Assessment of Fecundity and Germ Line Transmission in Two Transgenic Pig Lines Produced by Sleeping Beauty Transposition


Abstract
:
1. Introduction
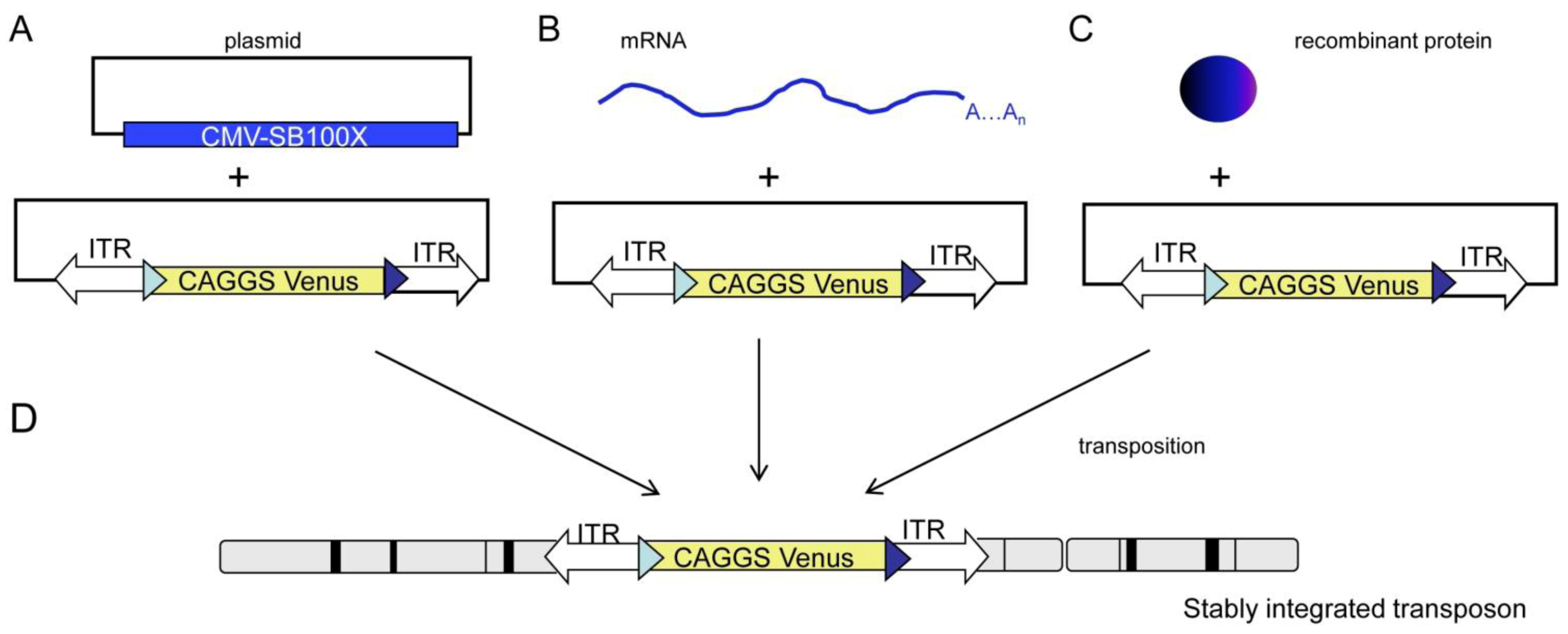
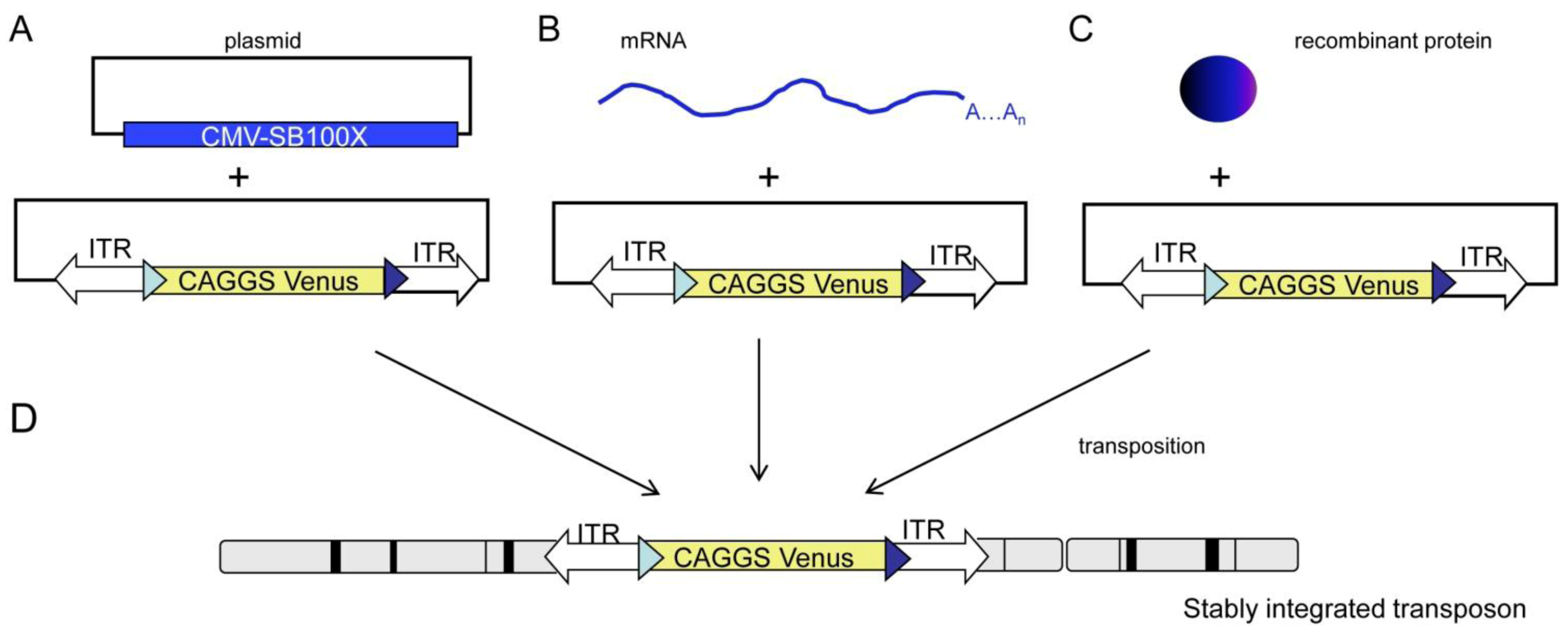
2. Results and Discussion
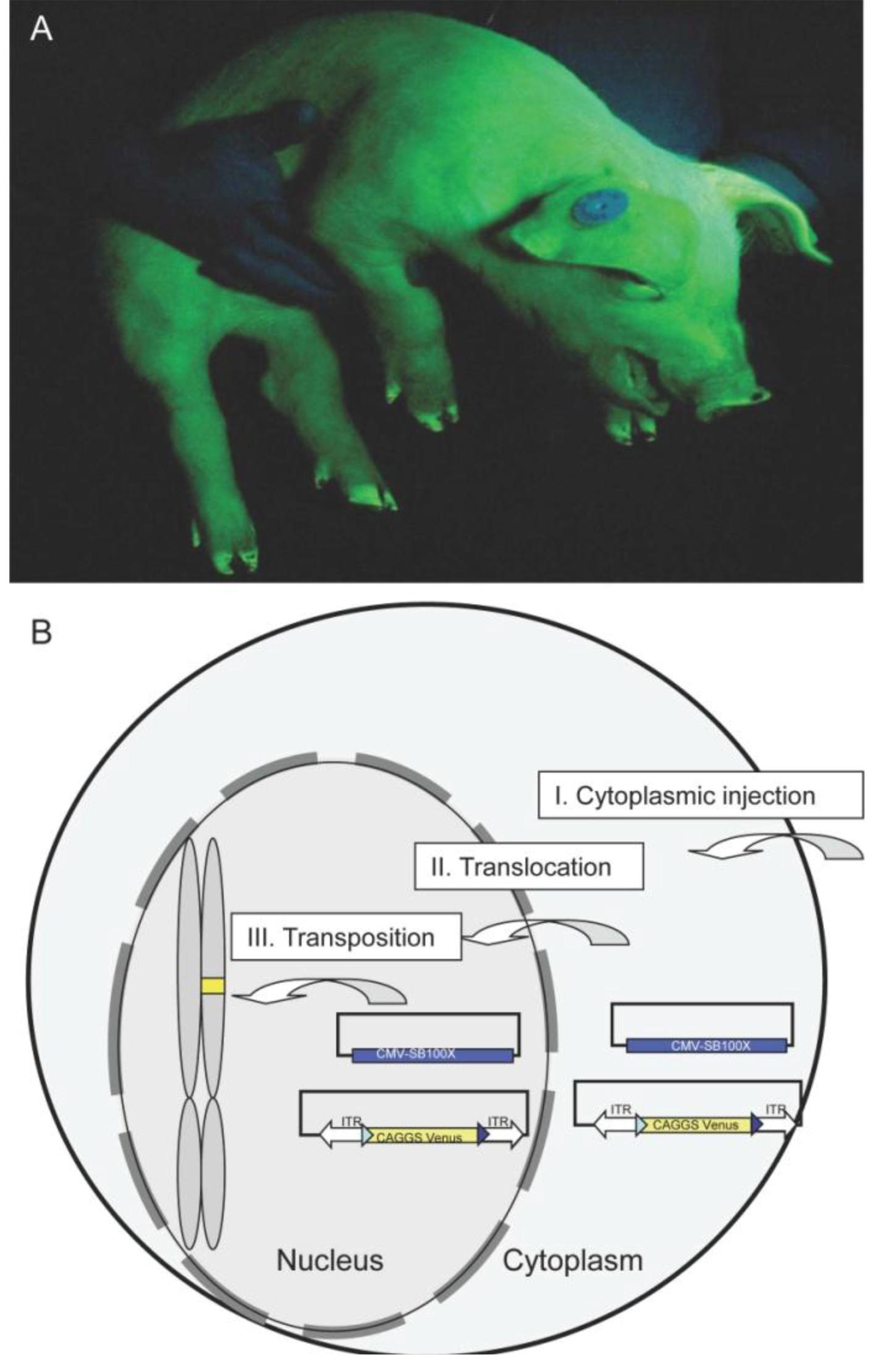
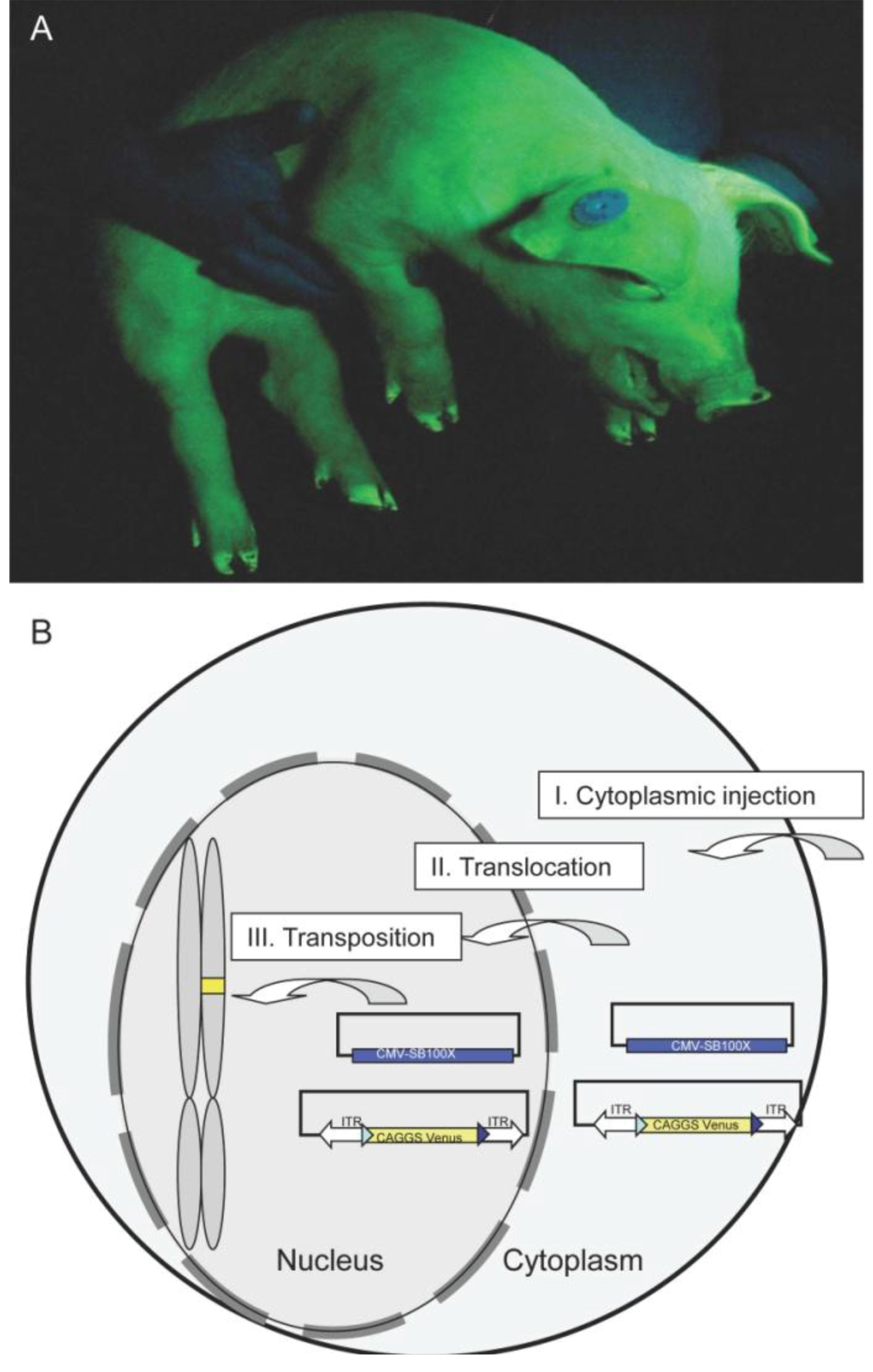
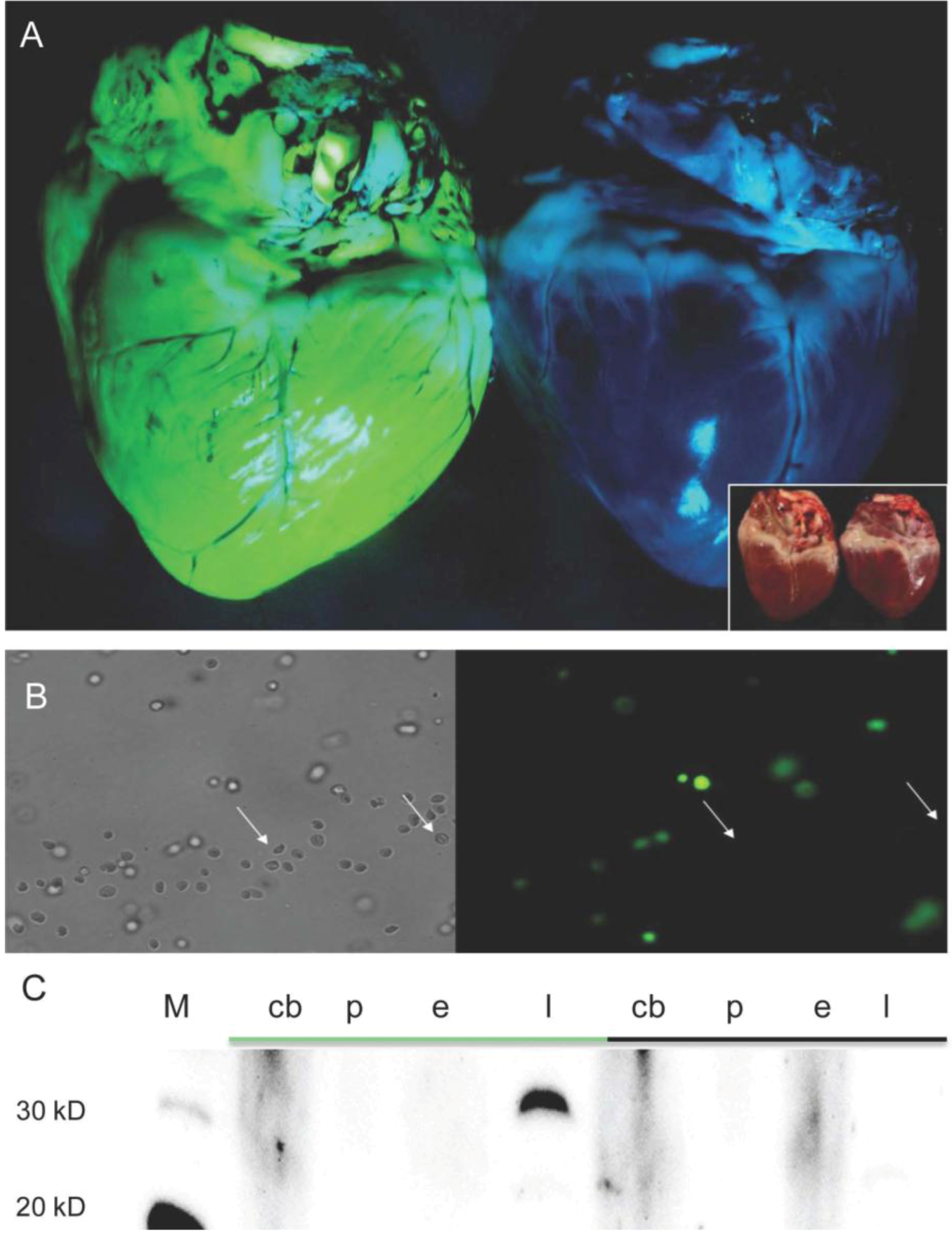
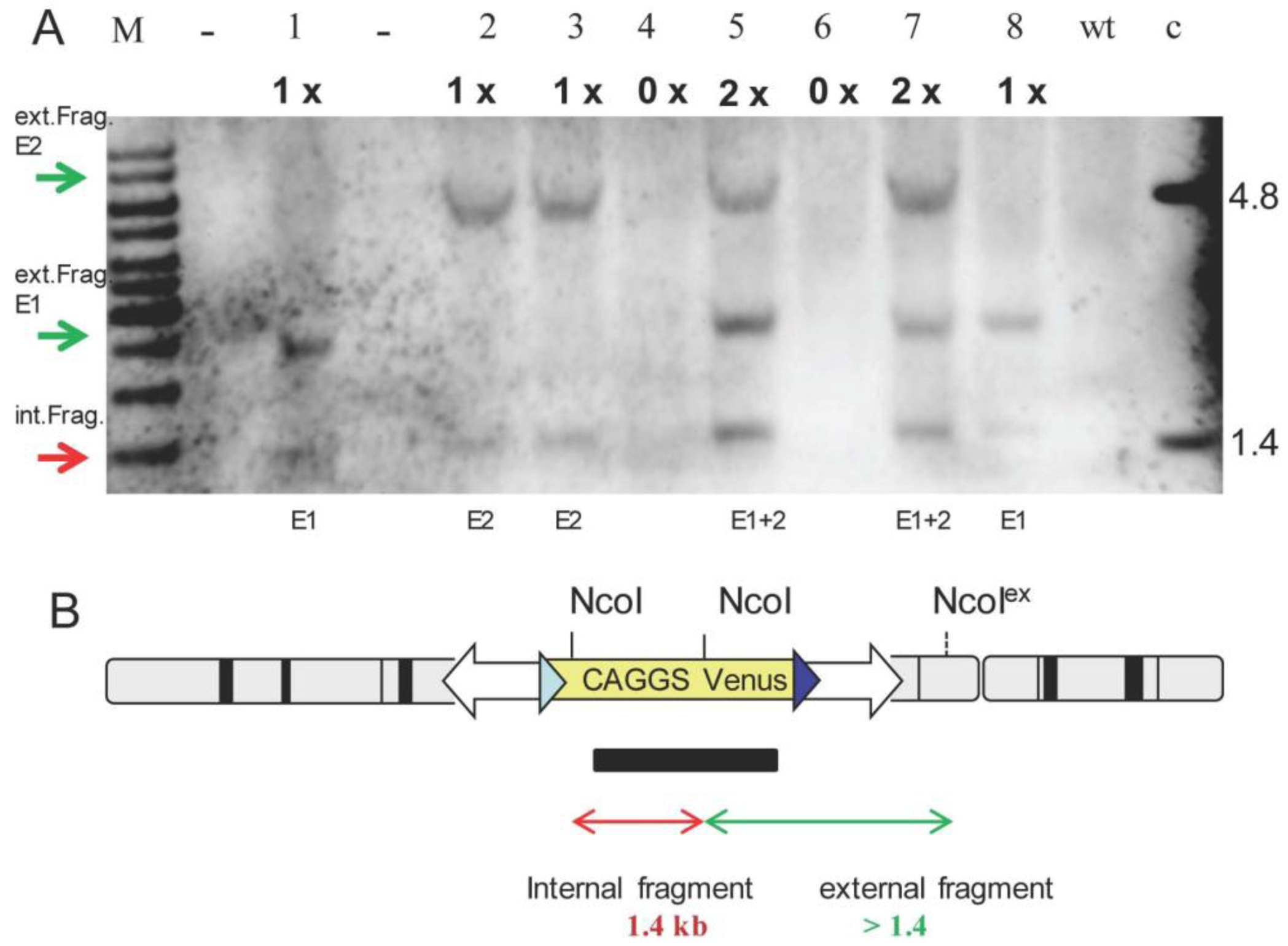
2.1. Generation and Analysis of Founder Animals
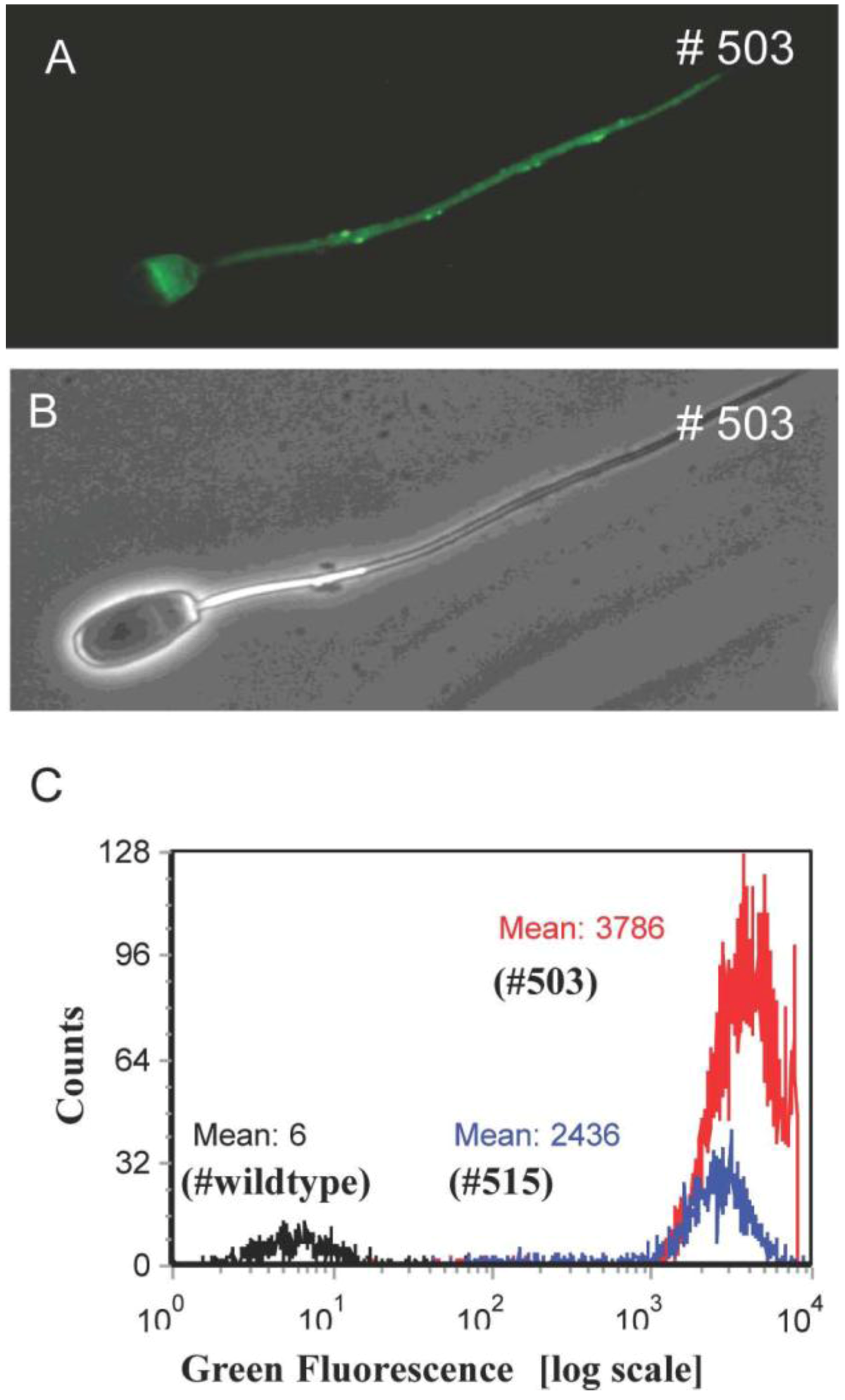
2.2. Analysis of Spermatozoa from Founder Boars
2.3. Assessment of Germ Line Transmission and Derivation of F1-lines

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
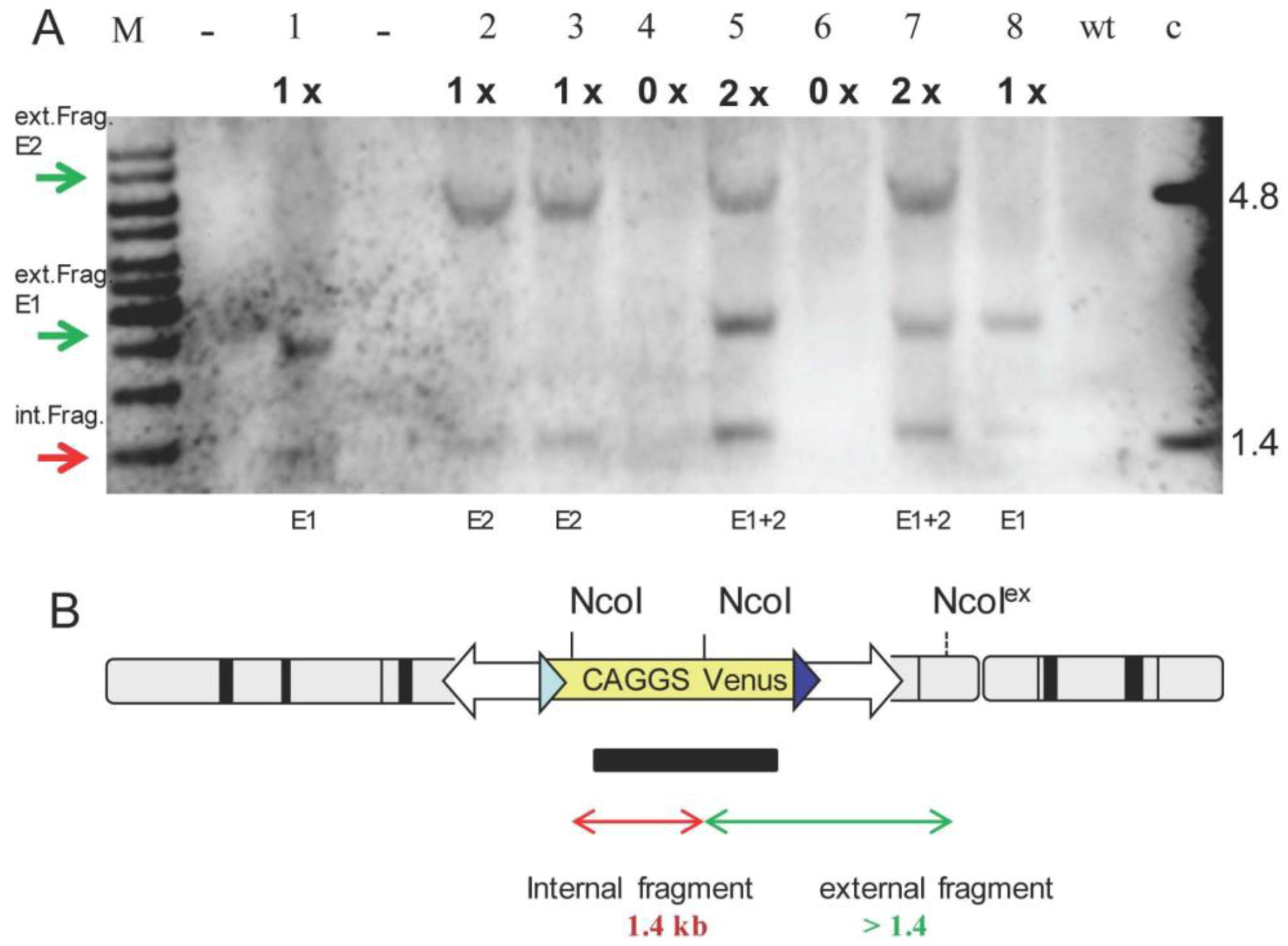
{kind=link}
| Breeding of boar / sow | Pregnancies after artificial insemination | Litter number | Born piglets | Ratio female / male | Transposon positive (%) | Ratio female / male of transposon positive piglets | Ratio female / male of non-transgenic piglets |
|---|---|---|---|---|---|---|---|
| F0 #503 / wt | 3 / 3 | 1 | 8 | 5 / 3 | 5 (63) | 3 / 2 | 2 / 1 |
| 2 | 13 | 3 / 10 | 12 ( 92) | 3 / 9 | 0 / 1 | ||
| 3 | 9 | 4 / 5 | 7 (78) | 3 / 4 | 1 / 1 | ||
| F0 #505 / wt | 2 / 3 | 4 | 6 | 1 / 5 | 5 (83) | 1 / 4 | 0 / 1 |
| 5 | 8 | 2 / 6 | 6 (75) | 2 / 4 | 0 / 2 | ||
| Subtotal | 5 | x = 8.8, n = 44 | 15 / 29 | 35 (80) | 12 / 23 | 3 / 6 | |
| F0#505 / | 1/1 | 6 | 10 | 3 / 7 | 8 (80) | 2 / 6 | 1 / 1 |
| F1#518 | |||||||
| F0#503 / | 2/2 | 7 | 3 | 2 / 1 | 2 (67) | 2 / 0 | 0 / 1 |
| F1#537 | 8 | 5 | 3 / 2 | 5 (100) | 3 / 2 | 0 / 0 | |
| F1#538 | |||||||
| Subtotal | 3 | x = 6.0, n = 18 | 8 / 10 | 15 (83) | 7 / 8 | 1 / 2 | |
| Total | 8 | x = 7.8, n = 62 | 23 / 39 | 50 (81) | 19 / 31 | 4 / 8 |


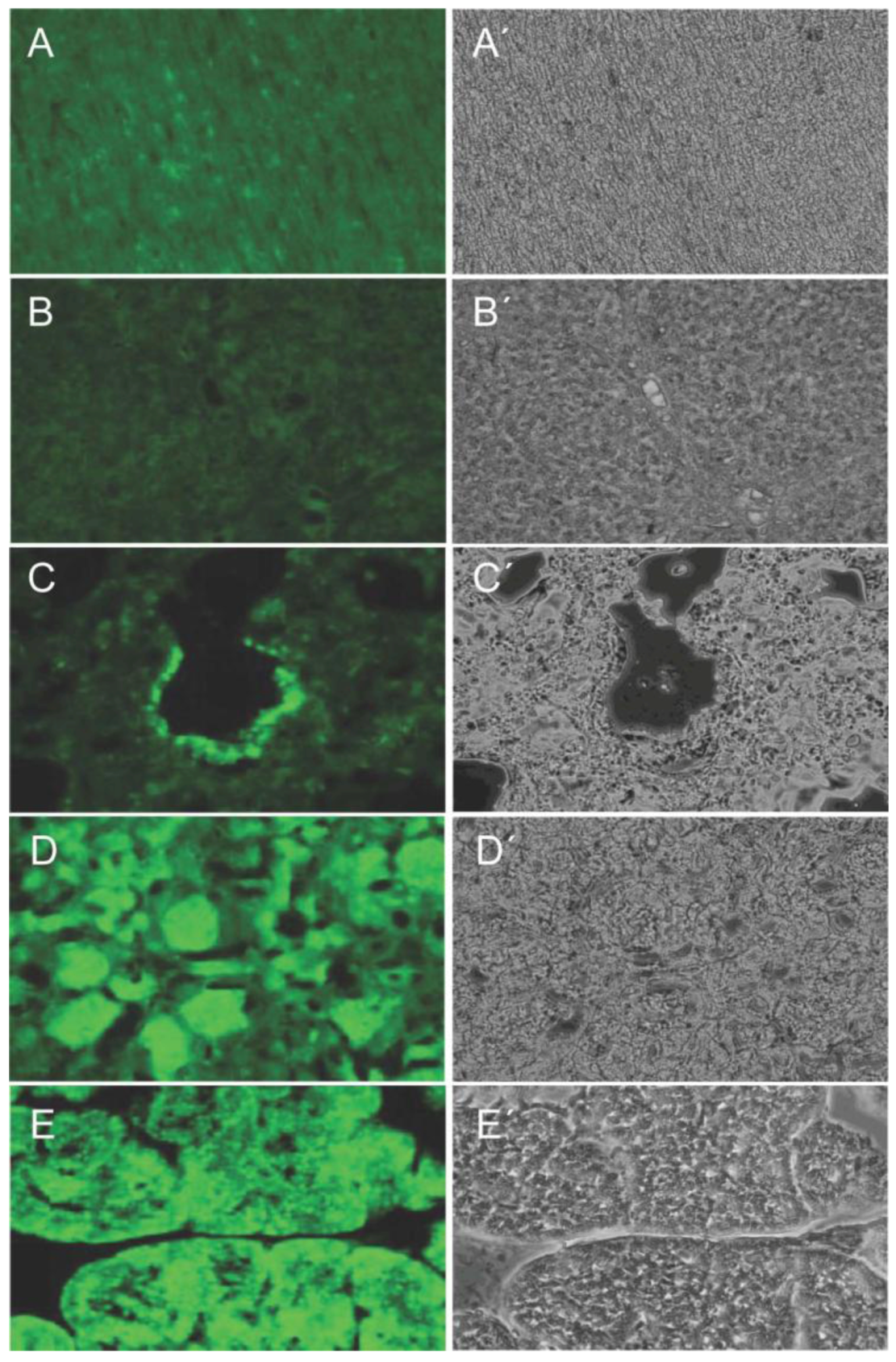
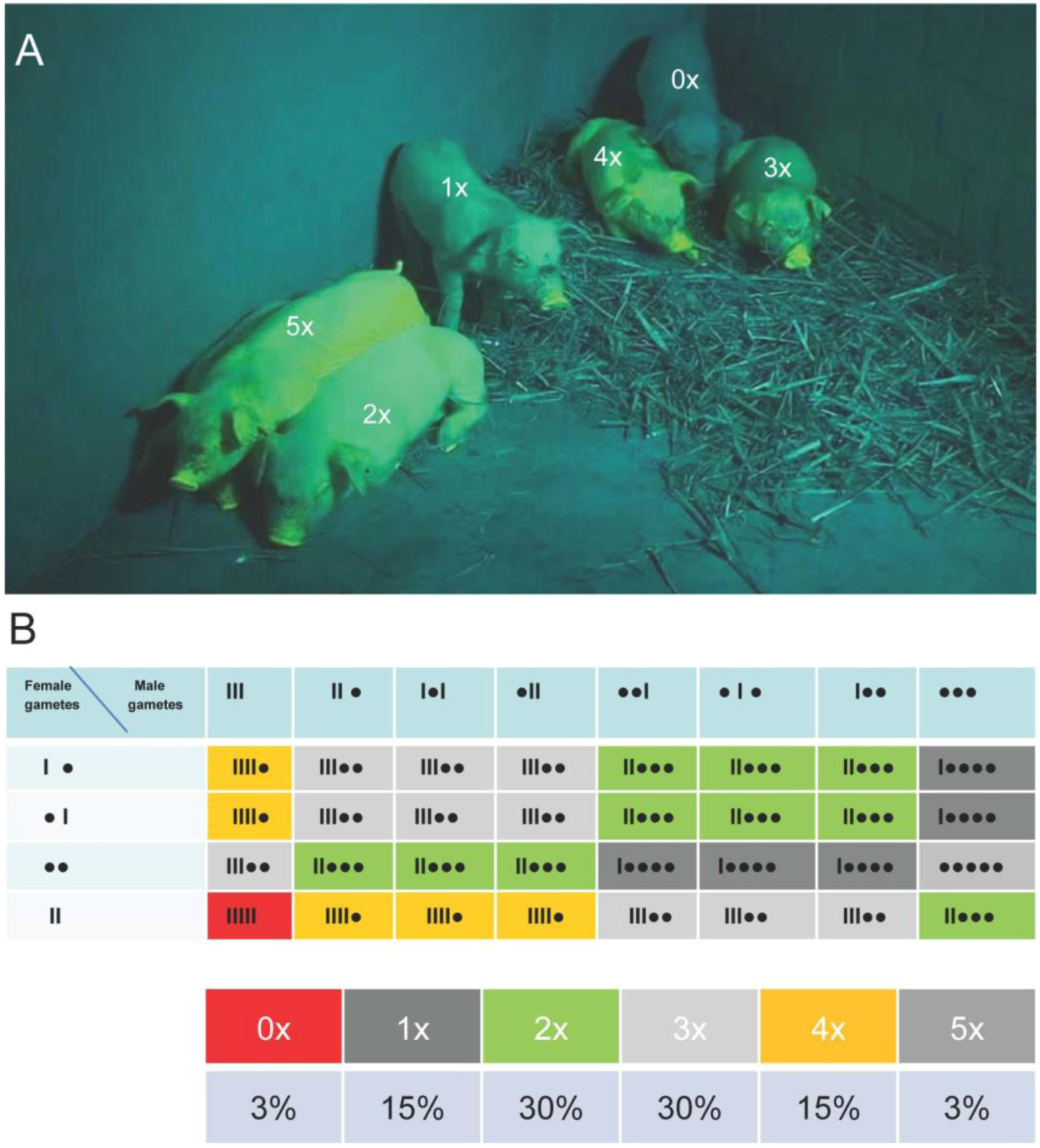
2.4. Crossbreeding to Generate Piglets with Maximized Transposon Copy Number and Venus Expression

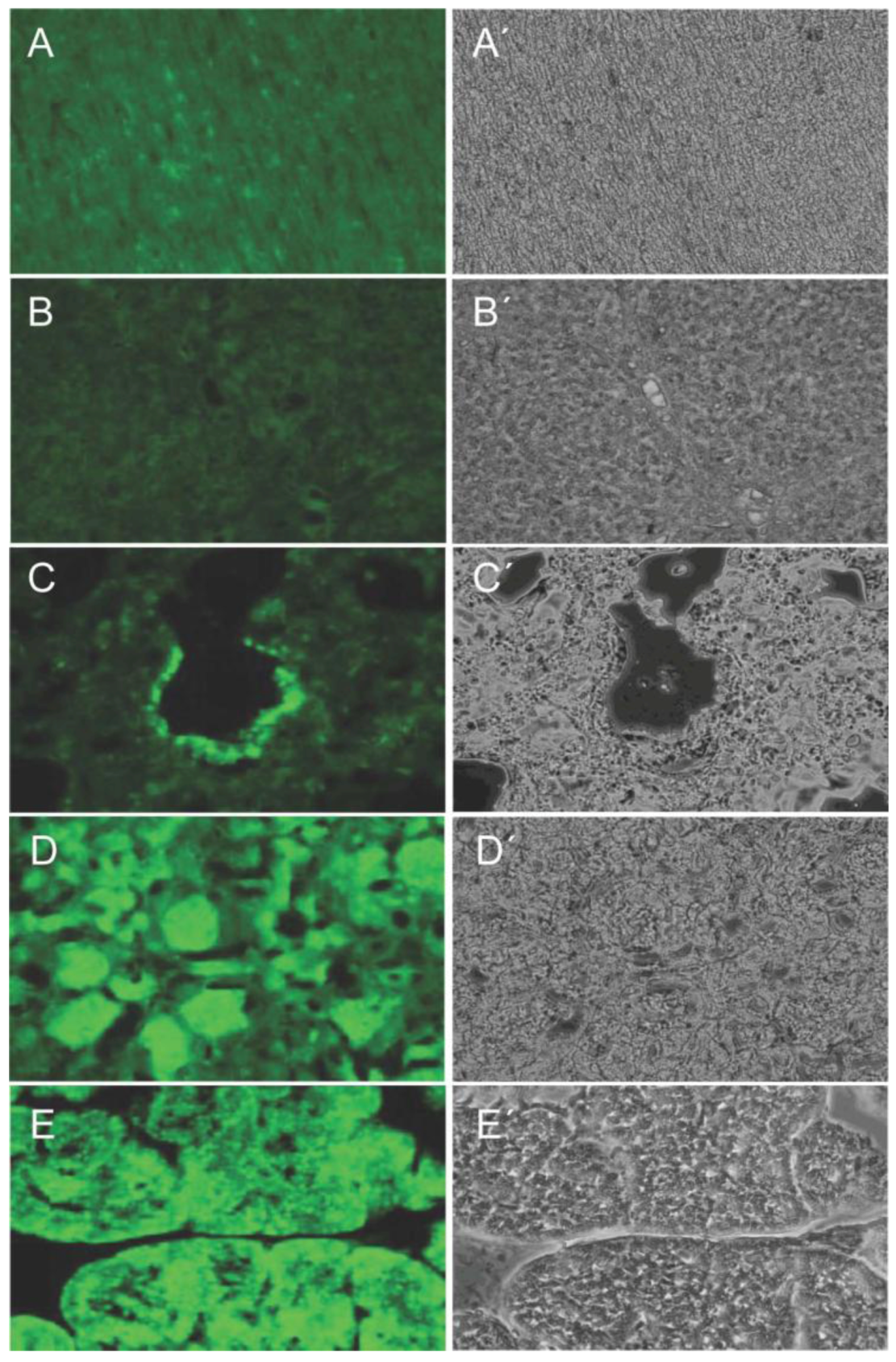
2.5. Generation of Porcine Models for Biomedical and Pharmaceutical Research
3. Experimental Section
3.1. Animals and Samples
3.2. Southern Blotting
3.3. Sperm Analysis
3.4. Western Blotting
3.5. Macroscopic Imaging and Fluorescence Microscopy
4. Conclusions
Acknowledgments
Conflict of Interests
References
- Luo, Y.; Li, J.; Liu, Y.; Lin, L.; Du, Y.; Li, S.; Yang, H.; Vajta, G.; Callesen, H.; Bolund, L.; Sorensen, C.B. High efficiency of BRCA1 knockout using rAAV-mediated gene targeting: Developing a pig model for breast cancer. Transgenic. Res. 2011, 20, 975–988. [Google Scholar] [CrossRef]
- Muller, M.; Brem, G. Transgenic strategies to increase disease resistance in livestock. Reprod. Fertil. Dev. 1994, 6, 605–613. [Google Scholar] [CrossRef]
- Tanila, H. Wading pools, fading memories-place navigation in transgenic mouse models of Alzheimer's disease. Front. Aging. Neurosci. 2012, 4, 11. [Google Scholar]
- Jacobsen, J.C.; Bawden, C.S.; Rudiger, S.R.; McLaughlan, C.J.; Reid, S.J.; Waldvogel, H.J.; MacDonald, M.E.; Gusella, J.F.; Walker, S.K.; Kelly, J.M.; Webb, G.C.; Faull, R.L.; Rees, M.I.; Snell, R.G. An ovine transgenic, Huntington's disease model. Hum. Mol. Genet. 2010, 19, 1873–1882. [Google Scholar] [CrossRef]
- Kragh, P.M.; Nielsen, A.L.; Li, J.; Du, Y.; Lin, L.; Schmidt, M.; Bogh, I.B.; Holm, I.E.; Jakobsen, J.E.; Johansen, M.G.; Purup, S.; Bolund, L.; Vajta, G.; Jorgensen, A.L. Hemizygous minipigs produced by random gene insertion and handmade cloning express the Alzheimer's disease-causing dominant mutation APPsw. Transgenic. Res. 2009, 18, 545–558. [Google Scholar] [CrossRef]
- Palmiter, R.D.; Brinster, R.L.; Hammer, R.E.; Trumbauer, M.E.; Rosenfeld, M.G.; Birnberg, N.C.; Evans, R.M. Dramatic growth of mice that develop from eggs microinjected with metallothionein-growth hormone fusion genes. Nature 1982, 300, 611–615. [Google Scholar] [CrossRef]
- Gama Sosa, M.A.; De Gasperi, R.; Elder, G.A. Animal transgenesis: An overview. Brain. Struct. Funct. 2010, 214, 91–109. [Google Scholar] [CrossRef]
- Wall, R.J.; Shani, M. Are animal models as good as we think? Theriogenology 2008, 69, 2–9. [Google Scholar]
- Habermann, F.A.; Wuensch, A.; Sinowatz, F.; Wolf, E. Reporter genes for embryogenesis research in livestock species. Theriogenology 2007, 68, S116–S124. [Google Scholar] [CrossRef]
- Laible, G.; Alonso-Gonzalez, L. Gene targeting from laboratory to livestock: Current status and emerging concepts. Biotechnol. J. 2009, 4, 1278–1292. [Google Scholar] [CrossRef]
- Nowak-Imialek, M.; Kues, W.; Carnwath, J.W.; Niemann, H. Pluripotent stem cells and reprogrammed cells in farm animals. Microsc. Microanal. 2011, 17, 474–497. [Google Scholar]
- Hammer, R.E.; Pursel, V.G.; Rexroad, C.E., Jr.; Wall, R.J.; Bolt, D.J.; Ebert, K.M.; Palmiter, R.D.; Brinster, R.L. Production of transgenic rabbits sheep and pigs by microinjection. Nature 1985, 315, 680–683. [Google Scholar] [CrossRef]
- Chang, K.; Qian, J.; Jiang, M.; Liu, Y.H.; Wu, M.C.; Chen, C.D.; Lai, C.K.; Lo, H.L.; Hsiao, C.T.; Brown, L.; Bolen, J., Jr.; Huang, H.I.; Ho, P.Y.; Shih, P.Y.; Yao, C.W.; Lin, W.J.; Chen, C.H.; Wu, F.Y.; Lin, Y.J.; Xu, J.; Wang, K. Effective generation of transgenic pigs and mice by linker based sperm-mediated gene transfer. BMC Biotechnol. 2002, 2, 5. [Google Scholar] [CrossRef]
- Watanabe, M.; Kurome, M.; Matsunari, H.; Nakano, K.; Umeyema, K.; Shiota, A.; Nakauchi, H.; Nagashima, H. The creation of transgenic pigs expressing human proteins using BAC-derived full-length genes and intracytoplasmic sperm injection-mediated gene transfer. Transgenic. Res. 2012, 21, 605–618. [Google Scholar] [CrossRef]
- Garcia-Vazquez, F.A.; Ruiz, S.; Matas, C.; Izquierdo-Rico, M.J.; Grullon, L.A.; De, O.A.; Vieira, L.; viles-Lopez, K.; Gutierrez-Adan, A.; Gadea, J. Production of transgenic piglets using ICSI-sperm-mediated gene transfer in combination with recombinase RecA. Reproduction. 2010, 140, 259–272. [Google Scholar] [CrossRef]
- Park, F. Lentiviral vectors: Are they the future of animal transgenesis? Physiol. Genomics. 2007, 31, 159–173. [Google Scholar] [CrossRef]
- Hofmann, A.; Kessler, B.; Ewerling, S.; Weppert, M.; Vogg, B.; Ludwig, H.; Stojkovic, M.; Boelhauve, M.; Brem, G.; Wolf, E.; Pfeifer, A. Efficient transgenesis in farm animals by lentiviral vectors. EMBO Rep. 2003, 4, 1054–1060. [Google Scholar] [CrossRef]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Schnieke, A.E.; Kind, A.J.; Ritchie, W.A.; Mycock, K.; Scott, A.R.; Ritchie, M.; Wilmut, I.; Colman, A.; Campbell, KH. Human factor IX transgenic sheep produced by transfer of nuclei from transfected fetal fibroblasts. Science 1997, 278, 2130–2133. [Google Scholar] [CrossRef]
- Petersen, B.; Lucas-Hahn, A.; Oropeza, M.; Hornen, N.; Lemme, E.; Hassel, P.; Queisser, A.L.; Niemann, H. Development and validation of a highly efficient protocol of porcine somatic cloning using preovulatory embryo transfer in peripubertal gilts. Cloning. Stem. Cells. 2008, 10, 355–362. [Google Scholar] [CrossRef]
- Garrels, W.; Ivics, Z.; Kues, W.A. Precision genetic engineering in large mammals. Trends. Biotechnol. 2012, 7, 386–393. [Google Scholar]
- Shinohara, E.T.; Kaminski, J.M.; Segal, D.J.; Pelczar, P.; Kolhe, R.; Ryan, T.; Coates, C.J.; Fraser, M.J.; Handler, A.M.; Yanagimachi, R.; Moisyadi, S. Active integration: New strategies for transgenesis. Transgenic. Res. 2007, 16, 333–339. [Google Scholar] [CrossRef] [Green Version]
- Fahrenkrug, S.C.; Blake, A.; Carlson, D.F.; Doran, T.; Van, E.A.; Faber, D.; Galli, C.; Gao, Q.; Hackett, P.B.; Li, N.; Maga, E.A.; Muir, W.M.; Murray, J.D.; Shi, D.; Stotish, R.; Sullivan, E.; Taylor, J.F.; Walton, M.; Wheeler, M.; Whitelaw, B.; Glenn, B.P. Precision genetics for complex objectives in animal agriculture. J Anim. Sci. 2010, 88, 2530–2539. [Google Scholar] [CrossRef]
- Staunstrup, N.H.; Madsen, J.; Primo, M.N.; Li, J.; Liu, Y.; Kragh, P.M.; Li, R.; Schmidt, M.; Purup, S.; Dagnaes-Hansen, F.; Svensson, L.; Petersen, T.K.; Callesen, H.; Bolund, L.; Mikkelsen, J.G. Development of transgenic cloned pig models of skin inflammation by DNA transposon-directed ectopic expression of human beta1 and alpha2 integrin. PLoS One 2012, 7, e36658. [Google Scholar]
- Garrels, W.; Mates, L.; Holler, S.; Dalda, A.; Taylor, U.; Petersen, B.; Niemann, H.; Izsvak, Z.; Ivics, Z.; Kues, W.A. Germline transgenic pigs by, Sleeping, Beauty transposition in porcine zygotes and targeted integration in the pig genome. PLoS One 2011, 6, e23573. [Google Scholar]
- Carlson, D.F.; Geurts, A.M.; Garbe, J.R.; Park, C.W.; Rangel-Filho, A.; O'Grady, S.M.; Jacob, H.J.; Steer, C.J.; Largaespada, D.A.; Fahrenkrug, S.C. Efficient mammalian germline transgenesis by cis-enhanced Sleeping Beauty transposition. Transgenic. Res. 2011, 20, 29–45. [Google Scholar] [CrossRef]
- McGrew, M.J.; Sherman, A.; Ellard, F.M.; Lillico, S.G.; Gilhooley, H.J.; Kingsman, A.J.; Mitrophanous, KA.; Sang, H. Efficient production of germline transgenic chickens using lentiviral vectors. EMBO Rep. 2004, 5, 728–733. [Google Scholar] [CrossRef]
- Tasic, B.; Hippenmeyer, S.; Wang, C.; Gamboa, M.; Zong, H.; Chen-Tsai, Y.; Luo, L. Site-specific integrase-mediated transgenesis in mice via pronuclear injection. Proc. Natl. Acad. Sci. USA 2011, 108, 7902–7907. [Google Scholar]
- Macdonald, J.; Taylor, L.; Sherman, A.; Kawakami, K.; Takahashi, Y.; Sang, H.M.; McGrew, M.J. Efficient genetic modification and germ-line transmission of primordial germ cells using piggyBac and Tol2 transposons. Proc. Natl. Acad. Sci. USA 2012, 109, E1466–E1472. [Google Scholar]
- Carlson, D.F.; Garbe, J.R.; Tan, W.; Martin, M.J.; Dobrinsky, J.R.; Hackett, P.B.; Clark, K.J.; Fahrenkrug, S.C. Strategies for selection marker-free swine transgenesis using the Sleeping Beauty transposon system. Transgenic. Res. 2011, 20, 1125–1137. [Google Scholar] [CrossRef]
- Jakobsen, J.E.; Li, J.; Kragh, P.M.; Moldt, B.; Lin, L.; Liu, Y.; Schmidt, M.; Winther, K.D.; Schyth, B.D.; Holm, I.E.; Vajta, G.; Bolund, L.; Callesen, H.; Jorgensen, A.L.; Nielsen, A.L.; Mikkelsen, J.G. Pig transgenesis by Sleeping Beauty DNA transposition. Transgenic. Res. 2011, 20, 533–545. [Google Scholar]
- Van Reenen, C.G.; Meuwissen, T.H.; Hopster, H.; Oldenbroek, K.; Kruip, T.H.; Blokhuis, H.J. Transgenesis may affect farm animal welfare: A case for systematic risk assessment. J. Anim. Sci. 2001, 79, 1763–1779. [Google Scholar]
- Clark, J.; Whitelaw, B. A future for transgenic livestock. Nat. Rev. Genet. 2003, 4, 825–833. [Google Scholar] [CrossRef]
- Carter, D.B.; Lai, L.; Park, K.W.; Samuel, M.; Lattimer, J.C.; Jordan, K.R.; Estes, D.M.; Besch-Williford, C.; Prather, R.S. Phenotyping of transgenic cloned piglets. Cloning. Stem. Cells 2002, 4, 131–145. [Google Scholar] [CrossRef]
- Greger, M. Trait selection and welfare of genetically engineered animals in agriculture. J. Anim. Sci. 2010, 88, 811–814. [Google Scholar] [CrossRef]
- Deppenmeier, S.; Bock, O.; Mengel, M.; Niemann, H.; Kues, W.; Lemme, E.; Wirth, D.; Wonigeit, K.; Kreipe, H. Health status of transgenic pigs expressing the human complement regulatory protein CD59. Xenotransplantation. 2006, 13, 345–356. [Google Scholar] [CrossRef]
- Huber, R.; Remuge, L.; Carlisle, A.; Lillico, S.; Sandoe, P.; Sorensen, D.; Withelaw, B.; Olosson, A. Welfare assessment in transgenic pigs expressing green fluorescent protein GFP. Transgenic. Res. 2012, 21, 773–784. [Google Scholar] [CrossRef]
- Reichenbach, M.; Lim, T.; Reichenbach, H.D.; Guengoer, T.; Habermann, F.A.; Matthiesen, M.; Hofmann, A.; Weber, F.; Zerbe, H.; Grupp, T.; Sinowatz, F.; Pfeifer, A.; Wolf, E. Germ-line transmission of lentiviral, PGK-EGFP integrants in transgenic cattle: new perspectives for experimental embryology. Transgenic. Res. 2010, 19, 549–556. [Google Scholar] [CrossRef]
- Pursel, V.G.; Wall, R.J.; Solomon, M.B.; Bolt, D.J.; Murray, J.E.; Ward, K.A. Transfer of an ovine metallothionein-ovine growth hormone fusion gene into swine. J Anim. Sci. 1997, 75, 2208–2214. [Google Scholar]
- Grupen, C.G.; Verma, P.J.; Du, Z.T.; McIlfatrick, S.M.; Ashman, R.J.; Nottle, M.B. Activation of in vivo- and in vitro-derived porcine oocytes by using multiple electrical pulses. Reprod. Fertil. Dev. 1999, 11, 457–462. [Google Scholar] [CrossRef]
- Hill, J.R.; Roussel, A.J.; Cibelli, J.B.; Edwards, J.F.; Hooper, N.L.; Miller, M.W.; Thompson, J.A.; Looney, C.R.; Westhusin, M.E.; Robl, J.M.; Stice, S.L. Clinical and pathologic features of cloned transgenic calves and fetuses (13 case studies). Theriogenology 1999, 51, 1451–1465. [Google Scholar]
- Chavatte-Palmer, P.; Camous, S.; Jammes, H.; Le, C.N.; Guillomot, M.; Lee, R.S. Review: Placental perturbations induce the developmental abnormalities often observed in bovine somatic cell nuclear transfer. Placenta 2012, 33, S99–S104. [Google Scholar] [CrossRef]
- Watanabe, S.; Nagai, T. Health status and productive performance of somatic cell cloned cattle and their offspring produced in Japan. J. Reprod. Dev. 2008, 54, 6–17. [Google Scholar] [CrossRef]
- Cao, Z.; Li, Y.; Wen, X.; Li, Z.; Mi, C.; Zhang, Z.; Li, N.; Li, Q. Recloned transgenic pigs possess normal reproductive performance and stable genetic transmission capacity. Zygote 2012, 12, 1–7. [Google Scholar]
- Iqbal, K.; Barg-Kues, B.; Broll, S.; Bode, J.; Niemann, H.; Kues, W.A. Cytoplasmic injection of circular plasmids allows targeted expression in mammalian embryos. Biotechniques 2009, 47, 959–968. [Google Scholar] [CrossRef]
- Mates, L.; Chuah, M.K.; Belay, E.; Jerchow, B.; Manoj, N.; Costa-Sanchez, A.; Grzela, D.P.; Schmitt, A.; Becker, K.; Matrai, J.; Ma, L.; Samara-Kuko, E.; Gysemans, C.; Pryputniewicz, D.; Miskey, C.; Fletcher, B.; Vandendriessche, T.; Ivics, Z.; Izsvak, Z. Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. Nat. Genet. 2009, 41, 753–761. [Google Scholar]
- Tamashiro, K.L.; Wakayama, T.; Blanchard, R.J.; Blanchard, D.C.; Yanagimachi, R. Postnatal growth and behavioral development of mice cloned from adult cumulus cells. Biol. Reprod. 2000, 63, 328–334. [Google Scholar] [CrossRef]
- Rhind, S.M.; King, T.J.; Harkness, L.M.; Bellamy, C.; Wallace, W.; DeSousa, P.; Wilmut, I. Cloned lambs-lessons from pathology. Nat. Biotechnol. 2003, 21, 744–745. [Google Scholar] [CrossRef]
- Garrels, W.; Holler, S.; Taylor, U.; Herrmann, D.; Struckmann, C.; Klein, S.; Barg-Kues, B.; Nowak-Imialek, M.; Ehling, C.; Rath, D.; Ivics, Z.; Niemann, H.; Kues, W.A. Genotype-independent transmission of transgenic fluorophore protein by boar spermatozoa. PLoS One 2011, 6, e27563. [Google Scholar]
- Pursel, V.G.; Pinkert, C.A.; Miller, K.F.; Bolt, D.J.; Campbell, R.G.; Palmiter, R.D.; Brinster, R.L.; Hammer, R.E. Genetic engineering of livestock. Science 1989, 244, 1281–1288. [Google Scholar]
- Nagai, T.; Ibata, K.; Park, E.S.; Kubota, M.; Mikoshiba, K.; Miyawaki, A. A variant of yellow fluorescent protein with fast and efficient maturation for cell-biological applications. Nat. Biotechnol. 2002, 20, 87–90. [Google Scholar] [CrossRef]
- Garrels, W.; Cleve, N., Niemann; Kues, W.A. Rapid non-invasive genotyping of reporter transgenic mammals. Biotechniques 2012, 0, 1–4. [Google Scholar]
- Kues, W.A.; Schwinzer, R.; Wirth, D.; Verhoeyen, E.; Lemme, E.; Herrmann, D.; Barg-Kues, B.; Hauser, H.; Wonigeit, K.; Niemann, H. Epigenetic silencing and tissue independent expression of a novel tetracycline inducible system in double-transgenic pigs. FASEB J. 2006, 20, 1200–1202. [Google Scholar] [CrossRef]
- Alonso-Gonzalez, L.; Couldrey, C.; Meinhardt, M.W.; Cole, S.A.; Wells, D.N.; Laible, G. Primary transgenic bovine cells and their rejuvenated cloned equivalents show transgene-specific epigenetic differences. PLoS One 2012, 7, e35619. [Google Scholar]
- Luo, Y.; Bolund, L.; Sorensen, C.B. Pig gene knockout by rAAV-mediated homologous recombination: Comparison of BRCA1 gene knockout efficiency in Yucatan and Gottingen fibroblasts with slightly different target sequences. Transgenic. Res. 2012, 21, 671–676. [Google Scholar] [CrossRef]
- Yang, D.; Wang, C.E.; Zhao, B.; Li, W.; Ouyang, Z.; Liu, Z.; Yang, H.; Fan, P.; O'Neill, A.; Gu, W.; Yi, H.; Li, S.; Lai, L.; Li, X.J. Expression of Huntington's disease protein results in apoptotic neurons in the brains of cloned transgenic pigs. Hum. Mol. Genet. 2010, 19, 3983–3994. [Google Scholar]
- Kues, W.A.; Niemann, H. The contribution of farm animals to human health. Trends. Biotechnol. 2004, 22, 286–294. [Google Scholar] [CrossRef]
- Yang, D.; Yang, H.; Li, W.; Zhao, B.; Ouyang, Z.; Liu, Z.; Zhao, Y.; Fan, N.; Song, J.; Tian, J.; Li, F.; Zhang, J.; Chang, L.; Pei, D.; Chen, Y.E.; Lai, L. Generation of PPAR gamma mono-allelic knockout pigs via zinc-finger nucleases and nuclear transfer cloning. Cell. Res. 2011, 21, 979–982. [Google Scholar] [CrossRef]
- Flisikowska, T.; Thorey, I.S.; Offner, S.; Ros, F.; Lifke, V.; Zeitler, B.; Rottmann, O.; Vincent, A.; Zhang, L.; Jenkins, S.; Niersbach, H.; Kind, A.J.; Gregory, P.D.; Schnieke, A.E.; Platzer, J. Efficient immunoglobulin gene disruption and targeted replacement in rabbit using zinc finger nucleases. PLoS One 2011, 6, e21045. [Google Scholar]
- Renner, S.; Fehlings, C.; Herbach, N.; Hofmann, A.; Von Waldthausen, D.C.; Kessler, B.; Ulrichs, K.; Chodnevskaja, I.; Moskalenko, V.; Amselgruber, W.; Goke, B.; Pfeifer, A.; Wanke, R.; Wolf, E. Glucose intolerance and reduced proliferation of pancreatic beta-cells in transgenic pigs with impaired glucose-dependent insulinotropic polypeptide function. Diabetes 2010, 59, 1228–1238. [Google Scholar] [CrossRef]
- Klymiuk, N.; Mundhenk, L.; Kraehe, K.; Wuensch, A.; Plog, S.; Emrich, D.; Langenmayer, M.C.; Stehr, M.; Holzinger, A.; Kroner, C.; Richter, A.; Kessler, B.; Kurome, M.; Eddicks, M.; Nagashima, H.; Heinritzi, K.; Gruber, A.D.; Wolf, E. Sequential targeting of CFTR by BAC vectors generates a novel pig model of cystic fibrosis. J. Mol. Med. (Berl.) 2012, 90, 597–608. [Google Scholar]
- Klymiuk, N.; Van Buerck, L.; Bahr, A.; Offers, M.; Kessler, B.; Wuensch, A.; Kurome, M.; Thormann, M.; Lochner, K.; Nagashima, H.; Herbach, N.; Wanke, R.; Seissler, J.; Wolf, E. Xenografted islet cell clusters from INSLEA29Y transgenic pigs rescue diabetes and prevent immune rejection in humanized mice. Diabetes 2012, 61, 1527–1532. [Google Scholar] [CrossRef]
- Wheeler, D.G.; Joseph, M.E.; Mahamud, S.D.; Aurand, W.L.; Mohler, P.J.; Pompili, V.J.; Dwyer, K.M.; Nottle, M.B.; Harrison, S.J.; d'Apice, A.J.; Robson, S.C.; Cowan, P.J.; Gumina, R.J. Transgenic swine: Expression of human CD39 protects against myocardial injury. J. Mol. Cell. Cardiol. 2012, 52, 958–961. [Google Scholar] [CrossRef]
- Rogers, C.S.; Stoltz, D.A.; Meyerholz, D.K.; Ostedgaard, L.S.; Rokhlina, T.; Taft, P.J.; Rogan, M.P.; Pezzulo, A.A.; Karp, P.H.; Itani, O.A.; Kabel, A.C.; Wohlford-Lenane, C.L.; Davis, G.J.; Hanfland, R.A.; Smith, T.L.; Samuel, M.; Wax, D.; Murphy, C.N.; Rieke, A.; Whitworth, K.; Uc, A.; Starner, T.D.; Brogden, K.A.; Shilyansky, J.; McCray, P.B., Jr.; Zabner, J.; Prather, R.S.; Welsh, M.J. Disruption of the CFTR gene produces a model of cystic fibrosis in newborn pigs. Science 2008, 321, 1837–1841. [Google Scholar]
- Sommer, J.R.; Estrada, J.L.; Collins, E.B.; Bedell, M.; Alexander, C.A.; Yang, Z.; Hughes, G.; Mir, B.; Gilger, B.C.; Grob, S.; Wei, X.; Piedrahita, J.A.; Shaw, P.X.; Petters, R.M.; Zhang, K. Production of ELOVL4 transgenic pigs: a large animal model for Stargardt-like macular degeneration. Br. J. Ophthalmol. 2011, 95, 1749–1754. [Google Scholar] [CrossRef]
- Mendicino, M.; Ramsoondar, J.; Phelps, C.; Vaught, T.; Ball, S.; LeRoith, T.; Monahan, J.; Chen, S.; Dandro, A.; Boone, J.; Jobst, P.; Vance, A.; Wertz, N.; Bergman, Z.; Sun, X.Z.; Polejaeva, I.; Butler, J.; Dai, Y.; Ayares, D.; Wells, K. Generation of antibody- and B cell-deficient pigs by targeted disruption of the, J-region gene segment of the heavy chain locus. Transgenic. Res. 2011, 20, 625–641. [Google Scholar] [CrossRef]
- Ramsoondar, J.; Mendicino, M.; Phelps, C.; Vaught, T.; Ball, S.; Monahan, J.; Chen, S.; Dandro, A.; Boone, J.; Jobst, P.; Vance, A.; Wertz, N.; Polejaeva, I.; Butler, J.; Dai, Y.; Ayares, D.; Wells, K. Targeted disruption of the porcine immunoglobulin kappa light chain locus. Transgenic. Res. 2011, 20, 643–653. [Google Scholar] [CrossRef]
- Suzuki, S.; Iwamoto, M.; Saito, Y.; Fuchimoto, D.; Sembon, S.; Suzuki, M.; Mikawa, S.; Hashimoto, M.; Aoki, Y.; Najima, Y.; Takagi, S.; Suzuki, N.; Suzuki, E.; Kubo, M.; Mimuro, J.; Kashiwakura, Y.; Madoiwa, S.; Sakata, Y.; Perry, A.C.F.; Ishikawa, F.; Onishi, A. Il2rg gene-targeted severe combined immunodeficiency pigs. Cell. Stem. Cell 2012, 10, 753–758. [Google Scholar] [CrossRef]
- Hackett, P.B., Jr.; Aronovich, E.L.; Hunter, D.; Urness, M.; Bell, J.B.; Kass, S.J.; Cooper, L.J.; McIvor, S. Efficacy and safety of Sleeping Beauty transposon-mediated gene transfer in preclinical animal studies. Curr. Gene. Ther. 2011, 11, 341–349. [Google Scholar]
- Luo, Y.; Kofod-Olsen, E.; Christensen, R.; Brandt-Sorensen, C.; Bolund, L. Targeted genome editing by recombinant adeno-associated virus (rAAV) vectors for generating genetically modified pigs. J. Genet. Genomics 2012, 39, 269–274. [Google Scholar] [CrossRef]
- Galli, C.; Lagutina, I.; Colleoni, S.; Duchi, R.; Luccini, F.; Lazzari, G. Somatic cell nuclear transfer and transgenesis in large animals: current and future insights. Reprod. Dom. Anim. 2012, 47, 2–11. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Garrels, W.; Holler, S.; Cleve, N.; Niemann, H.; Ivics, Z.; Kues, W.A. Assessment of Fecundity and Germ Line Transmission in Two Transgenic Pig Lines Produced by Sleeping Beauty Transposition. Genes 2012, 3, 615-633. https://doi.org/10.3390/genes3040615
Garrels W, Holler S, Cleve N, Niemann H, Ivics Z, Kues WA. Assessment of Fecundity and Germ Line Transmission in Two Transgenic Pig Lines Produced by Sleeping Beauty Transposition. Genes. 2012; 3(4):615-633. https://doi.org/10.3390/genes3040615
Chicago/Turabian StyleGarrels, Wiebke, Stephanie Holler, Nicole Cleve, Heiner Niemann, Zoltan Ivics, and Wilfried A. Kues. 2012. "Assessment of Fecundity and Germ Line Transmission in Two Transgenic Pig Lines Produced by Sleeping Beauty Transposition" Genes 3, no. 4: 615-633. https://doi.org/10.3390/genes3040615