Horizontal Transmission of the Entomopathogen Fungus Metarhizium anisopliae in Microcerotermes diversus Groups

Abstract
:1. Introduction
2. Experimental Section
2.1. Collection of Termites
2.2. Fungal Isolate
2.3. Preparation of Fungal Suspension
2.4. Experiment

2.5. Statistical Analysis
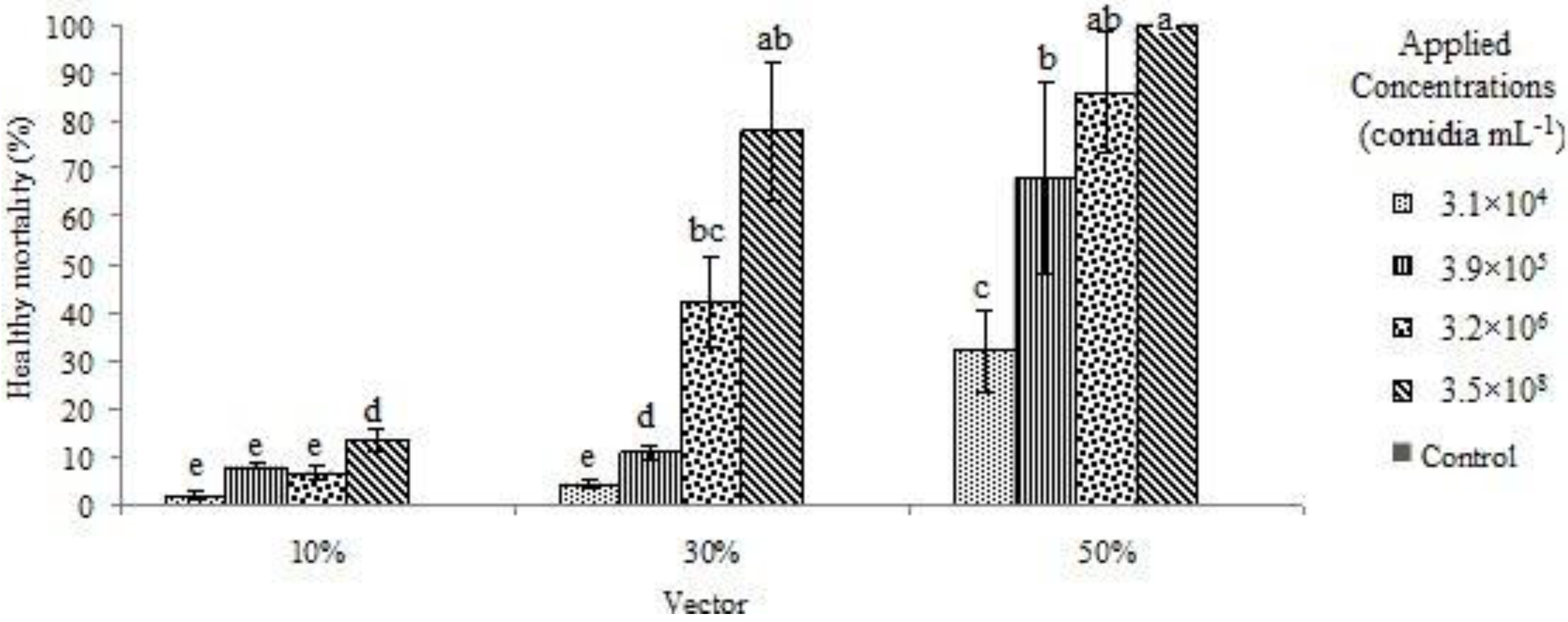
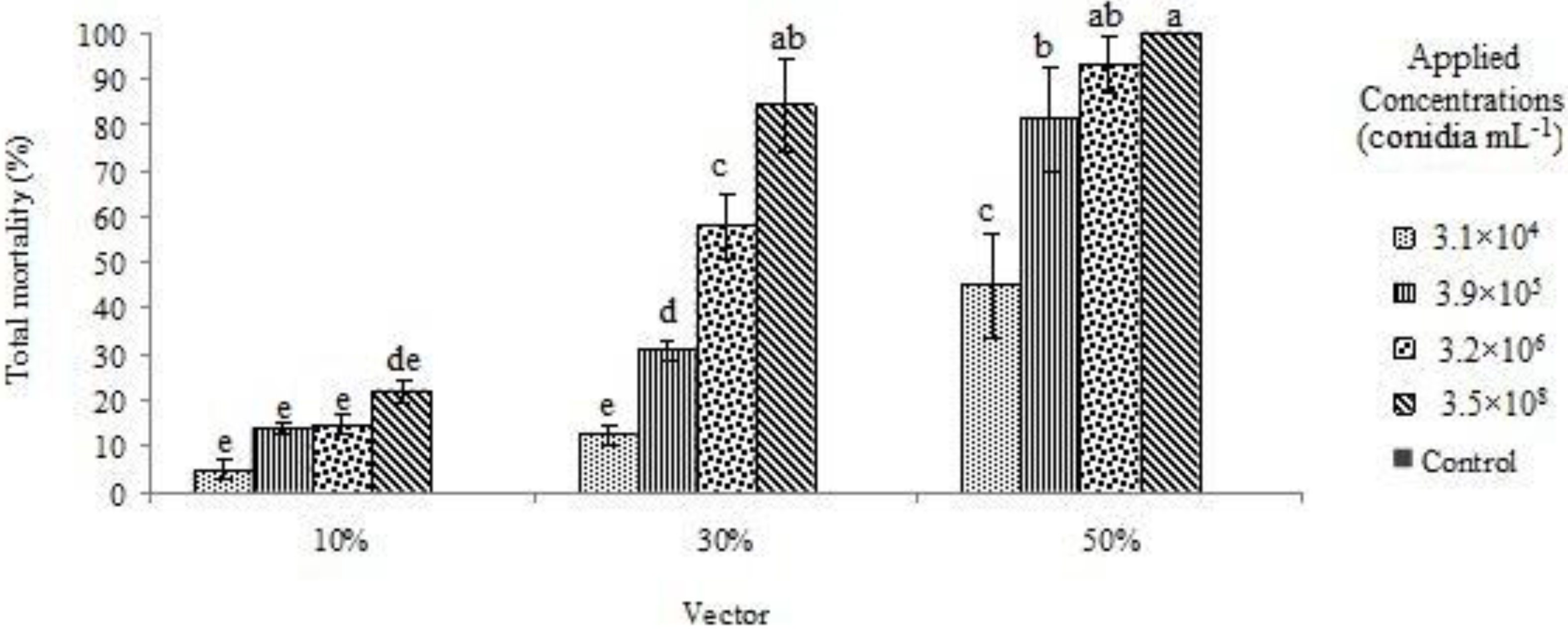
3. Results and Discussion
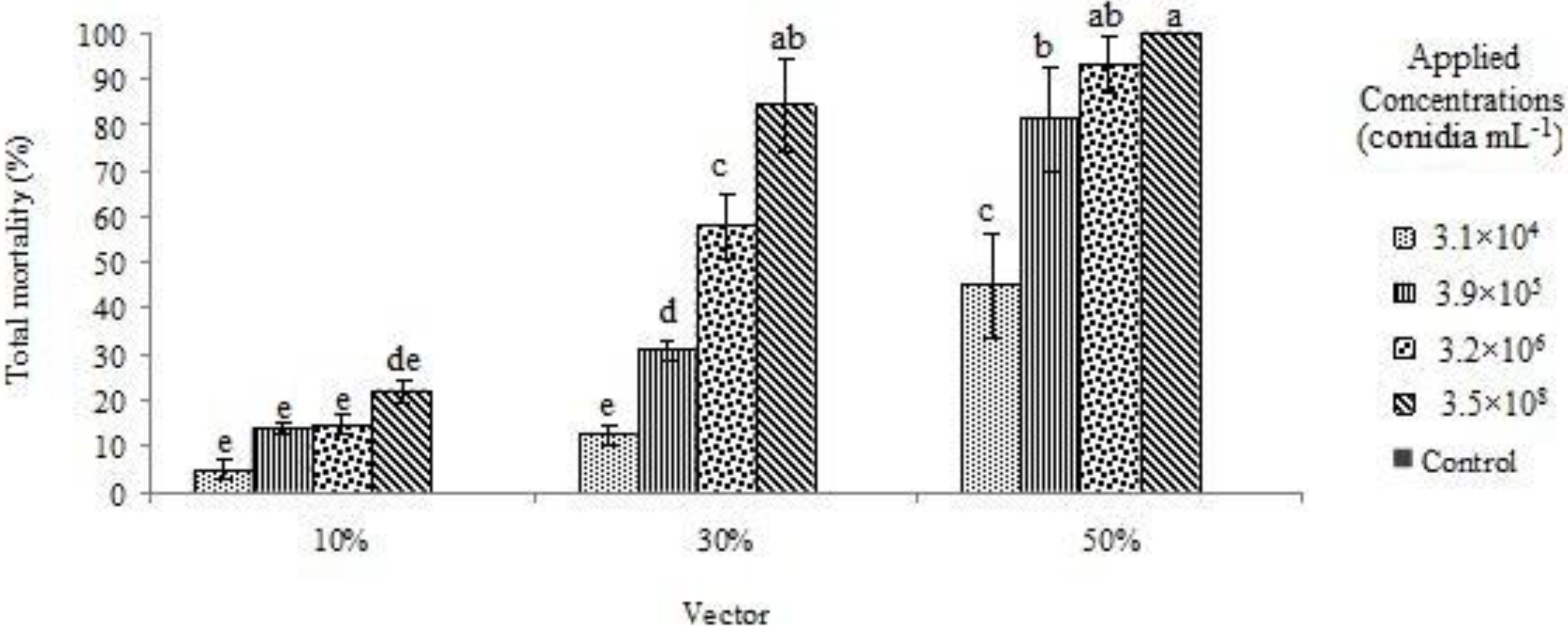

{kind=link}
{kind=link}
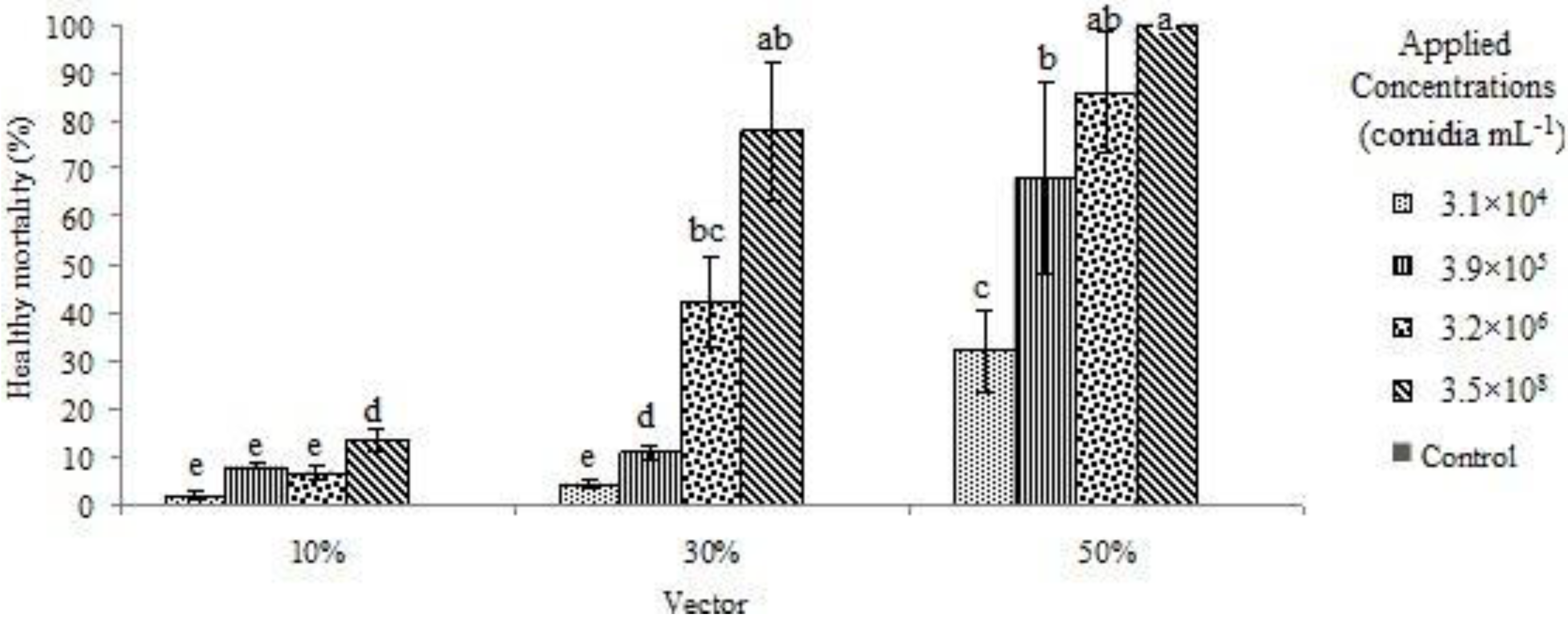{kind=link}
| Concentration | Vector (%) | LT50(day) | LT90(day) |
|---|---|---|---|
| (Conidia mL−1) | (95% Fiducial limits) | (95% Fiducial limits) | |
| 3.1 × 104 | 10 | -* | - |
| 30 | - | - | |
| 50 | 19.5 (17–23) | - | |
| 3.9 × 105 | 10 | - | - |
| 30 | 30 (21–55) | - | |
| 50 | 7.27 (6.4–8.29) | 27.81(21.22–41.63) | |
| 3.2 × 106 | 10 | - | - |
| 30 | 12.12 (10.32–15.04) | - | |
| 50 | 4.68 (3.51–5.85) | 23.49(15.96–48.05) | |
| 3.5 × 108 | 10 | - | - |
| 30 | 6.23 (4.51–8.58) | 47.32(24.77–215.03) | |
| 50 | 2.33 (1.20–3.32) | 12.86(8.82–26.99) |
4. Conclusion
Acknowledgments
References and Notes
- Rath, A.C. The use of entomopathogenic fungi for control of termites. Biocontrol. Sci. Techn. 2000, 10, 563–581. [Google Scholar] [CrossRef]
- Arab, A.; Costa-Leonardo, A.M.; Casarin, F.E.; Guaraldo, A.C.; Chaves, R.C. Foraging activity and demographic patterns of two termite species (Isoptera: Rhinotermitidae) living in urban landscapes in southeastern Brazil. Eur. J. Entomol. 2005, 102, 691–697. [Google Scholar]
- Roisin, Y. Queen replacement in the termite Microcerotermes papuanus. Entomol. Exp. Appl. 1990, 5, 83–90. [Google Scholar] [CrossRef]
- Edwards, R.; Mill, A.E. Termites in buildings: Their biology and control; Rentokil Limited: London, UK, 1986. [Google Scholar]
- Habibpour, B.; Mossadegh, M.S.; Henderson, G.; Moharramipour, S. Laboratory evaluation of two insect growth regulators (IGRs) on Microcerotermes diversus (Silvestri) (Isoptera: Termitidae) in southwest Iran. Sociobiology 2007, 50, 1199–1209. [Google Scholar]
- Habibpour, B. Laboratory of flurox, a chitin synthesis inhibitor, on the termite, Microcerotermes divers. J. Insect Sci. 2010, 10, 1–8. [Google Scholar] [CrossRef]
- Logan, J.W.M.; El Bakri, A. Termite damage to date palms (Phoenix dactylifera L.) in Northern Sudan with particular reference to the Dongle District. Trop. Sci. 1990, 30, 95–108. [Google Scholar]
- Habibpour, B. Laboratory and field evaluation of bait-toxicants for suppression subterranean termite populations in Ahvaz (Iran). PhD Thesis, Department of Plant Protection, College of Agriculture, Shahid Chamran University of Ahvaz, Ahvaz, Iran, 2006. [Google Scholar]
- Bayon, I.L.; Ansard, D.; Brunet, C.; Girardi, S.; Paulmier, I. Biocontrol of Reticulitermes santonensis by entomopathogenic fungi improvement of the contamination Process. In Presented at the International Research Group on Wood Protection (IRG/WP/DOC 00–10359), Kona, Hawaii, USA, 14-19 May 2000.
- Tajick Ghanbalani, M.A.; Asgharzadeh, A.; Hadizadeh, A.R.; Mohammadi Sharif, M. A quick method for Metarhizium anisopliae isolation from cultural soils. Am. J. Agr. Biol. Sci. 2009, 4, 152–155. [Google Scholar] [CrossRef]
- Kramm, K.R.; West, D.F.; Rockenbach, P.G. Termite pathogens: Transfer of the entomopathogen Metarhizium anisopliae between Reticulitermes sp. Termites. J. Invertebr. Pathol. 1982, 39, 1–5. [Google Scholar] [CrossRef]
- Ignoffo, C.M. Environmental factors affecting persistence of entomopathogens. Fla. Entomol. 1992, 75, 516–525. [Google Scholar] [CrossRef]
- Wang, C.; Powell, J.E. Cellulose bait improves the effectiveness of Metarhizium anisopliae as a microbial control of termites (Isoptera: Rhinotermitidae). Biol. Control 2004, 30, 523–529. [Google Scholar] [CrossRef]
- Chouvenc, T.; Su, N.Y. Apparent synergy among defense mechanisms in subterranean termites (Rhinotermitidae) against epizootic events: Limits and potential for biological control. J. Econ. Entomol. 2010, 103, 1327–1337. [Google Scholar] [CrossRef]
- Culliney, T.W.; Grace, J.K. Prospects for the biological control of subterranean termites (Iso.: Rhinotermitidae) with special reference to Coptotermes formosanus. B Entomol. Res. 2000, 90, 9–21. [Google Scholar] [CrossRef]
- Wright, M.S.; Osbrink, W.L.A.; Lax, A.R. Transfer of entomopathogenic fungi among Formosan subterranean termites and subsequence mortality. J. Appl. Entomol. 2002, 126, 20–23. [Google Scholar] [CrossRef]
- Grace, J.K.; Zoberi, M.H. Experimental evidence for transmission of Beauveria bassiana by Reticulitermes flavipes workers (Isoptera: Rhinotermitidae). Sociobiology 1992, 20, 23–28. [Google Scholar]
- Myles, T.G. Alarm, aggregation, and defense by Reticulitermes flavipes in response to a naturally occurring isolate of Metarhizium anisoplia. Sociobiology 2002, 40, 243–255. [Google Scholar]
- Desyanti, W.H.; Zulyusri, Y.; Yumarni, A.; Jasni, H. Transmission of entomopathogenic fungus Metarhizium anisopliae Petch and Myrothecium roridum Tode ex Steudel in colony of dry wood termites Coptotermes sp. (Blattodea: Kalotermitidae) using vector. In Proceeding of the Sixth Conference of the Pacific Rim Termite Research Group, Kyoto, Japan, 2–3 March 2009; pp. 16–19.
- Desyanti, W.H. Contagious test of the entomopathogenic fungus originated from west Sumatera Indonesia between individual in colony of subterranean termites Coptotermes gestroi Wasman (Blattodea: Rhinotermitidae). In In Proceeding of the Seventh Conference of the Pacific Rim Termite Research Group, Singapore, 1–2 March 2010; pp. 16–19.
- Myles, T.G. Laboratory studies on the transmission of Metarhizium anisopliae in the Eastern subterranean termite, Reticulitermes flavipes (Isoptera: Rhinotermitidae), with a method for applying appropriate doses of conidia to trapped termites for release. Sociobiology 2002, 40, 265–276. [Google Scholar]
- Quesada-Moraga, E.; Martin-Carballo, I.; Garrido-Jurado, I.; Santiago-Alvarez, C. Horizontal transmission of Metarhizium anisopliae among laboratory populations of Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Biol. Control 2008, 47, 115–124. [Google Scholar] [CrossRef]
- Roy, H.E.; Pell, J.K. Interactions between entomopathogenic fungi and other natural enemies: Implications for biological control. Biocontrol. Sci. Technol. 2000, 10, 737–752. [Google Scholar] [CrossRef]
- Vega, F.E.; Dowd, P.F.; Lacey, L.A.; Pell, J.K.; Jackson, D.M.; Klein, M.G. Dissemination of beneficial microbial agents by insects. In Field Manual of Techniques in Invertebrate Pathology; Lacey, L.A., Kaya, H.K., Eds.; Springer: London, UK, 2000; pp. 153–177. [Google Scholar]
- Baverstock, J.; Roy, H.E.; Pell, J.K. Entomopathogenic fungi and insect behavior: From unsuspecting hosts to targeted vectors. Biocontrol 2010, 55, 89–102. [Google Scholar] [CrossRef]
- Cory, J.S.; Hoover, K. Plant-mediated effects in insect-pathogen interactions. Trends Ecol. Evol. 2006, 21, 278–286. [Google Scholar] [CrossRef]
- Chouvenc, T.; Su, N.Y.; Elliott, M.L. Interaction between the subterranean termite Reticulitermes flavipes (Isoptera: Rhinotermitidae) and the entomopathogenic fungus Metarhizium anisopliae in foraging arenas. J. Econ. Entomol. 2008, 101, 885–893. [Google Scholar] [CrossRef]
- Staples, J.A.; Milner, R.J. A laboratory evaluation of the repellency of Metarhizium anisopliae conidia to Coptotermes lacteus (Isoptera: Rhinotermitidae). Sociobiology 2000, 36, 133–148. [Google Scholar]
- Meyling, N.V.; Pell, J.K. Detection and avoidance of an entomopathogenic fungus by a generalist insect predator. Ecol. Entomol. 2006, 31, 162–171. [Google Scholar] [CrossRef]
- Thompson, S.R.; Brandenburg, R.L. Tunneling responses of mole crickets (Orthoptera: Gryllotalpidae) to the entomopathogenic fungus, Beauveria bassiana. Environl. Entomol. 2005, 34, 140–147. [Google Scholar] [CrossRef]
- Villani, M.G.; Krueger, S.R.; Schroeder, P.C.; Consolie, F.; Consolie, N.H.; Preston-Wilsey, L.M.; Roberts, D.W. Soil application effects of Metarhizium anisopliae on Japanese-beetle (Coleoptera: Scarabaeidae) behavior and survival in turfgrass microcosms. Environ. Entomol. 1994, 23, 502–513. [Google Scholar]
- Yanagawa, A.; Yokohari, F.; Shimizu, S. Influence of fungal odor on grooming behavior of the termite, Coptotermes formosanus. J. Insect Sci. 2010, 10, 141–155. [Google Scholar]
- Thorne, B.L.; Breisch, N.L. Effects of sublethal exposure to imidacloprid on subsequent behavior of subterranean termite Reticulitermes virginicus (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2001, 94, 492–498. [Google Scholar] [CrossRef]
- Boucias, D.G.; Stokes, C.; Storey, G.; Pendland, J.C. The effects of imidacloprid on the termite Reticulitermes flavipes and its interaction with the mycopathogen Beauveria bassiana. Pflanzenschutz-Nachrichten Bayer 1996, 49, 103–144. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cheraghi, A.; Habibpour, B.; Mossadegh, M.S.; Sharififard, M. Horizontal Transmission of the Entomopathogen Fungus Metarhizium anisopliae in Microcerotermes diversus Groups. Insects 2012, 3, 709-718. https://doi.org/10.3390/insects3030709
Cheraghi A, Habibpour B, Mossadegh MS, Sharififard M. Horizontal Transmission of the Entomopathogen Fungus Metarhizium anisopliae in Microcerotermes diversus Groups. Insects. 2012; 3(3):709-718. https://doi.org/10.3390/insects3030709
Chicago/Turabian StyleCheraghi, Amir, Behzad Habibpour, Mohammad Saied Mossadegh, and Mona Sharififard. 2012. "Horizontal Transmission of the Entomopathogen Fungus Metarhizium anisopliae in Microcerotermes diversus Groups" Insects 3, no. 3: 709-718. https://doi.org/10.3390/insects3030709