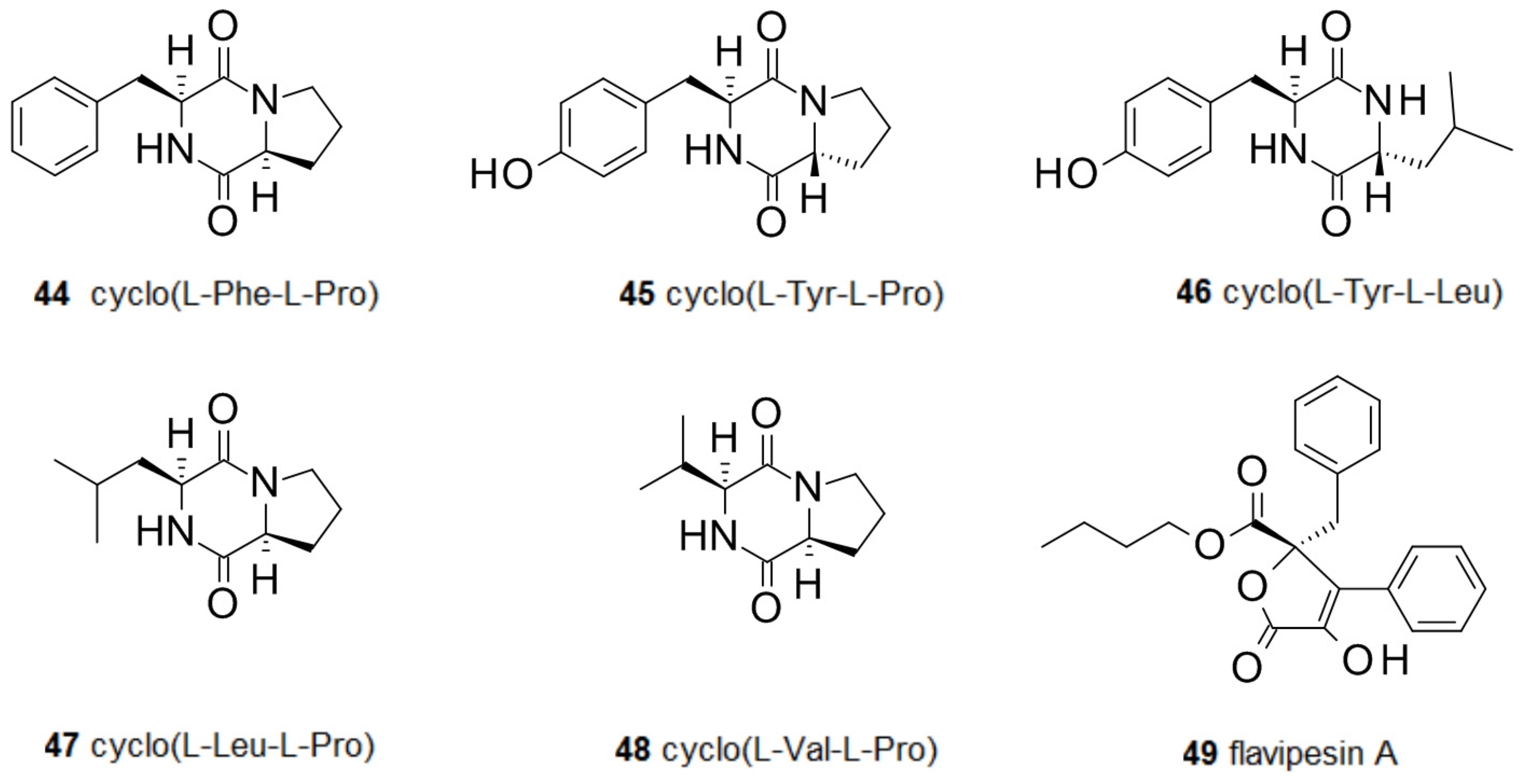
Figure 6.
Papulacandins and related antifungal compounds possessing a spiroketal moiety.
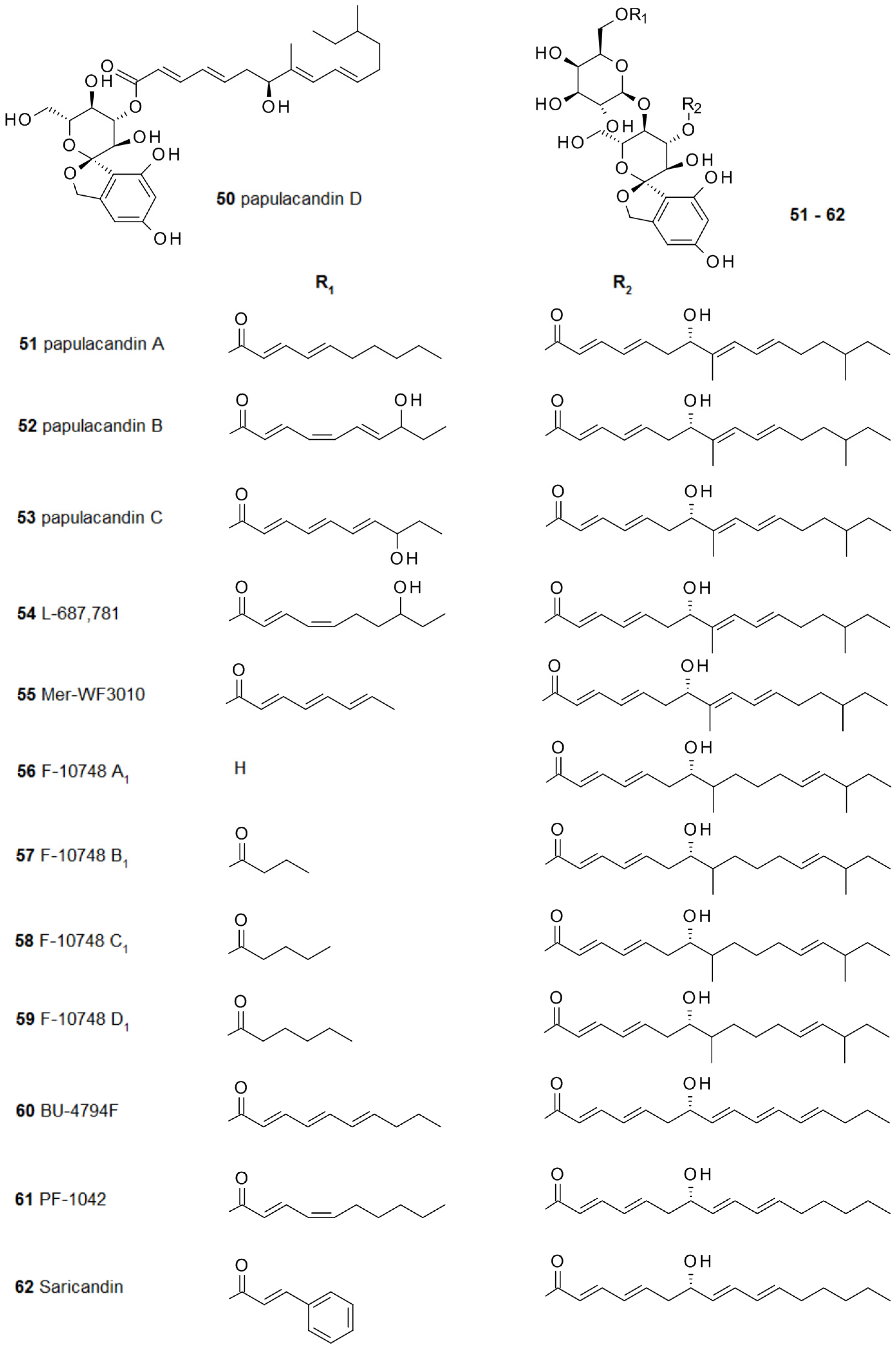
Figure 7.
Antifungal papulacandins and the galactosylated fusacandins lacking the spiroketal moiety.
Figure 7.
Antifungal papulacandins and the galactosylated fusacandins lacking the spiroketal moiety.

Figure 8.
Secondary metabolites of the echinocandin-type active against several fungal pathogens organized in biofilms. C14:0 = tetradecanoic acid, C16:0 = hexadecanoic acid, Me-14:0 = 12-methyl-tetradecanoic acid, diMe-14:0 = 10,12-dimethyl-tetradecanoic acid, diMe-16:0 = 12,14-dimethyl-hexadecanoic acid.
Figure 8.
Secondary metabolites of the echinocandin-type active against several fungal pathogens organized in biofilms. C14:0 = tetradecanoic acid, C16:0 = hexadecanoic acid, Me-14:0 = 12-methyl-tetradecanoic acid, diMe-14:0 = 10,12-dimethyl-tetradecanoic acid, diMe-16:0 = 12,14-dimethyl-hexadecanoic acid.
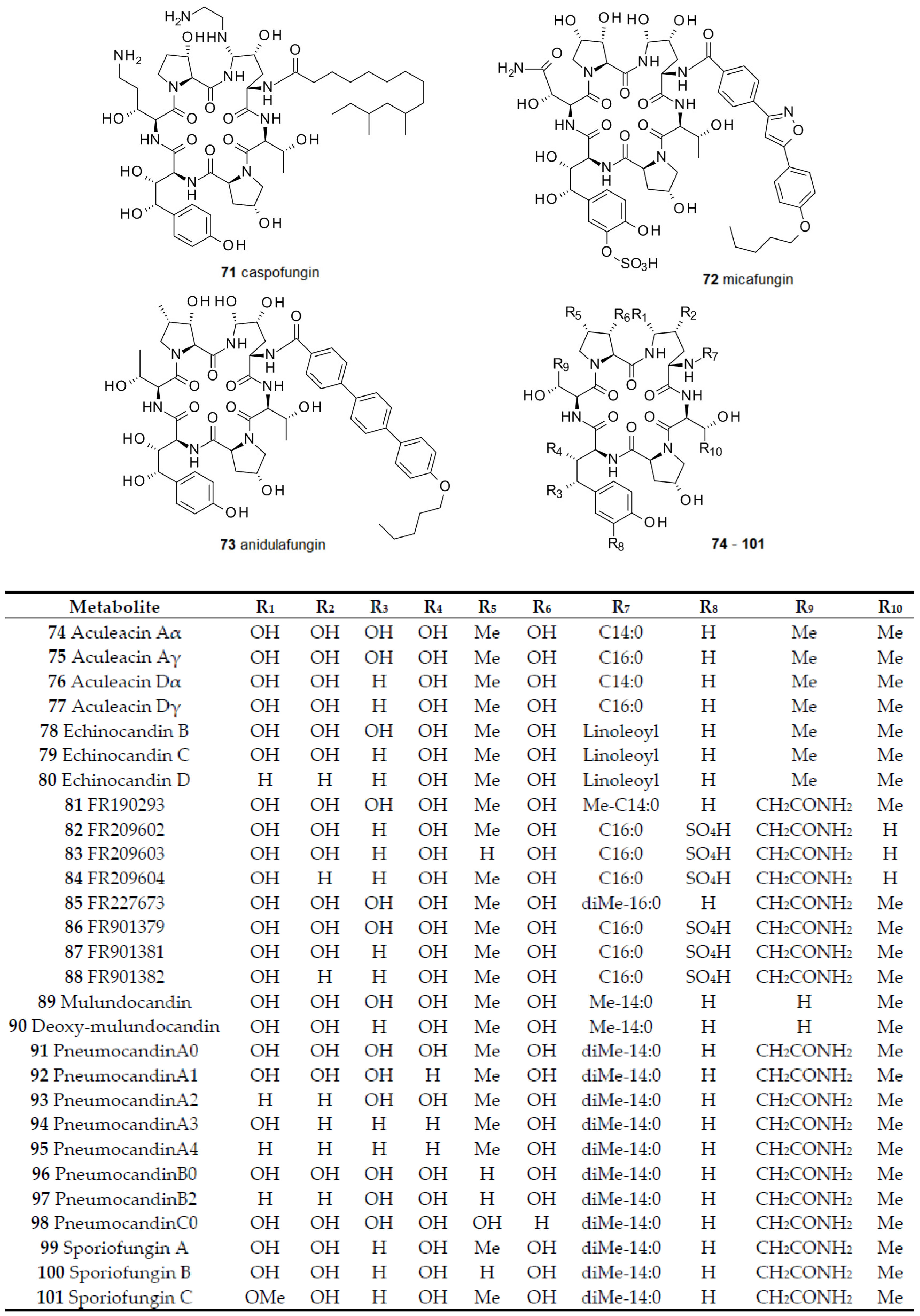
Figure 9.
Anti-biofilm mechanisms of fungal molecules against Gram positive bacteria, Gram negative bacteria and yeasts, with examples of active compounds. QS: quorum sensing; AHL: acyl homoserine lactone; AI-2: auto-inducer-2; agr: accessory gene regulator system; PQS: Pseudomonas quinolone signal; 3R hta: 3R-hydroxy-tetradecanoic acid.
Figure 9.
Anti-biofilm mechanisms of fungal molecules against Gram positive bacteria, Gram negative bacteria and yeasts, with examples of active compounds. QS: quorum sensing; AHL: acyl homoserine lactone; AI-2: auto-inducer-2; agr: accessory gene regulator system; PQS: Pseudomonas quinolone signal; 3R hta: 3R-hydroxy-tetradecanoic acid.
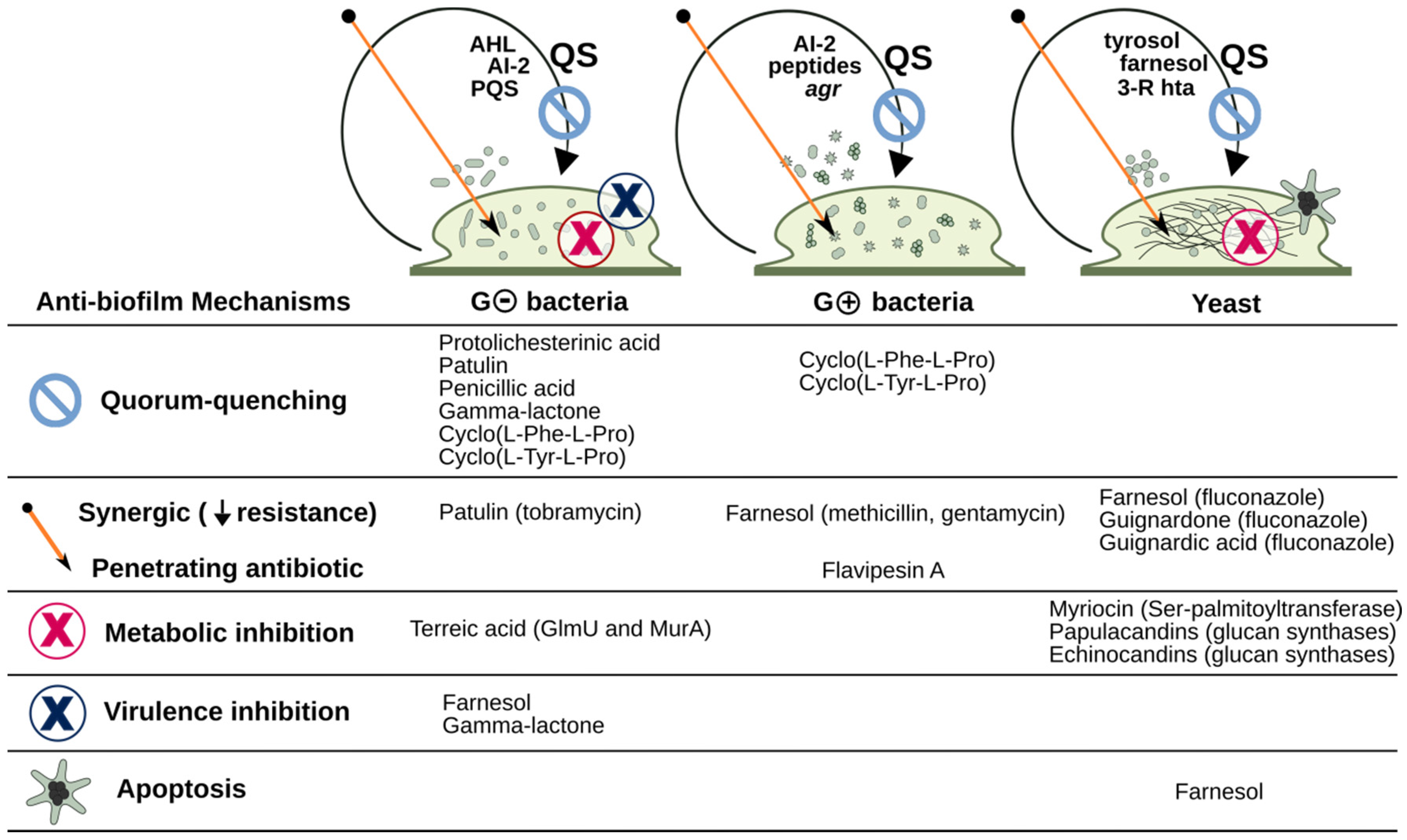

{kind=link}
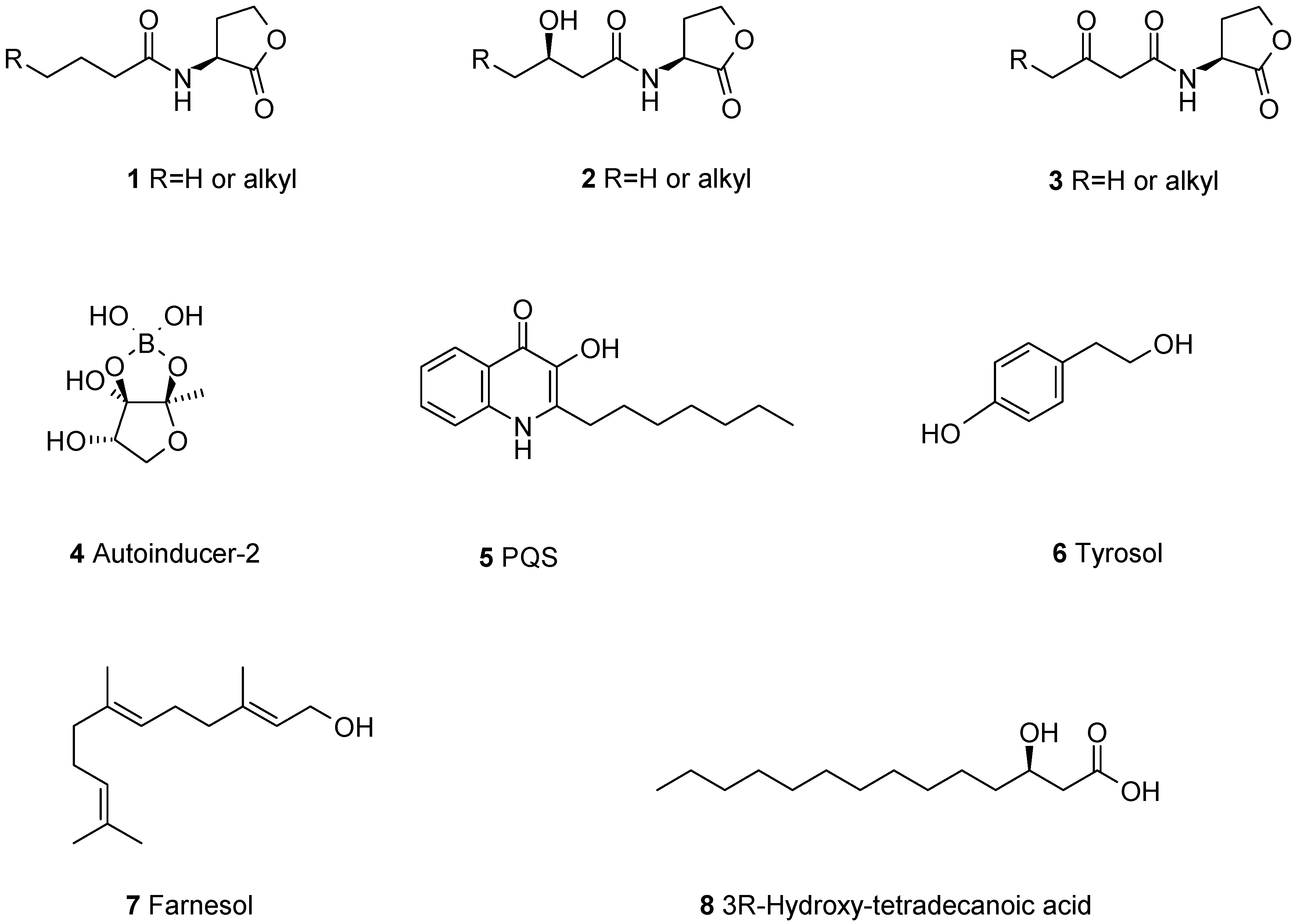{kind=link}
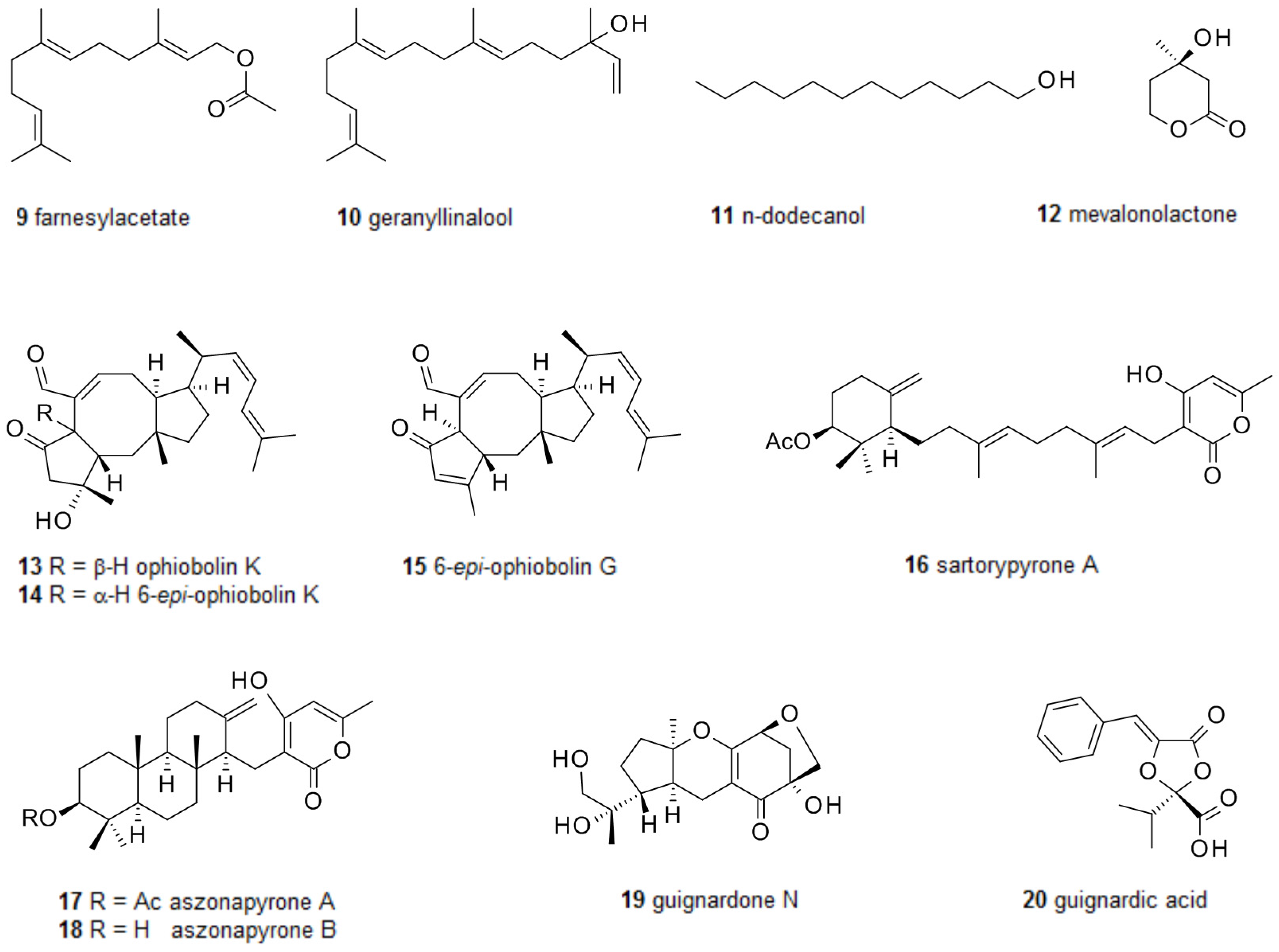{kind=link}
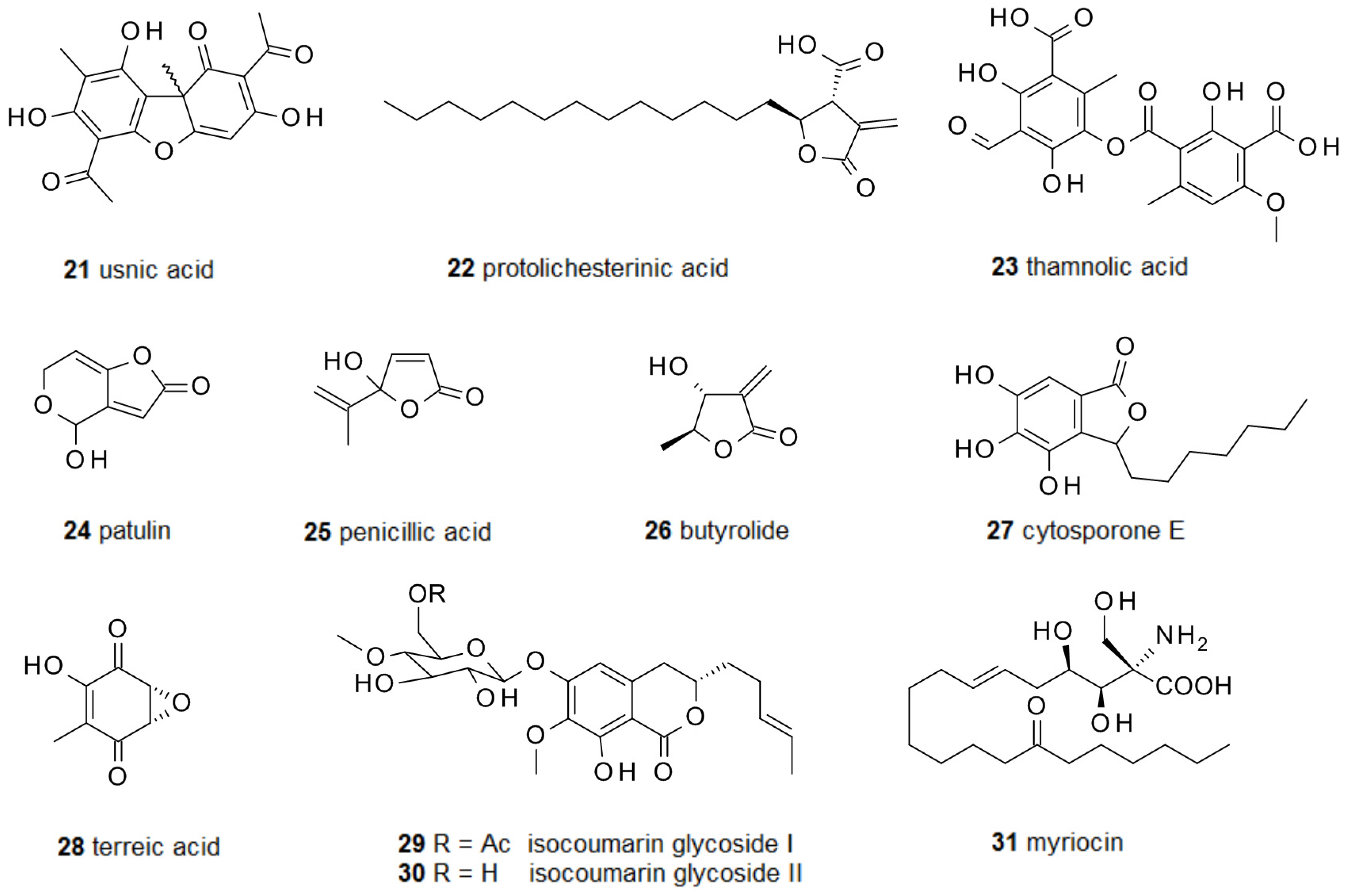{kind=link}
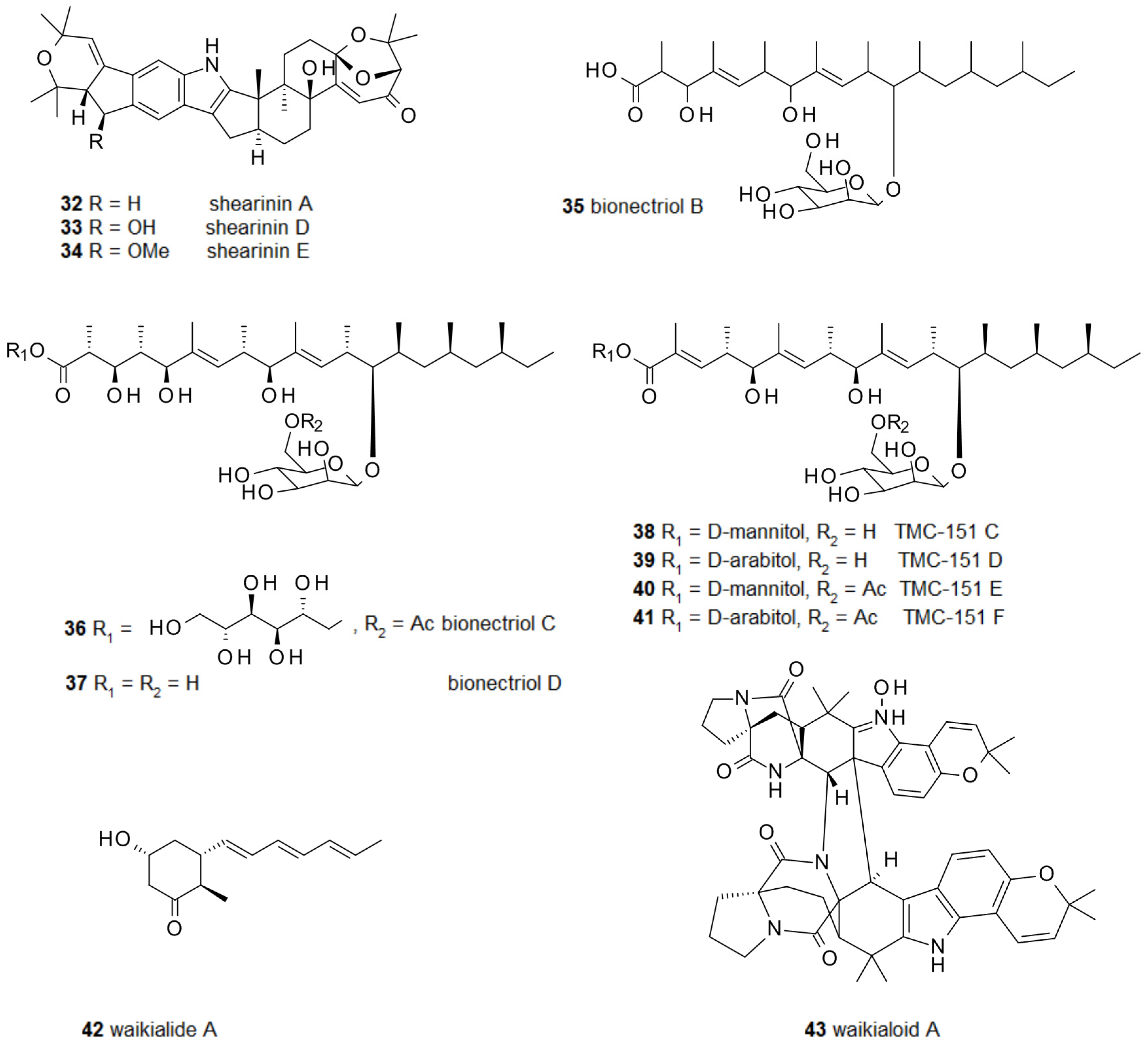{kind=link}
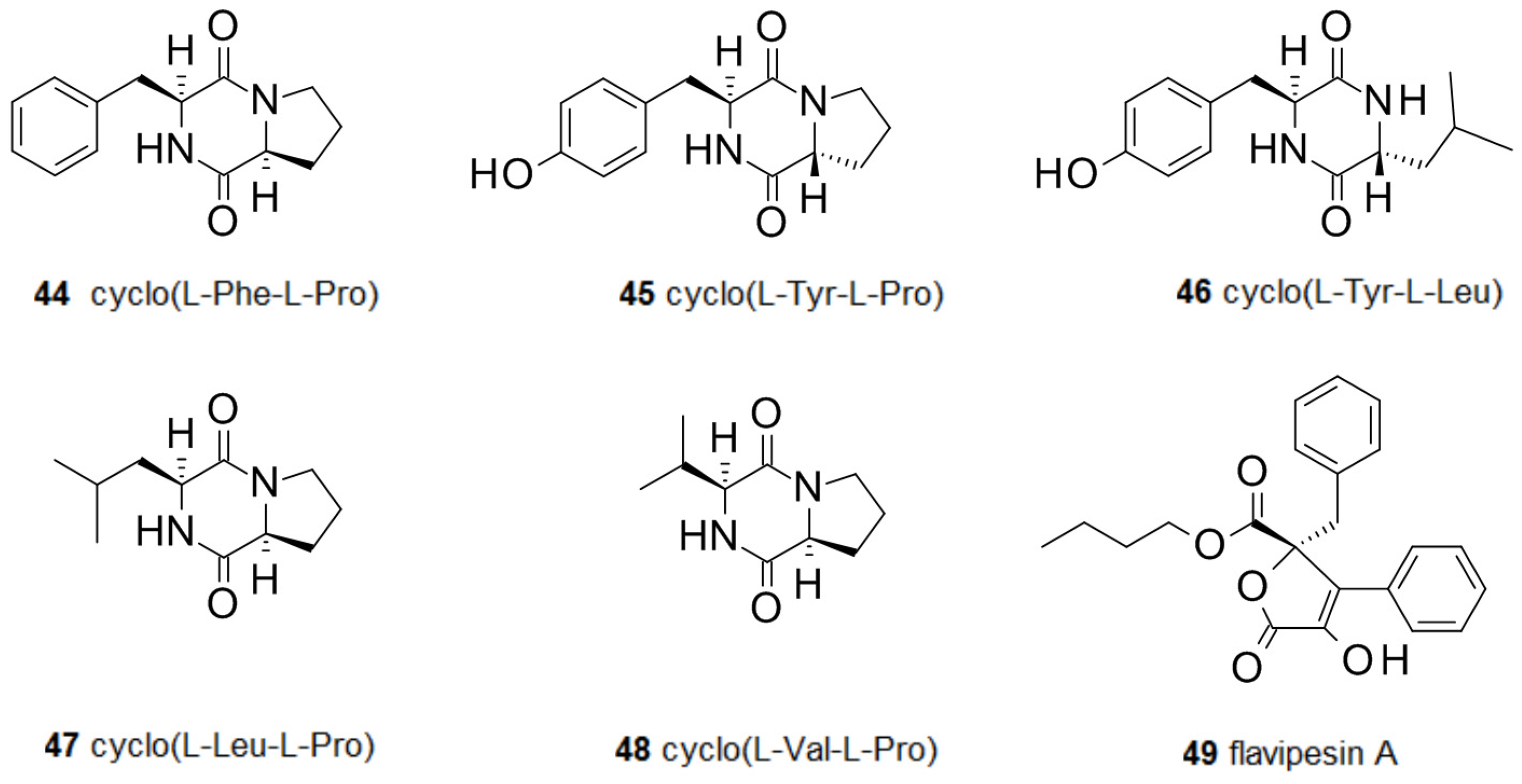{kind=link}
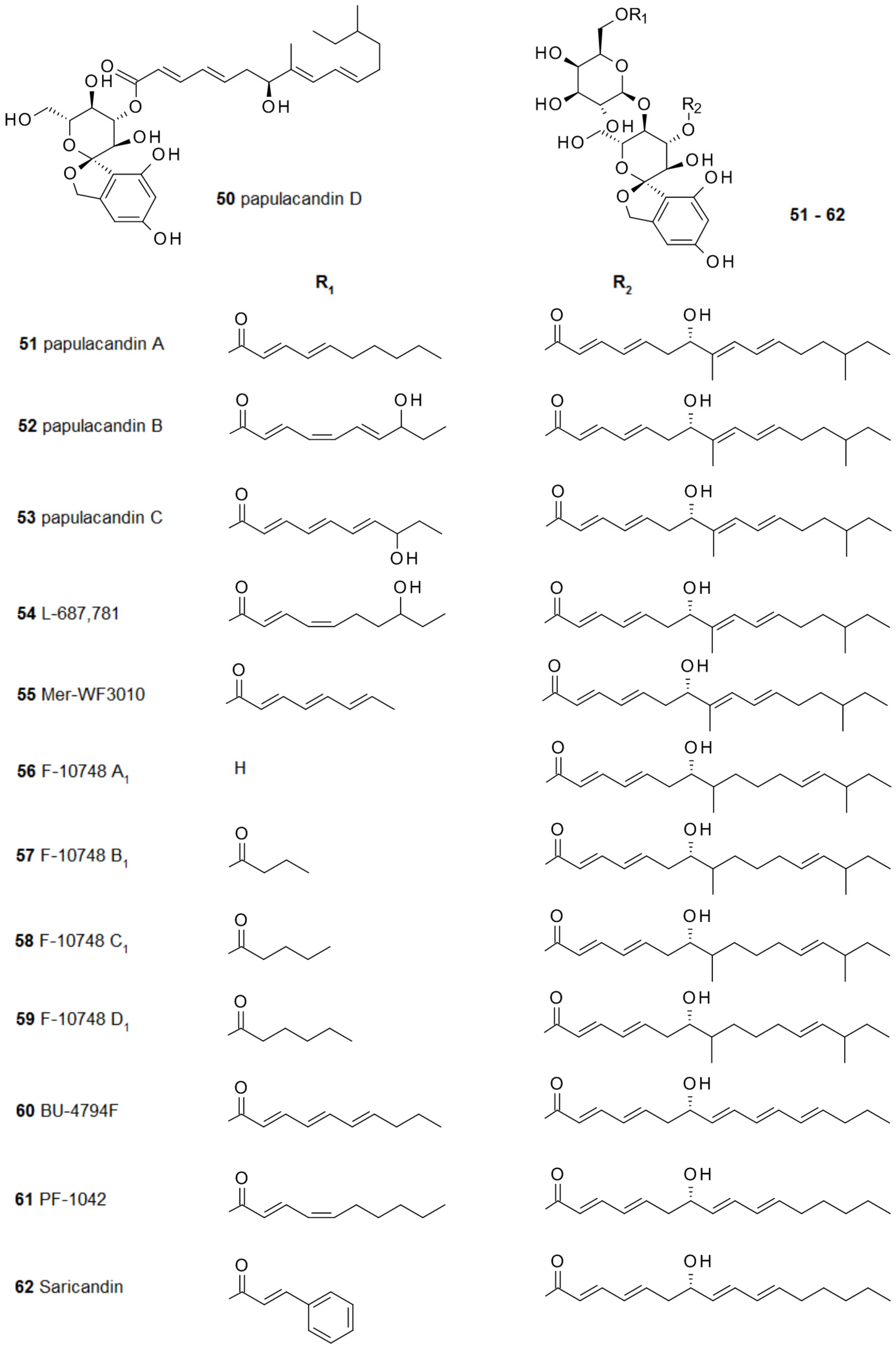{kind=link}
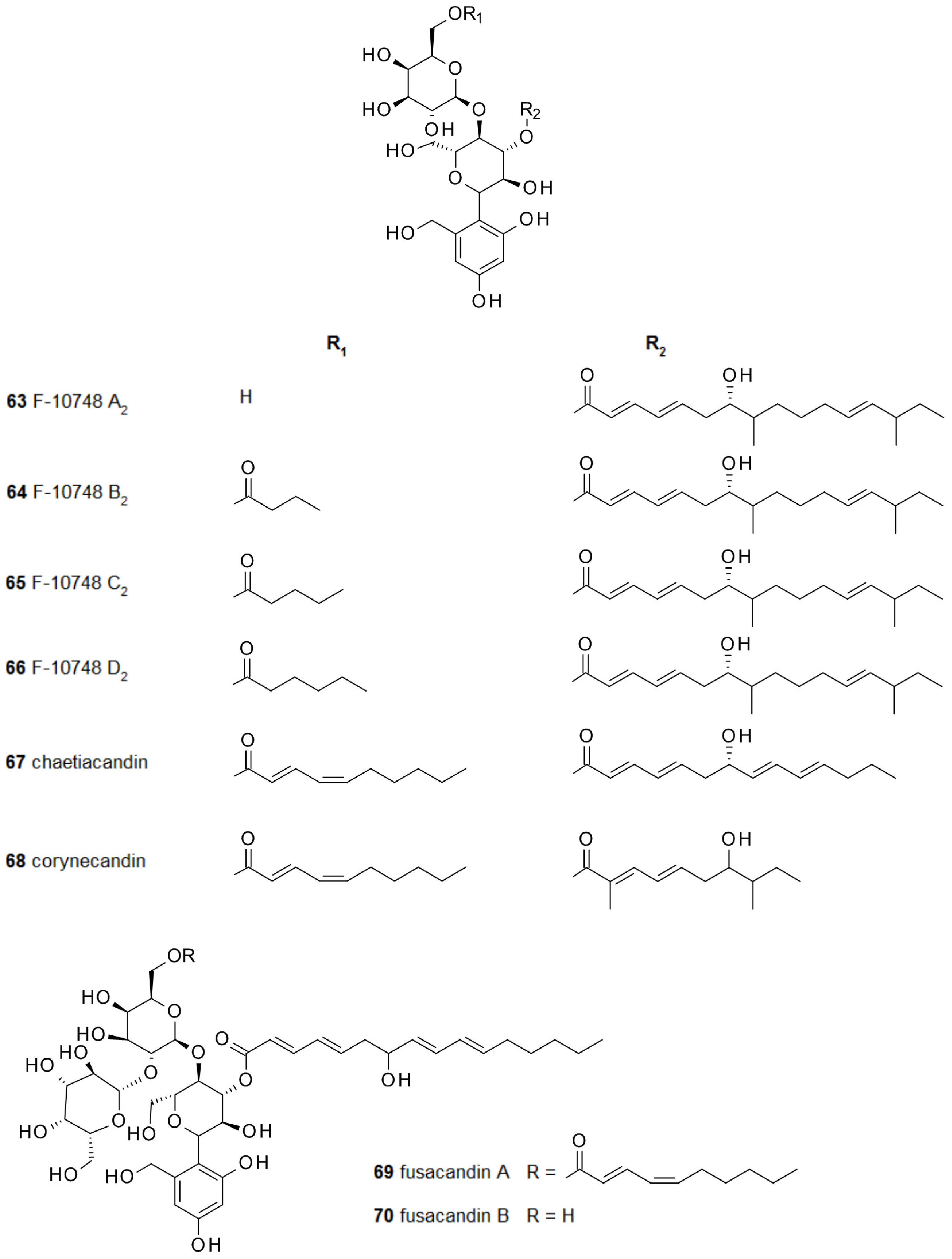{kind=link}
{kind=link}
{kind=link}
| Experimental Setup for Anti-Biofilm Tests | Target Organisms | Fungal Compounds and Combinations Tested | References |
|---|---|---|---|
| in vitro biofilm formation by clinical isolates | Aspergillus spp. | anid, casp, casp + DNase | [150,151] |
| Candida albicans | anid, casp, mica, farnesol, cas + voriconazole, cas + amb, shearinine, shearinine + amb | [137,150,152,153,154,155,156,157,158,159,160,161,162,163] | |
| Candida spp. (non-albicans) | anid, casp, mica, farnesol, casp + farnesol, mica + farnesol, tyrosol, tyrosol + amb, shearinine, shearinine + amb | [76,138,142,150,152,153,156,157,159,161,163,164,165,166] | |
| Trichosporon asahii | casp, casp + voriconazole | [167] | |
| Pseudomonas aeruginosa | mica | [168] | |
| Group A Streptococcus | usnic acid | [61] | |
| in vivo model of catheter biofilm in rabbit | C. albicans | casp, mica | [169,170] |
| in vivo model of subcutaneous device infection in rat | C. albicans | anid, casp, mica, casp + diclofenac | [171,172,173] |
| C. glabrata | anid, casp, mica | [174] | |
| mixed species oral biofilm, in vitro QS (quorum-sensing) interference in biofilm from dental plaque isolates | C. albicans, C. glabrata, S. mutans | tt-farnesol, tyrosol | [79,175] |
| Gram-positive bacteria | usnic acid | [176] | |
| in vivo model of biofilm infection by clinical isolates in Galleria mellonella larvae | C. albicans | anid | [177] |
| in vitro biofilms in catheters and biomaterials, simulated endocardial vegetation (SEV) | Candida spp. | casp, mica | [178,179,180,181] |
| Staphylococcus aureus, P. aeruginosa | usnic acid | [63] | |
| in vivo murine model of central venous catheter or subcutaneous catheter infection | C. albicans | casp | [182] |
| S. epidermidis | tt-farnesol | [183] | |
| in vivo model of foreign-body infection in guinea pig | C. albicans | casp, anid | [140] |
| in vivo murine model of cystic fibrosis | Burkholderia cepacia, P. aeruginosa | patulin | [184] |
| synthetic urine medium | C. albicans | casp | [185] |
Notes: anid: anidulafungin; casp: caspofungin; mica: micafungin; amb: amphotericin B; tt-farnesol: trans,trans-farnesol.
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Estrela, A.B.; Abraham, W.-R. Fungal Metabolites for the Control of Biofilm Infections. Agriculture 2016, 6, 37. https://doi.org/10.3390/agriculture6030037
AMA Style
Estrela AB, Abraham W-R. Fungal Metabolites for the Control of Biofilm Infections. Agriculture. 2016; 6(3):37. https://doi.org/10.3390/agriculture6030037
Chicago/Turabian StyleEstrela, Andréia Bergamo, and Wolf-Rainer Abraham. 2016. "Fungal Metabolites for the Control of Biofilm Infections" Agriculture 6, no. 3: 37. https://doi.org/10.3390/agriculture6030037
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.