
The Importance of Providing Multiple-Channel Sections in Dredging Activities to Improve Fish Habitat Environments


Abstract
:1. Introduction

2. Methods
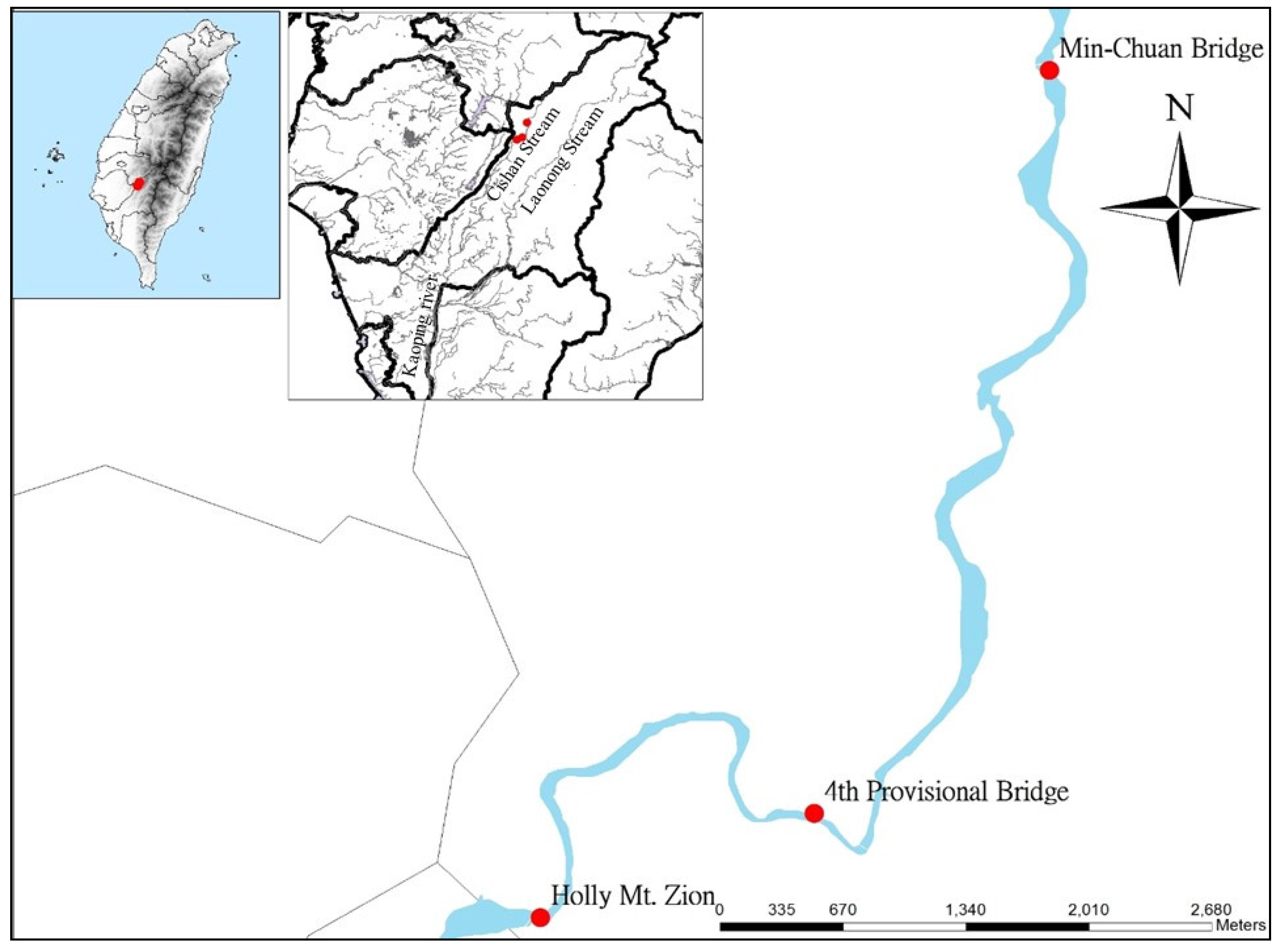
2.1. Area of Study
2.2. Fish Sampling
2.3. Habitat Survey
2.4. Data Analysis
3. Results
3.1. Fish Abundance

{kind=link}
{kind=link}
{kind=link}
| Fish Species | Multiple-Channel Section | Single-Channel Section | |
|---|---|---|---|
| Main Channel | Secondary Channel | ||
| H. formosanus | 2.91 | 3.55 | 1.32 |
| R. nantaiensis | 1.58 | 1.59 | 0.55 |
| A. paradoxus | 0.15 | 0.83 | 0.77 |
| O. alticorpus | 0.21 | 0.38 | 1.00 |
3.2. Environmental Factors
| Environmental Factor | Multiple-Channel Section | Single-Channel Section | ||||
|---|---|---|---|---|---|---|
| Main Channel | Secondary Channel | |||||
| Mean | CV (%) | Mean | CV (%) | Mean | CV (%) | |
| EC (µs/cm) | 423.98 a | 15.7 | 413.06 a | 7.2 | 421.06 a | 4.8 |
| Turbidity (NTU) | 123.96 a | 161.1 | 78.03 a | 182.3 | 71.17 a | 213.7 |
| Salinity (ppt) | 0.2 a | 11.1 | 0.2 a | 8.2 | 0.2 a | ― |
| DO (mg/L) | 8.59 a | 12.7 | 8.60 a | 9.4 | 8.35 a | 7.4 |
| pH | 8.34 a | 2.0 | 8.33 a | 1.9 | 8.38 a | 1.5 |
| Chlorophyll a (ppb) | 0.676 a | 68.8 | 0.534 a | 59.1 | 0.501 a | 69.1 |
| Flow velocity (m/s) | 0.7 b | 37.6 | 0.6 b | 54.4 | 0.5 a | 50.8 |
| Water depth (cm) | 33.4 b | 37.3 | 26.6 a | 36.6 | 39.0 c | 31.2 |
| Sand | 0.13 a | 182.1 | 0.18 a | 153.2 | 0.21 a | 131.6 |
| Gravel | 0.28 a | 98.9 | 0.33 a | 86.5 | 0.34 a | 73.9 |
| Pebbles | 0.21 b | 111.7 | 0.18 b | 103.6 | 0.08 a | 105.3 |
| Cobble | 0.17 a | 90.7 | 0.13 a | 69.6 | 0.11 a | 86.1 |
| Boulders | 0.20 a | 137.3 | 0.18 a | 117.6 | 0.26 a | 83.9 |
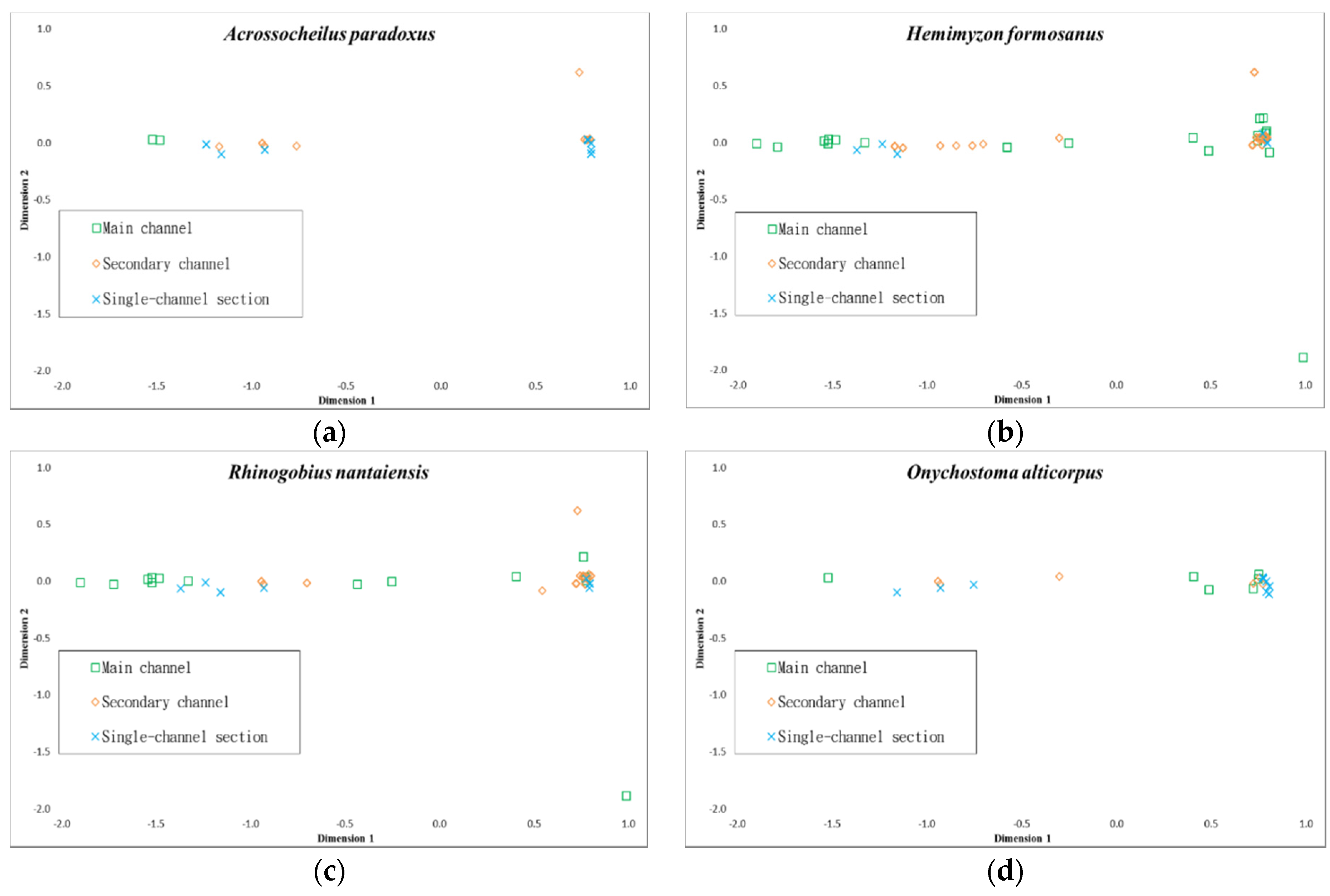
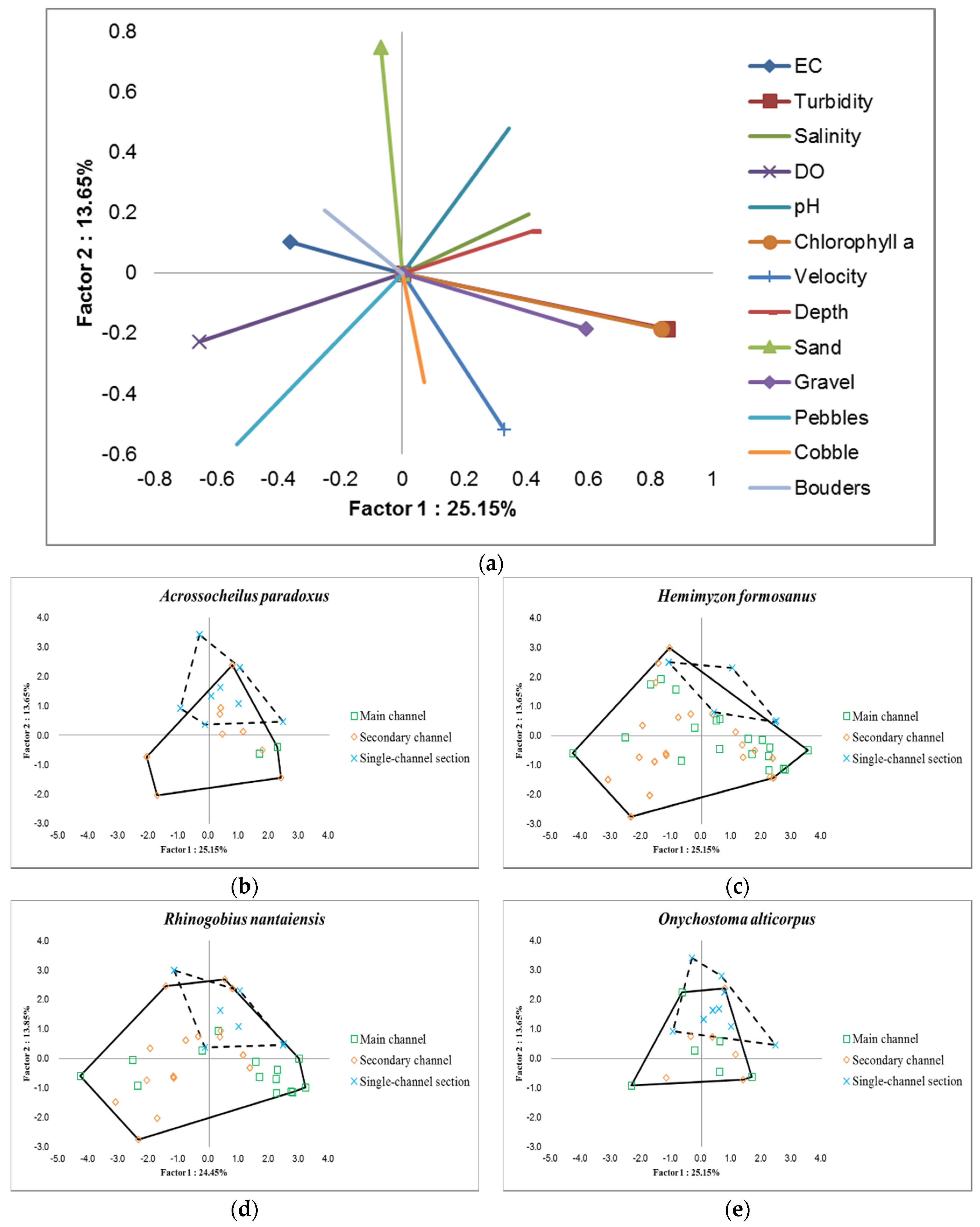
3.3. Habitat Characterization in Combination with Fish Species

3.4. Environmental Use for Fish Species

| Fish Species | Channel Type | −2 LL | Hosemer and Lemeshow Test | Equation | |
|---|---|---|---|---|---|
| Chi-Square | Sig. | ||||
| Acrossocheilus paradoxus | Main channel | 0.00 | 0.00 | 1.00 | −46.769 + 78.016 Chlorophyll a − 137.743 Flow velocity |
| Secondary channel | 28.90 | 5.07 | 0.54 | 46.796 − 233.323 Salinity − 5.258 Pebbles | |
| Single-channel section | ND | ND | ND | ND | |
| Hemimyzon formosanus | Main channel | 27.81 | 8.90 | 0.35 | 4.826 − 6.299 Boulders − 0.084 Water depth |
| Secondary channel | 17.79 | 4.27 | 0.64 | −0.916 + 20.067 Pebbles | |
| Single-channel section | ND | ND | ND | ND | |
| Rhinogobius nantaiensis | Main channel | 25.32 | 7.01 | 0.54 | 42.776 +3.439 Chlorophyll a − 229.978 Salinity |
| Secondary channel | 25.64 | 5.03 | 0.66 | 3.974 − 4.964 Flow velocity | |
| Single-channel section | 15.70 | 3.87 | 0.70 | −2.529 + 3.648 Chlorophyll a | |
| Onychostoma alticorpus | Main channel | 26.55 | 2.63 | 0.62 | −2.089 +3.154 Boulders |
| Secondary channel | ND | ND | ND | ND | |
| Single-channel section | ND | ND | ND | ND | |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Amoros, C. The concept of habitat diversity between and within ecosystems applied to river side-arm restoration. Environ. Manag. 2001, 28, 805–817. [Google Scholar] [CrossRef]
- Jähnig, S.C.; Lorenz, A.; Hering, D. Hydromorphological parameters indicating differences between single-and multiple-channel mountain rivers in Germany, in relation to their modification and recovery. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 1200–1216. [Google Scholar] [CrossRef]
- Kanno, Y.; Schmidt, C.U.; Cook, S.B.; Mattingly, H.T. Variation in microhabitat use of the threatened spotfin chub (Erimonax monachus) among stream sites and seasons. Ecol. Freshw. Fish 2012, 21, 363–374. [Google Scholar] [CrossRef]
- Wyżga, B.; Oglęcki, P.; Radecki-Pawlik, A.; Skalski, T.; Zawiejska, J. Hydromorphological complexity as a driver of the diversity of benthic invertebrate communities in the Czarny Dunajec River, Polish Carpathians. Hydrobiologia 2012, 696, 29–46. [Google Scholar] [CrossRef]
- Burge, L.M. Testing links between river patterns and in-channel characteristics using MRPP and ANOVA. Geomorphology 2004, 63, 115–130. [Google Scholar] [CrossRef]
- Seegrist, D.W.; Gard, R. Effects of floods on trout in Sagehen Creek, California. Trans. Am. Fish. Soc. 1972, 101, 478–482. [Google Scholar] [CrossRef]
- Matthews, W.J. Fish faunal structure in an Ozark stream: Stability, persistence and a catastrophic flood. Copeia 1986, 1986, 388–397. [Google Scholar] [CrossRef]
- Harvey, B.C. Susceptibility of young-of-the-year fishes to downstream displacement by flooding. Trans. Am. Fish. Soc. 1987, 116, 851–855. [Google Scholar] [CrossRef]
- Lamberti, G.A.; Gregory, S.V.; Ashkenas, L.R.; Wildman, R.C.; Moore, K.M. Stream ecosystem recovery following a catastrophic debris flow. Can. J. Fish. Aquat. Sci. 1991, 48, 196–208. [Google Scholar] [CrossRef]
- Valdez, R.A.; Hoffnagle, T.L.; McIvor, C.C.; McKinney, T.; Leibfried, W.C. Effects of a test flood on fishes of the Colorado River in Grand Canyon, Arizona. Ecol. Appl. 2001, 11, 686–700. [Google Scholar] [CrossRef]
- Wesner, J.S. Shoaling species drive fish assemblage response to sequential large floods in a small midwestern USA stream. Environ. Biol. Fishes 2011, 91, 231–242. [Google Scholar] [CrossRef]
- van der Nat, D.; Schmidt, A.P.; Tockner, K.; Edwards, P.J.; Ward, J.V. Inundation dynamics in braided floodplains: Tagliamento River, northeast Italy. Ecosystems 2002, 5, 636–647. [Google Scholar]
- Ward, J.V.; Tockner, K.; Arscott, D.B.; Claret, C. Riverine landscape diversity. Freshw. Biol. 2002, 47, 517–539. [Google Scholar] [CrossRef]
- Leopold, L.B.; Wolman, M.G. River Channel Patterns: Braided, Meandering and Straight; Geological Survey Professional Paper 282-B; United States Government Printing Office: Washington, DC, USA, 1957; pp. 39–85. [Google Scholar]
- Parker, G. On the cause and characteristic scales of meandering and braiding in rivers. J. Fluid Mech. 1976, 76, 457–480. [Google Scholar] [CrossRef]
- Nanson, G.C.; Croke, J.C. A genetic classification of floodplains. Geomorphology 1992, 4, 459–486. [Google Scholar] [CrossRef]
- Rosgen, D.L. A classification of natural rivers. Catena 1994, 22, 169–199. [Google Scholar] [CrossRef]
- Gray, D.; Scarsbrook, M.R.; Harding, J.S. Spatial biodiversity patterns in a large New Zealand braided river. N. Z. J. Mar. Freshw. Res. 2006, 40, 631–642. [Google Scholar] [CrossRef]
- Kiss, T.; Sipos, G. Braided-scale channel geometry changes in a sand-bedded river: Significance of low stages. Geomorphology 2007, 84, 209–221. [Google Scholar] [CrossRef]
- Soulsby, C.; Grant, J.; Gibbins, C.; Malcolm, I.A. Spatial and temporal variability of Atlantic salmon (Salmo salar L.) spawning activity in braided river channels: A preliminary assessment. Aquat. Sci. 2012, 74, 571–586. [Google Scholar] [CrossRef]
- Gray, D.; Harding, J.S. Braided river benthic diversity at multiple spatial scales: A hierarchical analysis of β diversity in complex floodplain systems. J. N. Am. Benthol. Soc. 2009, 28, 537–551. [Google Scholar] [CrossRef]
- Carson, M.A.; Griffiths, G.A. Bedload Transport in Gravel Channels; New Zealand Hydrological Society: Wellington, New Zealand, 1987. [Google Scholar]
- Gurnell, A.; Surian, N.; Zanoni, L. Multi-thread river channels: A perspective on changing European alpine river systems. Aquat. Sci. 2009, 71, 253–265. [Google Scholar] [CrossRef]
- Helfield, J.M.; Engström, J.; Michel, J.T.; Nilsson, C.; Jansson, R. Effects of river restoration on riparian biodiversity in secondary channels of the Pite River, Sweden. Environ. Manag. 2012, 49, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Brouder, M.J. Effects of flooding on recruitment of roundtail chub, Gila robusta, in a southwestern river. Southwest. Nat. 2001, 46, 302–310. [Google Scholar] [CrossRef]
- Fritz, K.M.; Tripe, J.A.; Guy, C.S. Recovery of three fish species to flood and seasonal drying in a tallgrass prairie stream. Trans. Kans. Acad. Sci. 2002, 105, 209–218. [Google Scholar] [CrossRef]
- Natsumeda, T. Effects of a severe flood on the movements of Japanese fluvial sculpin. Environ. Biol. Fishes 2003, 68, 417–424. [Google Scholar] [CrossRef]
- Morán-López, R.; da Silva, E.; Pérez-Bote, J.L.; Corbacho Amado, C. Associations between fish assemblages and environmental factors for Mediterranean-type rivers during summer. J. Fish Biol. 2006, 69, 1552–1569. [Google Scholar] [CrossRef]
- Lüderitz, V.; Speierl, T.; Langheinrich, U.; Völkl, W.; Gersberg, R.M. Restoration of the Upper Main and Rodach rivers—The success and its measurement. Ecol. Eng. 2011, 37, 2044–2055. [Google Scholar] [CrossRef]
- Welker, T.L.; Scarnecchia, D.L. River alteration and niche overlap among three native minnows (Cyprinidae) in the Missouri River hydrosystem. J. Fish Biol. 2006, 68, 1530–1550. [Google Scholar] [CrossRef]
- Glova, G.J.; Bonnett, M.L.; Docherty, C.R. Comparison of fish populations in riffles of three braided rivers of Canterbury, New Zealand. N. Z. J. Mar. Freshw. Res. 1985, 19, 157–165. [Google Scholar] [CrossRef]
- Jähnig, S.C.; Brunzel, S.; Gacek, S.; Lorenz, A.W.; Hering, D. Effects of re-braiding measures on hydromorphology, floodplain vegetation, ground beetles and benthic invertebrates in mountain rivers. J. Appl. Ecol. 2009, 46, 406–416. [Google Scholar] [CrossRef]
- Sukhodolov, A.; Bertoldi, W.; Wolter, C.; Surian, N.; Tubino, M. Implications of channel processes for juvenile fish habitats in Alpine rivers. Aquat. Sci. 2009, 71, 338–349. [Google Scholar] [CrossRef]
- Mosley, M.P. Analysis of the effect of changing discharge on channel morphology and instream uses in a braided river, Ohau River, New Zealand. Water Resour. Res. 1982, 18, 800–812. [Google Scholar] [CrossRef]
- Gido, K.B.; Propst, D.L.; Molles, M.C., Jr. Spatial and temporal variation of fish communities in secondary channels of the San Juan River, New Mexico and Utah. Environ. Biol. Fishes 1997, 49, 417–434. [Google Scholar] [CrossRef]
- Gurnell, A.M.; Petts, G.E. Island-dominated landscapes of large floodplain rivers, a European perspective. Freshw. Biol. 2002, 47, 581–600. [Google Scholar] [CrossRef]
- Kung, C.L.; Chyi, S.J. River terraces and landscape evolution in the Nantzu hsien Drainage Basin. J. Geogr. Sci. 2004, 2004, 47–62. [Google Scholar]
- Gualdoni, C.M.; Boccolini, M.F.; Oberto, A.M.; Principe, R.E.; Raffaini, G.B.; Corigliano, M.D.C. Potential habitats versus functional habitats in a lowland braided river (Córdoba, Argentina). Ann. Limnol. Int. J. Limnol. 2009, 45, 69–78. [Google Scholar] [CrossRef]
- Bain, M.B.; Finn, J.T.; Brooke, H.E. A quantitative method for sampling riverine microhabitats by electrofishing. N. Am. J. Fish. Manag. 1985, 5, 489–493. [Google Scholar] [CrossRef]
- Wintersberger, H. Spatial resource utilization and species assemblages of larval and juvenile fishes. Large Rivers 1996, 11, 29–44. [Google Scholar]
- Schiemer, F.; Keckeis, H.; Reckendorfer, W.; Winkler, G. The “inshore retention concept” and its significance for large rivers. Arch. Hydrobiol. Suppl. 2001, 135, 509–516. [Google Scholar]
- Platts, W.S.; Megahan, W.F.; Minshall, G.W. Methods for Evaluating Stream, Riparian, and Biotic Conditions; General Technical Report INT-138; U.S. Forest Service, Intermountain Forest and Range Experiment Station: Ogden, UT, USA, 1983. [Google Scholar]
- Hair, J.F.; Black, W.C.; Babin, B.J.; Anderson, R.E. Multivariate Data Analysis; Pearson Education, Inc.: Upper Saddle River, NJ, USA, 2009. [Google Scholar]
- Huitema, B. The Analysis of Covariance and Alternatives: Statistical Methods for Experiments, Quasi-Experiments, and Single-Case Studie; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2011; pp. 49–50. [Google Scholar]
- Pegg, M.A.; McClelland, M.A. Spatial and temporal patterns in fish communities along the Illinois River. Ecol. Freshw. Fish 2004, 13, 125–135. [Google Scholar] [CrossRef]
- King, G.; Zeng, L. Logistic regression in rare events data. Polit. Anal. 2001, 9, 137–163. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Hosmer, T.; Le Cessie, S.; Lemeshow, S. A comparison of goodness-of-fit tests for the logistic regression model. Stat. Med. 1997, 16, 965–980. [Google Scholar] [CrossRef]
- Jähnig, S.C.; Lorenz, A.W.; Hering, D. Restoration effort, habitat mosaics, and macroinvertebrates—Does channel form determine community composition? Aquat. Conserv. Mar. Freshw. Ecosyst. 2009, 19, 157–169. [Google Scholar] [CrossRef]
- Haase, P.; Hering, D.; Jähnig, S.C.; Lorenz, A.W.; Sundermann, A. The impact of hydromorphological restoration on river ecological status: A comparison of fish, benthic invertebrates, and macrophytes. Hydrobiologia 2013, 704, 475–488. [Google Scholar] [CrossRef]
- Jähnig, S.C.; Lorenz, A.W.; Lorenz, R.R.; Kail, J. A comparison of habitat diversity and interannual habitat dynamics in actively and passively restored mountain rivers of Germany. Hydrobiologia 2013, 712, 89–104. [Google Scholar] [CrossRef]
- Lee, P.Y.; Suen, J.P. Niche partitioning of fish assemblages in a mountain stream with frequent natural disturbances—An examination of microhabitat in riffle areas. Ecol. Freshw. Fish 2012, 21, 255–265. [Google Scholar] [CrossRef]
- Wu, M.Y.; Hsu, P.Y.; Chang, S.T. Feeding ecology of the endemic goby (Rhinogobius candidianus) in Chinshui Creek, Taiwan. Taiwan J. Biodivers. 2010, 12, 367–380. [Google Scholar]
- Tew, K.S.; Han, C.C.; Chou, W.R.; Fang, L.S. Habitat and fish fauna structure in a subtropical mountain stream in Taiwan before and after a catastrophic typhoon. Environ. Biol. Fish. 2002, 65, 457–462. [Google Scholar]
- Lyu, Y.S.; Suen, J.P. The relationship between the habitat preference of fish in different life stages and physical habitat parameters. J. Taiwan Agric. Eng. 2010, 56, 40–51. [Google Scholar]
- Han, C.C.; Tew, K.S.; Chen, I.S.; Su, L.Y.; Fang, L.S. Environmental biology of an endemic cyprinid, Varicorhinus alticorpus, in a subtropical mountain stream of Taiwan. Environ. Biol. Fish. 2000, 59, 153–161. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, H.-P.; Suen, J.-P. The Importance of Providing Multiple-Channel Sections in Dredging Activities to Improve Fish Habitat Environments. Water 2016, 8, 36. https://doi.org/10.3390/w8020036
Chiu H-P, Suen J-P. The Importance of Providing Multiple-Channel Sections in Dredging Activities to Improve Fish Habitat Environments. Water. 2016; 8(2):36. https://doi.org/10.3390/w8020036
Chicago/Turabian StyleChiu, Hung-Pin, and Jian-Ping Suen. 2016. "The Importance of Providing Multiple-Channel Sections in Dredging Activities to Improve Fish Habitat Environments" Water 8, no. 2: 36. https://doi.org/10.3390/w8020036
APA StyleChiu, H.-P., & Suen, J.-P. (2016). The Importance of Providing Multiple-Channel Sections in Dredging Activities to Improve Fish Habitat Environments. Water, 8(2), 36. https://doi.org/10.3390/w8020036