
Review of Microfluidic Photobioreactor Technology for Metabolic Engineering and Synthetic Biology of Cyanobacteria and Microalgae

Abstract
:
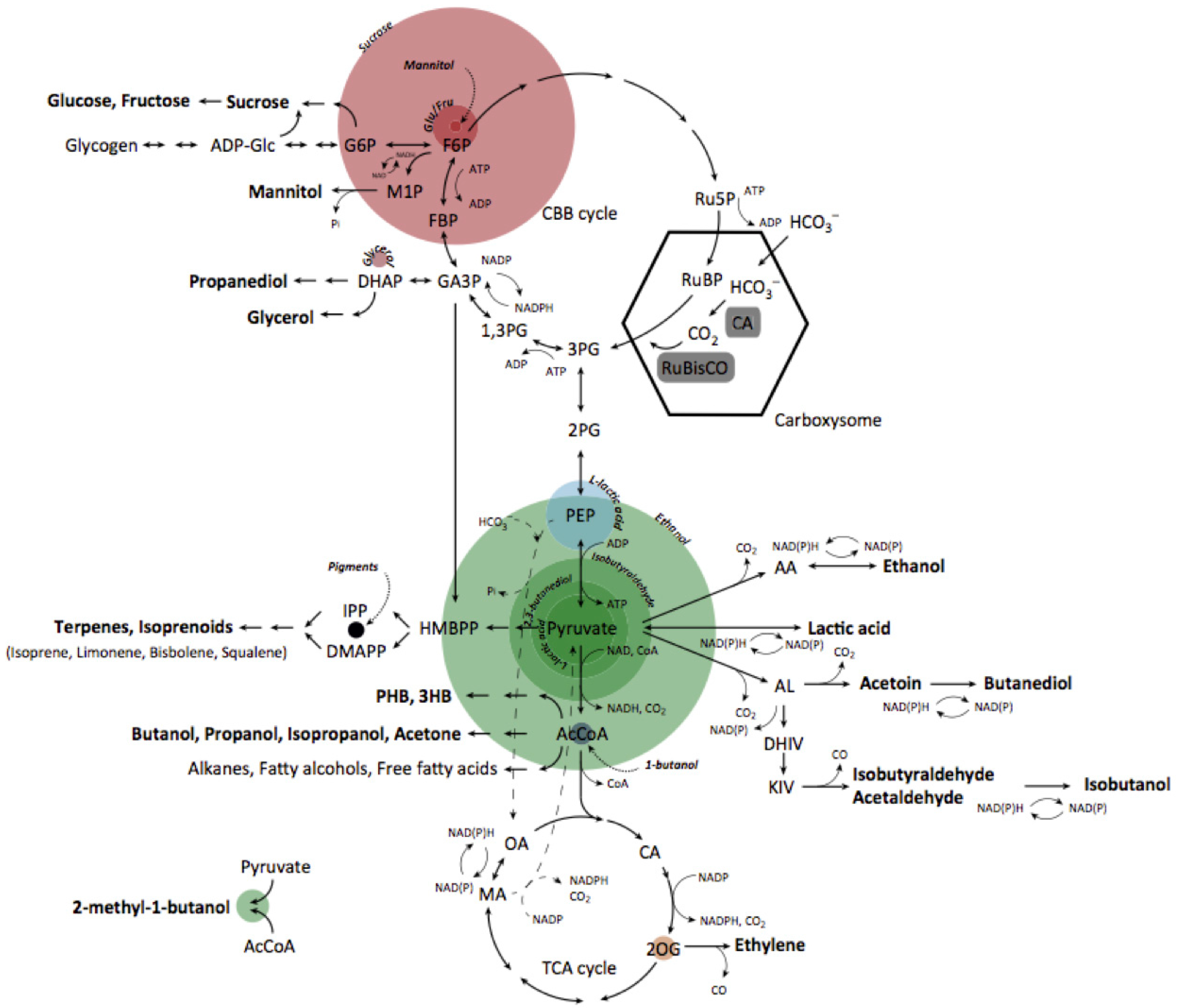
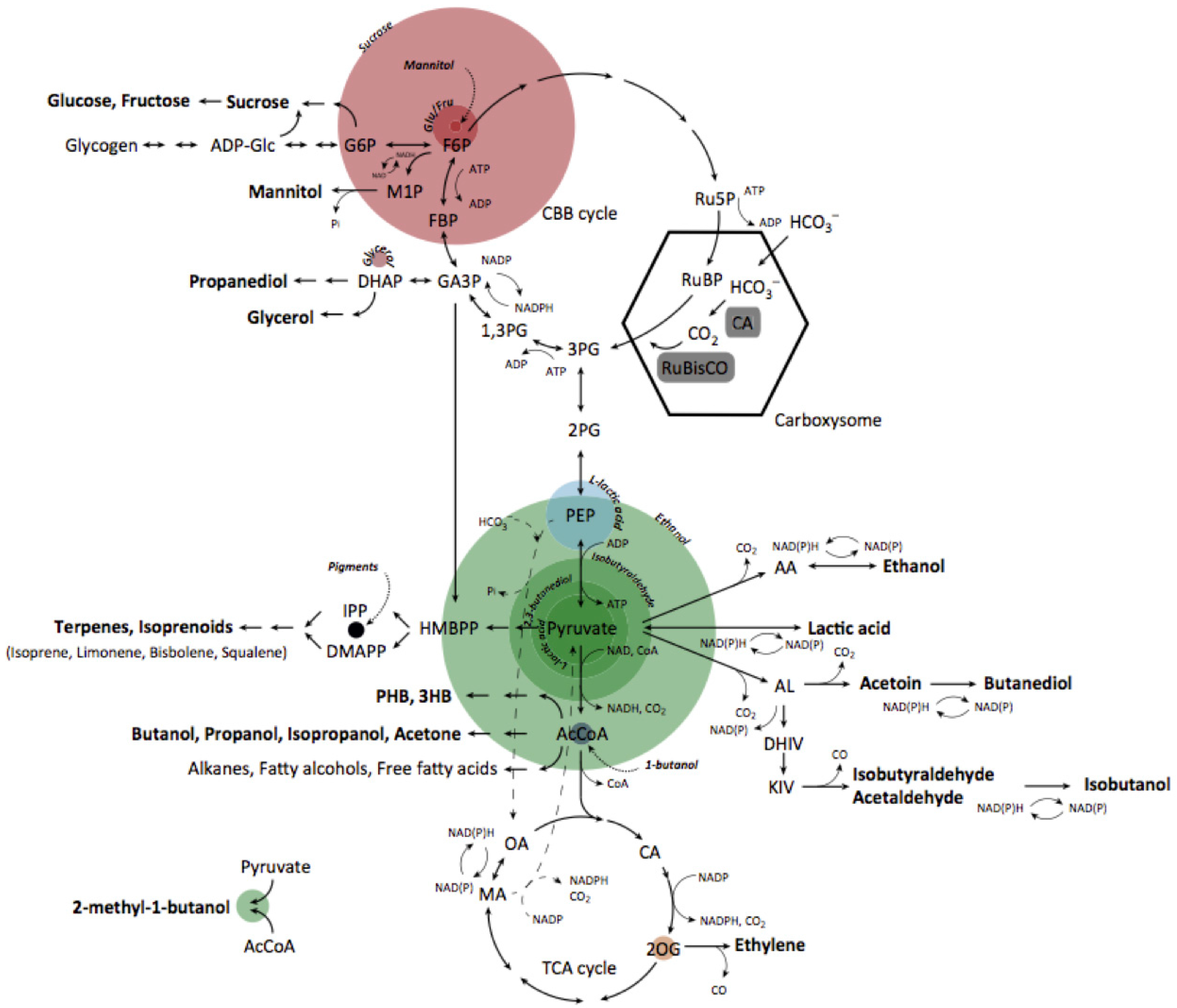
1. Introduction
2. Review of Main Body of Research
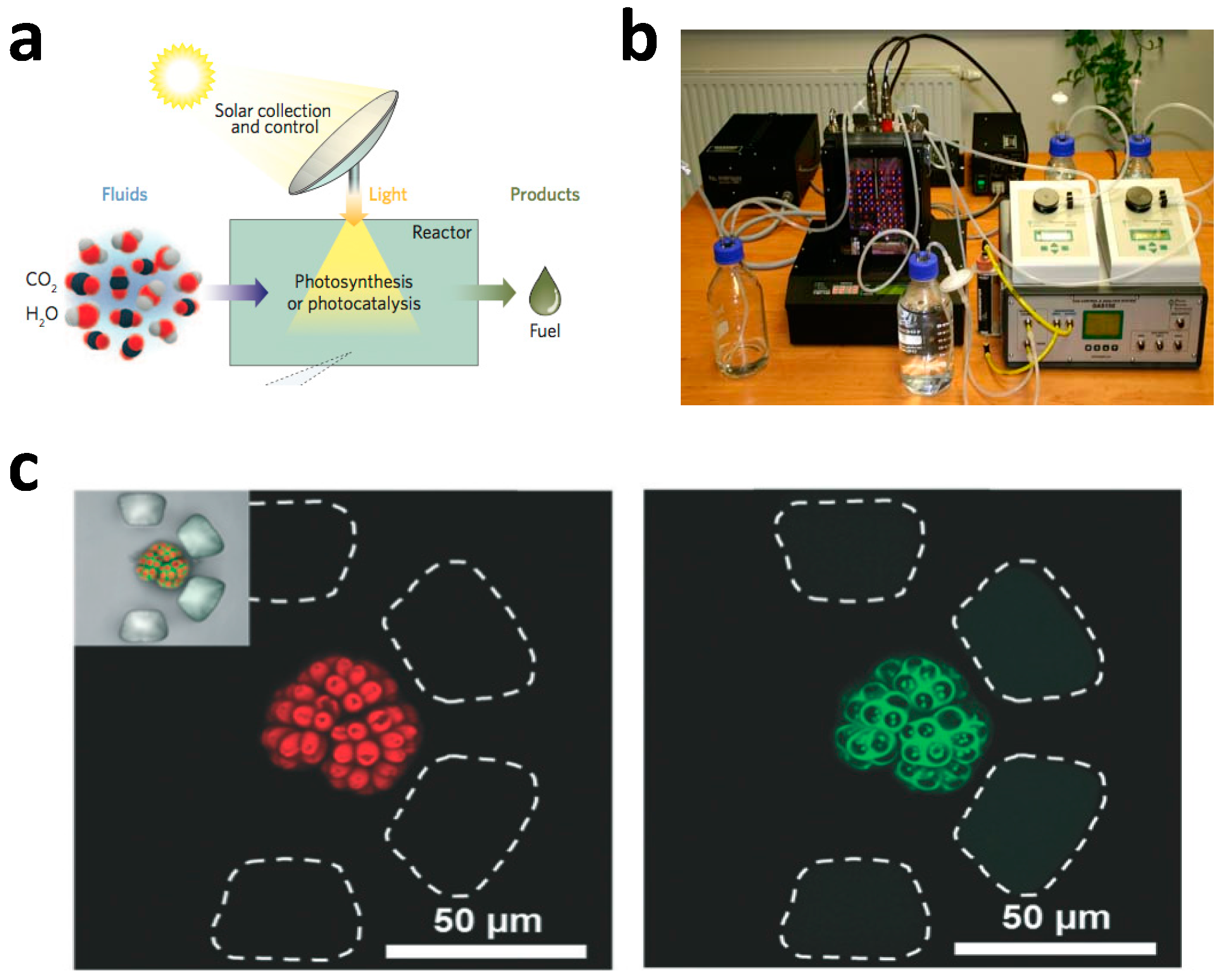
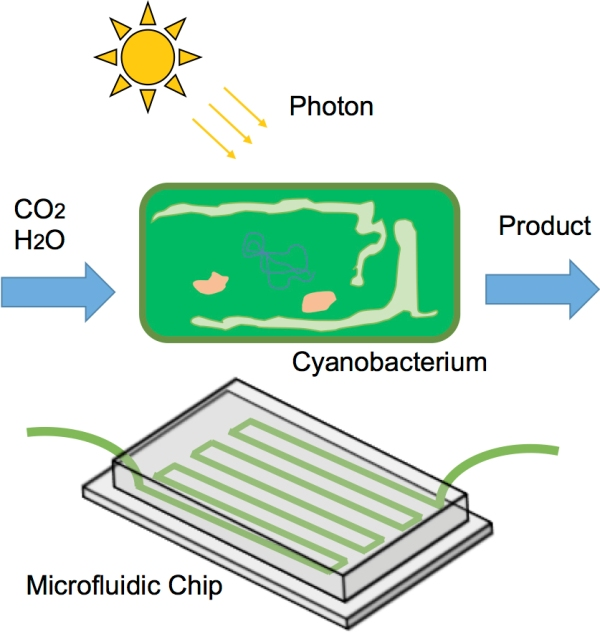
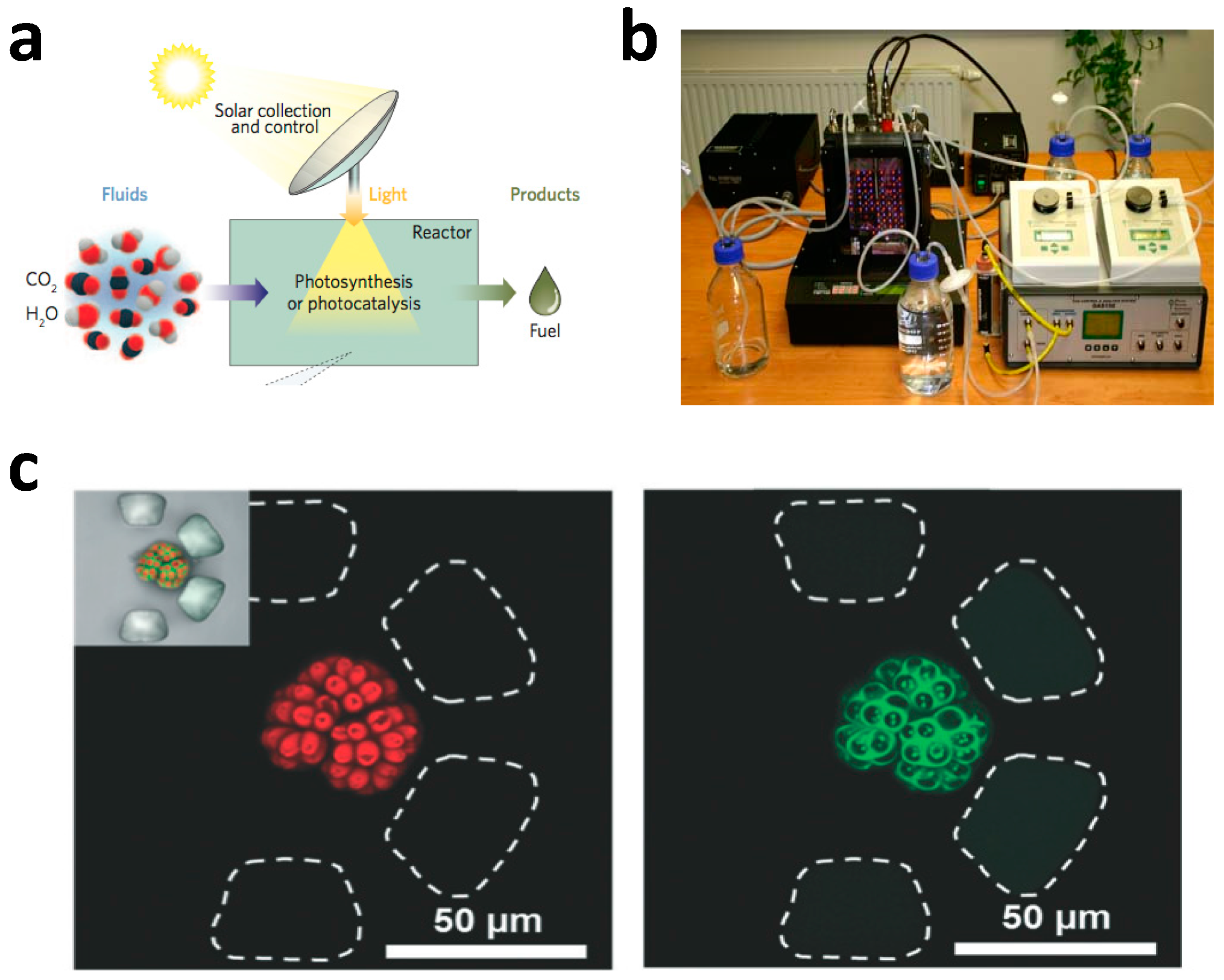
2.1. General Comments on Microfluidic Photobioreactors
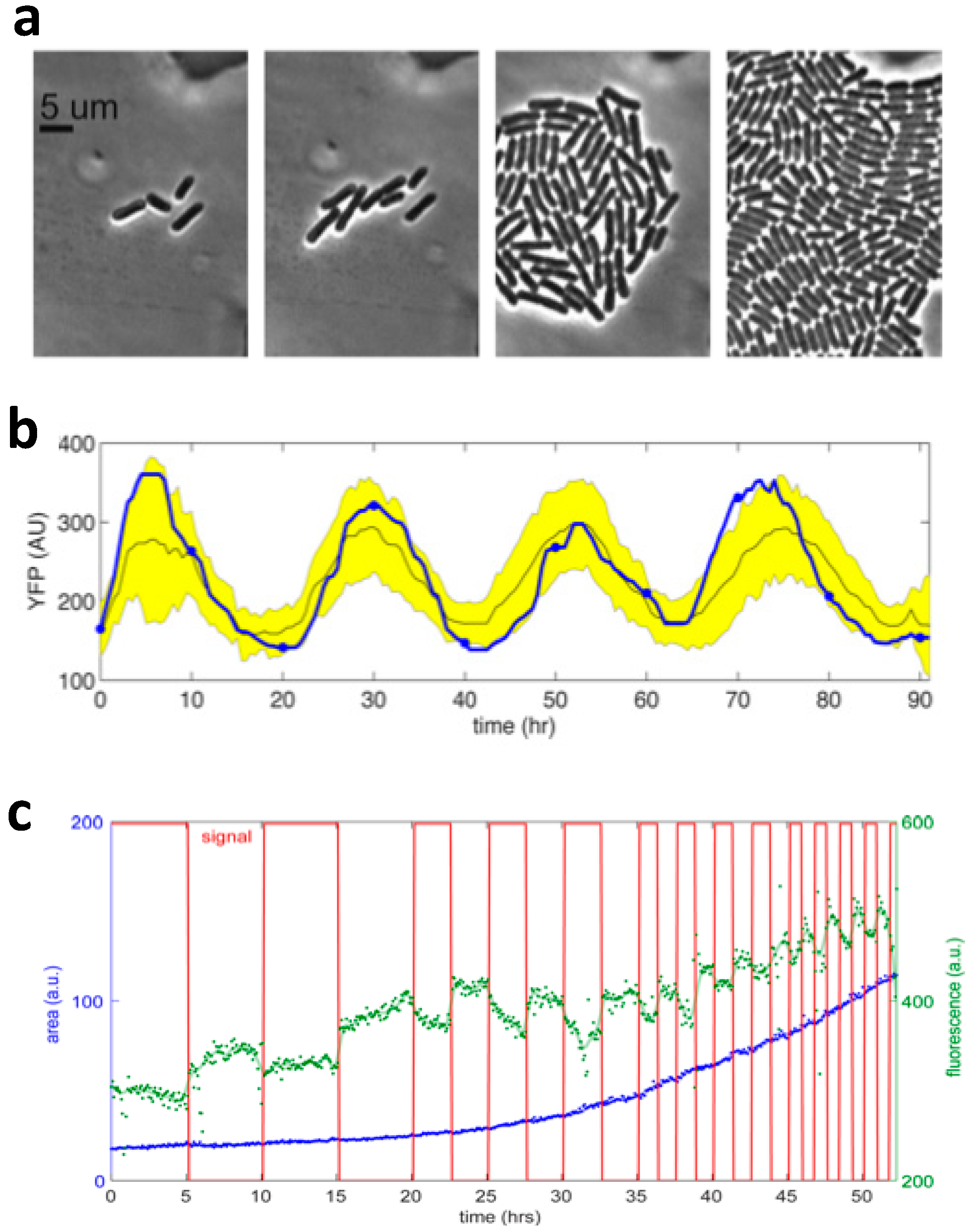
2.2. Microfluidic Photobioreactor Based on Microplate and Agar
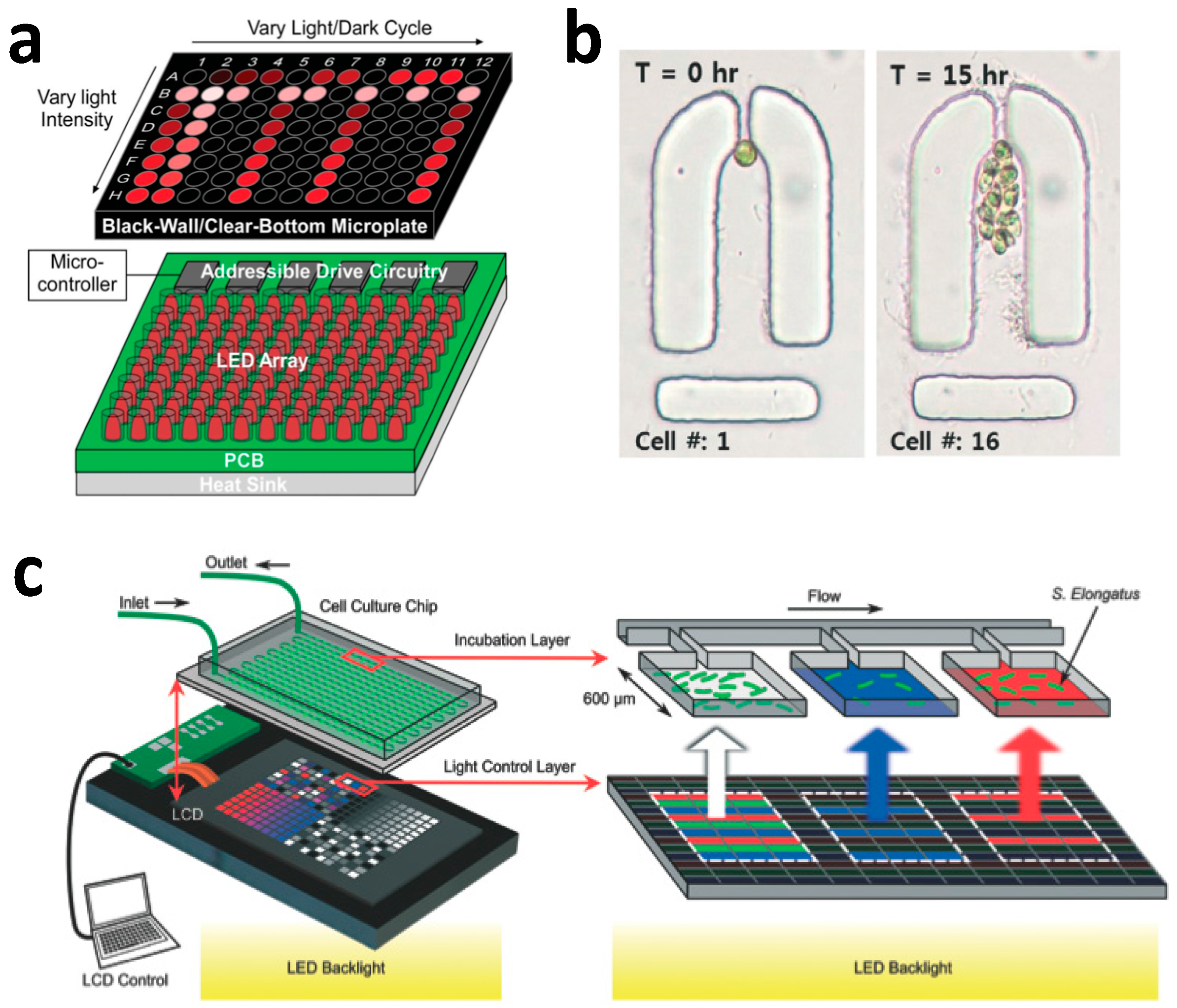
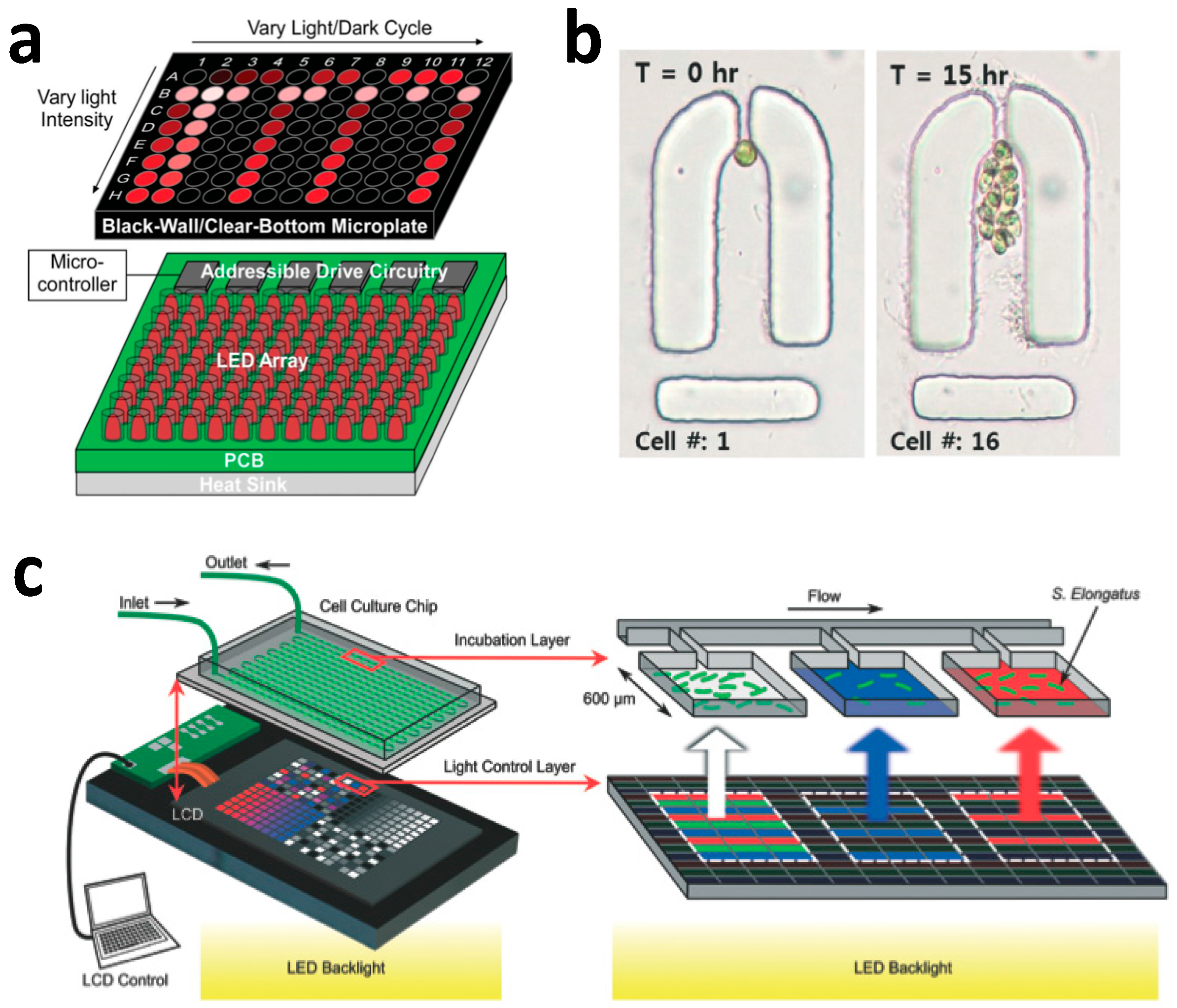
2.3. Microfluidic Photobioreactor Utilizing Flow-Based mLSI Technology
2.4. Microfluidic Photobioreactor Based on Droplet and Digital Microfluidics
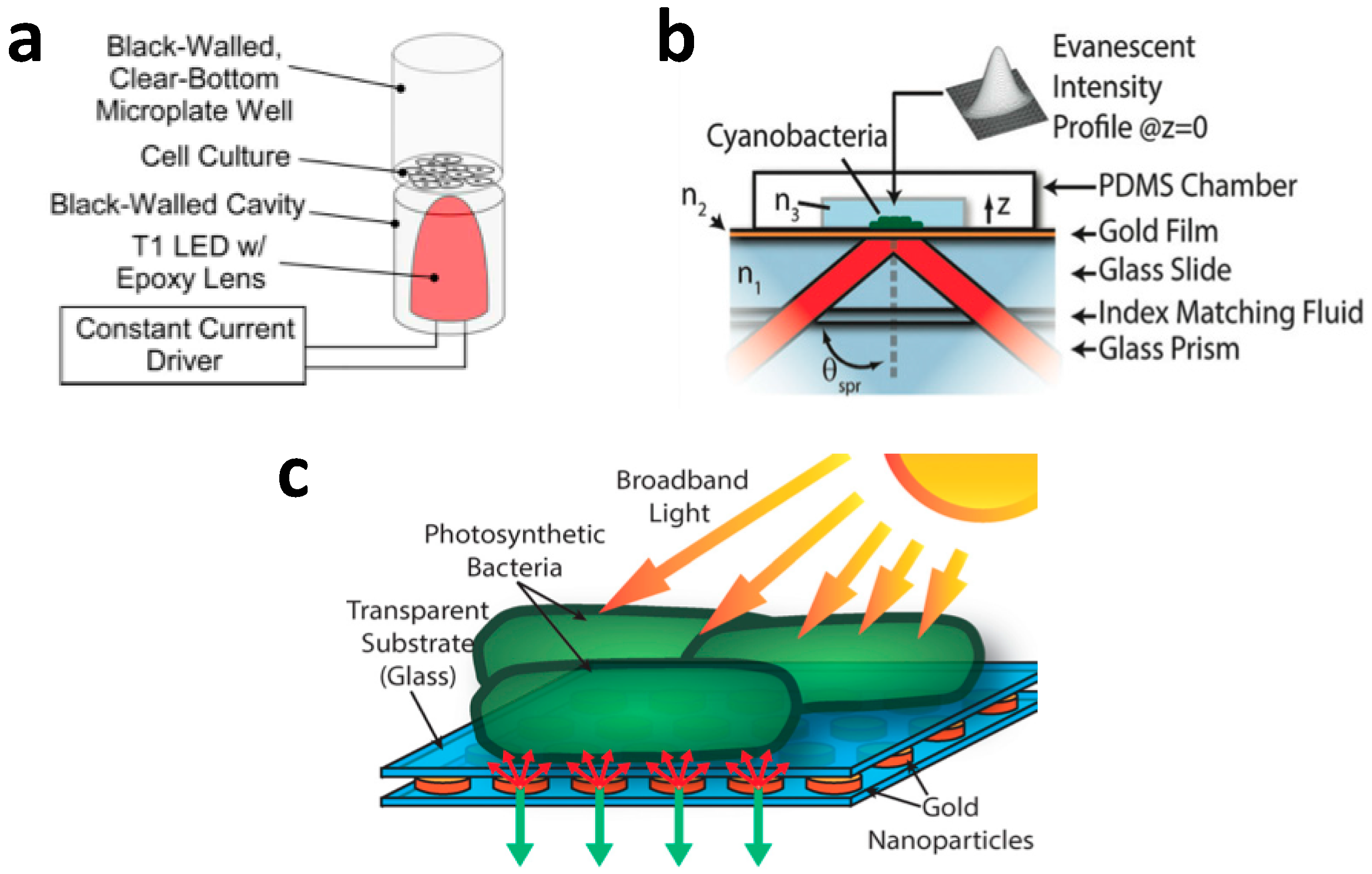
2.5. Microfluidic Photobioreactor with Alternative Illumination Method
3. Future Perspectives
3.1. Integration with Current Workflow of Synthetic Biology
3.2. Synthetic Photobiology and Optogenetics
3.3. End Point Titration and Metabolic Flux Measurement
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Hellingwerf, K.J.; Lindblad, P.; de Mattos, M.J.T. Energy biotechnology with cyanobacteria. Curr. Opin. Biotechnol. 2009, 20, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Rovira, A.G.; Hellingwerf, K.J. Metabolic engineering of cyanobacteria for the synthesis of commodity product. Trends Biotechnol. 2015, 33, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Berla, B.M.; Saha, R.; Immethuan, C.M.; Maranas, C.D.; Moon, T.S.; Pakrasi, H.B. Synthetic biology of cyanobacteria: Unique challenges and opportunities. Front. Microbiol. 2013, 4, 246. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.; Kondev, J.; Theriot, J. Physical Biology of the Cell, 2nd ed.; Garland Science: New York, NY, USA, 2012. [Google Scholar]
- Gao, Z.; Zhao, H.; Li, Z.; Tan, X.; Lu, X. Photosynthetic production of ethanol from carbon dioxide in genetically engineered cyanobacteria. Energy Environ. Sci. 2012, 5, 9857–9865. [Google Scholar] [CrossRef]
- Oliver, J.W.K.; Machado, I.M.P.; Yoneda, H.; Atsumi, S. Cyanobacterial conversion of carbon dioxide to 2,3-butanediol. Proc. Natl. Acad. Sci. USA 2013, 110, 1249–1254. [Google Scholar] [CrossRef] [PubMed]
- Angermayr, S.A.; Hellinwerf, K.J. On the use of metabolic control analysis in the optimization of cyanobacterial biosolar cell factories. J. Phys. Chem. B 2013, 117, 11169–11175. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.M.; Yu, Y.; You, L.; Tang, Y.J. Photoautotrophic production of d-lactic acid in an engineered cyanobacterium. Microb. Cell Fact. 2013, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Atsumi, S.; Higashide, W.; Liao, J.C. Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nat. Biotechnol. 2009, 27, 1177–1180. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, C.E.; Whited, G.M. Metabolic engineering for the microbial production of 1,3-propanediol. Curr. Opin. Biotechnol. 2003, 14, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Yim, H.; Haselbeck, R.; Niu, W.; Pujol-Baxley, C.; Burgard, A.; Boldt, J.; Khandurina, J.; Trawick, J.D.; Osterhout, R.E.; Stephen, R.; et al. Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nat. Chem. Biol. 2011, 7, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Densmore, D. Integration of microfluidics into synthetic biology design workflow. Lab Chip 2014, 14, 3459–3474. [Google Scholar] [CrossRef] [PubMed]
- Szita, A.N.; Polizzi, K.; Jaccard, N.; Baganz, F. Microfluidic approaches for systems and synthetic biology. Curr. Opin. Biotechnol. 2010, 21, 517–523. [Google Scholar] [CrossRef] [PubMed]
- Photo System Instruments. Available online: http://www.psi.cz/photobioreactors/ (accessed on 23 October 2015).
- Psaltis, D.; Quake, S.R.; Yang, C.H. Developing optofluidic technology through the fusion of microfluidics and optics. Nature 2006, 442, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Erickson, D.; Sinton, D.; Psaltis, D. Optofluidics for energy application. Nat. Photonics 2011, 5, 583–590. [Google Scholar] [CrossRef]
- Han, A.; Hou, H.; Li, L.; Kim, H.S.; de Figueiredo, P. Microfabricated devices in microbial bioenergy sciences. Trends Biotechnol. 2013, 31, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Weiss, T.L.; Thapa, H.R.; Devarenne, T.P.; Han, A. A microfluidic photobioreactor array demonstrating high-throughput screening for microalgal oil production. Lab Chip 2014, 14, 1415–1425. [Google Scholar] [CrossRef] [PubMed]
- Melin, J.; Quake, S.R. Microfluidic large-scale integration: The evaluation of design rules for biological automation. Annu. Rev. Biophys. Biomol. Struct. 2007, 7, 213–231. [Google Scholar] [CrossRef] [PubMed]
- Teh, S.Y.; Lin, R.; Hung, L.H.; Lee, A.P. Droplet microfluidics. Lab Chip 2008, 8, 198–200. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.; Ng, A.H.; Fobel, R.; Wheeler, A.R. Digital Microfluidics. Annu. Rev. Anal. Chem. 2012, 5, 413–440. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, A.R. Putting electrowetting to work. Science 2008, 322, 539–540. [Google Scholar] [CrossRef] [PubMed]
- Hegab, H.M.; ElMekawy, A.; Stakeborg, T. Review of microfluidic microbioreactor technology for high-throughput submerged microbiological cultivation. Biomicrofluidics 2013, 7, 021502. [Google Scholar] [CrossRef] [PubMed]
- Halldorsson, S.; Lucumi, E.; Gómez-Sjöberg, R.; Fleming, R.M.T. Advantages and challenges of microfluidic cell cuture in polydimethylsiloxane. Biosens. Bioelectron. 2015, 63, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Balagadde, F.K.; You, L.C.; Hansen, C.L.; Arnold, F.H.; Quake, S.R. Long-term monitoring of bacteria undergoing programmed population control in a microchemostat. Science 2005, 309, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Balagadde, F.K.; Song, H.; Ozaki, J.; Collins, C.H.; Barnet, M.; Arnold, F.H.; Quake, S.R.; You, L. A synthetic Eschericha coli predator-prey ecosystem. Mol. Syst. Biol. 2008, 4, 187. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Wu, J.; Polymenis, M.; Han, A. Microchemostat array with small-volume fraction replenishment for steady-state microbial culture. Lab Chip 2013, 13, 4217–4224. [Google Scholar] [CrossRef] [PubMed]
- Gu, G.; Lee, Y.W.; Chiang, C.C.; Yang, Y.T. A nanoliter microfluidic serial dilution bioreactor. Biomicrofluidics 2015, 9, 044126. [Google Scholar] [CrossRef] [PubMed]
- Groisman, A.; Lobo, C.; Cho, H.; Campbell, J.K.; Dufour, Y.S.; Stevens, A.M.; Levchenko, A. A microfluidic chemostat for experiments with bacterial and yeast cells. Nat. Method 2005, 9, 685–689. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, J.R.; Lee, J.B.; Cluzel, P. The single-cell chemostat: An agarose-based, microfluidic device for high-throughput, single-cell studies of bacteria and bacterial communities. Lab Chip 2012, 12, 1487–1494. [Google Scholar] [CrossRef] [PubMed]
- Long, Z.; Nugent, E.; Javer, A.; Cicuta, P.; Sclavi, B.; Lagomarsino, M.C.; Dorfman, K.D. Microfluidic chemostat for measuring single cell dynamics in bacteria. Lab Chip 2013, 13, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Karimi, A.; Karig, D.; Kumar, A.; Ardekani, A.M. Interplay of physical mechanisms and biofilm processes: Review of microfluidic methods. Lab Chip 2015, 15, 23–42. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Park, H.D.; Chung, S. Microfluidic approaches to bacterial biofilm formation. Molecules 2012, 17, 9818. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.H.; Hegde, M.; Kim, J.; Wang, X.; Jayaraman, A.; Wood, T.K. Synthetic quorum-sensing circuit to control consortial biofilm formation and dispersal in a microfluidic device. Nat. Commun. 2012, 3, 613. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.L.; Ghaderi, A.; Zhou, H.; Agresti, J.; Weitz, D.A.; Fink, G.R.; Stephanopoulos, G. Microfluidic high-throughput culturing of single cells for selection based on extracellular metabolite production or consumption. Nat. Biotechnol. 2014, 32, 473–480. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Mertiri, T.; Holland, T.; Basu, A.S. Optical microplates for high throughput screening of photosynthesis in lipid producing algae. Lab Chip 2012, 12, 3870–3874. [Google Scholar] [CrossRef] [PubMed]
- Teng, S.W.; Mukherji, S.; Moffitt, J.R.; de Buyl, S.; O’Shea, E.K. Robust circadian oscillations in growing cyanobacteria require transcriptional feedback. Science 2013, 340, 737–740. [Google Scholar] [CrossRef] [PubMed]
- Yokoo, R.; Hood, R.D.; Savage, D.F. Live cell imaging of cyanobacteria. Photosynth. Res. 2015, 126, 33–46. [Google Scholar] [CrossRef] [PubMed]
- Holcomb, R.; Mason, L.J.; Reardon, K.F.; Cropek, D.M.; Henry, C.S. Culturing and investigation of stress-induced lipid accumulation in microalgae using microfluidic device. Anal. Bioanal. Chem. 2011, 400, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Devarnne, T.P.; Han, A. A high-throughput microfluidic single-cell screening platform capable of selective cell extraction. Lab Chip 2015, 15, 2467–2475. [Google Scholar] [CrossRef] [PubMed]
- Eu, Y.J.; Park, H.S.; Kim, D.P.; Hong, J.W. A microfluidic perfusion platform for cultivation and screening study of motile microalgal cells. Biomicrofluidics 2014, 8, 024113. [Google Scholar] [CrossRef] [PubMed]
- Luke, C.S.; Selimkhanov, J.; Baumgart, L.; Cohen, S.E.; Golden, S.S.; Cookson, N.A.; Hasty, J. A microfluidic platform for long-term monitoring of algae in dynamic environment. ACS Synth. Biol. 2016, 5, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Graham, P.J.; Riordon, J.; Sinton, D. Microalgae on display: A microfluidic pixel-based irradiance assay for photosynthetic growth. Lab Chip 2015, 15, 3116–3124. [Google Scholar] [CrossRef] [PubMed]
- Qu, B.; Eu, Y.J.; Jeong, W.J.; Kim, D.P. Droplet electroporation in microfluidics for efficient cell transformation with or without cell wall removal. Lab Chip 2012, 12, 4483–4488. [Google Scholar] [CrossRef] [PubMed]
- Abalde-Cela, S.; Gould, A.; Liu, X.; Kazamia, E.; Smit, A.G.; Abell, C. High-throughput detection of ethanol- producing cyanobacteria in a microdroplet platform. J. R. Soc. Interface 2015, 12, 20150216. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.K.; Guzman, A.R.; Thapa, H.R.; Devarenne, T.P.; Han, A. A droplet microfluidics platform for rapid microalgal growth and oil production analysis. Biotechnol. Bioeng. 2016, 113, 1691–1701. [Google Scholar] [CrossRef] [PubMed]
- Hammar, P.; Angermayr, S.A.; Sjostrom, S.L.; van der Meer, J.; Hellingwerf, K.J.; Hudson, E.P.; Joensson, H.W. Single-cell screening of photosynthetic growth and lactate production by cyanobacteria. Biotechnol. Biofuels 2015, 8, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Au, S.; Shih, S.C.; Wheeler, A.R. Integrated microbioreactor for culture and analysis of bacteria, algae, and yeast. Biomed. Microdevices 2011, 13, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Shih, S.C.C.; Mufti, N.S.; Chamberlain, M.D.; Kim, J.; Wheeler, A.R. A droplet-based screen for wavelength-dependent lipid production in alage. Energy Environ. Sci. 2014, 7, 2366–2375. [Google Scholar] [CrossRef]
- Jung, E.E.; Kalontarov, M.; Doud, D.F.R.; Ooms, M.D.; Angenent, L.T.; Sinton, D.; Erickson, D. Slab waveguide photobioreactors for microalgae based biofuel production. Lab Chip 2012, 12, 3740–3745. [Google Scholar] [CrossRef] [PubMed]
- Pierobon, S.; Ooms, M.; Sinton, D. Evanescent cultivation of photosynthetic bacteria on thin waveguides. J. Micromach. Microeng. 2014, 24, 045017. [Google Scholar] [CrossRef]
- Novotny, L.; Hecht, B. Principle of Nano Optics, 2nd ed.; Cambridge University Press: Cambridge, UK, 2012. [Google Scholar]
- Stockman, M.I. Nanoplasmonics: The physics behind the applications. Phys. Today 2011, 64, 39–44. [Google Scholar] [CrossRef]
- Ooms, M.D.; Bajin, L.; Sinton, D. Culturing photosynthetic bacteria through surface plasmon resonance. Appl. Phys. Lett. 2012, 101, 253701. [Google Scholar] [CrossRef]
- Ooms, M.; Jeyaram, Y.; Sinton, D. Wavelength-selective plasmonics for enhanced cultivation of microalgae. Appl. Phys. Lett. 2015, 106, 063902. [Google Scholar] [CrossRef]
- Samsonoff, N.; Ooms, M.D.; Sinton, D. A photosynthetic-plasmonic-voltaic cell: Excitation of photosynthetic bacteria and currentcollection through a plasmonic substrate. Appl. Phys. Lett. 2014, 104, 043704. [Google Scholar] [CrossRef]
- Ramey, C.J.; Barón-Sola, Ã.; Aucoin, H.R.; Boyle, N.R. Genome engineering in cyanobacteria: Where we are and where we need to go. ACS Synth. Biol. 2015, 4, 1186–1196. [Google Scholar] [CrossRef] [PubMed]
- Hillson, N.J.; Rosengarten, R.D.; Keasling, J.D. j5 DNA assembly design automation software. ACS Synth. Biol. 2012, 1, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.C.; Liu, Y.J.; Yao, D.J. Core—Shell droplets for parallel DNA ligation of an ultra-micro volume using an EWOD microfluidic system. J. Lab. Autom. 2010, 15, 210–215. [Google Scholar] [CrossRef]
- Linshiz, G.; Stawski, N.; Goyal, G.; Bi, C.; Poust, S.; Sharma, M.; Mutalik, V.; Keasling, J.D.; Hillson, N.J. PR-PR: Cross-platform laboratory automation system. ACS Synth. Biol. 2014, 3, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Shih, S.C.C.; Goyal, G.; Kim, P.W.; Koutsoubelis, N.; Keasling, J.D.; Adams, P.D.; Hillson, N.J.; Singh, A.K. A versatile microfluidic device for automating synthetic biology. ACS Synth. Biol. 2015, 4, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.J.; Tabor, J.J. Optogenetic characterization methods overcome key challenges in synthetic and systems biology. Nat. Chem. Biol. 2014, 10, 502–511. [Google Scholar] [CrossRef] [PubMed]
- Levskaya, A.; Chevalier, A.A.; Tabor, J.J.; Simpson, Z.B.; Lavery, L.A.; Levy, M.; Davidson, E.A.; Scouras, A.; Andrew, D.; Ellington, A.D.; et al. Synthetic biology: Engineering Escherichia coli to see light. Nature 2005, 438, 441–442. [Google Scholar] [CrossRef] [PubMed]
- Tabor, J.J.; Levskaya, A.; Voigt, C.A. Multichromatic control of gene expression in Escherichia coli. J. Mol. Biol. 2011, 405, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Schmidl, S.R.; Sheth, R.U.; Wu, A.; Tabor, J.J. Refactoring and optimization of light-switchable Escherichia coli two-component system. ACS Synth. Biol. 2014, 3, 820–831. [Google Scholar] [CrossRef] [PubMed]
- Ohlendorf, R.; Vidavski, R.R.; Eldar, A.; Moffat, K.; Möglich, A. From dusk till dawn: One-plasmid systems for light regulated gene expression. J. Mol. Biol. 2012, 416, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Möglich, A.; Ayers, R.A.; Moffat, K. Design and signaling mechanism of light-regulated histidine kinases. J. Mol. Biol. 2009, 385, 1433–1444. [Google Scholar] [CrossRef] [PubMed]
- Olson, E.J.; Hartsough, L.A.; Landry, B.P.; Schroff, R.; Tabor, J.J. Characterizing bacterial gene circuit dynamics with optically programmed gene expression signals. Nat. Methods 2014, 11, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Link, H.; Fuherer, T.; Gerosa, L.; Zamboni, N.; Sauer, U. Real-time metabolome profiling of the metabolic switch between starvation and growth. Nat. Methods 2015, 12, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Young, J.D.; Shastri, A.A.; Stephanopoulos, G.; Morgan, J.A. Mapping photoaurotrophic metabolism with isotropically nonstationary 13C flux analysis. Metab. Eng. 2011, 13, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Bonnett, B.D.; Kimball, E.H.; Gao, M.; Osterhout, R.; Van Dien, S.J.; Rabinowitz, J.D. Absolute metabolite concentrations and implied enzyme active site occupancy in Eschricha coli. Nat. Chem. Biol. 2009, 8, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ren, L.; Zhang, X.; Shan, Y.; Wang, Y.; Ji, Y.; Yin, H.; Huang, W.E.; Xu, J.; Ma, B. Raman activated cell sorting based on dielectrophoretic single-cell trap and release. Anal. Chem. 2015, 87, 2282–2289. [Google Scholar] [CrossRef] [PubMed]
- Ji, Y.; He, Y.; Cui, Y.; Wang, T.; Wang, Y.; Huang, W.E.; Xu, J. Raman spectroscopy provides a rapid, non-invasive method for quantitation of starch in live, unicellular microalgae. Biotechnol. J. 2014, 9, 1512–1518. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.; Kim, C.W.; Choi, J.S.; Yang, J.W.; Seo, T.S. An integrated microfluidic device for the high-throuput screening of microalgal cell culture conditions that induces high growth rate and lipid content. Anal. Bioanal. Chem. 2013, 405, 9365–9374. [Google Scholar] [CrossRef] [PubMed]
- Kiviet, D.J.; Nghe, P.; Walker, N.; Boulineau, S.; Sunderlikova, V.; Tans, S.J. Stochasticity of metabolism and growth at the single cell level. Nature 2014, 514, 376–389. [Google Scholar] [CrossRef] [PubMed]
- Ackerman, M. A functional perspective on phenotypic heterogeneity in microorganisms. Nat. Rev. Microbiol. 2015, 13, 497–508. [Google Scholar] [CrossRef] [PubMed]
- Bar-Even, A.; Noor, E.; Lewis, N.E.; Milo, R. Design and analysis of synthetic carbon fixation pathway. Proc. Natl. Acad. Sci. USA 2009, 107, 8889–8894. [Google Scholar] [CrossRef] [PubMed]
- Siegel, J.B.; Smith, A.L.; Poust, S.; Wargacki, A.J.; Bar-Even, A.; Louw, C.; Shen, B.W.; Eiben, C.B.; Tran, H.M.; Noor, E.; et al. Computational protein design enables a novel one-carbon assimilation pathway. Proc. Natl. Acad. Sci. USA 2015, 112, 3704–3709. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Work Flow | Challenges | Microfluidic Technology | References |
|---|---|---|---|
| Gene assembly | Fast, accurate, construction of large genetic devices | Microfluidics DNA synthesis and assembly with integrated bacterial transformation | [60,62] |
| Verification | Large scale screening Dynamic chemical environment | Microfluidic photobioreactor supporting long term cell growth and single cell monitoring | [41,42,43] |
| End production titration | Low cost, in situ measurement to replace current technology | 1. Enzyme assay with fluorescent detection 2. Spectroscopy such as Raman integrated with microfluidics | [46,48,73,74] |
© 2016 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.-T.; Wang, C.Y. Review of Microfluidic Photobioreactor Technology for Metabolic Engineering and Synthetic Biology of Cyanobacteria and Microalgae. Micromachines 2016, 7, 185. https://doi.org/10.3390/mi7100185
Yang Y-T, Wang CY. Review of Microfluidic Photobioreactor Technology for Metabolic Engineering and Synthetic Biology of Cyanobacteria and Microalgae. Micromachines. 2016; 7(10):185. https://doi.org/10.3390/mi7100185
Chicago/Turabian StyleYang, Ya-Tang, and Chun Ying Wang. 2016. "Review of Microfluidic Photobioreactor Technology for Metabolic Engineering and Synthetic Biology of Cyanobacteria and Microalgae" Micromachines 7, no. 10: 185. https://doi.org/10.3390/mi7100185