Adenosine Triphosphate (ATP) Is a Candidate Signaling Molecule in the Mitochondria-to-Nucleus Retrograde Response Pathway

Abstract
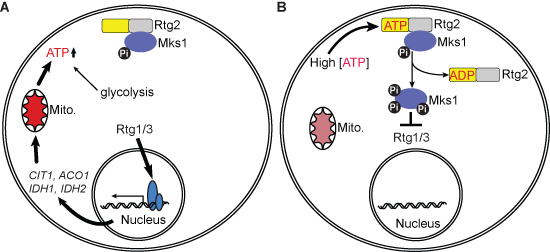
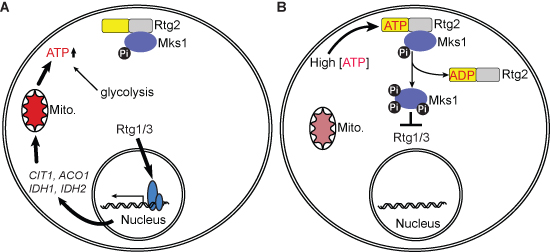
:
1. Introduction
2. Experimental Section
2.1. Strains, Plasmids and Growth Media and Growth Conditions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genotype | Source | Application |
|---|---|---|---|
| RBY915 | MATα ura3-52 leu2 lys2 RTG2-myc3 mks1::LEU2 | [28] | Figure 1, Figure 2 |
| TSY619 | MATα ura3-52 leu2 lys2 mks1::LEU2 | [38] | Figure 1 |
| PSY142 | MATα ura3-52 leu2 lys2 ura3::CIT2-lacZ | [41] | Figure 3, Figure 4 |
| PSY142-rtg2 | MATα ura3-52 leu2 lys2 ura3::CIT2-lacZ rtg2::ura3 | [41] | Figure 3, Figure 4 |
| ZLY145 | MATα ura3-52 leu2 lys2 ura3::CIT2-lacZ rtg2::ura3 mks1::kanMX4 | This study | Figure 4, Figure 5 |
| ZLY028 | MATα ura3-52 leu2 lys2 ura3::CIT2-lacZ mks1::kanMX4 | [28] | Figure 4 |
| Plasmid | Description | Source | Application |
|---|---|---|---|
| pZL1480 | pRS416-MKS1p-MKS1-HA, expressing HA-tagged Mks1 from the MKS1 promoter. | This study | Figure 1, Figure 2, Figure 4 |
| pTS215 | pRS416-MKS1, expressing MKS1 from its own promoter. | [28] | Figure 1 |
| pFZ142 | pRS416-RTG2p-RTG2(Kla)-HA, expressing HA-tagged Rtg2 homolog from K. lactis from the RTG2 promoter of S. cerevisiae. | This study | Figure 3 |
| pFZ136 | pRS416-RTG2p-RTG2(Kwa)-HA, expressing HA-tagged Rtg2 homolog from K. waltii from the RTG2 promoter of S. cerevisiae. | This study | Figure 3 |
| pZL927 | pS416-MKS1, expressing Mks1 from the endogenous promoter. | [28] | Figure 4 |
| pFZ138 | pS416-MKS1p-MKS1(Kla), expressing the Mks1 homolog from K. lactis from the MKS1 promoter of S. cerevisiae. | This study | Figure 4 |
| pFZ144 | pRS416-MKS1p-MKS1(Kwa), expressing the Mks1 homolog from K. waltii from the MKS1 promoter of S. cerevisiae. | This study | Figure 4 |
| pFZ134 | pRS416-MKS1p-MKS1(Kla)-HA3, expressing HA-tagged Mks1 homolog from K. lactis from the MKS1 promoter of S. cerevisiae. | This study | Figure 4 |
| pFL150 | pRS416-MKS1p-MKS1(Kwa)-HA3, expressing HA-tagged Mks1 homolog from K. waltii from the MKS1 promoter of S. cerevisiae. | This study | Figure 4 |
| pZL1951 | pRS415-RTG2-myc, expressing myc-tagged Rtg2 from the RTG2 promoter. | This study | Figure 5 |
| pFZ140 | pRS415-RTG2p-RTG2(Kla)-myc, expressing myc-tagged Rtg2 homolog from K. lactis from the RTG2 promoter of S. cerevisiae. | This study | Figure 5 |
| pFZ148 | pRS415-RTG2p-RTG2(Kwa)-myc, expressing myc-tagged Rtg2 homolog from K. waltii from the RTG2 promoter of S. cerevisiae. | This study | Figure 5 |
2.2. Cellular Extracts Preparation, Immunoprecipitation and Immunoblotting
2.3. Yeast Transformation and β-Galactosidase Activity Assays
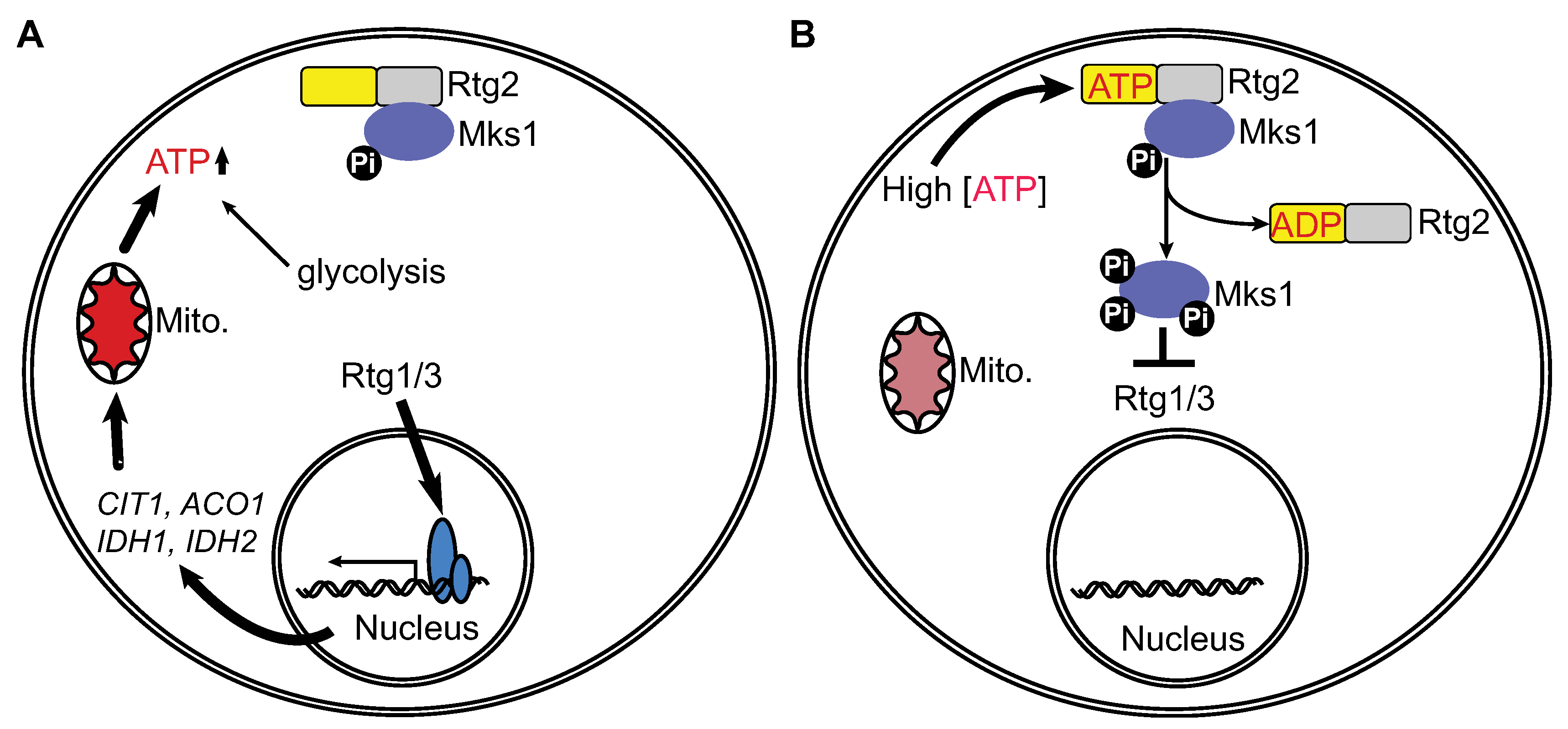
3. Results and Discussion
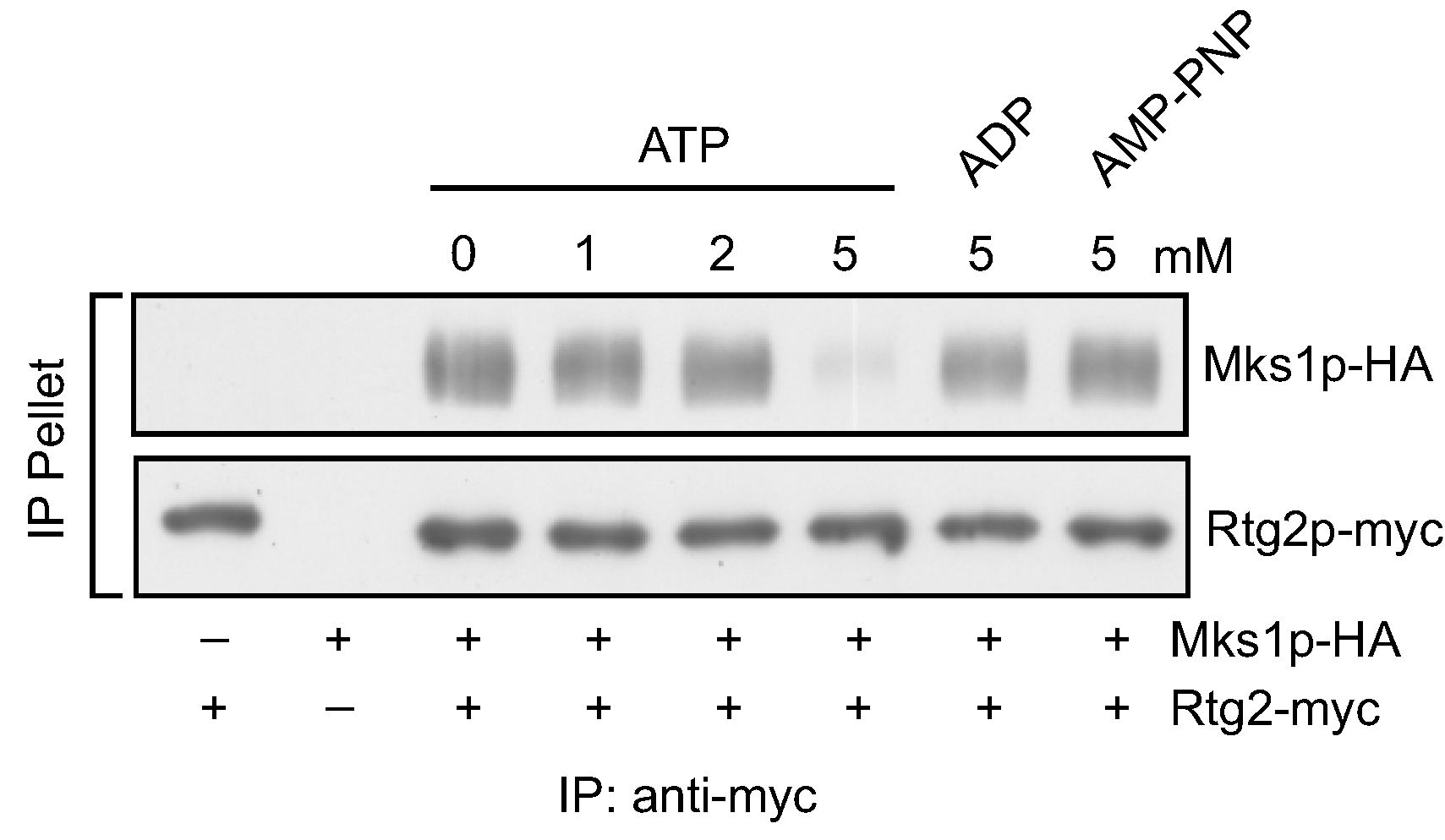
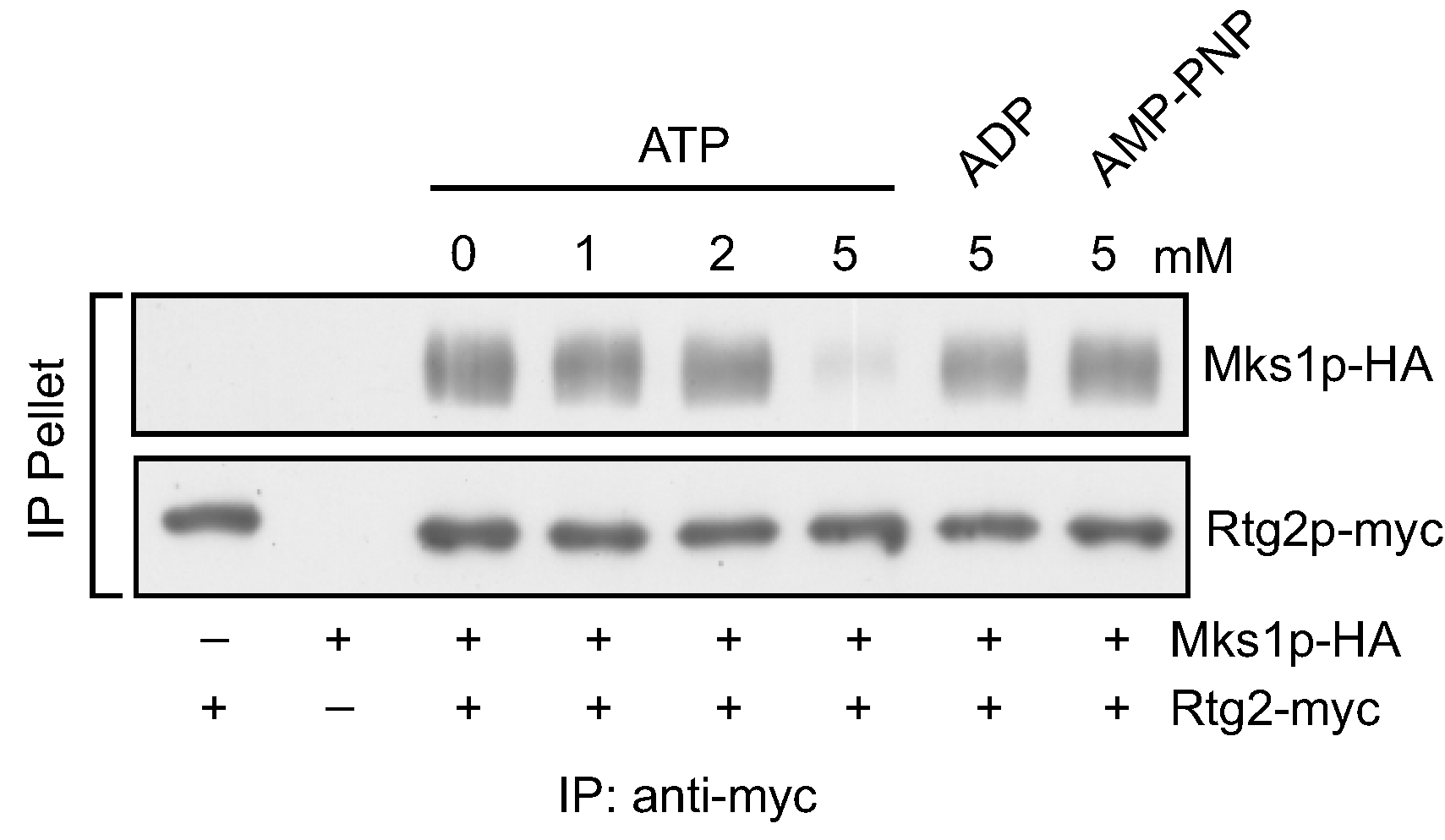
3.1. ATP, but not ADP or AMP-PNP, at High Concentrations Disrupts the Interaction between Rtg2 and Mks1 in Total Cellular Lysates
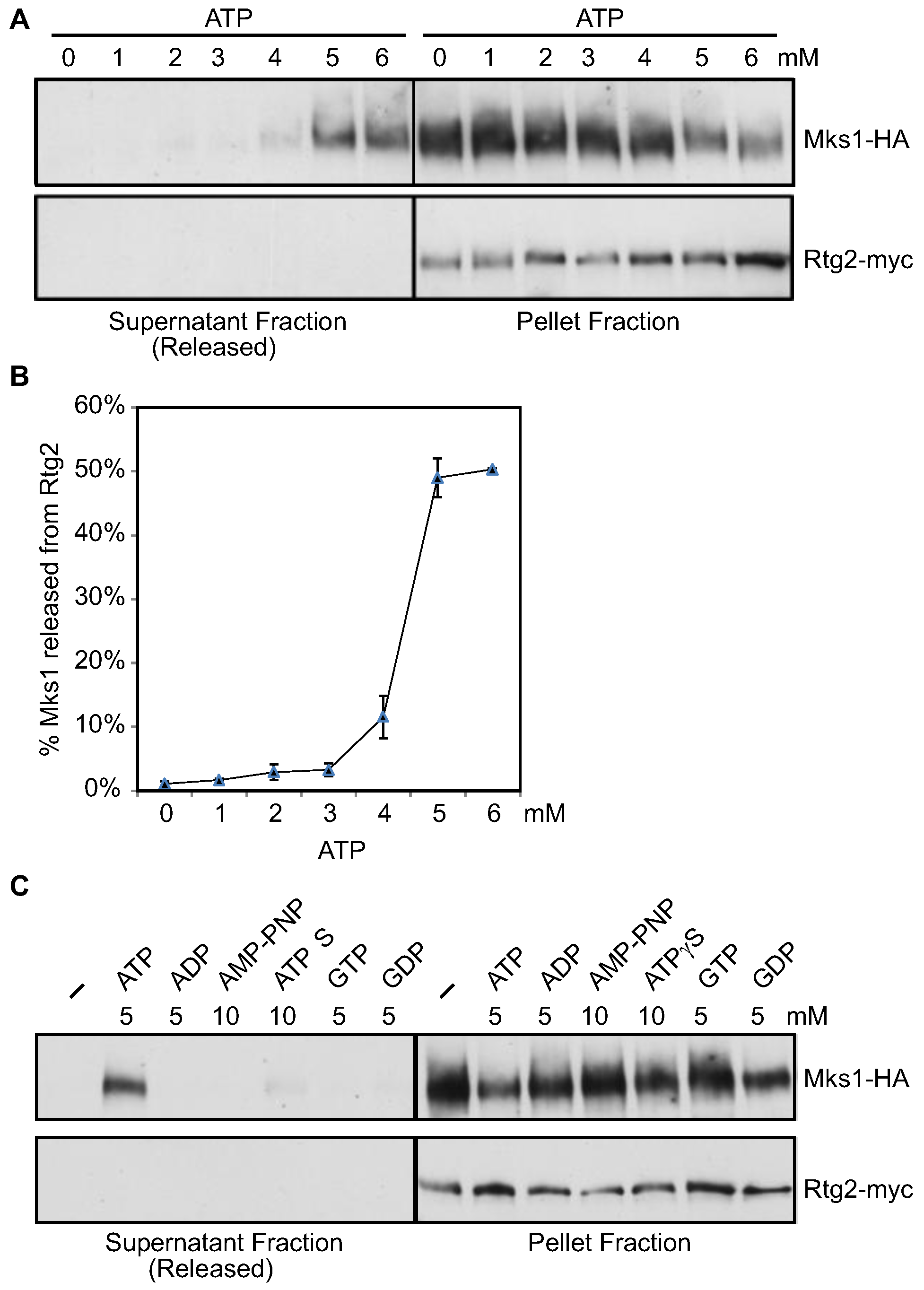
3.2. ATP within a Small Range of Physiological Concentrations Releases Mks1 from an Immunopurified Rtg2-Mks1 Complex

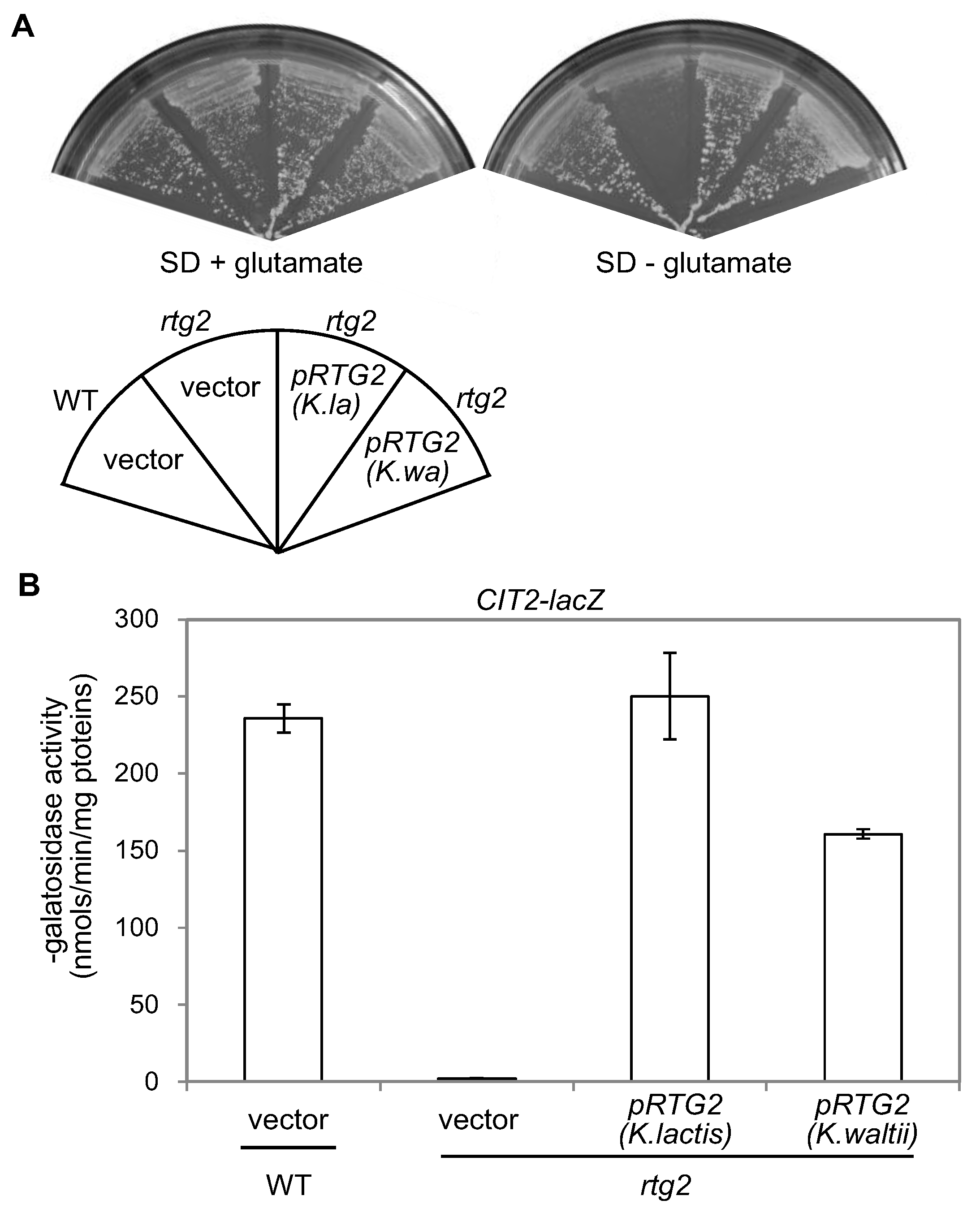
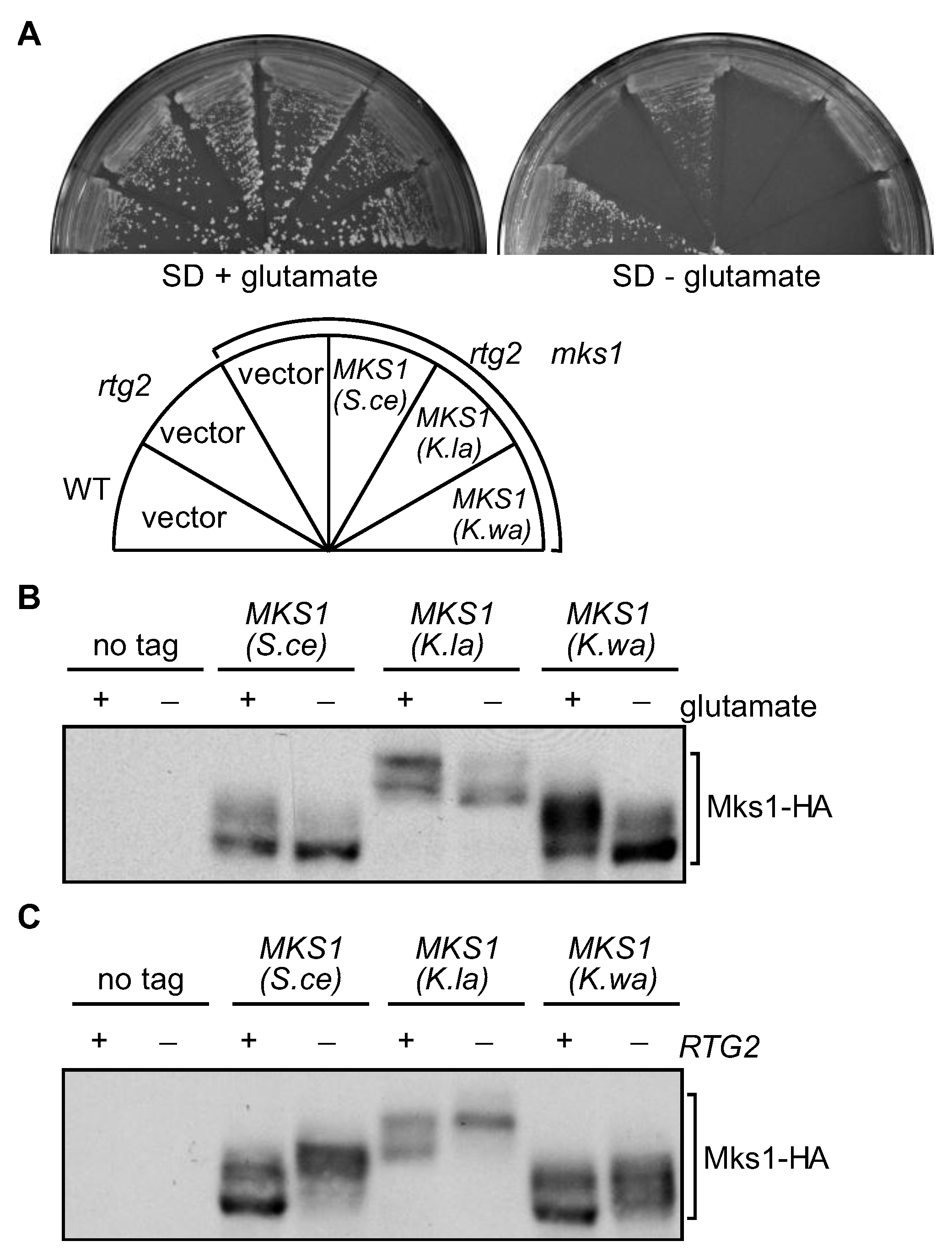
3.3. RTG2 Homologs from K. lactis and K. waltii Complement an rtg2Δ Mutation in S. cerevisiae

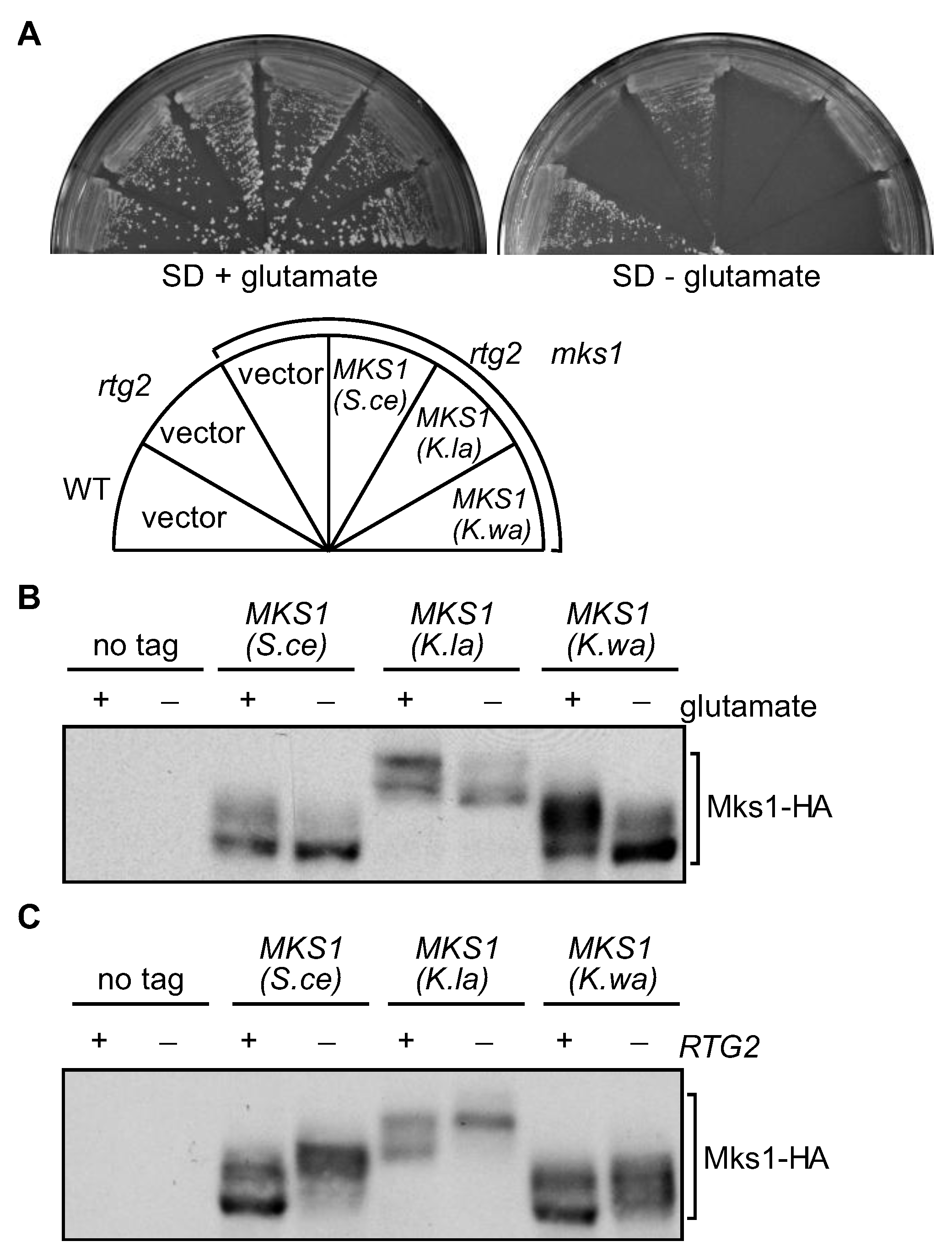
3.4. MKS1 Homologs from K. lactis and K. waltii Complement an mks1Δ Mutation in S. cerevisiae
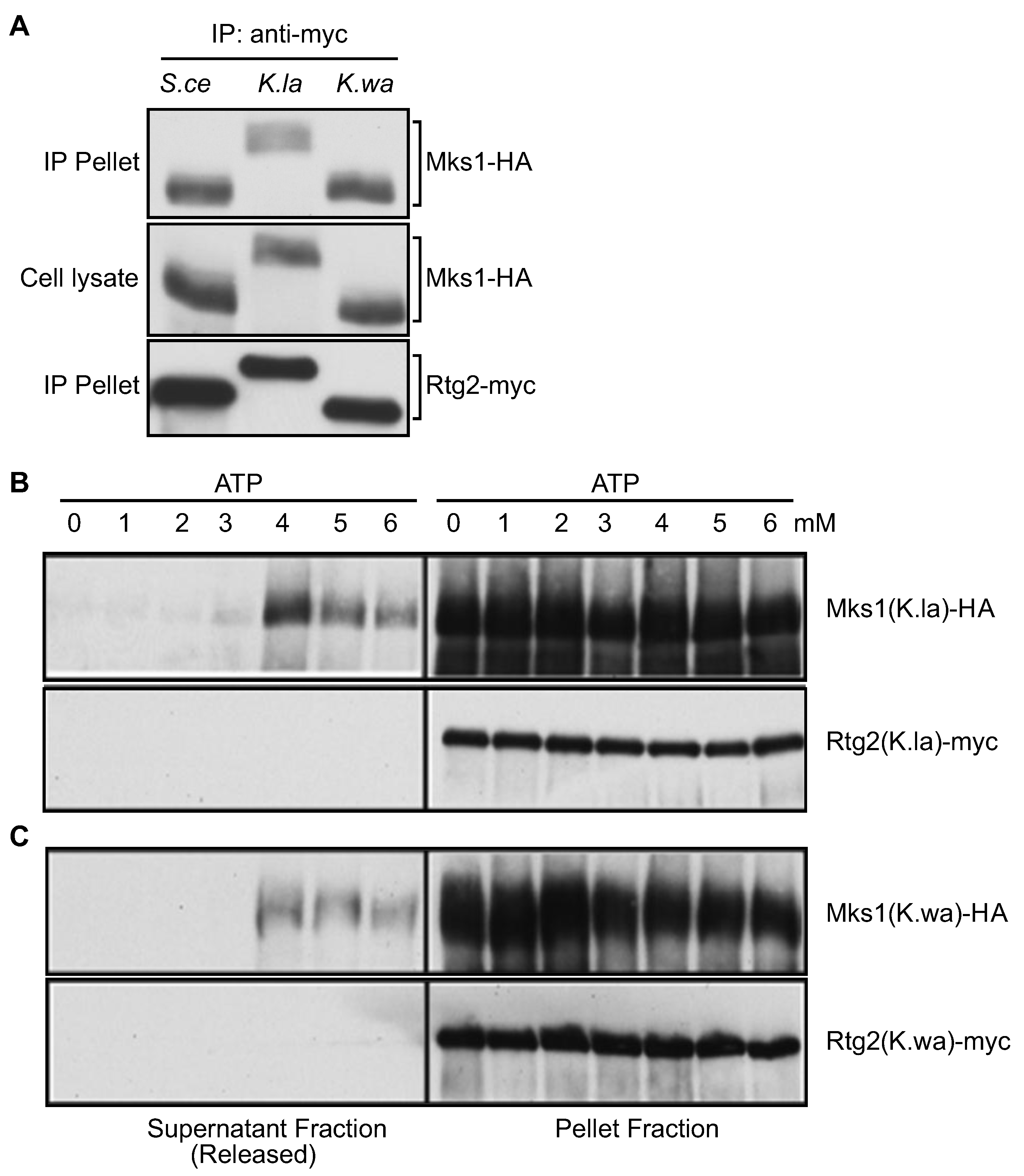
3.5. ATP at Physiological Concentrations Releases Mks1 Homologs from its Cognate Rtg2 Homologs from K. lactis and K. waltii
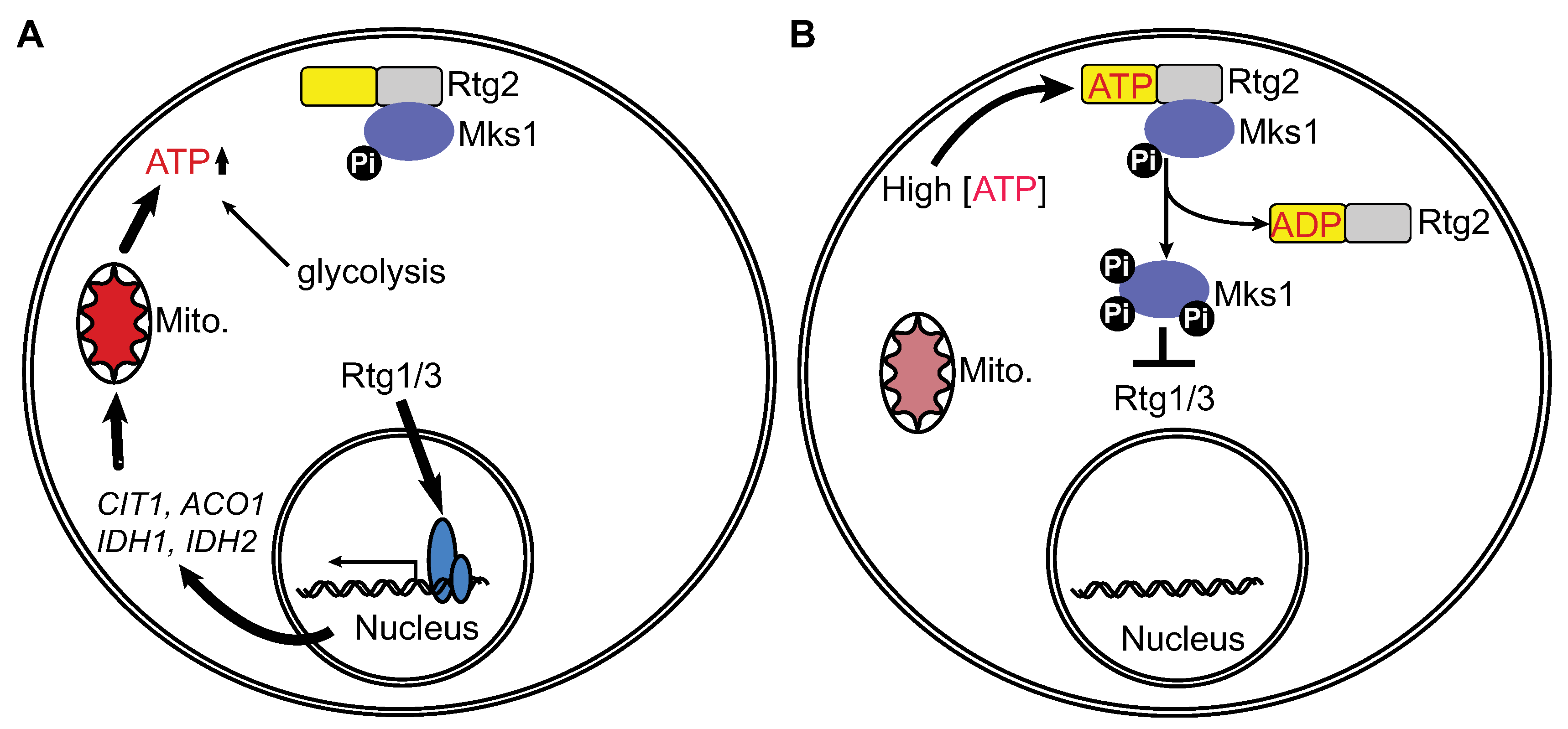
4. Conclusions
Conflict of Interest
Acknowledgments
References
- Jiang, X.; Wang, X. Cytochrome C-mediated apoptosis. Annu. Rev. Biochem. 2004, 73, 87–106. [Google Scholar] [CrossRef]
- Orrenius, S.; Gogvadze, V.; Zhivotovsky, B. Mitochondrial oxidative stress: Implications for cell death. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 143–183. [Google Scholar] [CrossRef]
- Seo, A.Y.; Joseph, A.M.; Dutta, D.; Hwang, J.C.; Aris, J.P.; Leeuwenburgh, C. New insights into the role of mitochondria in aging: Mitochondrial dynamics and more. J. Cell. Sci. 2010, 123, 2533–2542. [Google Scholar] [CrossRef]
- Wallace, D.C. A mitochondrial paradigm of metabolic and degenerative diseases, aging, and cancer: A dawn for evolutionary medicine. Annu. Rev. Genet. 2005, 39, 359–407. [Google Scholar] [CrossRef]
- Butow, R.A.; Avadhani, N.G. Mitochondrial signaling: the retrograde response. Mol. Cell. 2004, 14, 1–15. [Google Scholar] [CrossRef]
- Liu, Z.; Butow, R.A. Mitochondrial retrograde signaling. Annu. Rev. Genet. 2006, 40, 159–185. [Google Scholar] [CrossRef]
- Epstein, C.B.; Waddle, J.A.; Hale, W.T.; Dave, V.; Thornton, J.; Macatee, T.L.; Garner, H.R.; Butow, R.A. Genome-wide responses to mitochondrial dysfunction. Mol. Biol. Cell. 2001, 12, 297–308. [Google Scholar]
- Biswas, G.; Anandatheerthavarada, H.K.; Zaidi, M.; Avadhani, N.G. Mitochondria to nucleus stress signaling: A distinctive mechanism of NFkappaB/Rel activation through calcineurin-mediated inactivation of IkappaBbeta. J. Cell. Biol. 2003, 161, 507–519. [Google Scholar] [CrossRef]
- Biswas, G.; Guha, M.; Avadhani, N.G. Mitochondria-to-nucleus stress signaling in mammalian cells: Nature of nuclear gene targets, transcription regulation, and induced resistance to apoptosis. Gene 2005, 354, 132–139. [Google Scholar] [CrossRef]
- Guha, M.; Fang, J.K.; Monks, R.; Birnbaum, M.J.; Avadhani, N.G. Activation of Akt is essential for the propagation of mitochondrial respiratory stress signaling and activation of the transcriptional coactivator heterogeneous ribonucleoprotein A2. Mol. Biol. Cell. 2010, 21, 3578–3589. [Google Scholar] [CrossRef]
- Guha, M.; Tang, W.; Sondheimer, N.; Avadhani, N.G. Role of calcineurin, hnRNPA2 and Akt in mitochondrial respiratory stress-mediated transcription activation of nuclear gene targets. Biochim. Biophys Acta 2010, 1797, 1055–1065. [Google Scholar] [CrossRef]
- Jazwinski, S.M.; Kriete, A. The yeast retrograde response as a model of intracellular signaling of mitochondrial dysfunction. Front. Physiol. 2012, 3, 139. [Google Scholar]
- Dojcinovic, D.; Krosting, J.; Harris, A.J.; Wagner, D.J.; Rhoads, D.M. Identification of a region of the Arabidopsis AtAOX1a promoter necessary for mitochondrial retrograde regulation of expression. Plant. Mol. Biol. 2005, 58, 159–175. [Google Scholar] [CrossRef]
- Scheckhuber, C.Q.; Houthoofd, K.; Weil, A.C.; Werner, A.; De Vreese, A.; Vanfleteren, J.R.; Osiewacz, H.D. Alternative oxidase dependent respiration leads to an increased mitochondrial content in two long-lived mutants of the aging model Podospora anserina. PLoS One 2011, 6, e16620. [Google Scholar]
- Rhoads, D.M.; Subbaiah, C.C. Mitochondrial retrograde regulation in plants. Mitochondrion 2007, 7, 177–194. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, M.; Yu, J. Mitochondrial retrograde regulation tuning fork in nuclear genes expressions of higher plants. J. Genet. Genomics 2008, 35, 65–71. [Google Scholar] [CrossRef]
- Traven, A.; Wong, J.M.; Xu, D.; Sopta, M.; Ingles, C.J. Interorganellar communication. Altered nuclear gene expression profiles in a yeast mitochondrial dna mutant. J. Biol. Chem. 2001, 276, 4020–4027. [Google Scholar]
- Liu, Z.; Butow, R.A. A transcriptional switch in the expression of yeast tricarboxylic acid cycle genes in response to a reduction or loss of respiratory function. Mol. Cell. Biol. 1999, 19, 6720–6728. [Google Scholar]
- Moye-Rowley, W.S. Retrograde regulation of multidrug resistance in Saccharomyces cerevisiae. Gene 2005, 354, 15–21. [Google Scholar] [CrossRef]
- McCammon, M.T.; Epstein, C.B.; Przybyla-Zawislak, B.; McAlister-Henn, L.; Butow, R.A. Global transcription analysis of Krebs tricarboxylic acid cycle mutants reveals an alternating pattern of gene expression and effects on hypoxic and oxidative genes. Mol. Biol. Cell. 2003, 14, 958–972. [Google Scholar] [CrossRef]
- Woo, D.K.; Poyton, R.O. The absence of a mitochondrial genome in rho0 yeast cells extends lifespan independently of retrograde regulation. Exp. Gerontology 2009, 44, 390–397. [Google Scholar] [CrossRef]
- Miceli, M.V.; Jiang, J.C.; Tiwari, A.; Rodriguez-Quinones, J.F.; Jazwinski, S.M. Loss of mitochondrial membrane potential triggers the retrograde response extending yeast replicative lifespan. Front. Genet. 2011, 2, 102. [Google Scholar]
- Liao, X.; Butow, R.A. RTG1 and RTG2: two yeast genes required for a novel path of communication from mitochondria to the nucleus. Cell 1993, 72, 61–71. [Google Scholar] [CrossRef]
- Liao, X.S.; Small, W.C.; Srere, P.A.; Butow, R.A. Intramitochondrial functions regulate nonmitochondrial citrate synthase (CIT2) expression in Saccharomyces cerevisiae. Mol. Cell. Biol. 1991, 11, 38–46. [Google Scholar]
- Jia, Y.; Rothermel, B.; Thornton, J.; Butow, R.A. A basic helix-loop-helix-leucine zipper transcription complex in yeast functions in a signaling pathway from mitochondria to the nucleus. Mol. Cell. Biol. 1997, 17, 1110–1117. [Google Scholar]
- Sekito, T.; Thornton, J.; Butow, R.A. Mitochondria-to-nuclear signaling is regulated by the subcellular localization of the transcription factors Rtg1p and Rtg3p. Mol. Biol. Cell. 2000, 11, 2103–2115. [Google Scholar]
- Dilova, I.; Powers, T. Accounting for strain-specific differences during RTG target gene regulation in Saccharomyces cerevisiae. FEMS Yeast Res. 2006, 6, 112–119. [Google Scholar] [CrossRef]
- Liu, Z.; Sekito, T.; Spirek, M.; Thornton, J.; Butow, R.A. Retrograde signaling is regulated by the dynamic interaction between Rtg2p and Mks1p. Mol. Cell. 2003, 12, 401–411. [Google Scholar] [CrossRef]
- Koonin, E.V. Yeast protein controlling inter-organelle communication is related to bacterial phosphatases containing the Hsp 70-type ATP-binding domain. Trends Biochem. Sci. 1994, 19, 156–157. [Google Scholar] [CrossRef]
- Bork, P.; Sander, C.; Valencia, A. An ATPase domain common to prokaryotic cell cycle proteins, sugar kinases, actin, and hsp70 heat shock proteins. Proc. Natl. Acad. Sci USA 1992, 89, 7290–7294. [Google Scholar] [CrossRef]
- Ruiz-Roig, C.; Noriega, N.; Duch, A.; Posas, F.; de Nadal, E. The Hog1 SAPK controls the Rtg1/Rtg3 transcriptional complex activity by multiple regulatory mechanisms. Mol. Biol. Cell. 2012, 23, 4286–4296. [Google Scholar] [CrossRef]
- Dilova, I.; Aronova, S.; Chen, J.C.; Powers, T. Tor signaling and nutrient-based signals converge on Mks1p phosphorylation to regulate expression of Rtg1.Rtg3p-dependent target genes. J. Biol. Chem. 2004, 279, 46527–46535. [Google Scholar]
- Dilova, I.; Chen, C.Y.; Powers, T. Mks1 in concert with TOR signaling negatively regulates RTG target gene expression in S. cerevisiae. Curr. Biol. 2002, 12, 389–395. [Google Scholar] [CrossRef]
- Komeili, A.; Wedaman, K.P.; O'Shea, E.K.; Powers, T. Mechanism of metabolic control. Target of rapamycin signaling links nitrogen quality to the activity of the Rtg1 and Rtg3 transcription factors. J. Cell. Biol. 2000, 151, 863–878. [Google Scholar] [CrossRef]
- Sekito, T.; Liu, Z.; Thornton, J.; Butow, R.A. RTG-dependent mitochondria-to-nucleus signaling is regulated by MKS1 and is linked to formation of yeast prion [URE3]. Mol. Biol. Cell. 2002, 13, 795–804. [Google Scholar] [CrossRef]
- Tate, J.J.; Cox, K.H.; Rai, R.; Cooper, T.G. Mks1p is required for negative regulation of retrograde gene expression in Saccharomyces cerevisiae but does not affect nitrogen catabolite repression-sensitive gene expression. J. Biol. Chem. 2002, 277, 20477–20482. [Google Scholar]
- Ferreira Junior, J.R.; Spirek, M.; Liu, Z.; Butow, R.A. Interaction between Rtg2p and Mks1p in the regulation of the RTG pathway of Saccharomyces cerevisiae. Gene 2005, 354, 2–8. [Google Scholar] [CrossRef]
- Liu, Z.; Spirek, M.; Thornton, J.; Butow, R.A. A novel degron-mediated degradation of the RTG pathway regulator, Mks1p, by SCFGrr1. Mol. Biol. Cell. 2005, 16, 4893–4904. [Google Scholar] [CrossRef]
- Chen, E.J.; Kaiser, C.A. LST8 negatively regulates amino acid biosynthesis as a component of the TOR pathway. J. Cell. Biol. 2003, 161, 333–347. [Google Scholar] [CrossRef]
- Chen, E.J.; Kaiser, C.A. Amino acids regulate the intracellular trafficking of the general amino acid permease of Saccharomycescerevisiae. Proc. Natl. Acad. Sci. USA 2002, 99, 14837–14842. [Google Scholar] [CrossRef]
- Liu, Z.; Sekito, T.; Epstein, C.B.; Butow, R.A. RTG-dependent mitochondria to nucleus signaling is negatively regulated by the seven WD-repeat protein Lst8p. Embo. J. 2001, 20, 7209–7219. [Google Scholar] [CrossRef]
- Breitkreutz, A.; Choi, H.; Sharom, J.R.; Boucher, L.; Neduva, V.; Larsen, B.; Lin, Z.Y.; Breitkreutz, B.J.; Stark, C.; Liu, G.; Ahn, J.; Dewar-Darch, D.; Reguly, T.; Tang, X.; Almeida, R.; Qin, Z.S.; Pawson, T.; Gingras, A.C.; Nesvizhskii, A.I.; Tyers, M. A global protein kinase and phosphatase interaction network in yeast. Science 2010, 328, 1043–1046. [Google Scholar] [CrossRef]
- Amberg, D.C.; Burke, D.J.; Strathern, J.N. Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual; Cold Spring Harbor Laboratory: New York, NY, USA, 2005. [Google Scholar]
- Yaffe, M.P.; Schatz, G. Two nuclear mutations that block mitochondrial protein import in yeast. Proc. Natl. Acad. Sci. USA 1984, 81, 4819–4823. [Google Scholar] [CrossRef]
- Paulus, B.F.; Bryant, F.R. Time-dependent inhibition of recA protein-catalyzed ATP hydrolysis by ATPgammaS: Evidence for a rate-determining isomerization of the recA-ssDNA complex. Biochemistry 1997, 36, 7832–7838. [Google Scholar] [CrossRef]
- Yu, X.; Egelman, E.H. Direct visualization of dynamics and co-operative conformational changes within RecA filaments that appear to be associated with the hydrolysis of adenosine 5'-O-(3-thiotriphosphate). J. Mol. Biol. 1992, 225, 193–216. [Google Scholar] [CrossRef]
- Larsson, C.; Nilsson, A.; Blomberg, A.; Gustafsson, L. Glycolytic flux is conditionally correlated with ATP concentration in Saccharomyces cerevisiae: A chemostat study under carbon- or nitrogen-limiting conditions. J. Bacteriol. 1997, 179, 7243–7250. [Google Scholar]
- Sauer, U.; Schlattner, U. Inverse metabolic engineering with phosphagen kinase systems improves the cellular energy state. Metab. Eng. 2004, 6, 220–228. [Google Scholar] [CrossRef]
- Liu, Z.; Butow, R.A. Mitochondrial Retrograde Signaling. Annu. Rev. Genet. 2006, 40, 159–185. [Google Scholar] [CrossRef]
- Hardie, D.G. AMP-activated protein kinase: An energy sensor that regulates all aspects of cell function. Genes Dev. 2011, 25, 1895–1908. [Google Scholar] [CrossRef]
- Carling, D.; Thornton, C.; Woods, A.; Sanders, M.J. AMP-activated protein kinase: New regulation, new roles? Biochem. J. 2012, 445, 11–27. [Google Scholar] [CrossRef]
- Schuller, H.J. Transcriptional control of nonfermentative metabolism in the yeast Saccharomyces cerevisiae. Curr. Genet. 2003, 43, 139–160. [Google Scholar]
- Zaman, S.; Lippman, S.I.; Zhao, X.; Broach, J.R. How Saccharomyces responds to nutrients. Annu. Rev. Genet. 2008, 42, 27–81. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Zhang, F.; Pracheil, T.; Thornton, J.; Liu, Z. Adenosine Triphosphate (ATP) Is a Candidate Signaling Molecule in the Mitochondria-to-Nucleus Retrograde Response Pathway. Genes 2013, 4, 86-100. https://doi.org/10.3390/genes4010086
Zhang F, Pracheil T, Thornton J, Liu Z. Adenosine Triphosphate (ATP) Is a Candidate Signaling Molecule in the Mitochondria-to-Nucleus Retrograde Response Pathway. Genes. 2013; 4(1):86-100. https://doi.org/10.3390/genes4010086
Chicago/Turabian StyleZhang, Feng, Tammy Pracheil, Janet Thornton, and Zhengchang Liu. 2013. "Adenosine Triphosphate (ATP) Is a Candidate Signaling Molecule in the Mitochondria-to-Nucleus Retrograde Response Pathway" Genes 4, no. 1: 86-100. https://doi.org/10.3390/genes4010086