
The Occurrence and Toxicity of Indospicine to Grazing Animals



{kind=link}
Abstract
:1. Introduction
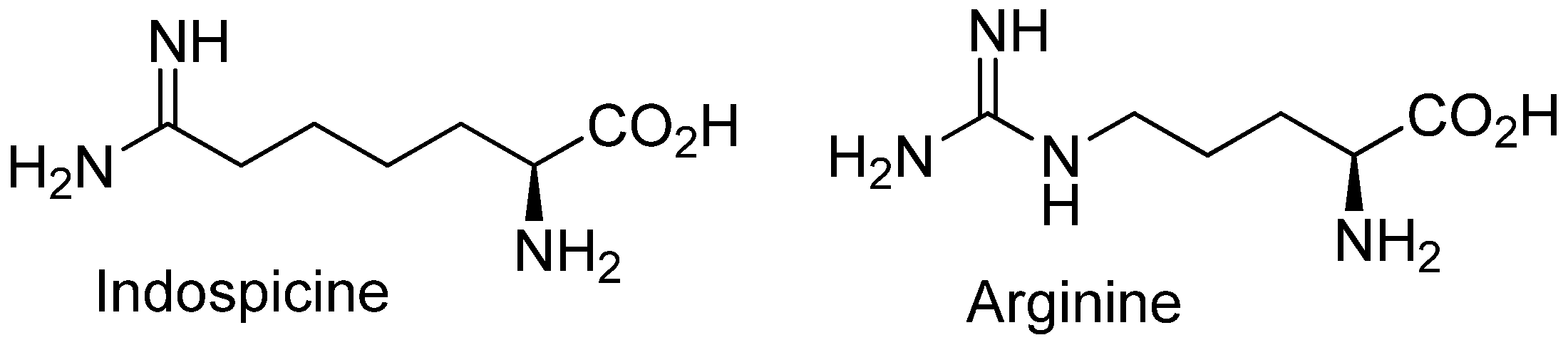
2. Plant Sources of Indospicine
2.1. Indospicine Content of Indigofera Species
2.2. Prevalence of Indospicine-Containing Indigofera Species
3. Etiology of Indospicine Toxicosis
4. Persistence of Indospicine in Animal Tissues
5. Indospicine Effects on Animals Consuming Indigofera Plants
5.1. Cattle
5.2. Sheep
5.3. Horses
5.4. Camels
5.5. Goats
5.6. Poultry/Birds
6. Conclusions
Author Contributions
Conflicts of Interest
References
- Hegarty, M.P.; Kelly, W.R.; McEwan, D.; Williams, O.J.; Cameron, R. Hepatotoxicity to dogs of horse meat contaminated with indospicine. Aust. Vet. J. 1988, 65, 337–340. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, L.M.; Fletcher, M.T.; Paul, A.E.H.; Mansfield, C.S.; O’Hara, A.J. Hepatotoxicosis in dogs consuming a diet of camel meat contaminated with indospicine. Aust. Vet. J. 2011, 89, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Pearn, J.H.; Hegarty, M.P. Indospicine—The teratogenic factor from Indigofera spicata extract causing cleft palate. Br. J. Exp. Pathol. 1970, 51, 34–36. [Google Scholar] [PubMed]
- Hegarty, M.P.; Pound, A.W. Indospicine, a hepatotoxic amino acid from Indigofera spicata: Isolation, structure, and biological studies. Aust. J. Biol. Sci. 1970, 23, 831–842. [Google Scholar]
- Skerman, P.J.; Cameron, D.G.; Riveros, F. Tropical Forage Legumes, 2nd ed.; Food and Agricultural Organisation of the United Nations: Rome, Italy, 1988. [Google Scholar]
- Emmel, M.W.; Ritchey, G.E. The toxicity of Indigofera endecaphylla Jacq. for rabbits. J. Am. Soc. Agron. 1941, 33, 675–677. [Google Scholar] [CrossRef]
- Aylward, J.H.; Court, R.D.; Haydock, K.P.; Strickland, R.W.; Hegarty, M.P. Indigofera species with agronomic potential in the tropics. Rat toxicity studies. Aust. J. Agric. Res. 1987, 38, 177–186. [Google Scholar] [CrossRef]
- Hassen, A.; Rethman, N.F.G.; van Niekerk, W.A.; Tjelele, T.J. Influence of season/year and species on chemical composition and in vitro digestibility of five Indigofera accessions. Anim. Feed Sci. Technol. 2007, 136, 312–322. [Google Scholar] [CrossRef]
- Hassen, A.; Rethman, N.F.G.; Apostolides, Z.; van Niekerk, W.A. Forage production and potential nutritive value of 24 shrubby Indigofera accessions under field conditions in South Africa. Trop. Grasslands 2008, 42, 96–103. [Google Scholar]
- Du Puy, D.J.; Labat, J.N.; Schrire, B.D. The separation of two previously confused species in the Indigofera spicata complex (Leguminosae: Papilionoideae). Kew Bull. 1993, 48, 727–733. [Google Scholar]
- Ossedryver, S.M.; Baldwin, G.I.; Stone, B.M.; McKenzie, R.A.; van Eps, A.W.; Murray, S.; Fletcher, M.T. Indigofera spicata (creeping indigo) poisoning of three ponies. Aust. Vet. J. 2013, 91, 143–149. [Google Scholar] [CrossRef] [PubMed]
- Pollitt, S. Residue Implications of Indospicine, a Toxic, Non-Protein Amino Acid. Ph.D. Thesis, Department of Physiology and Pharmacology, The University of Queensland, Brisbane, Australia, 2001. [Google Scholar]
- Tan, E.T.T.; Silcock, R.G.; Al Jassim, R.; D’Arcy, B.R.; Fletcher, M.T.; The University of Queensland, Brisbane, Queensland, Australia. Unpublished work. 2015.
- Gardner, D.; Riet-Correa, F. Analysis of the toxic amino acid indospicine by liquid chromatography-tandem mass spectrometry. Int. J. Poisonous Plant Res. 2011, 1, 20–27. [Google Scholar]
- Charlwood, B.V.; Morris, G.S.; Grenham, M.J. A chemical database for the Leguminosae. In Database in Systematics; Alkin, R., Bisby, F.A., Eds.; Academic Press: London, UK, 1984; pp. 201–208. [Google Scholar]
- Aylward, J.H. Screening legumes for toxicity. In CSIRO Division of Tropcal Crops and Pastures Annual Report 1981–1982; CSIRO: New South Wales, Australia, 1982; pp. 80–81. [Google Scholar]
- Miller, R.W.; Smith, C.R., Jr. Seeds of Indigofera species. Their content of amino acids that may be deleterious. J. Agric. Food Chem. 1973, 21, 909–912. [Google Scholar] [CrossRef] [PubMed]
- Duke, J.A. Handbook of Legumes of World Economic Importance; Plenum Press: New York, NY, USA, 1981. [Google Scholar]
- Schrire, B.D.; Lavin, M.; Barker, N.P.; Forest, F. Phylogeny of the tribe Indigofereae (Leguminosae–Papilionoideae): Geographically structured more in succulent-rich and temperate settings than in grass-rich environments. Am. J. Bot. 2009, 96, 816–852. [Google Scholar] [CrossRef] [PubMed]
- PIER (Pacific Islands Ecosystems at Risk). Indigofera Hendecaphylla Jacq., Fabaceae. Available online: http://www.hear.org/pier/species/indigofera_hendecaphylla.htm (accessed on 9 July 2015).
- PIER (Pacific Islands Ecosystems at Risk). Indigofera spicata Forssk., Fabaceae. Available online: http://www.hear.org/pier/species/indigofera_spicata.htm (accessed on 9 July 2015).
- Wagner, W.L.; Herbst, D.R.; Lorence, D.H. Flora of the Hawaiian Islands website. Available online: http://botany.si.edu/pacificislandbiodiversity/hawaiianflora/index.htm (accessed on 9 July 2015).
- Bezabih, M.; Pellikaan, W.F.; Tolera, A.; Khan, N.A.; Hendriks, W.H. Chemical composition and in vitro total gas and methane production of forage species from the Mid Rift Valley grasslands of Ethiopia. Grass Forage Sci. 2014, 69, 635–643. [Google Scholar] [CrossRef]
- GRIN (USDA Germplasm Resources Information Network). Taxon: Indigofera linnaei Ali. Available online: http://www.ars-grin.gov/cgi-bin/npgs/html/taxon.pl?70382 (accessed on 9 July 2015).
- Gracie, A.; Brown, A.; Saville, P. AgNote: Birdsville Disease (Northern Territory Government). Available online: http://www.nt.gov.au/d/Content/File/p/Anim_Dis/633.pdf (accessed on 9 July 2015).
- Dowling, R.M.; McKenzie, R.A. Poisonous Plants: A Field Guide; Department of Primary Industries: Brisbane, Australia, 1993. [Google Scholar]
- GRIN (USDA Germplasm Resources Information Network). Taxon: Indigofera lespedezioides Kunth. Available online: http://www.ars-grin.gov/cgi-bin/npgs/html/taxon.pl?20025 (accessed on 9 July 2015).
- Hastings, R.B. Medicinal legumes of Mexico: Fabaceae, Papilionoideae, Part One. Econ. Bot. 1990, 44, 336–348. [Google Scholar] [CrossRef]
- Lima, E.F.; Riet-Correa, F.; Gardner, D.R.; Barros, S.S.; Medeiros, R.M.T.; Soares, M.P.; Riet-Correa, G. Poisoning by Indigofera lespedezioides in horses. Toxicon 2012, 60, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.V.R. Drugs and Foods from Little-Known Plants: Notes in Harvard University Herbaria; Harvard University Press: Cambridge, MA, USA, 1973. [Google Scholar]
- GRIN (USDA Germplasm Resources Information Network). Taxon: Indigofera vicioides Jaub. & Spach. Available online: http://www.ars-grin.gov/cgi-bin/npgs/html/tax_search.pl (accessed on 9 July 2015).
- GRIN (USDA Germplasm Resources Information Network). Taxon: Indigofera volkensii Taub. Available online: http://www.ars-grin.gov/cgi-bin/npgs/html/tax_search.pl (accessed on 9 July 2015).
- Pearn, J.H. Report of a new site-specific cleft palate teratogen. Nature 1967, 215, 980–981. [Google Scholar] [CrossRef] [PubMed]
- Pearn, J.H. Studies on a site-specific cleft palate teratogen. The toxic extract from Indigofera spicata Forssk. Br. J. Exp. Pathol. 1967, 48, 620–626. [Google Scholar] [PubMed]
- Nordfeldt, S.; Henke, L.A.; Morita, K.; Matsumoto, H.; Takahash, M.; Younge, O.R.; Willers, E.H.; Cross, R.F. Feeding Tests with Indigofera Endecaphylla Jacq. (Creeping Indigo) and Some Observations on its Poisonous Effects on Domestic Animals; Technical Bulletin No. 15; Hawaii Agriculture Experiment Station, University of Hawaii: Honolulu, HI, USA, 1952; pp. 5–23. [Google Scholar]
- Christie, G.S.; Madsen, N.P.; Hegarty, M.P. Acute biochemical changes in rat liver induced by the naturally-occurring amino acid indospicine. Biochem. Pharmacol. 1969, 18, 693–700. [Google Scholar] [CrossRef]
- Madsen, N.P.; Christie, G.S.; Hegarty, M.P. Effect of indospicine on incorporation of l-arginine-14C into protein and transfer ribonucleic acid by cell-free systems from rat liver. Biochem. Pharmacol. 1970, 19, 853–857. [Google Scholar] [CrossRef]
- Madsen, N.P.; Hegarty, M.P. Inhibition of rat liver homogenate arginase activity in vitro by the hepatotoxic amino acid indospicine. Biochem. Pharmacol. 1970, 19, 2391–2393. [Google Scholar] [CrossRef]
- Christie, G.S.; de Munk, F.G.; Madsen, N.P.; Hegarty, M.P. Effects of an arginine antagonist on stimulated human lymphocytes in culture. Pathology 1971, 3, 139–144. [Google Scholar] [CrossRef] [PubMed]
- De Munk, F.G.; Christie, G.S.; Hegarty, M.P. The effects of indospicine on bone marrow cells in liquid culture. Pathology 1972, 4, 133–137. [Google Scholar]
- Hutton, E.M.; Windrum, G.M.; Kratzing, C.C. Studies on the toxicity of Indigofera endecaphylla: II. Toxicity for mice. J. Nutr. 1958, 65, 429–440. [Google Scholar] [PubMed]
- Pollitt, S.; Hegarty, M.P.; Pass, M.A. Analysis of the amino acid indospicine in biological samples by high performance liquid chromatography. Nat. Toxins 1999, 7, 233–240. [Google Scholar] [CrossRef]
- Hegarty, M.P. Non-metallic chemical residues in toxic plants with potential importance to animal and human health. In Vet Update 92; Osborne, H.G., Ed.; Continuing Professional Education, University of Queensland: Brisbane, Australia, 1992; pp. 323–332. [Google Scholar]
- Arid Zone Research Institute. Indospicine in beef. In Northern Territory Department of Primary Industries and Fisheries Technical Annual Report 1987–1988; Technical Bulletin No. 150; NTDPIF: Alice Springs, Australia, 1989; p. 39. [Google Scholar]
- Young, M.P. Investigation of the Toxicity of Horsemeat due to Contamination by Indospicine. Ph.D. Thesis, School of Veterinary Science, University of Queensland, Brisbane, Australia, 1992. [Google Scholar]
- Bogdan, A.V. Observations on palatability of some leguminous plants of Kenya. East Afr. Agric. J. 1949, 15, 38–40. [Google Scholar]
- Jeganathan, P. Toxic effects of feeding Indigofera endecaphylla (Jacq.) to calves. Ceylon Vet. J. 1953, 1, 83–85. [Google Scholar]
- Dalziel, J.M. The Useful Plants of West Tropical Africa; The Crown Agents for the Colonies: London, UK, 1937; pp. 243–247. [Google Scholar]
- Nordfeldt, S.; Young, O.R. Toxicity of creeping indigo to livestock. Hawaii Agric. Expt. Station Univ. Hawaii Prog. Notes 1949, 55, 1–2. [Google Scholar]
- Yelf, J.D. The toxicity of creeping indigo in Fiji. Agric. J. 1959, 29, 9–10. [Google Scholar]
- Maxwell, H.S.; Nettleton, P.; Kirkland, P.D. Overview of congenital and inherited anomalies. Merck Veterinary Manual. Available online: http://www.merckvetmanual.com/mvm/ (accessed on 9 July 2015).
- Keeler, R.F. Plant metabolites that are teratogenic in offspring and toxic in the dam. Toxicon 1983, 21, 221–225. [Google Scholar] [CrossRef]
- Australia’s Virtual Herbarium. Available online: http://avh.chah.org.au/ (accessed on 9 July 2015).
- Burns, B.M.; Fordyce, G.; Holroyd, R.G. A review of factors that impact on the capacity of beef cattle females to conceive, maintain a pregnancy and wean a calf—Implications for reproductive efficiency in northern Australia. Anim. Reprod. Sci. 2010, 122, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Bunter, K.L.; Johnston, D.J.; Wolcott, M.L.; Fordyce, G. Factors associated with calf mortality in tropically adapted beef breeds managed in extensive Australian production systems. Anim. Prod. Sci. 2014, 54, 25–36. [Google Scholar] [CrossRef]
- Maskasame, C. Toxicity and Nutritional Value of Some Promising Pasture Legumes in Rats and Sheep. Master’s Thesis, Department of Physiology and Pharmacology, The University of Queensland, Brisbane, Australia, 1984. [Google Scholar]
- Nath, K.; Malik, N.; Singh, O. Chemical composition and nutritive value of Indigofera enneaphylla and I. cordifolia as sheep feeds. Aust. J. Exp. Agric. 1971, 11, 178–185. [Google Scholar] [CrossRef]
- Srinivas, B.; Swain, N. Seasonal dynamics in vegetation and rumen microbial nitrogen production and nutritional status of grazing sheep in a semiarid rangeland in eastern Rajasthan, India. Grassland Sci. 2011, 57, 219–224. [Google Scholar] [CrossRef]
- Krishna, L.; Vaid, J.; Singh, B. Pathological study on Indigofera teysmanni toxicity in sheep. Indian J. Comp. Microbiol. Immunol. Infect. Dis. 1986, 7, 14–17. [Google Scholar]
- Morton, J.F. Creeping indigo (Indigofera spicata Forsk.) (Fabaceae): A hazard to herbivores in Florida. Econ. Bot. 1989, 43, 314–327. [Google Scholar] [CrossRef]
- Majak, W.; Benn, M.; McEwan, D.; Pass, M.A. Three nitropropanoyl esters of glucose from Indigofera linnaei. Phytochemistry 1992, 31, 2393–2395. [Google Scholar]
- Williams, M.C. Nitro compounds in Indigofera species. Agron. J. 1981, 73, 434–436. [Google Scholar] [CrossRef]
- Cooke, A.R. The toxic constituent of Indigofera endecaphylla. Arch. Biochem. Biophys. 1955, 55, 114–120. [Google Scholar] [CrossRef]
- Majak, W.; Pass, M.A. Aliphatic nitrocompounds. In Toxicants of Plant Origin, Volume II, Glycosides; Cheeke, P.R., Ed.; CRC Press: Boca Raton, FL, USA, 1989; pp. 143–159. [Google Scholar]
- Shear, D.A.; Haik, K.L.; Dunbar, G.L. Creatine reduces 3-nitropropionic-acid-induced cognitive and motor abnormalities in rats. NeuroReport 2000, 11, 1833–1837. [Google Scholar] [CrossRef] [PubMed]
- Northern Territory Dept Primary Industries Fisheries & Mines. Northern Territory of Australia Meat Industries Act. Available online: http://www.austlii.edu.au/au/legis/nt/consol_act/mia184.txt/cgi-bin/download.cgi/download/au/legis/nt/consol_act/mia184.rtf#_Toc263691324 (accessed on 9 July 2015).
- Dörges, B.; Heucke, J.; Dance, R. The Palatability of Central Australian Plant Species to Camels; Technote No. 116; Department of Primary Industry, Fisheries and Resources, Northern Territory Government: Alice Springs, Australia, 2003. Available online: www.nt.gov.au/d/Content/File/p/Technote/TN116.pdf.
- Tan, E.T.T.; Al Jassim, R.A.M.; Cawdell-Smith, J.A.; Ossedryver, S.M.; Fletcher, M.T.; The University of Queensland, Brisbane, Queensland, Australia. Unpublished work. 2015.
- Suliman, H.B.; Wasfi, I.A.; Tartour, G.; Adam, S.E.I. The effects of Indigofera hochstetteri on goats. Rev. Elev. Med. Vet. Pays Trop. 1983, 36, 393–402. [Google Scholar] [PubMed]
- Rosenberg, M.M.; Zoebisch, O.C. A chick test for toxicity in forage legumes. Agron. J. 1952, 44, 315–318. [Google Scholar] [CrossRef]
- Morris, M.P.; Pagan, C.; Warmke, H.E. Hiptagenic acid, a toxic component of Indigofera endecaphylla. Science 1954, 119, 322–323. [Google Scholar] [CrossRef] [PubMed]
- Britten, E.J.; Matsumoto, H.; Palafox, A.L. Comparative toxic effects of 3-nitropropionic acid, sodium nitrite and Indigofera endecaphylla on chicks. Agron. J. 1959, 51, 462–464. [Google Scholar] [CrossRef]
- Britten, E.J.; Palafox, A.L.; Matsumoto, H. Selection for toxicity in single plants of Indigofera endecaphylla by biological assay. Agron. J. 1959, 51, 651–654. [Google Scholar] [CrossRef]
- Britten, E.J.; Palafox, A.L.; Frodyma, M.M.; Lynd, F.T. Level of 3-nitropropanoic acid in relation to toxicity of Indigofera spicata in chicks. Crop Sci. 1963, 3, 415–416. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fletcher, M.T.; Al Jassim, R.A.M.; Cawdell-Smith, A.J. The Occurrence and Toxicity of Indospicine to Grazing Animals. Agriculture 2015, 5, 427-440. https://doi.org/10.3390/agriculture5030427
Fletcher MT, Al Jassim RAM, Cawdell-Smith AJ. The Occurrence and Toxicity of Indospicine to Grazing Animals. Agriculture. 2015; 5(3):427-440. https://doi.org/10.3390/agriculture5030427
Chicago/Turabian StyleFletcher, Mary T., Rafat A. M. Al Jassim, and A. Judith Cawdell-Smith. 2015. "The Occurrence and Toxicity of Indospicine to Grazing Animals" Agriculture 5, no. 3: 427-440. https://doi.org/10.3390/agriculture5030427