The Roles of Parasitoid Foraging for Hosts, Food and Mates in the Augmentative Control of Tephritidae

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Foraging Behaviors
2.1. Foraging for Hosts
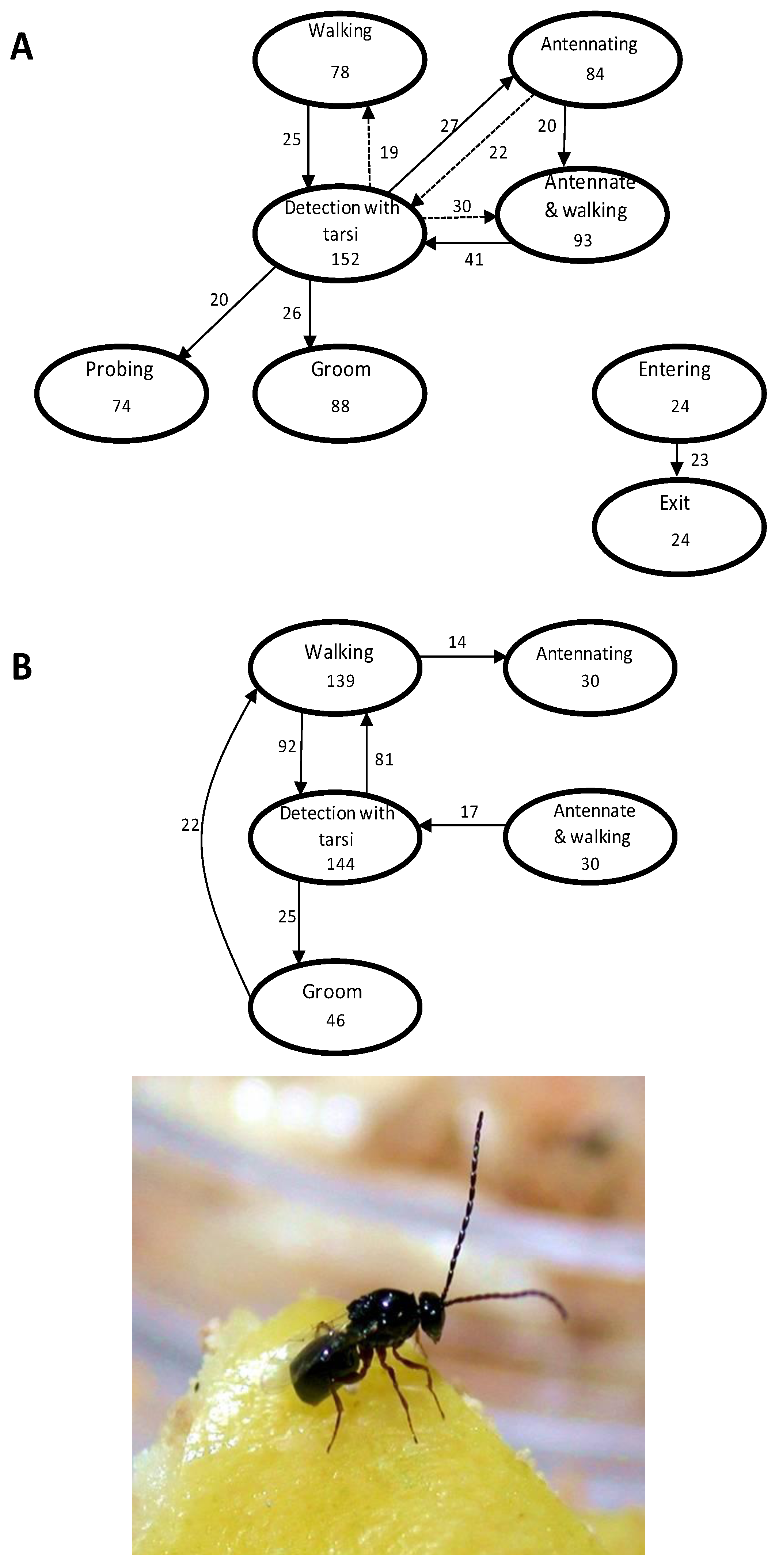
2.1.1. Visual Cues
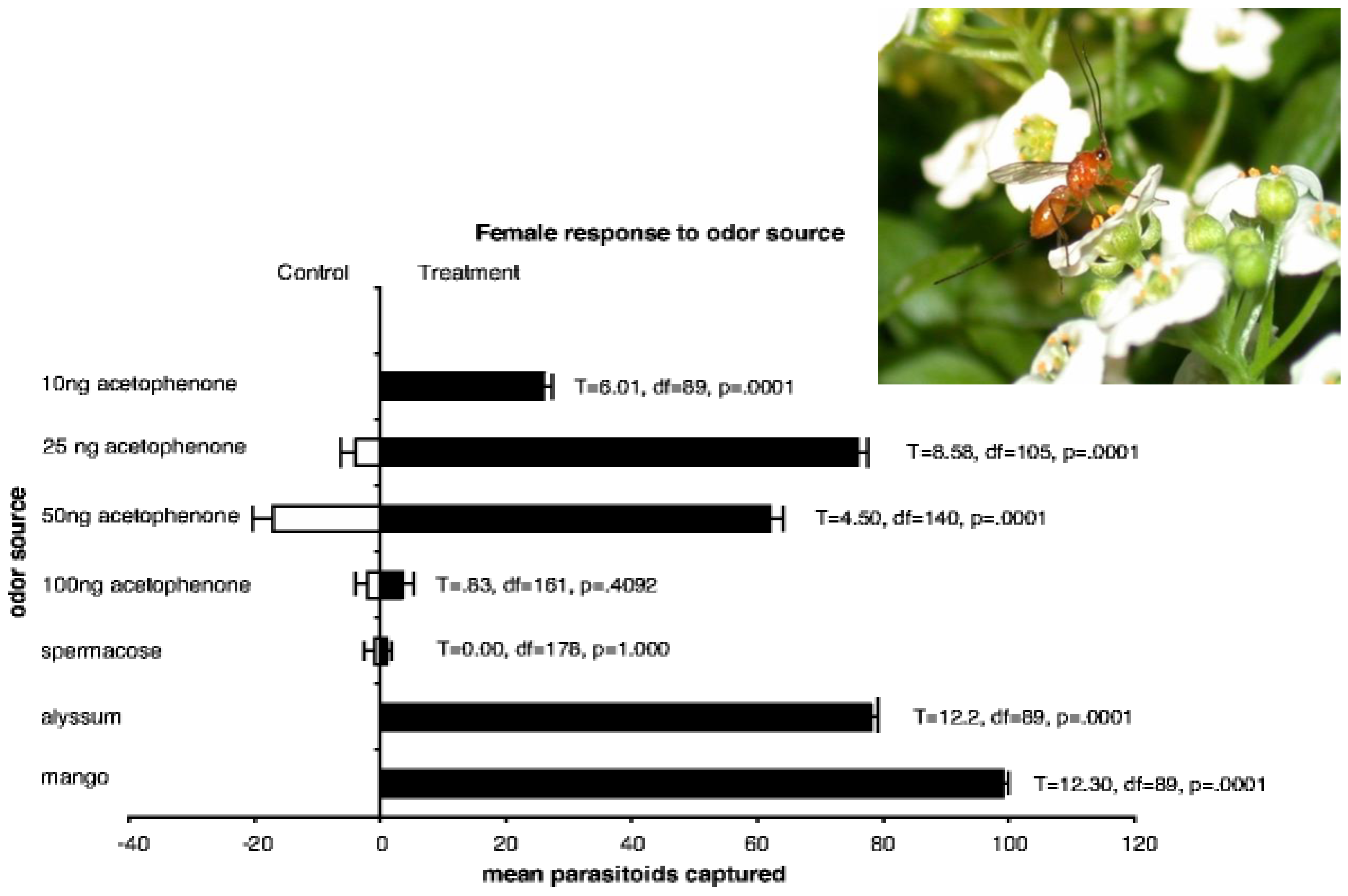
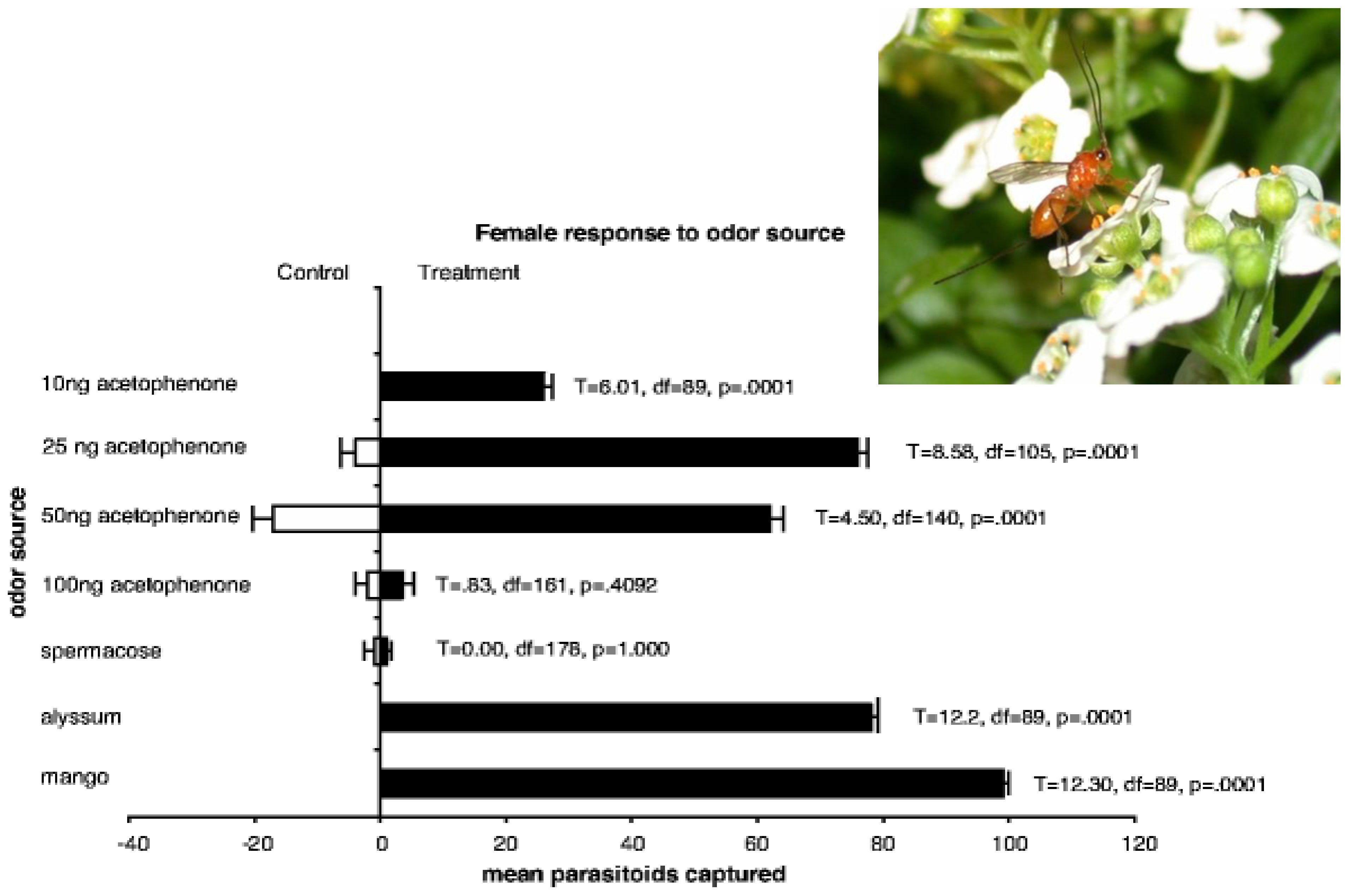
2.1.2. Chemical Cues
2.1.3. Vibration Cues
2.1.4. Plant Produced Signals
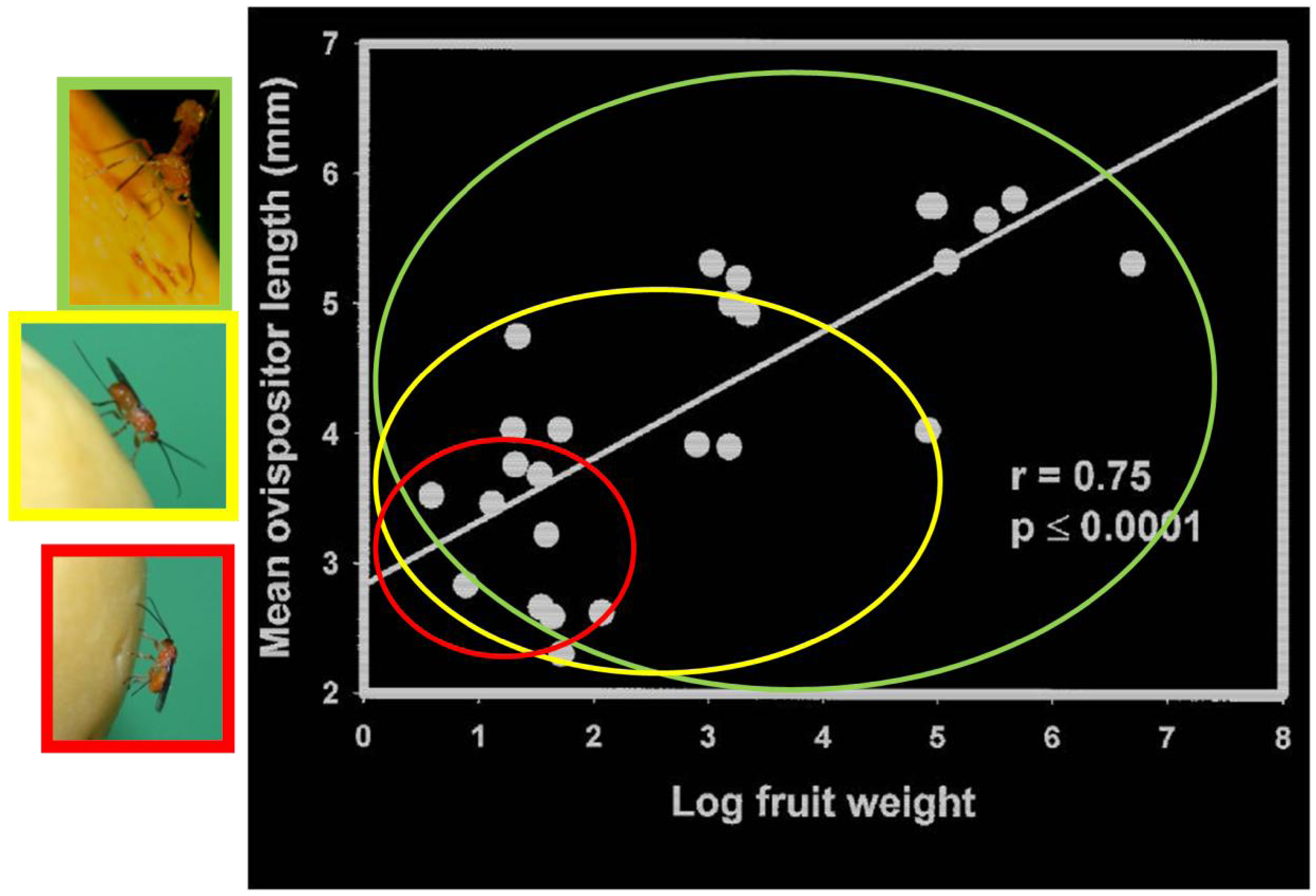
2.1.5. Host Vulnerability
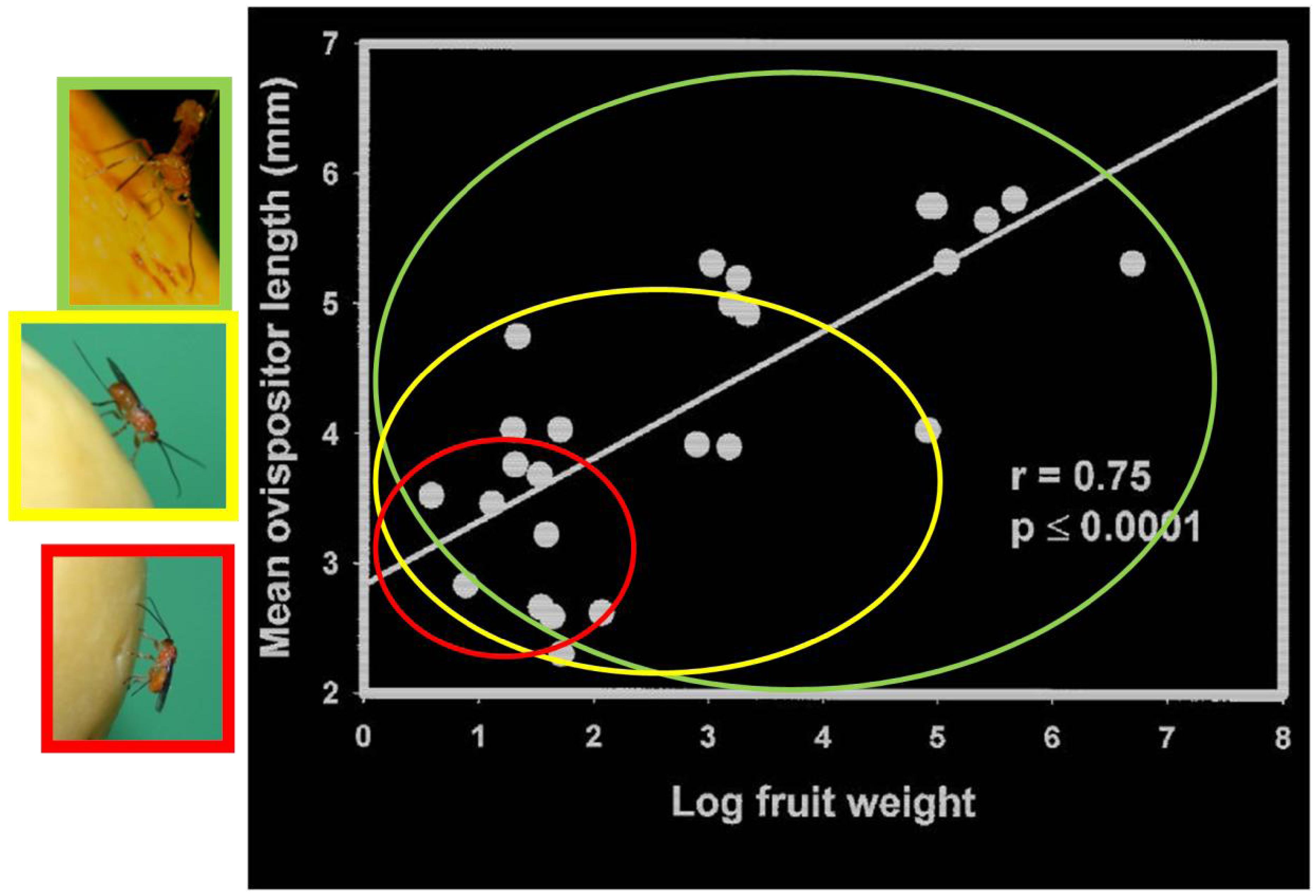
2.1.6. Competition Among Parasitoids
2.1.7. Host Range Breadth
2.1.8. Abiotic Influences on Foraging
2.1.9. Foraging for Hosts and Augmentative Biological Control
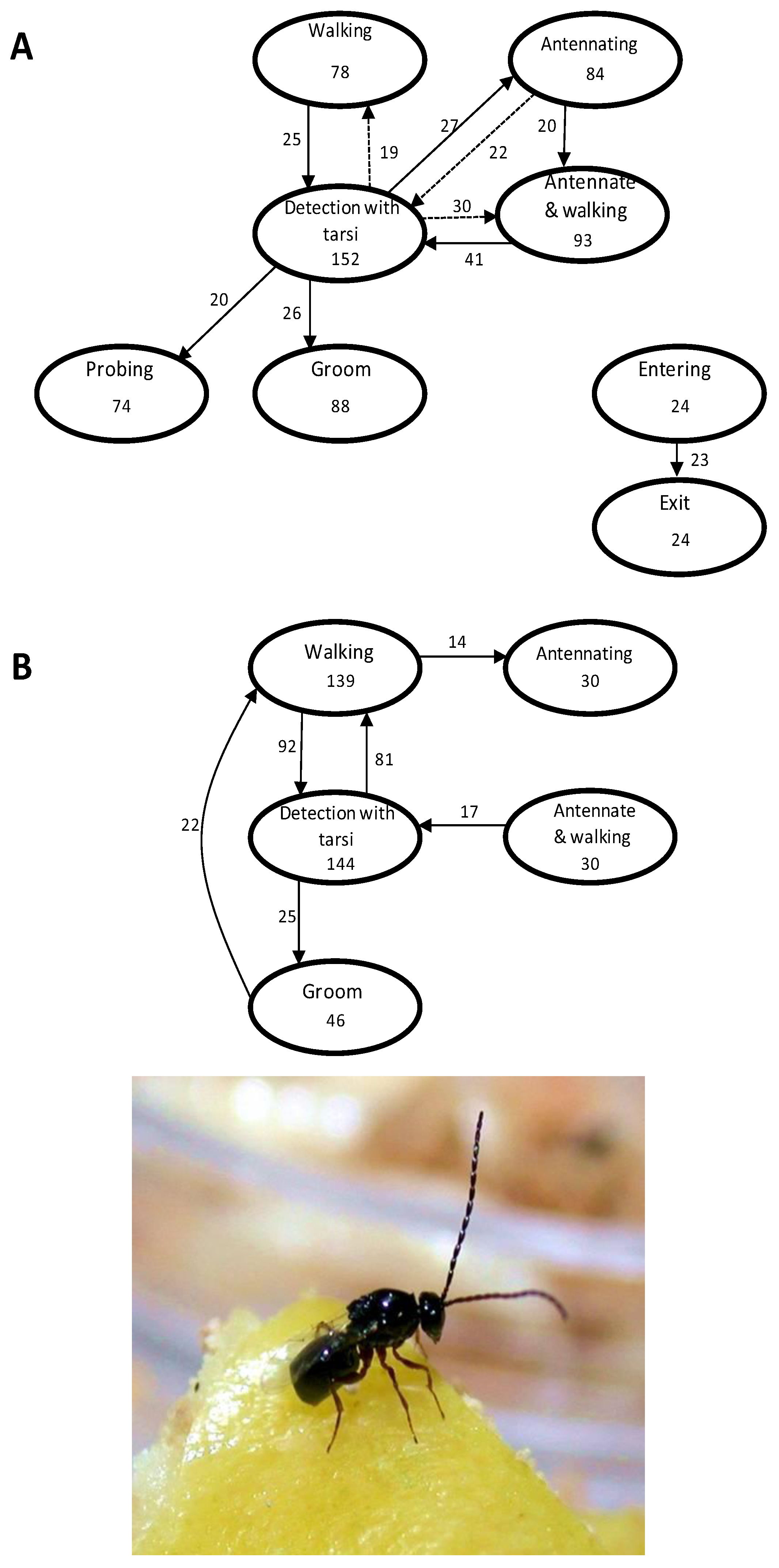
2.2. Foraging for Food
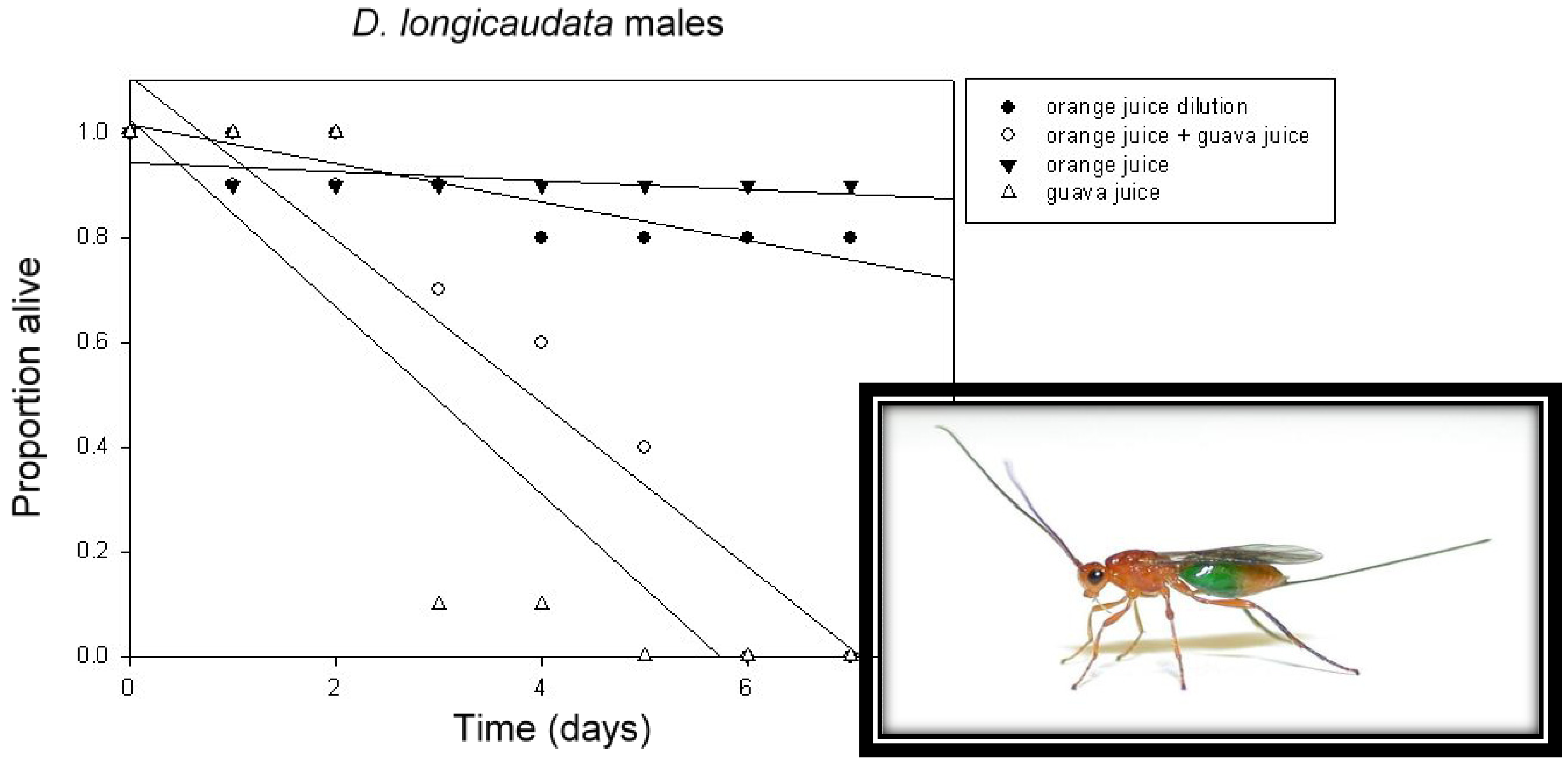
2.2.1. Foraging for Food and Augmentative Biological Control
2.3. Foraging for Mates
2.3.1. Mating Systems
2.3.2. Sex Pheromones
2.3.3. Acoustic Signals
2.3.4. Foraging for Mates and Augmentative Biological Control
3. Conclusions
Acknowledgements
References
- Ovruski, S.; Aluja, M.; Sivinski, J.; Wharton, R. Hymenopteran parasitoids on fruit-infesting Tephritidae (Diptera) in Latin America and the southern United States: Diversity, distribution, taxonomic status and their use in fruit fly biological control. Integr. Pest Manage. Rev. 2000, 5, 81–107. [Google Scholar] [CrossRef]
- Gilstrap, F.; Hart, W. Biological Control of the Mediterranean Fruit Fly in the United States and Central America; USDA-ARS.: Washingtong, DC, 1987; pp. 25–32. [Google Scholar]
- Glas, P.; Vet, L. Host-habitat location and host location by Diachasma alloem Muesebeck (Hym.: Braconidae), a parasitoid of Rhagoletis pommonella Walsh (Dipt.: Tephritidae). Neth. J. Zool. 1982, 33, 41–54. [Google Scholar] [CrossRef]
- Wäckers, F.; Lewis, W. Olfactory and visual learning and their combined influence on host site location by the parasitoid Microplitis croceipes (Cresson). Biol. Control. 1994, 4, 195–112. [Google Scholar]
- Baeder, J.; King, B. Associative learning of color by males of the parasitoid wasp Nasonia vitripennis (Hymenoptera: Pteromalidae). J. Insect Behav. 2004, 17, 201–213. [Google Scholar] [CrossRef]
- Messing, R.; Jang, E. Response of the fruit fly parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) to host-fruit stimuli. Environ. Entomol. 1992, 21, 1189–1195. [Google Scholar]
- Leyva, J.; Browning, H.; Gilstrap, H. Effect of host fruit species, size and color on parasitism of Ananstrepha ludens (Diptera: Tephritidae) by Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 1991, 20, 1469–1474. [Google Scholar]
- Rousse, P.; Chiroleu, F.; Domberg, C.; Quilici, S. Naïve Fopius arisanus females respond mainly to achromatic cues. Biol. Control. 2007, 43, 41–48. [Google Scholar] [CrossRef]
- Pérez, J.; Rojas, J.; Montoya, P.; Liedo, P.; González, F.; Castillo, A. Size, shape and hue moderate attraction and landing responses of the braconid parasitoid Fopius arisanus to fruit odour-baited visual targets. BioControl 2012, in press.. [Google Scholar]
- Benelli, G.; Canale, A. Learning of visual cues in the fruit fly parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). BioControl 2012. [Google Scholar] [CrossRef]
- Segura, D.; Viscarret, M.; Ovrusi, S.; Cladera, J. Response of the fruit fly parasitoid Diachasmimorpha longicaudata to host and host-habitat volatile cues. Entomol. Exp. Appl. 2012, 143, 164–176. [Google Scholar] [CrossRef]
- Greany, P.; Tumlinson, J.; Chambers, D.; Bousch, G. Chemically mediated host finding by Biosteres [Opius] longicaudatus, a parasitoid of tephritid fruit fly larvae. J. Chem. Ecol. 1977, 3, 189–195. [Google Scholar] [CrossRef]
- Eben, A.; Benray, B.; Sivinski, J.; Aluja, M. Host species and host plant effects on preference and performance of Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 2000, 29, 87–94. [Google Scholar] [CrossRef]
- Rohrig, E.; Sivinski, J.; Teal, P.; Stuhl, C.; Aluja, M. A floral-derived compound attractive to the tephritid fruit fly parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae). J. Chem. Ecol. 2008, 34, 549–557. [Google Scholar] [CrossRef]
- Stuhl, C.; Sivinski, J.; Teal, P.; Paranhos, B.; Aluja, M. A compound produced by fruigivorous Tephritidae (Diptera) larvae promotes oviposition behavior by the biological control agent Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Environ. Entomol. 2011, 40, 727–736. [Google Scholar] [CrossRef]
- Altizar, A.; Montoya, P.; Rojas, J. Response of Fopius arisanus (Hymenoptera: Braconidae) to fruit volatiles in a wind tunnel. Fla. Entomol. 2004, 87, 616–618. [Google Scholar] [CrossRef]
- Messing, R.; Klungness, M.; Jang, E.; Nishijima, K. Response of the melon fly parasitoid Psyttalia fletcheri [Hymenoptera: Braconidae] to host-habitat stimuli. J. Insect Behav. 1996, 9, 933–945. [Google Scholar] [CrossRef]
- Nishida, T. An experimental study of ovipositional behavior of Opius fletcheri. P. Hawaii Entomol. Soc. 1956, 16, 126–134. [Google Scholar]
- Whitman, D.; Eller, F. Parasitic wasps orient to green leaf volatiles. Chemecology 1990, 1, 69–75. [Google Scholar] [CrossRef]
- Prokopy, R.; Webster, R. Oviposition deterring pheromone of Rhagoletis pommonela- a kairomone for its parasitoid Opius lectus. J. Chem. Ecol. 1978, 4, 481–494. [Google Scholar] [CrossRef]
- Roitberg, B.; Lalonde, R. Host marking enhances parasitism risk for a fruit-infesting fly, Rhagoletis basiola. Oikos 1991, 61, 389–393. [Google Scholar] [CrossRef]
- Eitam, A.; Holler, T.; Sivinski, J.; Aluja, M. Use of host fruit chemical cues for laboratory rearing of Doryctobracon areolatus (Hymenoptera: Braconidae), a parasitoid of Anastrepha spp. (Diptera: Tephritdae). Fla. Entomol. 2003, 86, 211–216. [Google Scholar] [CrossRef]
- Bokonon-Ganta, A.; Ramadan, M.; Messing, R. Reproductive biology of Fopius ceratitivorous (Hymenoptera: Braconidae), an egg-larval parasitoid of the Mediterranean fruit fly, Ceratitis capitata (Diptera: Tephritidae). Biol. Control. 2007, 41, 361–367. [Google Scholar] [CrossRef]
- Rousse, P.; Ciroleu, F.; Veslot, J.; Quilici, S. The host- and microhabitat olfactory location by Fopius arisanus suggests a broad potential host range. Physiol. Entomol. 2007, 32, 313–321. [Google Scholar] [CrossRef]
- Granchietti, A.; Sacchetti, P.; Rosi, M.; Belcari, A. Fruit fly larval trail acts as a cue in the host location process of the pupal parasitoid Coptera occidentalis. Biol. Control. 2012, 61, 7–14. [Google Scholar]
- Baeza, G.; Sivinski, J.; Holler, T.; Aluja, M. The ability of Coptera haywardi [ogloblin] [Hymenoptera: Diapriidae] to locate and attack the pupae of the Mediterranean fruit fly, Ceratitis capitata [Wiedemann] [Diptera: Tephritidae], under seminatural conditions. Biol. Control. 2002, 23, 213–218. [Google Scholar]
- Guillén, L.; Aluja, M.; Equihua, M.; Sivinski, J. Performance of two fruit fly (Diptera: Tephritidae) pupal parasitoids [Coptera haywardi (Hymenoptera: Diapriidae) and Pachycrepoideus vindemiae (Hymenoptera: Pteromalidae)] under different soil conditions. Biol. Control. 2002, 23, 219–227. [Google Scholar]
- Lawrence, P. Host vibration- a cue to host location by the parasite. Biosteres longicaudatus. Oecologia 1981, 48, 249–251. [Google Scholar]
- Duan, J.; Messing, R. Effects of host substrate and vibration cues on ovipositor-probing behavior in two larval parasitoids of tephritid fruit flies. J. Insect Behav. 2000, 13, 175–186. [Google Scholar]
- Canale, A. Psyttalia concolor [Hymenoptera: Baraconidae]: role of host movement and host substrate in ovipositor-probing behavior. Bull. Insectology 2003, 56, 211–213. [Google Scholar]
- Djemai, I.; Casas, J.; Magal, C. Matching host reactions to parasitod wasp vibrations. Proc. Biol. Sci. 2000, 268, 2403–2408. [Google Scholar]
- Webb, J.; Landolt, P. Detecting insects in fruit by vibrations produced. J. Environ. Sci. Health 1984, 19, 367–375. [Google Scholar]
- Sivinski, J.; Calkins, C. Sexually dimorphic developmental rates in the Caribbean fruit-fly (Diptera, Tephritidae). Environ. Entomol. 1990, 19, 1491–1495. [Google Scholar]
- Mankin, R.; Hagstrum, D.; Smith, M.; Roda, A.; Kairo, M. Perspective and promise: a century of insect acoustic detection and monitoring. Am. Entomol. 2011, 57, 30–44. [Google Scholar]
- Vet, L.; van Alphen, J. A comparative functional approach to the host detection behavior of parasitic wasps. I. A quantitative study of Eucoilidae and Allysiinae. Oikos 1985, 44, 478–486. [Google Scholar]
- Van Dijken, M.; van Alphen, J. The ecological significance of differences in host detection behavior in coexisting parasitoid species. Ecol. Entomol. 1998, 23, 265–270. [Google Scholar]
- Canale, A.; Loni, A. Host location and acceptance in Psyttalia concolor: Role of host instar. Bull. Insectology 2006, 59, 7–10. [Google Scholar]
- Turlings, T.; Wäckers, F. Recruitment of predators and parasitoids by herbivore-injured plants. In Advances in Insect Chemistry; Carde, R., Millar, J., Eds.; Cambridge University Press: Cambridge, 2004; pp. 21–75. [Google Scholar]
- Bernasconi, M.; Turlings, T.; Ambrosetti, L.; Bassetti, P.; Dorn, S. Herbivore-induced emissions of maize volatiles repel the corn leaf aphid, Rhopalosiphum maidis. Ent. Exp. Appl. 1998, 87, 133–142. [Google Scholar]
- Hoffmeister, T.; Gienapp, P. Exploitation of the host’s chemical communication in a parasitoid searching for a concealed host. Ethology 1999, 105, 223–232. [Google Scholar]
- Rendon, P.; Sivinski, J.; Holler, T.; Bloem, K.; Lopez, M.; Martinez, A.; Aluja, M. The effects of sterile males and two braconid parasitoids, Fopius arisanus (Sonan) and Diachasmimorpha krausii (Fullaway) (Hymenoptera), on caged populations of Mediterranean fruit flies, Ceratitis capitata (Wied.) (Diptera: Tephritidae) at various sites in Guatemala. Biol. Control. 2006, 36, 224–231. [Google Scholar] [CrossRef]
- Aluja, M.; Ovruski, S.; Guillén, L.; Oroño, L.; Sivinski, J. Comparison of the host searching and oviposition behaviors of the tephritid (Diptera) parasitoids Aganaspis pelleranoi and Odontosema anastrephae [Hymenoptera: Figitidae: Eucoilinae]. J. Insect. Behav. 2009, 22, 423–451. [Google Scholar] [CrossRef]
- Sivinski, J. The influence of host fruit morphology on parasitization rrates in the Caribbean fruit fly. Anastrepha suspensa. Entomophaga 1991, 36, 447–454. [Google Scholar] [CrossRef]
- Sivinski, J.; Vulinec, K.; Aluja, M. Ovipositor length in a guild of parasitoids (Hymenoptera: Braconidae) attacking Anastrepha spp. fruit flies (Diptera: Tephritidae) in southern Mexico. Ann. Entomol. Soc. Am. 2001, 94, 886–895. [Google Scholar] [CrossRef]
- Lopez, M.; Aluja, M.; Sivinski, J. Hymenopterous larval-pupal and pupal parasitoids of Anastrepha flies in Mexico. Biol. Control. 1999, 15, 119–129. [Google Scholar] [CrossRef]
- Wang, X.; Johnson, M.; Daane, K.; Yokoyama, V. Larger fruit size reduces the efficacy of Psyttalia concolor, a parasitoid of the olive fruit fly. Biol. Control. 2009, 49, 45–51. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M. The evolution of ovipositor length in the parasitic Hymenoptera and the search for predictability in biological control. Fla. Entomol. 2003, 86, 143–2003. [Google Scholar] [CrossRef]
- Moretti, R.; Calvitti, M. Mortality by parasitization in the association between the egg-pupal parasitoid Fopius arisanus and Ceratitis capitata. BioControl 2003, 48, 275–291. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M.; Lopez, M. Spatial and temporal distributions of parasitoids of Mexican Anastrepha species (Diptera: Tephritidae) within the canopies of fruit trees. Ann. Entomol. Soc. Am. 1997, 90, 604–618. [Google Scholar]
- Sivinski, J.; Aluja, M.; Holler, T. The distributions of the Caribbean fruit fly, Anastrepha suspensa (Tephritidae) and its parasitoids (Hymenoptera: Braconidae) within the canopies of fruit trees. Fla. Entomol. 1999, 82, 72–81. [Google Scholar] [CrossRef]
- Hawkins, B. Patterns and Process in Host-Parasitoid Interactions.; Cambridge University Press: Cambridge, 1994. [Google Scholar]
- García-Medel, D.; Sivinski, J.; Díaz-Fleischer, F.; Rameriez-Romero, R.; Aluja, M. Foraging behavior by six fruit fly parasitoids (Hymenoptera: Braconidae) released as single- or multiple-species cohorts in field cages: influence of fruit location and host density. Biol. Control. 2007, 43, 12–22. [Google Scholar] [CrossRef]
- Stelinski, L.; Boina, D.; Meyer, W. Oviposition marking behavior of Diachasma alloeum, (Hymenoptera: Braconidae), parasitizing Rhagoletis pomonella (Diptera: Tephritidae). J. Insect Behav. 2010, 23, 419–430. [Google Scholar] [CrossRef]
- Boivin, G.; Brodeur, J. Intra- and inter-specific interactions among parasitoids. In Trophic and Guild Interactions in Biological Control.; Brodeur, J., Boivin, G., Eds.; Springer: The Netherlands, 2006; pp. 124–144. [Google Scholar]
- Wang, X.; Messing, R. Intra- and interspecific competition by Fopius arisanus and Diachasmimorpha tryoni (Hymenoptera: Braconidae), parasitoids of tephritid fruit flies. Biol. Control. 2003, 27, 251–259. [Google Scholar] [CrossRef]
- Cancino, J.; Liedo, P.; Ruiz, L.; López, G.; Montoya, P.; Barrera, J.; Sivinski, J.; Aluja, M. Discrimination by Coptera haywardi (Hymenoptera: Diapriidae) of hosts previously attacked by conspecifics or by the larval parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae). Biocontrol Sci. Technol. 2012. [Google Scholar] [CrossRef]
- Aluja, M.; Ovruski, S.; Sivinski, J.; Córdova-García, G.; Schliserman, P.; Nuñez-Campero, S. Ecol. Entomol. 2012. submitted for publication..
- Strand, M.; Obrycki, J. Host specificity of parasitoids and predators. BioScience 1996, 46, 422–429. [Google Scholar] [CrossRef]
- Aluja, M.; Mangan, R. Fruit fly (Diptera: Tephritidae) host status determination, critical conceptual, methodological, and regulatory considerations. Annu. Rev. Entomol. 2008, 53, 473–502. [Google Scholar] [CrossRef]
- Mena-Correa, J.; Sivinski, J.; Anzures-Dadda, A.; Ramírez-Romero, R.; Gates, M.; Aluja, M. Consideration of Eurytoma sivinskii Gates and Grissell, a eurytomid (Hymenoptera) with unusual foraging behaviors, as a biological control agent of tephritid (Diptera) fruit flies. Biol. Control. 2010, 53, 9–17. [Google Scholar] [CrossRef]
- Bokonon-Ganta, A.; Ramadan, M.; Wang, X.; Messing, R. Biological performance and potential of Fopius ceratitivorous (Hymenoptera:Braconidae), an egg-larval parasitoid of tephritid fruit flies newly imported to Hawaii. Biol. Control. 2005, 33, 238–247. [Google Scholar] [CrossRef]
- Stelinski, L.; Liburd, O. Behavioral evidence for host fidelity among populations of the parasitic wasp, Diachasma alloeum (Mueseback). Naturwissenschaften. 2005, 92, 65–68. [Google Scholar] [CrossRef]
- Sivinski, J. personnel communication, 2012. USDA-ARS, Center for Medical, Agricultural and Veterinary Entomology, Gainesville, FL..
- Snowball, G.; Lukins, R. Status of introduced parasites of Queensland fruit fly (Strumeta tryoni), 1960–1962. Aust. J. Agr. Res. 1964, 15, 586–608. [Google Scholar] [CrossRef]
- Spinner, J.; Cowling, A.; Gurr, G.; Jessup, A.; Reynolds, O. Parasitoid fauna of Queensland fruit fly, Bactrocera tryoni Froggart (Diptera: Tephritidae) in inland New South Wales, Australia and its potential use in augmentative biological control. Aust. J. Entomol. 2011, 50, 445–452. [Google Scholar] [CrossRef]
- Darby, H.; Knapp, E. Studies on the Mexican Fruit Fly, Ananstrepha ludens (Loew). USDA, Tech. Bull. 1934, #444. [Google Scholar]
- Wang, X.; Levy, K.; Son, Y.; Johnson, M; Daane, K. Comparison of the thermal performance between a population of the olive fruit fly and its co-adapted parasitoids. Biol. Control. 2012, 60, 247–254. [Google Scholar] [CrossRef]
- Eitam, A.; Sivinski, J.; Holler, T.; Aluja, M. Biogeography of braconid parasitoids of the Caribbean fruit fly [Diptera: Tephritidae] in Florida. Ann. Entomol. Soc. Am. 2004, 97, 928–939. [Google Scholar] [CrossRef]
- Aluja, M.; Lopez, M.; Sivinski, J. Ecological evidence for diapause in four native and one exotic species of larval-pupal fruit fly (Diptera: Tephritidae) parasitoids in tropical environments. Ann. Entomol. Soc. Am. 1998, 91, 821–833. [Google Scholar]
- Sivinski, J.; Piňero, J.; Aluja, M. The distributions of parasitoids (Hymenoptera) of Anastrepha fruit flies (Diptera: Tephritdae) along an altitudinal gradient in Veracruz, Mexico. Biol. Control. 2000, 18, 258–269. [Google Scholar] [CrossRef]
- Sivinski, J.; Holler, T.; Pereira, R.; Romero, M. The thermal environment of immature Caribbean fruit flies, Anastrepha suspensa (Diptera: Tephritidae). Fla. Entomol. 2007, 90, 347–357. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M.; Piñero, J.; Ojeda, M. Novel analysis of spatial and temporal patterns of resource use in a group of tephritid flies of the genus Anastrepha. Ann. Entomol. Soc. Am. 2004, 97, 504–512. [Google Scholar] [CrossRef]
- Force, D. Ecology of insect host-parasitoid communities. Science 1974, 184, 624–632. [Google Scholar]
- Heimpel, G.; Casa, J. Parasitoid foraging and oviposition in the field. In Behavioral Ecology of Insect Parasitoids from Theoretical Approaches to Field Applications; Wajnberg, É., Bernstein, C., van Alphen, J., Eds.; Blackwell Pub: Malden, MA, 2008; pp. 51–70. [Google Scholar]
- Purcell, M.; Herr, J.; Messing, R.; Wong, T. Interactions between argumentatively released Diachasmimorpha longicaudata (Hymenoptera: Braconidae) and a complex of opiine parasitoids in a commercial guava grove. Biol. Contr. Sci. Technol. 1998, 8, 139–151. [Google Scholar]
- Sivinski, J.; Calkins, C.; Baranowski, R.; Harris, D.; Brambila, J.; Diaz, J.; Burns, R.; Holler, T.; Dodson, G. Suppression of a Caribbean fruit fly, Ananstrepha suspensa (Loew) (Diptera: Tephritidae) population through augmentative release of the parasitoid Diachasmimorpha longicaudata (Ashmead) (Hymenoptera; Braconidae). Biol. Control. 1996, 6, 177–185. [Google Scholar] [CrossRef]
- Turlings, T.; Wäckers, F.; Vet, L.; Jewis, W.; Tumlinson, J. Learning of host-finding cues by hymenopterous parasitoids. In Insect Learning; Papaj, D., Lewis, A., Eds.; Chapman and Hall: London, 1993; pp. 51–78. [Google Scholar]
- Dukas, R.; Duan, J. Potential fitness consequences of associative learning in a parasitoid wasp. Behav. Ecol. 2000, 11, 536–543. [Google Scholar] [CrossRef]
- Canale, A.; Benelli, G. Impact of mass-rearing on the host seeking behavior and parasitism by the fruit fly parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae). J. Pest Sci. 2012, 85, 65–74. [Google Scholar] [CrossRef]
- Stouthammer, R. The use of sexual versus asexual wasps in biological control. Entomophaga 1993, 38, 3–6. [Google Scholar] [CrossRef]
- Tagami, Y.; Miura, K.; Stouthamer, R. How does infection with parthenogenesis-inducing Wolbachia reduce fitness of Trichogramma? J. Invertebr. Pathol. 2001, 78, 267–271. [Google Scholar] [CrossRef]
- Silva, I.; Van Meer, M.; Roskam, M.; Hoogenboom, A.; Gort, G.; Stouthhammer, R. Biological control potential of Wolbachia-infected versus uninfected wasps: laboratory and greenhouse evaluation of Trichogramma cordubensis and T. deion strains. Biol. Control Sci. Technol. 2000, 10, 223–238. [Google Scholar]
- Ramirez-Romero, R.; Sivinski, J.; Copeland, C.; Aluja, M. Are individuals from thelytokous and arrhenotokous populations equally adept as biocontrol agents? Orientation and host searching behavior of a fruit fly parasitoid. BioControl 2012. [Google Scholar] [CrossRef]
- Davies, A.; Sivinski, J.; Aluja, M. Biol. Cont. 2012. submitted for publication..
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The endosymbiotic bacterium Wolbachia induces resistance to dengue virus in Aedes aegypti. Plos Pathog. 2010, 6, e1000833. [Google Scholar] [CrossRef]
- Messing, R.; Wang, X. Competitor-free space mediates non-target impact of an introduced bniological control agent. Ecol. Entomol. 2009, 34, 107–113. [Google Scholar] [CrossRef]
- Messing, R.; Roitberg, B.; Broduer, J. Measuring and predicting indirect impacts of biological control: competition, displacement and secondary interactions. In Environmental Impacts of Invertebrates for Biological Control of Arthropods; Methods and Risk Assessment; Bigler, F., Babendreier, D., Kuhlmann, U., Eds.; CABI Pub: Cambridge, MA, 2006; pp. 64–77. [Google Scholar]
- Vorsino, A.; Wieczorek, A.; Wright, M.; Messing, R. Using evolutionary tools to facilitate the prediction and prevention of host-based differentiation in biological control: A review and perspective. Ann. Appl. Biol. 2012. [Google Scholar] [CrossRef]
- Sivinski, J.; Vulinec, K.; Menezes, E.; Aluja, M. The bionomics of Coptera haywardi (Ogloblin) (Hymenoptera : Diapriidae) and other pupal parasitoids of tephritid fruit flies (Diptera). Biol. Control. 1998, 11, 193–202. [Google Scholar] [CrossRef]
- Forbes, A.; Satar, S.; Hamerlinck, G.; Nelson, A.; Smith, J. DNA barcoding and targeted sampling methods identify a new species and cryptic patterns of host specialization among North American Coptera (Hymenoptera: Diapriidae). Ann. Entomol. Soc. Am. 2012, 105, 608–612. [Google Scholar] [CrossRef]
- Wang, X.; Messing, R.; Bautista, R. Competitive superiority of early acting species: a case study of opiine fruit fly parasitoids. Biol. Control. Sci. Technol. 2003, 13, 391–402. [Google Scholar]
- Aluja, M.; Sivinski, J.; Van Driesche, R.; Anzures-Dadda, A. Biol. Cont. 2012. submitted for publication..
- Sivinski, J.; Wahl, D.; Holler, T.; Al Dobai, S.; Sivinski, R. Conserving natural enemies with flowering plants: Estimating floral attractiveness to parasitic Hymenoptera and attraction's relationship to flower and plant morphology. Biol. Control. 2011, 58, 208–214. [Google Scholar] [CrossRef]
- Hoelmer, K.; Kirk, A. Selecting biological control agents against arthropod pests: can science be improved to decease the risk of releasing ineffective agents? Biol. Control. 2005, 34, 255–264. [Google Scholar] [CrossRef]
- Baeza-Larios, G.; Sivinski, J.; Holler, T.; Aluja, M. The effects of chilling on the fecundity and life span of mass-reared parasitoids (Hymenoptera: Braconidae) of the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae). Biol. Cont. Sci. Technol. 2002, 12, 205–215. [Google Scholar]
- Sivinski, J.; Jeronimo, J.; Holler, T. Development of aerial releases of Diachasmimorpha tyroni (Cameron) (Hymenoptera: Braconidae), a parasitoid that attacks the Mediterranean fruit fly, Ceratitis capitata (Wiedemann) (Diptera: Tephritidae) in the Guatemalan highlands. Biol. Cont. Sci. Technol. 2000, 10, 15–25. [Google Scholar]
- Aluja, M.; Sivinski, J.; Ovruski, S.; Guillén, L.; López, M.; Cancino, J.; Torres-Anaya, A.; Gallegos-Chan, G.; Ruíz, L. Colonization and domestication of seven species of native New World hymenopterous larval-prepupal and pupal fruit fly [Diptera: Tephritidae] parasitoids. Biol. Contr. Sci. Technol. 2009, 19 (Suppl. 1), 49–29. [Google Scholar]
- Bautista, R.; Harris, E.; Vargas, R. The fruit fly parasitoid Fopius arisanus: reproductive attributes of pre-released females and the use of added sugar as a potential food supplement in the field. Ent. Exp. Appl. 2001, 101, 247–255. [Google Scholar]
- Sivinski, J.; Calkins, C.; Baranowski, R. A container for eclosion and holding of adult insects prior to mass release. Fla. Entomol. 1994, 77, 513–515. [Google Scholar] [CrossRef]
- Stuhl, C.; Cicero, L.; Sivinski, J.; Teal, P.; Lapointe, S.; Paranhos, B.; Aluja, M. Longevity of multiple species of tephritid (Diptera) parasitoids (Hymenoptera: Braconidae: Opiinae) provided exotic and sympatric-fruit based diets. J. Insect Physiol. 2011, 57, 1463–147. [Google Scholar] [CrossRef]
- Sivinski, J.; Aluja, M.; Holler, T. Food sources for adult Diachasmimorpha longicaudata, a parasitoid of tephritid fruit flies: Effects on longevity and fecundity. Ent. Exp. Appl. 2006, 118, 193–202. [Google Scholar] [CrossRef]
- Bernstein, C.; Jervis, M. Food-searching in parasitoids: The dilemma of choosing between ‘immediate’ or future fitness gains. In Behavioral Ecology of Parasitoids; Wajnberg, E., Bernstein, C., van Alphen, J., Bernstein, C., Jervis, M., Eds.; Blackwell Pub: Malden, MA, 2008; pp. 129–171. [Google Scholar]
- Narváez, A.; Cancino, J.; Canal Daza, N.; Wyckhuys, K. Effect of different dietary resources on longevity, carbohydrate metabolism, and ovarian development in two fruit fly parasitoids. Arthropod-Plant Inte. 2012, in press.. [Google Scholar]
- Wang, X.; Johnson, M.; Opp, S.; Krugner, R.; Daane, K. Honeydew and insecticide bait as competing food resources for a fruit fly and common natural enemies in the olive ecosystem. Ent. Exp. Appl. 2011, 139, 128–137. [Google Scholar] [CrossRef]
- Carrasco, M.; Montoya, P.; Cruz-Lopez, L.; Rojas, J. Response of the fruit fly parasitoid Diachasmimorpha longicaudata (Hymenoptera: Braconidae) to mango fruit volatiles. Environ. Entomol. 2005, 34, 576–583. [Google Scholar] [CrossRef]
- Quimo, G.; Walter, G. Swarming, delayed sexual maturation of males, and mating behavior of Fopius arisanus (Sonan) (Hymenoptera: Braconidae). J. Insect Behav. 2000, 13, 797–813. [Google Scholar] [CrossRef]
- Sivinski, J. unpublished work, 1995; USDA-ARS, Center for Medical, Agricultural and Veterinary Entomology, FL.
- Sivinski, J.; Petersson, E. Mate choice and species isolation in swarming insects. In The Evolution of Mating Systems in Insects and Arachnids; Choe, J., Crespi, B., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 294–309. [Google Scholar]
- Ramadan, M.; Wong, T.; Wong, M. Influence of parasitoid size and age on male mating success of Opiinae (Hymenoptera: Braconidae), larval parasitoids of fruit flies (Diptera: Tephritidae). Biol. Control. 1991, 1, 248–255. [Google Scholar] [CrossRef]
- Hagen, K. A premating period in certain species of the genus Opius (Hymenoptera: Braconidae). P. Hawaii Entomol. Soc. 1953, 15, 115–116. [Google Scholar]
- Williams, H.; Wong, M.; Wharton, R.; Vinson, S. Hagen’s gland morphology and chemical content analysis for three species of parasitic wasps (Hymenoptera: Braconidae). J. Chem. Ecol. 1988, 14, 1727–1736. [Google Scholar] [CrossRef]
- Buckingham, G.; Sharkey, M. Abdominal exocrine glands in Braconidae (Hymenoptera). Adv. Parasit. Hymenopt. Res. 1988, 199–242. [Google Scholar]
- Sivinski, J.; Epsky, N. unpublished work, 2000; USDA-ARS, Center for Medical, Agricultural and Veterinary Entomology, Gainesville, FL.
- Boush, G.; Baerwald, R. Courtship behavior and evidence for a sex pheromone in the apple maggot parasite Opius alloeus [Hymenoptera: Braconidae]. Ann. Entomol. Soc. Am. 1967, 60, 805–806. [Google Scholar]
- Canale, A.; Benelli, G.; Lucchi, A. Female borne cues affecting Psyttalia concolor (Hymenoptera, Braconidae) male behavior during courtship and mating. Insect Sci. 2012. [Google Scholar] [CrossRef]
- Sivinski, J.; Webb, J.C. Acoustic signals produced during courtship in Diachasmimorpha (= Biosteres) longicaudata (Hymenoptera: Braconidae) and other Braconidae. Ann. Entomol. Soc. Am. 1989, 82, 116–120. [Google Scholar]
- Knipling, E. Principles of Insect Parasitism Analyzed from New Perspectives. In Agriculture Handbook—United States Department of Agriculture # 693; United States Department of Agriculture: Washington, D.C, 1995. [Google Scholar]
- Cáceres, C.; McInnis, D.; Shelly, T.; Jang, E.; Robinson, A.; Hendrichs, J. Quality management systems for fruit fly (Diptera: Tephritdae) Sterile Insect Technique. Fla. Entomol. 2007, 90, 1–9. [Google Scholar] [CrossRef]
- Joyce, A.; Aluja, M.; Sivinski, J.; Vinson, B.; Ramirez-Romero, R.; Bernal, J.; Guillen, L. Effect of continuous rearing on courtship acoustics of five braconid parasitoids, candidates for augmentative biological control of Anastrepha species. BioControl 2010, 55, 573–582. [Google Scholar] [CrossRef]
- Benelli, G.; Bonsignori, G.; Stefanini, C.; Canale, A. Courtship and mating behavior in the fruit fly parasitoid Psyttalia concolor (Szépligeti) (Hymenoptera: Braconidae): The role of wing fanning. J. Pest Sci. 2012, 85, 55–63. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Sivinski, J.; Aluja, M. The Roles of Parasitoid Foraging for Hosts, Food and Mates in the Augmentative Control of Tephritidae. Insects 2012, 3, 668-691. https://doi.org/10.3390/insects3030668
Sivinski J, Aluja M. The Roles of Parasitoid Foraging for Hosts, Food and Mates in the Augmentative Control of Tephritidae. Insects. 2012; 3(3):668-691. https://doi.org/10.3390/insects3030668
Chicago/Turabian StyleSivinski, John, and Martin Aluja. 2012. "The Roles of Parasitoid Foraging for Hosts, Food and Mates in the Augmentative Control of Tephritidae" Insects 3, no. 3: 668-691. https://doi.org/10.3390/insects3030668