1. Introduction
Natural river floodplains consist of complex habitats differing in hydrological connectivity, which affects fish community dynamics [
1,
2,
3]. In the temperate climate zone, large undisturbed floodplains have often been disrupted by channel regulation that exerted direct and indirect effects on habitat heterogeneity, successional trajectories and, ultimately, the ecological integrity of rivers [
4,
5].
The functional feature of intact alluvial floodplains of meandering rivers is a mosaic of lotic and lentic ecosystems, including the river and its side channels, tributary streams and cut-off channels. Due to the variation in the connectivity, laterally across the floodplain, a distinct zonation of the habitats has been widely reported both for tropical [
6,
7,
8] and temperate rivers [
9,
10]. Differences in the connection of floodplain lakes with the river channel determine the availability of nutrients and the degree to which processes such as primary productivity and decomposition are controlled by the river [
11,
12,
13]. Floodplain ecosystems connected to the river are particularly open to exchange of matter with the river which leads to higher concentrations of macroelements, while ecosystems isolated from the nearby river for most of the year acquire a lentic character that promotes autogenic, mainly organic, matter cycling. An increased flow rate throughout bi-connected water bodies is beneficial to organisms in that it transports food, oxygen, nutrients, and particulate and dissolved organic matter [
14,
15].
The hydrological integration between river and cut-off channels is a significant habitat parameter for species that require different aquatic microhabitats in the course of their life cycle, e.g., certain species of fish [
4,
16,
17,
18]. Fish communities in European lowland riverine ecosystems are composed of rheophilic (require flowing water to spawn), eurytopic (habitat generalists) and limnophilic (found in stagnant and strongly vegetated floodplain water bodies) fish species guilds [
4,
19,
20] that contribute to the overall high species diversity [
4]. For example, the ide
Leuciscus idus requires flowing water habitats, whereas the crucian carp
Carassius carassius needs a single stagnant floodplain lake that exists over a long period.
Floodplain lakes are ecosystems with diverse fish species adapted to periods of low and high water stages (flood-pulse), which affect any wetland water quality parameters [
11,
21,
22,
23]. Higher water stages promote greater nutrient availability, aquatic primary production, allochthonous inputs, and secondary production, which are especially beneficial for early life stages of fish in floodplain habitats. In contrast, low-water conditions lead to the contraction of marginal aquatic habitats, decay of aquatic macrophytes, and higher densities of aquatic organisms, including phytoplankton and zooplankton in floodplain water bodies [
24]. During low water periods, non-flowing ecosystems have been recognized as having limited conditions for light penetration and thereby limited photosynthesis. In turn, summer oxygen deficits are attributed to shading by emergent and floating vegetation, high biological oxygen demand and limited aeration [
25].
The natural hydrological regime is one of the key drivers of ichthyofauna development [
2,
8]. Floodplain lakes typically serve as nursery for young fish whereas adults live in the main channel or connected side arms [
4,
18]. Welcomme and Halls [
8] observed that extensive flooding increased the area available for spawning sites and provided fish with more food and better shelter opportunities, whereas the duration of the flood influenced the time during which fish could grow and find shelter from predators. In several studies, fish migrated to floodplain water bodies, in particular to lentic habitats, in search of refuge during floods [
26,
27,
28]. When water levels drop, fish either migrate back to the river and become a source of food for resident piscivores or remain in isolated floodplain water bodies [
2,
6,
29].
An increase in the reductions in landscape connectivity, ecological functioning and ecosystem biodiversity has driven initiatives to improve the ecological status of rivers, e.g., the European Union Water Framework Directive (2000/60/EEC) [
30], and to protect biological diversity, e.g., the Habitats Directive (92/43/EEC) [
31] and Agenda 21 of the Rio Convention and the Convention on Biological Diversity. According to Welcomme
et al. [
20], fish environmental guilds could be used as a tool for assessing the ecological status of rivers. Further, the knowledge of differences in the responses of functional fish groups to environmental factors is useful for predicting the effects of future environmental manipulations (e.g. changes in hydrology and connectivity) on fish communities in various aquatic systems. Achieving good ecological status by promoting fish abundance and diversity involves the creation of habitats that are functionally similar to natural lowland river-floodplain ecosystems. Nevertheless, this approach requires a relevant reference area to test whether environmental parameters are affecting the qualitative and quantitative structure of hydrobionts. The Biebrza river provides an opportunity to realize this test as it is an undisturbed system presenting variable levels of lateral connectivity with sequential shift in fish community composition from rheophilic to eurytopic to limnophilic fish species guilds, as it was reported
ca. 30 years ago by Witkowski [
14,
32]. Moreover, unlike many rivers in Europe, the floodplain lakes in the middle and lower section of the Biebrza River have not been disturbed by hydraulic structures, excessive nutrients or sediments introduced by runoffs from the surrounding farmland.
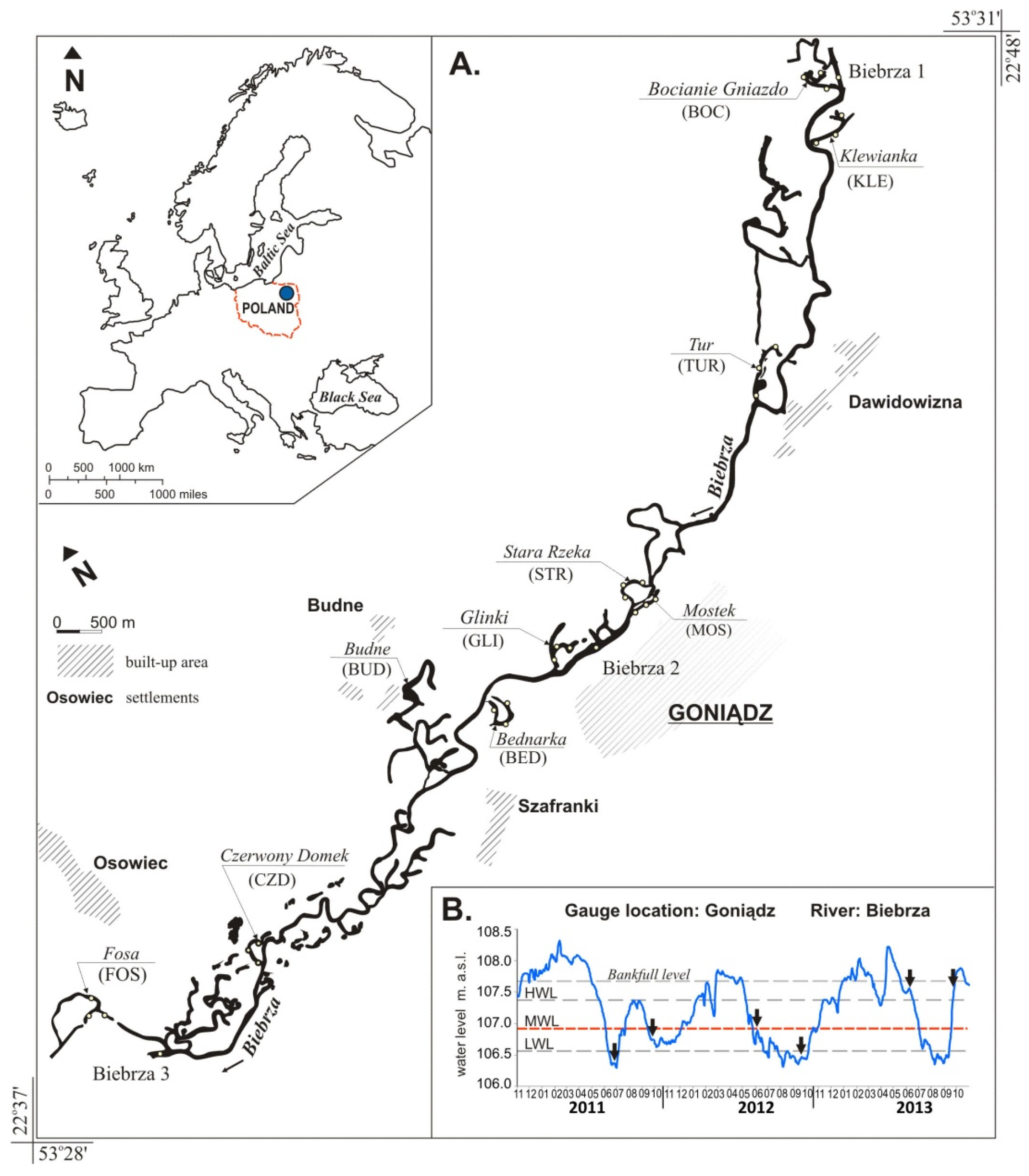
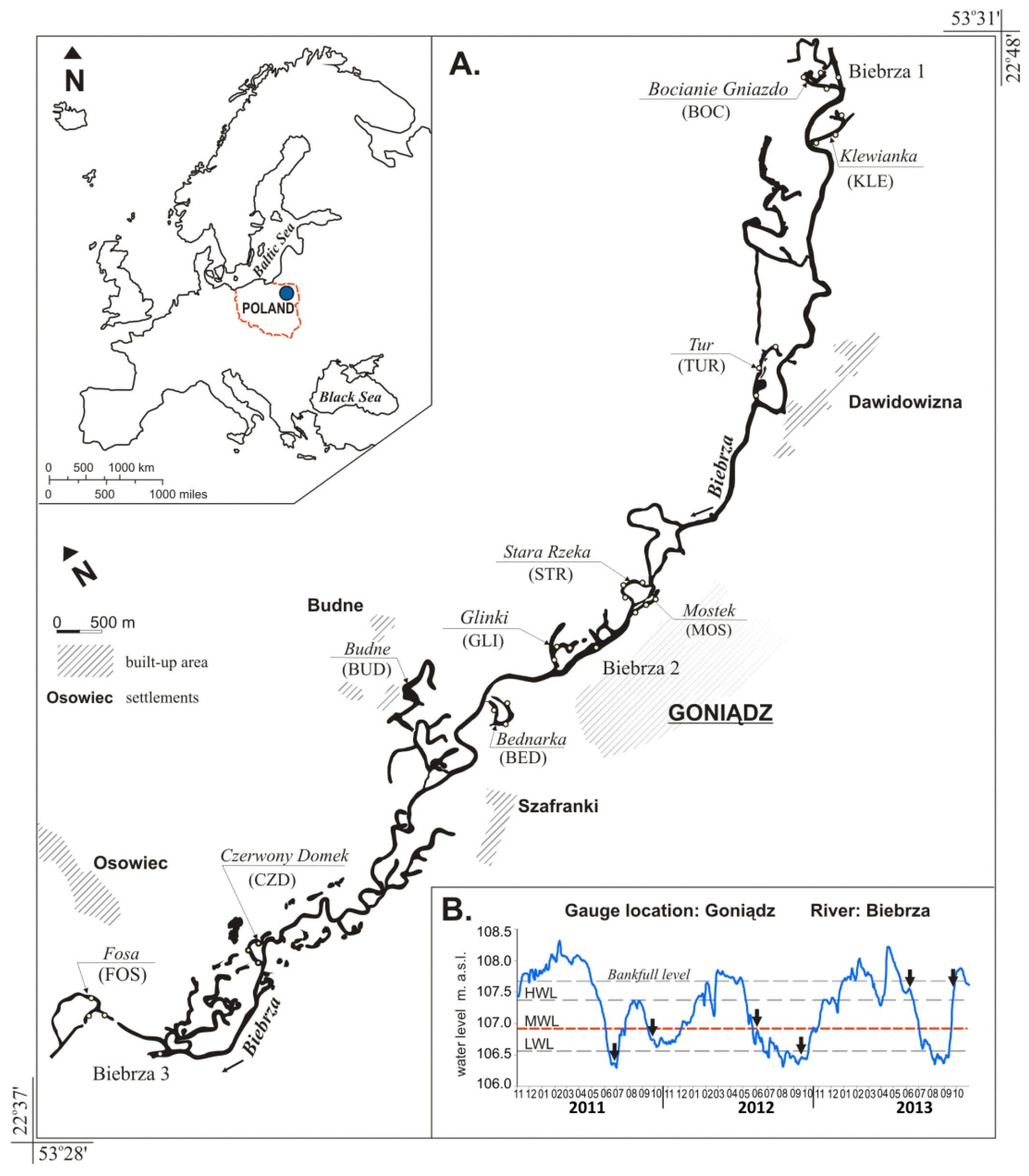
The aim of this study was to determine whether lateral connectivity and environmental parameters are influencing the qualitative and quantitative structure of fish communities in floodplain lakes. For this purpose, we sampled 10 natural floodplain lakes and the river channel in the middle section of the Biebrza River, (NE Poland) depending on their connectivity and habitat diversity.
3. Results
3.1. Water Quality
The surveyed floodplain lakes differed significantly in size, depth, range of water level fluctuations and the gradient of lateral connectivity with the adjacent river channel, which produced a dataset with a broad range of physicochemical parameters (
Table 2). Water mineralization was average with mean electrolytic conductivity of 542 ± 110 μS·cm
−1. Lower values in the range represented lotic habitats, and higher values were associated with lentic lakes. The pH of water was slightly basic (pH 7.77), and it decreased significantly when water levels were high, in particular in parapotamic water bodies (
r = −0.62;
p = 0.001). Floodplain lakes were generally abundant in organic matter with an average TOC content estimated at 12.59 ± 4.64 mg·L
−1 and a significant share of DOC (10.50 ± 3.18 mg·L
−1). In addition to autogenic organic matter, the accumulation of humic compounds and decomposed organic matter from adjacent peatlands significantly contributed to an increase in DOC in isolated lakes (48.92 ± 21.50 mg·L
−1). High concentrations of suspended solids in paleopotamic habitats reduced water transparency to 1.1 ± 0.3 m. In para- and plesiopotamic lakes, Secchi disc visibility was significantly higher at 1.5 ± 0.6 and 2.0 ± 0.9 m, respectively. Overall productivity of the ecosystems was related to the concentrations of chlorophyll-a, which, in paleopotamic and plesipotamic lakes, was twice as much (~17.50 μg·L
−1) as in parapotamic habitats (8.04 ± 4.03 μg·L
−1) or in the river (7.31 ± 0.80 μg·L
−1).
The mean concentrations of total phosphorus in the floodplain lakes were determined at 0.35 mg·L−1 and no significant differences were observed between the evaluated water bodies. Phosphate phosphorus levels were higher in parapotamic lakes (0.12 ± 0.05 mg·L−1; post-hoc Duncan’s test, ANOVA; p ≤ 0.05) than in paleopotamic lakes (0.07 ± 0.04 mg L−1). Total nitrogen concentrations increased along the isolation gradient from 1.07 mg·L−1 in parapotamic lakes to 1.39 mg·L−1 in paleopotamic lakes. The content of total inorganic nitrogen (TIN= NO2–N + NO3–N + NH4–N) was similar among the studied floodplain lakes (0.34–1.55 mg·L−1), but the contribution of each N-form differed significantly relative to water retention time: ammonium nitrogen was the predominant form in stagnant water of paleopotamic habitats (69%), whereas nitrate nitrogen in flowing water in parapotamic water bodies (62%).
The correlations between TP and TN concentrations vs. water levels varied across the examined lakes (correlations are not shown in the tables). TP values were positively correlated with water levels in plesiopotamic (r = 0.41, p = 0.02) and parapotamic lakes (r = 0.45, p = 0.02), but no such correlations were observed in paleopotamic lakes. Unlike TP, the rise in water levels significantly reduced TN concentrations in all lake types (r = −0.58; p = 0.001). DO was significantly higher and more stable in parapotamic lakes (7.45–1.54 mg·L−1; post-hoc Duncan’s test, ANOVA; p ≤ 0.05) than in plesiopotamic (6.51–2.12 mg·L−1) and paleopotamic habitats (6.13–2.31 mg·L−1).
3.2. Fish Assemblages
A total of 18,399 fish from 23 species belonging to 6 families were sampled but the number of specimens varied significantly across species. Cyprinidae was the most abundant and diverse family represented by 15 species (
Table 3). The family Cobitidae was represented by three species, Percidae by two species and the families Esocidae, Siluridae and Gadidae by one species each. The roach
R. rutilus define and the define rudd
S. erythrophthalmus were eudominant species that accounted for 36% and 12% of all captured specimens, respectively. The pike
E. lucius, bitterling
Rhodeus sericeus amarus, silver bream
Blicca bjoerkna and sunbleak
L. delineates were dominants. Other species, including the perch
Perca fluviatilis, Prussian carp C.
auratus gibelio, tench
Tinca tinca, bream
Abramis brama and bleak
Alburnus alburnus, were less abundant (3%–5%), but they had a stable share of the assemblage.
Among eurytopic fish, the roach R. rutilus and the pike E. lucius were encountered most frequently (euconstants found in >75% of the sites). Other eurytopic constant species (50% < occurrence < 75%) were the perch and silver bream, as well as the rudd, bitterling and tench in the group of limnophilic fish. Rheophilic species were far less abundant and amounted to 2.3% in plesiopotamic, 3.1% in paleopotamic and 8.2% in parapotamic water bodies.
Amount of rheophils in lotic habitats was 50% lower than that in the Biebrza river channel. Along the lateral connectivity gradient of parapotamic–plesiopotamic–paleopotamic lakes, the proportions of rheophilic species were determined at 10:5:1, and limnophilic species at 1:2:5.
Five out of the 23 identified species have been placed on the IUCN Red List of Threatened Species, and four are listed in Annex II to the EU Habitat Directive (92/43/EEC). They include three rheophilic species: the asp
Aspius aspius, spined loach
Cobitis taenia, and stone loach
Barbatula barbatula and two limnophylic taxa: weatherfish
Misgurnus fossilis and the bitterling
R. sericeus amarus. The above species had less than 2.5% share of the fish assemblage, excluding the bitterling, which was a dominant and frequently observed species. The Prussian carp was the only non-native species identified in the study. However, the study of van Damme
et al. [
42] showed evidence of the bitterling as non-native for the Polish watercourses.
As many as six species colonizing the Biebrza floodplain lakes were piscivores (
Table 3 and
Table 4) that accounted for nearly 12.4% of all specimens. Pike represented 60% and perch 35.7% of the identified piscivores. Other predatory species, such as the wels catfish
Silurus glanis, burbot
Lota lota, and the chub
Squalius cephalus, were less populous.
3.3. Fish Response to Habitats along the Connectivity Gradient
The composition of fish species in parapotamic, plesiopotamic, and paleopotamic floodplain lakes relative to the location of the sampling site is presented in
Table 4.
Parapotamic lakes (lotic side-channels) were inhabited by 21 of the 23 identified fish species, and the only missing taxa were the Prussian carp and the stone loach (
Appendix A). Mean fish density in the lakes amounted to 1995 specimens per ha, which was lower in comparison with other types of lakes, and accounted for 23.9% of captured fish (
Table 4). Fish species contribution decreased between upstream arms (9.1%) and downstream arms (6.5%). The biodiversity index of parapotamic habitats was determined at H′ = 2.45, and species evenness at
J = 0.79. Fish biomass in lakes of that type amounted to 78.4 kg·ha
−1, which constitutes ~34% of total fish biomass, but it was the highest near the downstream connections with the river (85.1 kg ha
−1).
Active exchange of water in habitats provides similar conditions for ichthyofauna to those noted in the Biebrza River. Eurytops accounted for 71%, limnophils for 21%, and rheophils for only 8% of the species in parapotamic lakes. In the group of parapotamic lakes, the roach and pike were eudominants, whereas the rudd, perch and silver bream were dominants (
Appendix A). No rheophilic species were eudominants or dominants in the studied group of water bodies. Among rheophilic species, only
L. idus was a subdominant.
The ichthyofauna in plesiopotamic lakes was represented by all of the 23 identified fish species. As many as 19 species were found in both upstream and middle arms, whereas 18 taxa were observed near the connection with the river channel. The diversified habitats along plesiopotamic lakes showed decreased water exchange, aeration and depth gradients, and they were colonized by 41% of captured fish and characterized by the highest biodiversity index (H′ = 2.51). One-side connected lakes showed mean fish density determined at 2687 specimens ha−1. Fish biomass in plesiopotamic lakes amounted to 85 kg·ha−1 and was the highest (36.5% of total biomass) among the studied lakes types. Although eurytops prevailed in the species composition (72.3%), an increase in limnophils by 5%, when compared to parapotamic lakes, has been noted. The number of limnophilic taxa (rudd, bitterling and tench) increased with a distance from the river channel. Eudominants in the plesiopotamic lakes were represented by roach, rudd, and pike, while dominants by silver bream bitterling, and perch. Due to stagnant water, no psammophils were identified, but the presence of ostracophils (bitterling) and litho-pelagophils (burbot) was noted in the downstream section in the vicinity of the river connection.
Fish assemblages in paleopotamic lakes were represented by the lowest number of species (17) and, consequently, the lowest biodiversity index (H’ = 2.23) in comparison with the lakes connected to the river (
Table 4). Simultaneously, isolated water bodies were characterized by the highest evenness index (J’) of 0.82. A clear increase in the share of limnophilic species (up to 63.1%) could be attributed to lakes' isolation from the river channel. The roach was the eudominant species with the highest D index. Paleopotamic lakes provided particularly supportive habitats for sunbleak and Prussian carp (
Appendix A). Among dominant species were pike, rudd, bitterling and tench. The number of specimens captured was almost 50% higher than that caught in bi-connected side-channels. Fish density reached 2936 specimens·ha
−1. Despite high fish abundance, fish biomass in isolated habitats was lower (70 kg·ha
−1, ~29% of total) than in lakes connected to the river channel.
3.4. Influence of Environmental Factors on Fish Abundance and Diversity
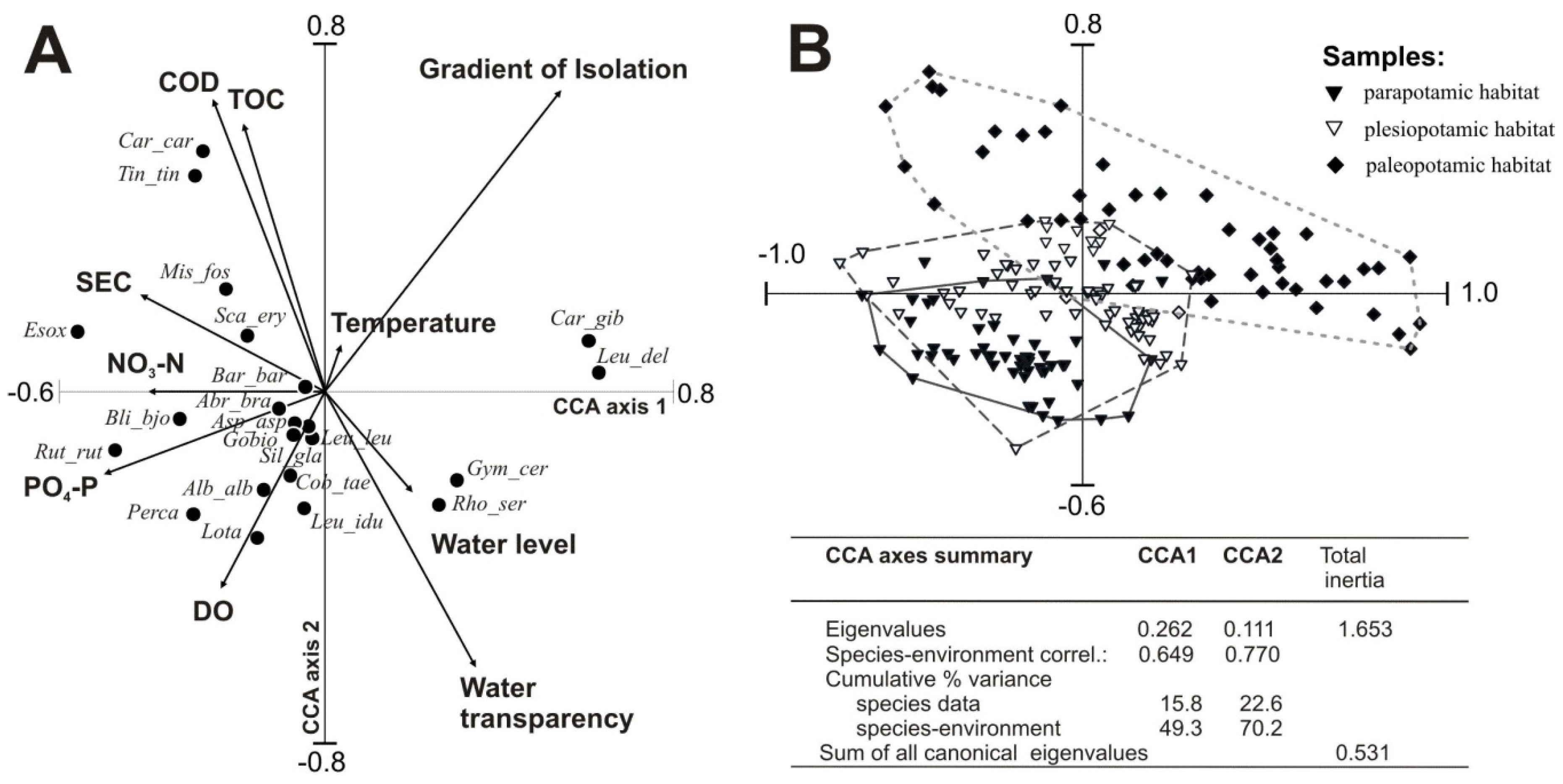
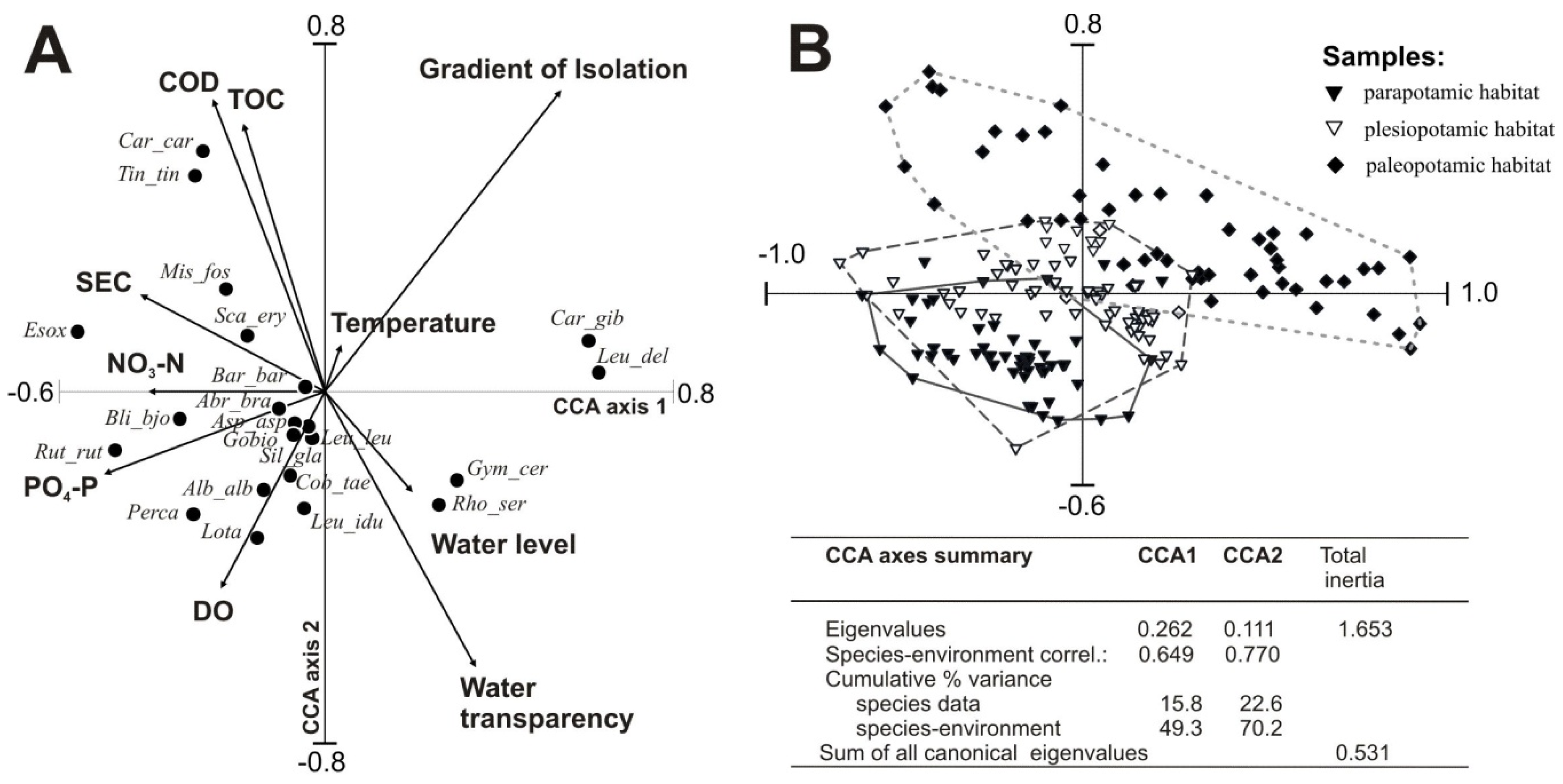
Three types of floodplain lakes (parapotamic, plesiopotamic, and paleopotamic), classified based on hydrological connectivity, differed significantly in physicochemical parameters of water and fish responses to varied habitat conditions. The canonical correspondence analysis (CCA) biplot (
Figure 2A) demonstrates the correlations between environmental factors as lateral connectivity, water level, water quality parameters and the abundance of fish fauna in the studied floodplain lakes. The results of the CCA (
Figure 2A) showed that eigenvalues of the first (λ
CC1 = 26.2%) and second (λ
CC2 = 11.1%) CCA axes accounted for 70.2% of the cumulative variation in the environmental data. The final model accounted for 22.6% of the total variance in fish composition and all canonical axes were significant (Monte Carlo test,
p = 0.002).
Ten out of 17 input variables were retained as significant contributors to the CCA model (
Table 5). The non-retained seven were redundant or did not increase the significance. The hydrological set of variables (including isolation gradient and water-level variability) accounted for λ1 = 18% of the species variability, while retained physical and chemical variables, including SEC, DO, COD
Cr, temperature and TOC explained in total 36%. Trophic variables (NO
3–N and PO
4–P) explained 11% of the variability.
The ordination space of factors CCA1 and CCA2 were clearly distinguished by environmental variables and fish species composition, confirming previous analyses. The first factor (CCA1) showed a gradient from highly eutrophicated lentic sites to less eutrophicated parapotamic sites (
Figure 2A). It correlated negatively with NO
3–N, SEC and PO
4–P while positively with water level. Organic matter content, expressed by COD
Cr and TOC, was positively correlated, whereas DO and water transparency were negatively correlated with CCA2 (20.9% of the variance). Significant variance explained by the lake isolation gradient (λ1 = 15%) was confirmed by three clusters of samples comprising parapotamic, plesiopotamic, and paleopotamic habitats (
Figure 2B). Lakes situated near the river with transparent and well-aerated waters were characterized by a predominance of rheophils, including the burbot, ide, wels catfish, asp, gudgeon, bleak and dace. Stagnant water habitats attracted mostly the rudd, Prussian carp, crucian carp, tench and weatherfish.
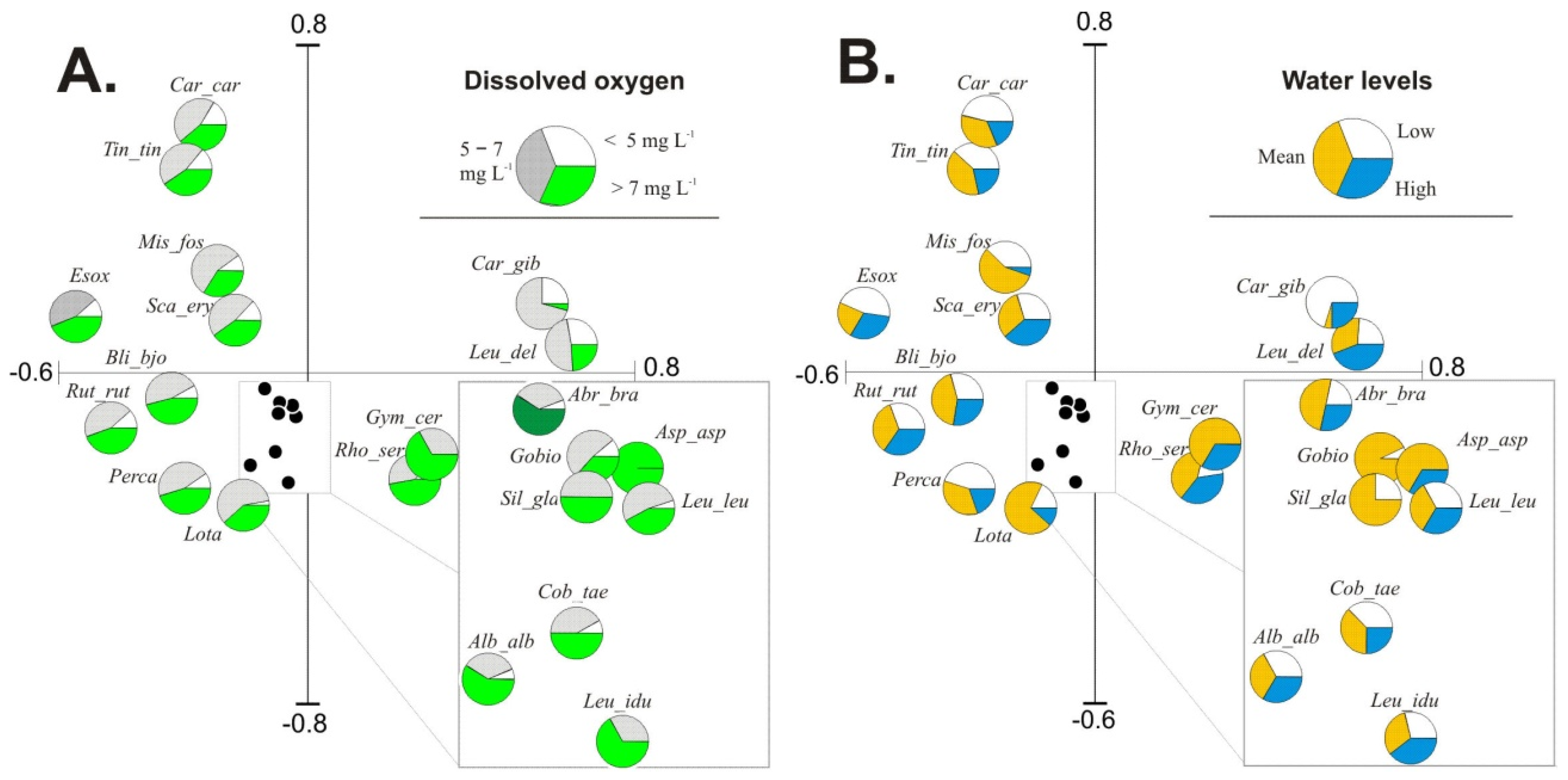
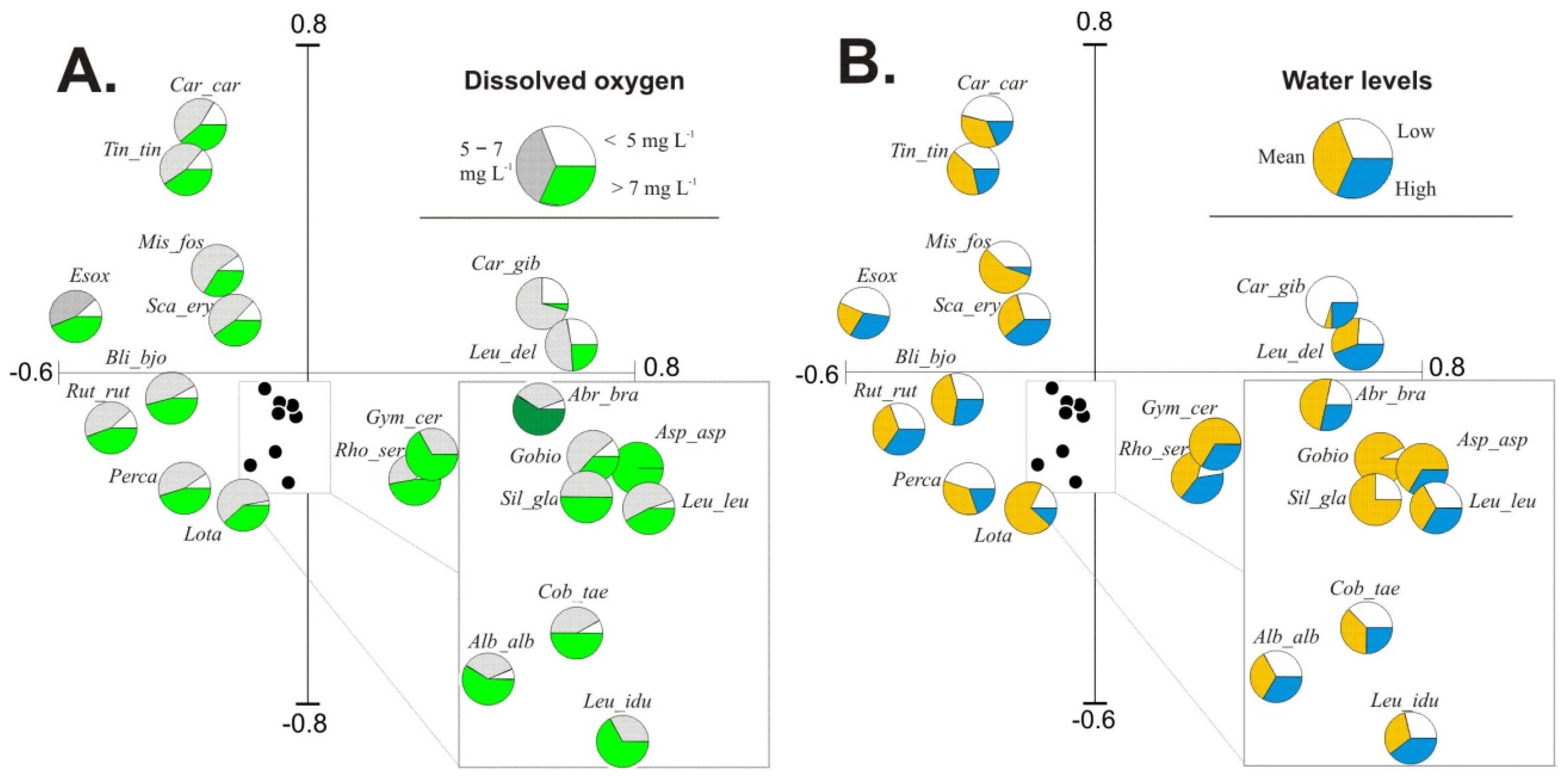
Forward selection revealed that in the group of input environmental variables, hydrochemical parameters were significant in explaining fish occurrence and abundance patterns in the studied floodplain lakes. White bream and pike were related to habitats with higher NO
3–N concentrations and lower water levels. Limnophilic species (crucian carp and tench) preferred habitats with higher COD and TOC and lower DO content, which are characteristic of lakes isolated from the river. In contrast, rheophilic (burbot) and eurytopic (perch and bleak) species preferred transparent and well aerated water. The availability of dissolved oxygen (DO) is the key contributor to fish yield in lakes. The highest share of fish specimens in water containing > 7 mg DO L
−1 confirmed distinct preferences of fish for inhabiting water bodies supplied with river water; this is presented in the form of species pies charts in
Figure 3A. The diagrams of pies charts based on water levels, as presented in
Figure 3B, showed that water stage is a significant factor that changes the quantitative structure of fish species in floodplain lakes.
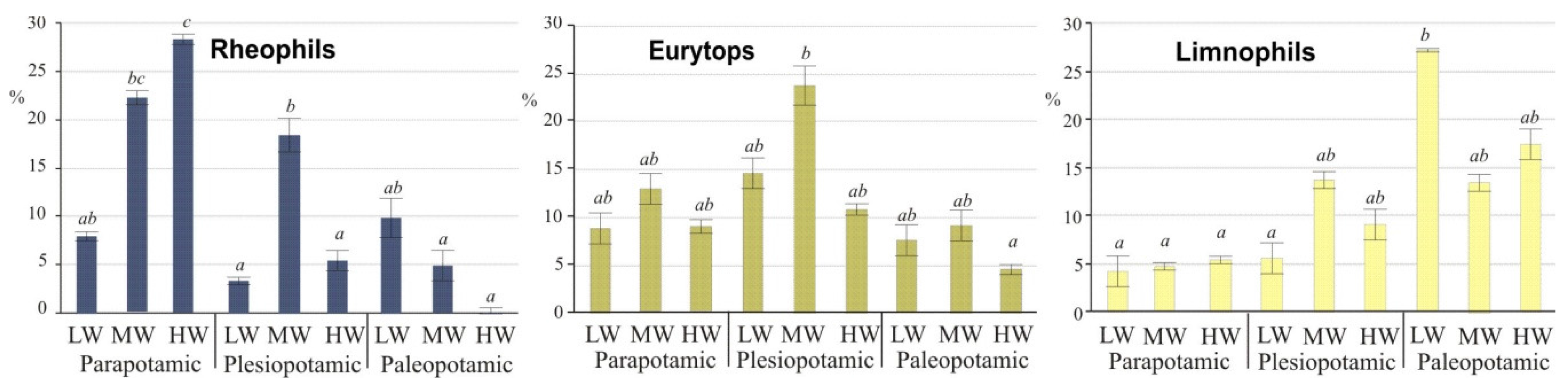
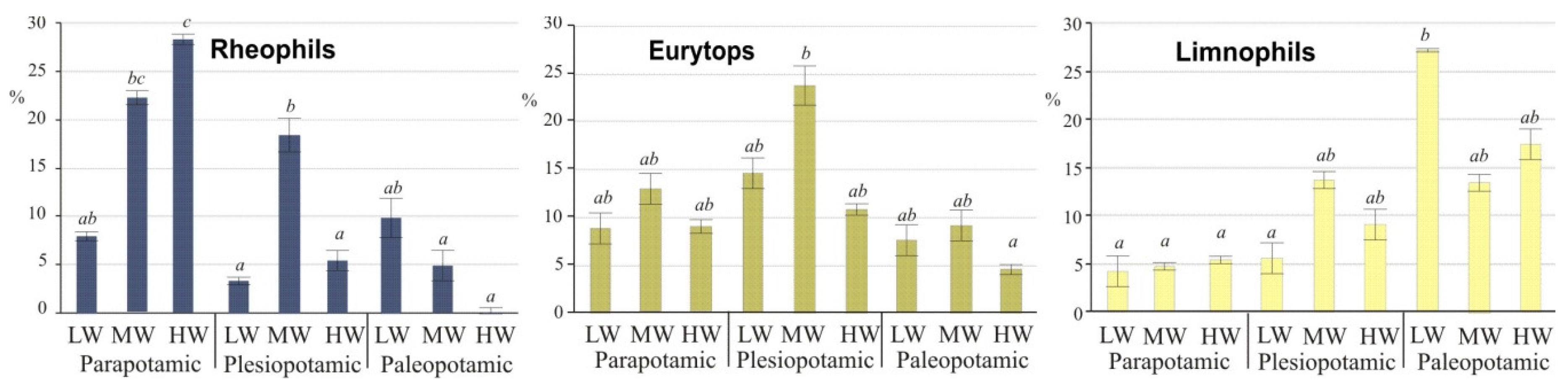
As typical inhabitants of isolated lakes, limnophils differed significantly (ANOVA,
post-hoc Duncan’s test,
p ≤ 0.05) from other guilds during low water stages (
Figure 4). In the group of rheophilic species, the chub predominated at low water (50% of specimens), mainly in lakes of high water exchange. High water (
potamophase) did not contribute to an increase in species abundance. Only two species, the ide and asp, adapted to higher water stages, and their share during that period reached 65% and 60%, respectively (
Figure 3A). The above taxa contributed the group of rheophils that were characterized by significant differences (ANOVA,
post-hoc Duncan’s test,
p ≤ 0.05) during high water. Eurytops were more opportunistic and had no water stage preferences, but their share in lentic lakes decreased during floods (
Figure 4).
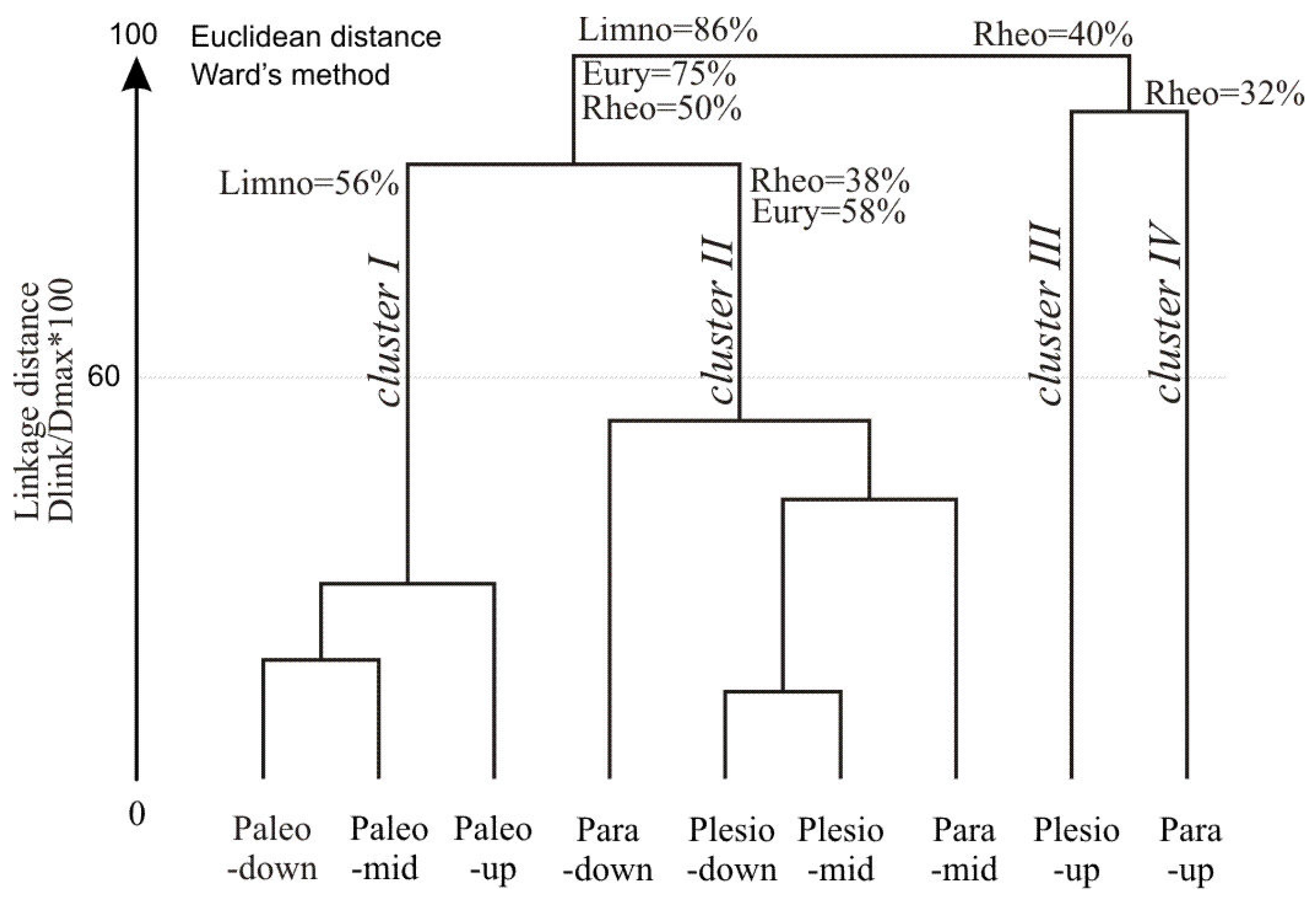
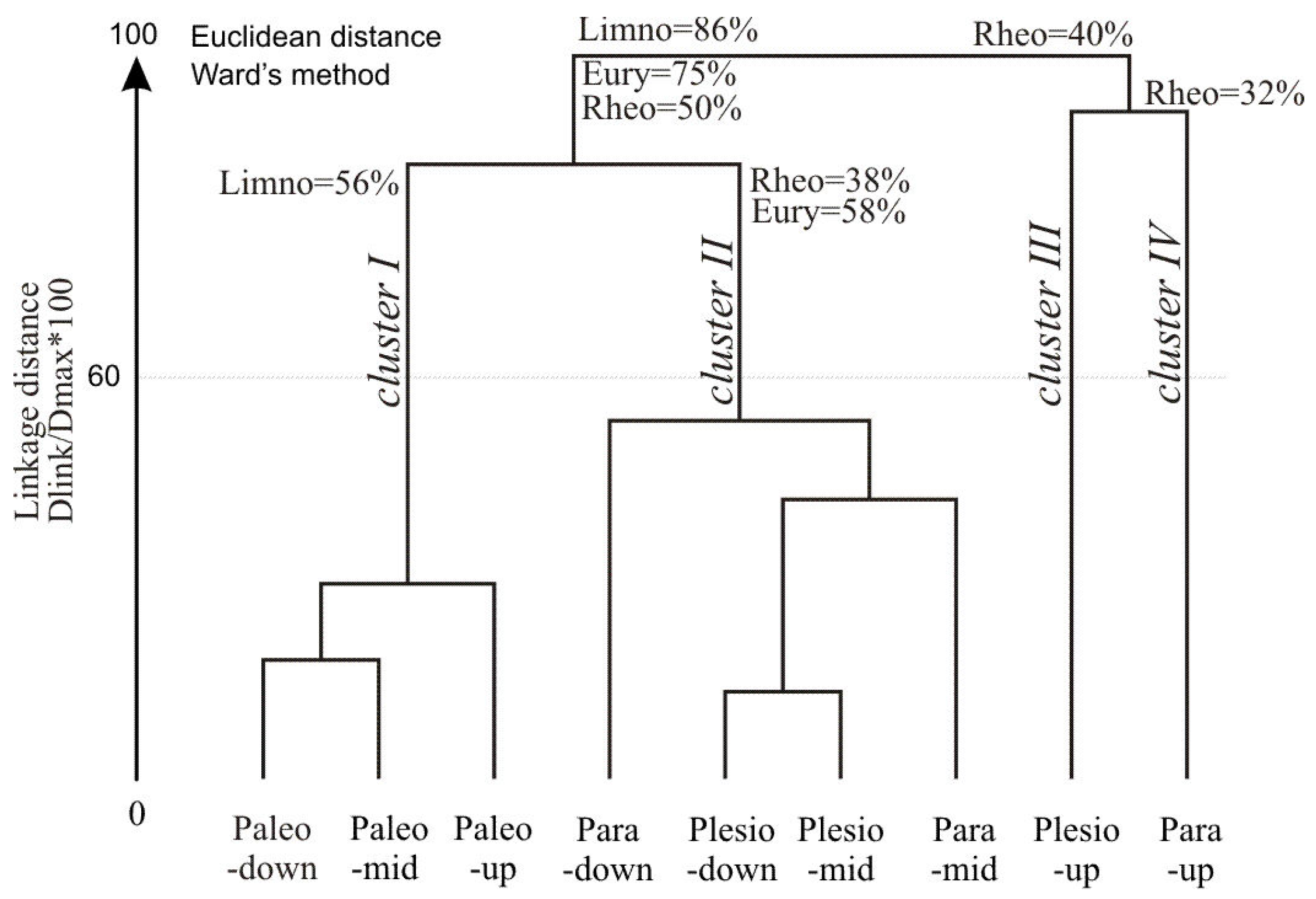
Hierarchical cluster analysis (HCA) performed on the percentage of species abundance in nine lake-sites produced four clusters of objects (
Figure 5). Cluster I covered lentic sites with a high percentage of limnophils (56%) and complementary species, mainly eurytops. The cluster comprised the predominant species in lentic habitats: the Prussian carp, crucian carp, sunbleak, bitterling and weatherfish. Cluster II was characterized by a significant share (58%) of eurytopic species. Clusters III and IV had a similar share of rheophils (40% in total), including the asp, gudgeon, chub, dace and spine loach.
4. Discussion
The present study showed that variations in fish abundance and community structure were related to environmental variables in floodplain lakes of the Middle Basin of the Biebrza River. Furthermore, the diversity of environmental conditions in the lakes produced distinctive fish guilds. Major environmental gradients related to the structuring of fish communities involved degree of isolation from the river, water transparency, SEC, DO, COD, temperature and water levels, whereas nutrients such as PO4–P, NO3–N and TOC played a minor role.
Lateral connectivity was found to be a key driver in shaping fish assemblage attributes in the studied floodplain lakes. It promotes fish migration, and thus species exchange, when the surface pathway is maintained between river channel and water ecosystems. Kwak [
43] reported that the increase in lateral movement of fishes between the river channel and floodplain habitats is enhanced by the increasing river discharge. In the case of the Biebrza River, the fish colonization of isolated water bodies probably occurs annually in spring when frequency and duration of connection is greatest. Flood-pulse of water is an essential factor that determines nutrient cycling, which is consistent with the hypothesis postulated by, e.g., Amoros and Bornette [
44] or Junk and Watzen [
23]. The connection of lakes with the main river channel during periods of high water provides the input of well-aerated river water, which protects the lakes against severe oxygen deficits. It is a period of somewhat “deep breath”, which enables many organisms to survive in isolation, and may be crucial for maintaining their populations [
15]. During low water periods, particularly in summer, the decreasing gradient of oxygen resources is the most distinct: from optimal aeration (
ca. 7 mg DO·L
−1) in parapotamic lakes towards paleopotamic lakes with significant DO deficits (<2 mg·L
−1). For example, DO measurements performed in the summer of 2011 in vertical profiles of isolated lakes during a low water stage revealed a shortage of oxygen already 0.4 m below the water table. According to estimates, more than two-thirds of the water volume in isolated lakes could experience anaerobic conditions in summer (not shown in the figure drawing), which could have significant implications for fish populations in those ecosystems.
Lateral gradient of lake connectivity is directly related to fish densities, which increased from
ca. 2000 in lotic to 3000 specimens·ha
−1 in lentic habitats. Parapotamic lakes were inhabited by 24%, plesiopotamic by 41% and paleopotamic lakes by 35% of the evaluated fish population. Similar to the study of Winemiller
et al. [
15], we observed higher values of the biodiversity index (H′) in floodplain lakes than in the Biebrza River. In our study, H′ was higher in plesiopotamic lakes than in parapotamic and paleopotamic lakes. The evaluated lake types were arranged in the following sequence based on the values of H′: plesiopotamic > parapotamic > paleopotamic. The results of this study confirm the findings of Tockner
et al. [
21] or Guti [
9] for the Danube, where the structure of ecological fish guilds changed from rheophilic to limnophilic and the diversity of fish species decreased in the lateral transect along with the increased distance from the main river channel and a decrease in hydrological connectivity. Along the lateral connectivity gradient of parapotamic–plesiopotamic–paleopotamic lakes, the proportions of rheophilic species were determined at 10:5:1, and limnophilic species-at 1:2:5. High species diversity in the analyzed floodplain lakes resulted from the co-occurrence of eurytopic, limnophilic and rheophilic guilds.
The ichthyofauna in the Biebrza floodplain lakes was most abundant in eurytops (59% of all specimens), that live under both lotic and lentic conditions. A wide range of microhabitats in semi-lotic lakes creates greater opportunities for eurytopic species, which are capable of living in varied habitats, than lotic or lentic environments. The predominant eurytopic species were the roach, pike, rudd, bream and bleak. Long-term isolation allows for a succession of the fish assemblage towards floodplain specialist limnophilic species [
10]. Limnophilic species accounted for 37% of the evaluated fish guilds, where the Prussian carp was the eudominant, and the crucian carp, bitterling and tench were the dominants. The above species are tolerant of habitats with low oxygen resources and high organic matter content (e.g., Prussian carp) that are found in isolated water bodies. Limnophils abundance was highly correlated with water quality descriptors (COD, TOC, DOC, NH
4–N and TN) characteristic of fertile and productive ecosystems. Other variables, including pH, DO, turbidity and macrophyte cover, also influenced fish assemblages inhabiting isolated water bodies as reported in temperate floodplains [
15,
17,
45]. Adaptation to anoxic conditions of the floodplain specialists is often accompanied by poor competitive abilities and predation avoidance, resulting in low population densities and high mortalities in multispecies fish assemblages, as in crucian carp [
46]. Thus, long-term isolated floodplain lakes in advanced successional stages with a tendency to temporary anoxia provide important refuge habitats for these species [
45]. Advanced successional stages of such lakes provide most suitable habitats for still-water species and specialist species that have evolved physiological adaptations and strategies to survive hypoxic and anoxic conditions [
10]. Accordingly, limnophilic fish are essential elements of the typical fish community of floodplain aquatic ecosystems and thus can serve as an indicator of their ecological integrity. We share the opinion of Welcomme
et al [
20] that isolated lakes are at least as important as the lakes with high rates of water exchange as fish habitats. Isolated lakes offer a rich array of habitat types at different stages of succession, and they are in direct need of statutory protection.
Our findings are consistent with the results of previous studies on fish assemblage metrics among floodplain lakes. In a study of fish assemblage structure among oxbow lakes of the Brazos River (Texas) species richness, diversity, and evenness were greatest in the connected oxbow lake [
15]. Similarly, Galat
et al [
47], Miranda [
12,
45], and Petry
et al [
48] noted more species in oxbow lakes that are connected to the Mississippi, Missouri, and the Parana River (Brazil), respectively, than in lakes permanently separated from the river. They observed greater species richness in connected floodplain water bodies than in isolated water bodies. Also Dembkowski and Miranda [
13], based on the monitoring of fish assemblages in two segments of an oxbow lake, one connected to and the other isolated from the Yazoo River, stated that greater species richness, diversity, and evenness were observed in the connected segment than isolated one.
Although the environmental variables play an initial role in structuring the fish assemblage, biological interactions are then superimposed on that structure. Thus, the influence of biotic factors, such as predator–prey interactions [
18,
49], which are significant determinants of assemblage composition, cannot be ruled out. The present study has suggested associations between environmental variables and fish abundance, but both interspecific and intra-specific biotic interactions remain to be studied in this system. Although the analyzed floodplain area is characterized by nearly pristine conditions as part of the Biebrza National Park, our results could be underestimated because legal angling and the significant increase in the number of poached fish exert pressure on the local fish assemblage. Unfortunately, fish loss is difficult to estimate. It should be noted, however, that fish abundance and biomass exceed the values reported in other temperate floodplain lakes, such as the lower section of the Vistula River [
50], but any comparisons should be performed with great caution due to differences in the applied equipment and calculations methods.
To protect and improve the biotic integrity of these relict ecosystems, knowledge about the influence of historic and contemporary connectivity with adjacent rivers on fish species richness, diversity, and assemblage composition is essential [
15]. Nevertheless, long-term data about connectivity are seldom available as these lakes are often located in remote areas and lack continued monitoring. An interesting background to a trend assessment can be drawn up from previous studies preformed within the Biebrza River. A comparison of our results with the findings of the most recent study that investigated fish communities in the Biebrza River in 1997 and 1998 [
51] indicates that the composition of various fish guilds in the quantitative structure of fish communities was not affected by anthropogenic changes. An analysis of historical [
14,
30] and present data revealed that the fish assemblage in the Middle Biebrza floodplain is still characterized by high species diversity (23 species at present, 26 species in 1983) and even higher fish abundance (4% increase in connected backwaters and 23% increase in disconnected backwaters) and fish biomass (10% increase in connected backwaters and 54% increase in disconnected backwaters) than in the past. The results point to the effectiveness of the nature conservation program covering the floodplain area, which has been included in the Biebrza National Park (established in 1993), the largest nature reserve in Poland. Results of that comparison support our hypothesis that fish populations are important bioindicators of ecological integrity in the river network under natural flood-pulsing conditions. We suggest that maintenance or restoration of connection should be an integral part of the fluvial ecosystem management plans.


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}