Clostridium perfringens Epsilon Toxin: A Malevolent Molecule for Animals and Man?



Abstract
:1. Introduction
2. Clostridium perfringens Toxins: Major and Minor (A Brief Overview)

{kind=link}
| Toxin | C. perfringens Type | Cellular Target (mode of action) | ||||
|---|---|---|---|---|---|---|
| A | B | C | D | E | ||
| Alpha | + | + | + | + | + | Membrane (phospholipid destruction) |
| Beta | + | + | Membrane (pore formation) | |||
| Epsilon | + | + | Membrane (pore formation) | |||
| Iota | + | Actin (cytoskeleton destruction) | ||||
| Toxin Type | Disease |
|---|---|
| A | Myonecrosis (gas gangrene in humans and animals); Necrotic enteritis of fowl plus piglets; Human food poisoning and antibiotic-associated diarrhea |
| B | Hemorrhagic enteritis in calves, foals and sheep; Dysentery in lambs |
| C | Necrotizing enteritis in humans (also popularly called pigbel, darmbrand or fire-belly), as well as in pigs, calves, goats and foals; Enterotoxemia in sheep (alias struck) |
| D | Enterotoxemia in lambs (known as pulpy kidney disease), goats and cattle |
| E | Enterotoxemia in calves and lambs. Similar enteric disease induced by iota-like toxin in rabbits, caused by Clostridium spiroforme |
2.1. Alpha Toxin
2.2. Beta Toxin
2.3. Iota Toxin
| Toxin/Enzyme | Activity |
|---|---|
| Beta 2 | ? |
| Delta | Cytolysin |
| Eta | ? |
| Gamma | ? |
| Kappa | Collagenase |
| Lambda | Protease |
| Mu | Hyaluronidase |
| Nu | Deoxyribonuclease |
| NanI, NanJ, NanH | Neuraminidase |
| NetB | Hemolysin |
| Theta (perfringolysin O) | Oxygen-labile Hemolysin |
| TpeL | Glucosylation of Ras |
3. Epsilon Toxin (Etx): C. perfringens Most Toxic Toxin
3.1. Natural Occurrence and a Potential Biological Warfare/Terrorism Agent?
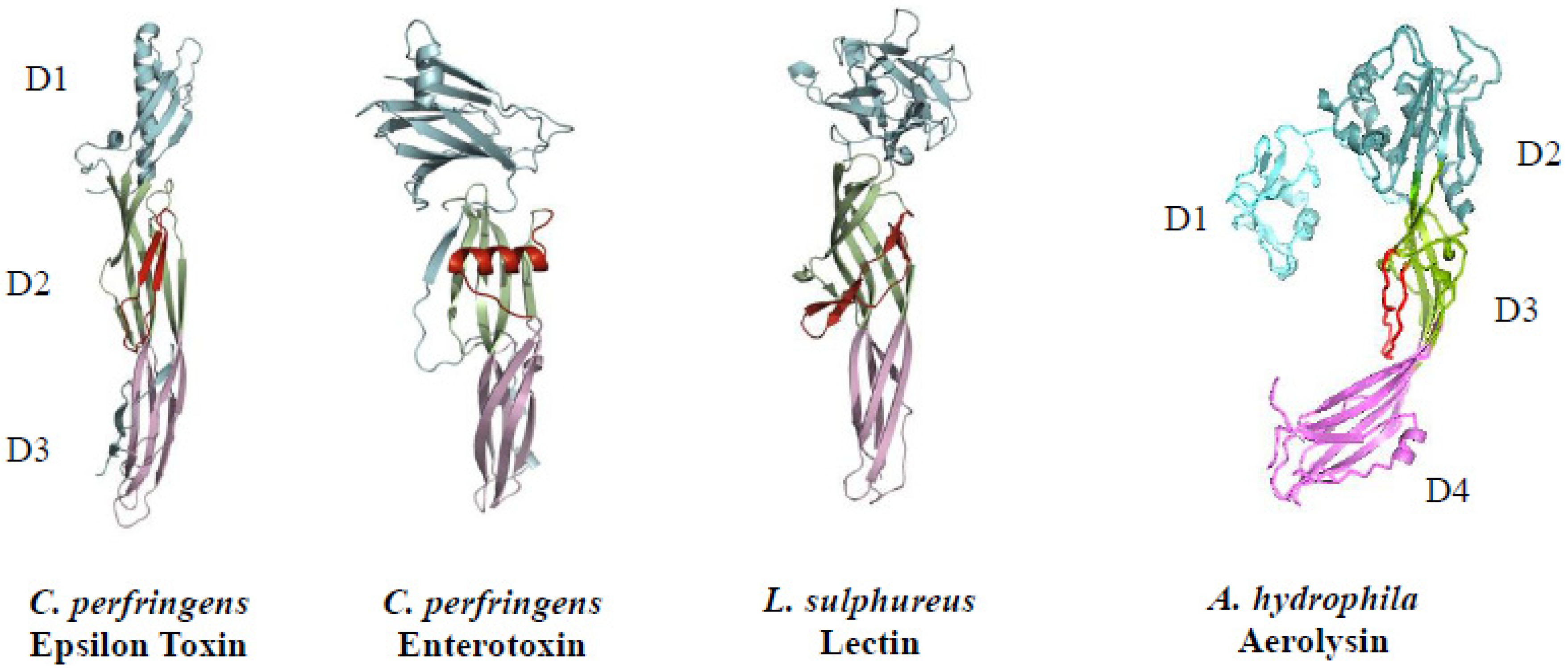
3.2. Chemical and Physical Properties
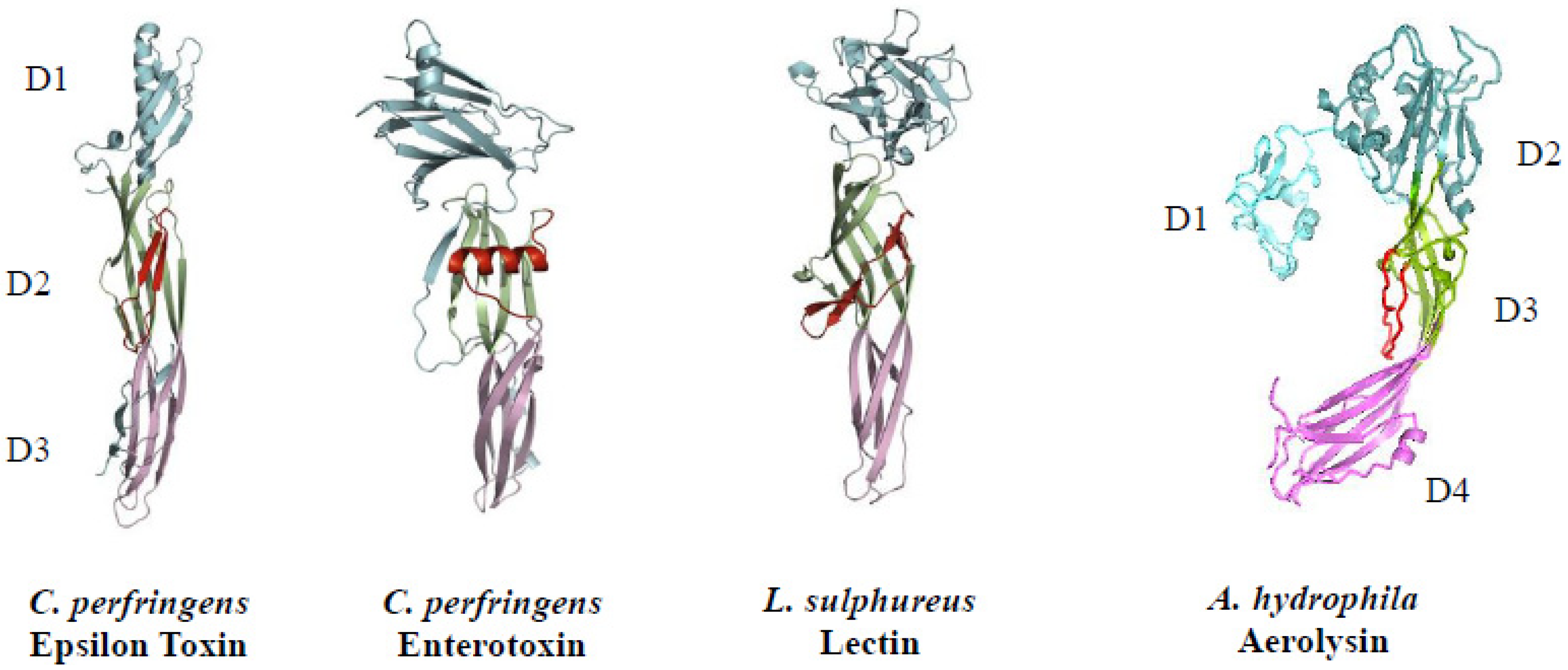
3.3. Cellular Target (Mode of Action)
4. A Veterinary Perspective on Etx: Field and Laboratory Findings
4.1. Small Animal Models
4.2. Detection
5. Management of Etx Intoxication: Therapy and Prophylaxis
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Songer, J.G. Clostridial enteric diseases of domestic animals. Clin. Microbiol. Rev. 1996, 9, 216–234. [Google Scholar]
- McDonel, J.L. Clostridium perfringens toxins (Type A, B, C, D, E). Pharmacol. Ther. 1980, 10, 617–655. [Google Scholar] [CrossRef]
- Oakley, C.L.; Warrack, G.H. Routine typing of Clostridium welchii. J. Hyg. Camb. 1953, 51, 102–107. [Google Scholar] [CrossRef]
- Goldstein, M.R.; Kruth, S.A.; Bersenas, A.M.; Holowaychuk, M.K.; Weese, J.S. Detection and characterization of Clostridium perfringens in the feces of healthy and diarrheic dogs. Can. J. Vet. Res. 2012, 76, 161–165. [Google Scholar]
- Albini, S.; Brodard, I.; Jaussi, A.; Wollschlaeger, N.; Frey, J.; Miserez, R.; Abril, C. Real-time multiplex PCR assays for reliable detection of Clostridium perfringens toxin genes in animal isolates. Vet. Microbiol. 2008, 127, 179–185. [Google Scholar] [CrossRef]
- Layana, J.E.; Fernandez-Miyakawa, M.E.; Uzal, F.A. Evaluation of different fluids for detection of Clostridium perfringens type D epsilon toxin in sheep with experimental enterotoxemia. Anaerobe 2006, 12, 204–206. [Google Scholar] [CrossRef]
- Carman, R.J.; Stevens, A.L.; Lyerly, M.W.; Hiltonsmith, M.F.; Stiles, B.G.; Wilkins, T.D. Clostridium difficile binary toxin (CDT) and diarrhea. Anaerobe 2011, 17, 161–165. [Google Scholar] [CrossRef]
- Macias Rioseco, M.; Beingesser, J.; Uzal, F.A. Freezing or adding trypsin inhibitor to equine intestinal contents extends the lifespan of Clostridium perfringens beta toxin for diagnostic purposes. Anaerobe 2012, 18, 357–360. [Google Scholar] [CrossRef]
- Hale, M.L.; Stiles, B.G. Detection of Clostridium perfringens alpha toxin using a capture antibody ELISA. Toxicon 1999, 37, 471–484. [Google Scholar] [CrossRef]
- Seyer, A.; Fenaille, F.; Feraudet-Tarisse, C.; Volland, H.; Popoff, M.R.; Tabet, J.C.; Junot, C.; Becher, F. Rapid quantification of clostridial epsilon toxin in complex food and biological matrixes by immunopurification and ultraperformance liquid chromatography-tandem mass spectrometry. Anal. Chem. 2012, 84, 5103–5109. [Google Scholar] [CrossRef]
- Langley, F.H.; Winkelstein, L.B. Gas gangrene: A study of 96 cases treated in an evacuation hospital. JAMA 1945, 128, 783–792. [Google Scholar] [CrossRef]
- Bryant, A.E.; Stevens, D.L. The Pathogenesis of Gas Gangrene. In The Clostridia: Molecular Biology and Pathogenesis; Rood, J.I., McClane, B.A., Songer, J.G., Titball, R.W., Eds.; Academic Press: San Diego, CA, USA, 1997; Chapter 11; pp. 185–196. [Google Scholar]
- Smith, L.D.; Gardner, M.V. The occurrence of vegetative cells of Clostridium perfringens in soil. J. Bacteriol. 1949, 58, 407–408. [Google Scholar]
- MacLennan, J.D.; MacFarlane, M.G. The treatment of gas gangrene. Br. Med. J. 1944, 1, 683–685. [Google Scholar] [CrossRef]
- Smith, L.D. Clostridia in gas gangrene. Bact. Rev. 1949, 13, 233–254. [Google Scholar]
- Evans, D.G.; Perkins, F.T. Fifth international standard for gas-gangrene antitoxin (perfringens) (Clostridium welchii type A antitoxin). Bull. World Health Organ. 1963, 29, 729–735. [Google Scholar]
- Williamson, E.D.; Titball, R.W. A genetically engineered vaccine against the alpha-toxin of Clostridium perfringens protects mice against experimental gas gangrene. Vaccine 1993, 11, 1253–1258. [Google Scholar] [CrossRef]
- Stevens, D.L.; Titball, R.W.; Jepson, M.; Bayer, C.R.; Hayes-Schroer, S.M.; Bryant, A.E. Immunization with the C-domain of alpha-toxin prevents lethal infection, localizes tissue injury, and promotes host response to challenge with Clostridium perfringens. J. Infect. Dis. 2004, 190, 767–773. [Google Scholar] [CrossRef]
- Naylor, C.E.; Eaton, J.T.; Howells, A.; Justin, N.; Moss, D.S.; Titball, R.W.; Basak, A.K. Structure of the key toxin in gas gangrene. Nat. Struct. Biol. 1998, 5, 738–746. [Google Scholar] [CrossRef]
- Sakurai, J.; Nagahama, M.; Oda, M. Clostridium perfringens alpha-toxin: Characterization and mode of action. J. Biochem. 2004, 136, 569–574. [Google Scholar] [CrossRef]
- MacFarlane, M.G.; Knight, B.C. The biochemistry of bacterial toxins. The lecithinase activity of Cl. welchii toxins. Biochem. J. 1941, 35, 884–902. [Google Scholar]
- Oda, M.; Kabura, M.; Takagishi, T.; Suzue, A.; Tominaga, K.; Urano, S.; Nagahama, M.; Kobayashi, K.; Furukawa, K.; Furukawa, K.; et al. Clostridium perfringens alpha-toxin recognizes the GM1a/TrkA complex. J. Biol. Chem. 2012, 287, 33070–33079. [Google Scholar] [CrossRef]
- Hunter, S.E.C.; Brown, J.E.; Oyston, P.C.F.; Sakurai, J.; Titball, R.W. Molecular genetic analysis of beta-toxin of Clostridium perfringens reveals sequence homology with alpha-toxin, gamma-toxin, and leukocidin of Staphylococcus aureus. Infect. Immun. 1993, 61, 3958–3965. [Google Scholar]
- Sakurai, J.; Duncan, C.L. Purification of beta-toxin from Clostridium perfringens type C. Infect. Immun. 1977, 18, 741–745. [Google Scholar]
- Walker, P.D.; Batty, I.; Egerton, J.R. The typing of Cl. perfringens and the veterinary background. Papua New Guinea Med. J. 1979, 22, 50–56. [Google Scholar]
- Shatursky, O.; Bayles, R.; Rogers, M.; Jost, B.H.; Songer, J.G.; Tweten, R.K. Clostridium perfringens beta-toxin forms potential-dependent, cation-selective channels in lipid bilayers. Infect. Immun. 2000, 68, 5546–5551. [Google Scholar] [CrossRef]
- Nagahama, M.; Morimitsu, S.; Kihara, A.; Akita, M.; Setsu, K.; Sakurai, J. Involvement of tachykinin receptors in Clostridium perfringens beta-toxin-induced plasma extravasation. Br. J. Pharmacol. 2003, 138, 23–30. [Google Scholar] [CrossRef]
- Bosworth, T. On a new type of toxin produced by Clostridium welchii. J. Comp. Path. 1943, 53, 245–255. [Google Scholar] [CrossRef]
- Stiles, B.G.; Wigelsworth, D.J.; Popoff, M.R.; Barth, H. Clostridial binary toxins: Iota and C2 family portraits. Front. Cell. Infect. Microbiol. 2011. [Google Scholar] [CrossRef]
- Briggs, D.C.; Naylor, C.E.; Smedley, J.G.; Lukoyanova, N.; Robertson, S.; Moss, D.S.; McClane, B.A.; Basak, A.K. Structure of the food-poisoning Clostridium perfringens enterotoxin reveals similarity to the aerolysin-like pore-forming toxins. J. Mol. Biol. 2011, 413, 138–149. [Google Scholar] [CrossRef]
- Popoff, M.R. Epsilon toxin: A fascinating pore-forming toxin. FEBS J. 2011, 278, 4602–4615. [Google Scholar] [CrossRef]
- Huebner, K.D.; Wannemacher, R.W.; Stiles, B.G.; Popoff, M.R.; Poli, M.A. Textbook of Military Medicine: Medical Aspects of Biological Warfare; Dembek, Z.F., Ed.; Office of The Surgeon General, Borden Institute: Washington, DC, USA, 2007; Chapter 17; p. 355. [Google Scholar]
- HHS and USDA Select Agents and Toxins. 7 CFR Part 331, 9 CFR Part 121, and 42 CFR Part 73. Available online: http://www.selectagents.gov/resources/List_of_Select_Agents_and_Toxins_2012-12-4-English.pdf (accessed on 1 October 2013).
- Cole, A.R.; Gibert, M.; Popoff, M.R.; Moss, D.S.; Titball, R.W.; Basak, A.K. Clostridium perfringens epsilon-toxin shows structural similarity to the pore-forming toxin aerolysin. Nat. Struct. Mol. Biol. 2004, 11, 797–798. [Google Scholar] [CrossRef]
- Knapp, O.; Stiles, B.G.; Popoff, M.R. The aerolysin-like toxin family of cytolytic, pore-forming toxins. Open Toxinol. J. 2010, 3, 53–68. [Google Scholar] [CrossRef]
- Mancheno, J.M.; Tateno, H.; Goldstein, I.J.; Martinez-Ripoll, M.; Hermoso, J.A. Structural analysis of the Laetiporus sulphureus hemolytic pore-forming lectin in complex with sugars. J. Biol. Chem. 2005, 280, 17251–17259. [Google Scholar] [CrossRef]
- Parker, M.W.; Buckley, J.T.; Postma, J.P.; Tucker, A.D.; Leonard, K.; Pattus, F.; Tsernoglou, D. Structure of the Aeromonas toxin proaerolysin in its water-soluble and membrane-channel states. Nature 1994, 367, 292–295. [Google Scholar] [CrossRef]
- Miyata, S.; Matsushita, O.; Minami, J.; Katayama, S.; Shimamoto, S.; Okabe, A. Cleavage of a C-terminal peptide is essential for heptamerization of Clostridium perfringens epsilon-toxin in the synaptosomal membrane. J. Biol. Chem. 2001, 276, 13778–13783. [Google Scholar]
- Petit, L.; Gibert, M.; Henri, C.; Lorin, V.; Baraige, F.; Carlier, J.P.; Popoff, M.R. Molecular basis of the activity of Clostridium perfringens toxins. Curr. Topics Biochem. Res. 1999, 1, 19–35. [Google Scholar]
- Jin, F.; Matsushita, O.; Katayama, S.; Jin, S.; Matsushita, C.; Minami, J.; Okabe, A. Purification, characterization, and primary structure of Clostridium perfringens lambda-toxin, a thermolysin-like metalloprotease. Infect. Immun. 1996, 64, 230–237. [Google Scholar]
- Harkness, J.M.; Li, J.; McClane, B.A. Identification of a lambda toxin-negative Clostridium perfringens strain that processes and activates epsilon prototoxin intracellularly. Anaerobe 2012, 18, 546–552. [Google Scholar] [CrossRef]
- Petit, L.; Gibert, M.; Gourch, A.; Bens, M.; Vandewalle, A.; Popoff, M.R. Clostridium perfringens epsilon toxin rapidly decreases membrane barrier permeability of polarized MDCK cells. Cell. Microbiol. 2003, 5, 155–164. [Google Scholar] [CrossRef]
- Miyata, S.; Minami, J.; Tamai, E.; Matsushita, O.; Shimamoto, S.; Okabe, A. Clostridium perfringens epsilon-toxin forms a heptameric pore within the detergent-insoluble microdomains of Madin-Darby canine kidney cells and rat synaptosomes. J. Biol. Chem. 2002, 277, 39463–39468. [Google Scholar]
- Lafont, F.; Abrami, L.; van der Goot, F.G. Bacterial subversion of lipid rafts. Curr. Opin. Microbiol. 2004, 7, 4–10. [Google Scholar]
- Fennessey, C.M.; Sheng, J.; Rubin, D.H.; McClain, M.S. Oligomerization of Clostridium perfringens epsilon toxin is dependent upon caveolins 1 and 2. PLoS One 2012, 7, e46866. [Google Scholar]
- Petit, L.; Gibert, M.; Gillet, D.; Laurent-Winter, C.; Boquet, P.; Popoff, M.R. Clostridium perfringens epsilon-toxin acts on MDCK cells by forming a large membrane complex. J. Bacteriol. 1997, 179, 6480–6487. [Google Scholar]
- Nagahama, M.; Itohayashi, Y.; Hara, H.; Higashihara, M.; Fukatani, Y.; Takagishi, T.; Oda, M.; Kobayashi, K.; Nakagawa, I.; Sakurai, J. Cellular vacuolation induced by Clostridium perfringens epsilon-toxin. FEBS J. 2011, 278, 3395–3407. [Google Scholar] [CrossRef]
- Li, J.; Sayeed, S.; Robertson, S.; Chen, J.; McClane, B.A. Sialidases affect the host cell adherence and epsilon toxin-induced cytotoxicity of Clostridium perfringens type D strain CD3718. PLoS Pathog. 2011, 7, e1002429. [Google Scholar] [CrossRef] [Green Version]
- Galen, J.E.; Ketley, J.M.; Fasano, A.; Richardson, S.H.; Wasserman, S.S.; Kaper, J.B. Role of Vibrio cholerae neuraminidase in the function of cholera toxin. Infect. Immun. 1992, 60, 406–415. [Google Scholar]
- Petit, L.; Maier, E.; Gibert, M.; Popoff, M.R.; Benz, R. Clostridium perfringens epsilon toxin induces a rapid change of cell membrane permeability to ions and forms channels in artificial lipid bilayers. J. Biol. Chem. 2001, 276, 15736–15740. [Google Scholar]
- Donelli, G.; Fiorentini, C.; Matarrese, P.; Falzano, L.; Cardines, R.; Mastrantonio, P.; Payne, D.W.; Titball, R.W. Evidence for cytoskeletal changes secondary to plasma membrane functional alterations in the in vitro cell response to Clostridium perfringens epsilon-toxin. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 145–156. [Google Scholar] [CrossRef]
- Zhu, C.; Ghabriel, M.N.; Blumbergs, P.C.; Reilly, P.L.; Manavis, J.; Youssef, J.; Hatami, S.; Finnie, J.W. Clostridium perfringens prototoxin-induced alteration of endothelial barrier antigen (EBA) immunoreactivity at the blood-brain barrier (BBB). Exp. Neurol. 2001, 169, 72–82. [Google Scholar] [CrossRef]
- Finnie, J.W.; Hajduk, P. An immunohistochemical study of plasma albumin extravasation in the brain of mice after the administration of Clostridium perfringens type D epsilon toxin. Aust. Vet. J. 1992, 69, 261–262. [Google Scholar] [CrossRef]
- Uzal, F.A.; Songer, J.G. Diagnosis of Clostridium perfringens intestinal infections in sheep and goats. J. Vet. Diagn. Invest. 2008, 20, 253–265. [Google Scholar] [CrossRef]
- Uzal, F.A.; Pasini, I.; Olaechea, F.V.; Robles, C.A.; Elizondo, A. An outbreak of enterotoxaemia caused by Clostridium perfringens type D in goats in Patagonia. Vet. Rec. 1994, 135, 279–280. [Google Scholar]
- Nagahama, M.; Sakurai, J. Distribution of labeled Clostridium perfringens epsilon toxin in mice. Toxicon 1991, 29, 211–217. [Google Scholar] [CrossRef]
- Soler-Jover, A.; Blasi, J.; Gomez de Aranda, I.; Navarro, P.; Gibert, M.; Popoff, M.R.; Martin-Satue, M. Effect of epsilon toxin-GFP on MDCK cells and renal tubules in vivo. J. Histochem. Cytochem. 2004, 52, 931–942. [Google Scholar] [CrossRef]
- Tamai, E.; Ishida, T.; Miyata, S.; Matsushita, O.; Suda, H.; Kobayashi, S.; Sonobe, H.; Okabe, A. Accumulation of Clostridium perfringens epsilon-toxin in the mouse kidney and its possible biological significance. Infect. Immun. 2003, 71, 5371–5375. [Google Scholar] [CrossRef]
- Gardner, D.E. Pathology of Clostridium welchii type D enterotoxaemia. 3. Basis of the hyperglycaemic response. J. Comp. Pathol. 1973, 83, 525–529. [Google Scholar] [CrossRef]
- Wioland, L.; Dupont, J.L.; Bossu, J.L.; Popoff, M.R.; Poulain, B. Attack of the nervous system by Clostridium perfringens epsilon toxin: From disease to mode of action on neural cells. Toxicon 2013, 75, 122–135. [Google Scholar] [CrossRef]
- Finnie, J.W. Pathogenesis of brain damage produced in sheep by Clostridium perfringens type D epsilon toxin: A review. Aust. Vet. J. 2003, 81, 219–221. [Google Scholar] [CrossRef]
- Finnie, J.W.; Manavis, J.; Blumbergs, P.C. Aquaporin-4 in acute cerebral edema produced by Clostridium perfringens type D epsilon toxin. Vet. Pathol. 2008, 45, 307–309. [Google Scholar] [CrossRef]
- Verkman, A.S. Aquaporins in clinical medicine. Annu. Rev. Med. 2012, 63, 303–316. [Google Scholar] [CrossRef]
- Uzal, F.A.; Kelly, W.R. Effects of the intravenous administration of Clostridium perfringens type D epsilon toxin on young goats and lambs. J. Comp. Pathol. 1997, 116, 63–71. [Google Scholar] [CrossRef]
- Uzal, F.A.; Kelly, W.R.; Morris, W.E.; Assis, R.A. Effects of intravenous injection of Clostridium perfringens type D epsilon toxin in calves. J. Comp. Pathol. 2002, 126, 71–75. [Google Scholar] [CrossRef]
- Uzal, F.A.; Kelly, W.R. Experimental Clostridium perfringens type D enterotoxemia in goats. Vet. Pathol. 1998, 35, 132–140. [Google Scholar] [CrossRef]
- Uzal, F.A.; Kelly, W.R.; Morris, W.E.; Bermudez, J.; Baison, M. The pathology of peracute experimental Clostridium perfringens type D enterotoxemia in sheep. J. Vet. Diagn. Invest 2004, 16, 403–411. [Google Scholar] [CrossRef]
- Finnie, J.W. Neurological disorders produced by Clostridium perfringens type D epsilon toxin. Anaerobe 2004, 10, 145–150. [Google Scholar] [CrossRef]
- Fernandez-Miyakawa, M.E.; Uzal, F.A. The early effects of Clostridium perfringens type D epsilon toxin in ligated intestinal loops of goats and sheep. Vet. Res. Commum. 2003, 27, 231–241. [Google Scholar] [CrossRef]
- Miyamoto, O.; Sumitani, K.; Nakamura, T.; Yamagami, S.; Miyata, S.; Itano, T.; Negi, T.; Okabe, A. Clostridium perfringens epsilon-toxin causes excessive release of glutamate in the mouse hippocampus. FEMS Microbiol. Lett. 2000, 189, 109–113. [Google Scholar] [CrossRef]
- Finnie, J.W. Ultrastructural changes in the brain of mice given Clostridium perfringens type D epsilon toxin. J. Comp. Path. 1984, 94, 445–452. [Google Scholar] [CrossRef]
- Sayeed, S.; Fernandez-Miyakawa, M.E.; Fisher, D.J.; Adams, V.; Poon, R.; Rood, J.I.; Uzal, F.A.; McClane, B.A. Epsilon-toxin is required for most Clostridium perfringens type D vegetative culture supernatants to cause lethality in the mouse intravenous injection model. Infect. Immun. 2005, 73, 7413–7421. [Google Scholar] [CrossRef]
- Garcia, J.P.; Adams, V.; Beingesser, J.; Hughes, M.L.; Poon, R.; Lyras, D.; Hill, A.; McClane, B.A.; Rood, J.I.; Uzal, F.A. Epsilon toxin is essential for the virulence of Clostridium perfringens type D infection in sheep, goats and mice. Infect. Immun. 2013, 81, 2405–2414. [Google Scholar] [CrossRef]
- Goldstein, J.; Morris, W.E.; Loidl, C.F.; Tironi-Farinatti, C.; McClane, B.A.; Uzal, F.A.; Fernandez-Miyakawa, M.E. Clostridium perfringens epsilon toxin increases the small intestinal permeability in mice and rats. PLoS One 2009, 4, e7065. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Miyakawa, M.E.; Sayeed, S.; Fisher, D.J.; Poon, R.; Adams, V.; Rood, J.I.; McClane, B.A.; Saputo, J.; Uzal, F.A. Development and application of an oral challenge mouse model for studying Clostridium perfringens type D infection. Infect. Immun. 2007, 75, 4282–4288. [Google Scholar] [CrossRef]
- Uzal, F.A.; Kelly, W.R.; Thomas, R.; Hornitzky, M.; Galea, F. Comparison of four techniques for the detection of Clostridium perfringens type D epsilon toxin in intestinal contents and other body fluids of sheep and goats. J. Vet. Diagn. Invest. 2003, 15, 94–99. [Google Scholar] [CrossRef]
- Payne, D.W.; Williamson, E.D.; Havard, H.; Modi, N.; Brown, J. Evaluation of a new cytotoxicity assay for Clostridium perfringens type D epsilon toxin. FEMS Microbiol. Lett. 1994, 116, 161–168. [Google Scholar] [CrossRef]
- Fernandez-Miyakawa, M.E.; Zabal, O.; Silberstein, C. Clostridium perfringens epsilon toxin is cytotoxic for human renal tubular epithelial cells. Hum. Exp. Toxicol. 2010, 30, 275–282. [Google Scholar] [CrossRef]
- Kumar, B.; Alam, S.I.; Kumar, O. Host response to intravenous injection of epsilon toxin in mouse model: A proteomic view. Proteomics 2012, 13, 89–107. [Google Scholar] [CrossRef]
- Ferry, T.; Thomas, D.; Perpoint, T.; Lina, G.; Monneret, G.; Mohammedi, I.; Chidiac, C.; Peyramond, D.; Vandenesch, F.; Etienne, J. Analysis of superantigenic toxin Vbeta T-cell signatures produced during cases of staphylococcal toxic shock syndrome and septic shock. Clin. Microbiol. Infect. 2008, 14, 546–554. [Google Scholar] [CrossRef]
- Lonchamp, E.; Dupont, J.L.; Wioland, L.; Courjaret, R.; Mbebi-Liegois, C.; Jover, E.; Doussau, F.; Popoff, M.R.; Bossu, J.-L.; Barry, J.; Poulain, B. Clostridium perfringens epsilon toxin targets granule cells in the mouse cerebellum and stimulates glutamate release. PLoS One 2011, 5, e13046. [Google Scholar]
- Miyamoto, O.; Minami, J.; Toyoshima, T.; Nakamura, T.; Masada, T.; Nagao, S.; Negi, T.; Itano, T.; Okabe, A. Neurotoxicity of Clostridium perfringens epsilon-toxin for the rat hippocampus via the glutamatergic system. Infect. Immun. 1998, 66, 2501–2508. [Google Scholar]
- Lewis, M.; Weaver, C.D.; McClain, M.S. Identification of small molecule inhibitors of Clostridium perfringens epsilon-toxin cytotoxicity using a cell-based high-throughput screen. Toxins 2010, 2, 1825–1847. [Google Scholar] [CrossRef]
- Aulinger, B.A.; Roehrl, M.H.; Mekalanos, J.J.; Collier, R.J.; Wang, J.Y. Combining anthrax vaccine and therapy: A dominant-negative inhibitor of anthrax toxin is also a potent and safe immunogen for vaccines. Infect. Immun. 2005, 73, 3408–3414. [Google Scholar] [CrossRef]
- Wai, S.N.; Westermark, M.; Oscarsson, J.; Jass, J.; Maier, E.; Benz, R.; Uhlin, B.E. Characterization of dominantly negative mutant ClyA cytotoxin proteins in Escherichia coli. J. Bacteriol. 2003, 185, 5491–5499. [Google Scholar]
- Genisset, C.; Galeotti, C.L.; Lupetti, P.; Mercati, D.; Skibinski, D.A.; Barone, S.; Battistutta, R.; de Bernard, M.; Telford, J.L. A Helicobacter pylori vacuolating toxin mutant that fails to oligomerize has a dominant negative phenotype. Infect. Immun. 2006, 74, 1786–1794. [Google Scholar] [CrossRef]
- Pelish, T.M.; McClain, M.S. Dominant-negative inhibitors of the Clostridium perfringens ε-toxin. J. Biol. Chem. 2009, 284, 29446–29453. [Google Scholar] [CrossRef]
- Buxton, D. Use of horseradish peroxidase to study the antagonism of Clostridium welchii (Cl. perfringens) type D epsilon toxin in mice by the formalinized epsilon protoxin. J. Comp. Pathol. 1976, 86, 67–72. [Google Scholar] [CrossRef]
- Dorca-Arevalo, J.; Martin-Satue, M.; Blasi, J. Characterization of the high affinity binding of epsilon toxin from Clostridium perfringens to the renal system. Vet. Microbiol. 2012, 157, 179–189. [Google Scholar] [CrossRef]
- Nagahama, M.; Sakurai, J. High-affinity binding of Clostridium perfringens epsilon-toxin to rat brain. Infect. Immun. 1992, 60, 1237–1240. [Google Scholar]
- Ivie, S.E.; Fennessey, C.M.; Sheng, J.; Rubin, D.H.; McClain, M.S. Gene-trap mutagenesis identifies mammalian genes contributing to intoxication by Clostridium perfringens epsilon-toxin. PLoS One 2011, 6, e17787. [Google Scholar]
- Ivie, S.E.; McClain, M.S. Identification of amino acids important for binding of Clostridium perfringens epsilon toxin to host cells and to HAVCR1. Biochemistry 2012, 51, 7588–7595. [Google Scholar] [CrossRef]
- Bokori-Brown, M.; Kokkinidou, M.D.; Savva, C.G.; Fernandes da Costa, S.P.; Naylor, C.E.; Cole, A.R.; Moss, D.S.; Basak, A.K.; Titball, R.W. Clostridium perfringens epsilon toxin H149A mutant as a platform for receptor binding studies. Protein Sci. 2013, 22, 650–659. [Google Scholar] [CrossRef] [Green Version]
- De la Rosa, C.; Hogue, D.E.; Thonney, M.L. Vaccination schedules to raise antibody concentrations against epsilon-toxin of Clostridium perfringens in ewes and their triplet lambs. J. Anim. Sci. 1997, 75, 2328–2334. [Google Scholar]
- Lobato, F.C.; Lima, C.G.; Assis, R.A.; Pires, P.S.; Silva, R.O.; Salvarani, F.M.; Carmo, A.O.; Contigli, C.; Kalapothakis, E. Potency against enterotoxemia of a recombinant Clostridium perfringens type D epsilon toxoid in ruminants. Vaccine 2010, 28, 6125–6127. [Google Scholar] [CrossRef]
- Uzal, F.A.; Wong, J.P.; Kelly, W.R.; Priest, J. Antibody response in goats vaccinated with liposome-adjuvanted Clostridium perfringens type D epsilon toxoid. Vet. Res. Commun. 1999, 23, 143–150. [Google Scholar] [CrossRef]
- Odendaal, M.W.; Visser, J.J.; Botha, W.J.; Prinsloo, H. The passive protection of lambs against Clostridium perfringens type D with semi-purified hyperimmune serum. Onderstepoort J. Vet. Res. 1988, 55, 47–50. [Google Scholar]
- Clarkson, M.J.; Faull, W.B.; Kerry, J.B. Vaccination of cows with clostridial antigens and passive transfer of clostridial antibodies from bovine colostrum to lambs. Vet. Rec. 1985, 116, 467–469. [Google Scholar]
- El-Enbaawy, M.I.; Abdalla, Y.A.; Hussein, A.Z.; Osman, R.M.; Selim, S.A. Production and evaluation of a monoclonal antibody to Clostridium perfringens type D epsilon toxin. Egypt. J. Immunol. 2003, 10, 77–81. [Google Scholar]
- McClain, M.S.; Cover, T.L. Functional analysis of neutralizing antibodies against Clostridium perfringens epsilon-toxin. Infect. Immun. 2007, 75, 1785–1793. [Google Scholar] [CrossRef]
- Walker, P.D. Bacterial vaccines: Old and new, veterinary and medical. Vaccine 1992, 10, 977–990. [Google Scholar] [CrossRef]
- Titball, R.W. Clostridium perfringens vaccines. Vaccine 2009, 27, D44–D47. [Google Scholar] [CrossRef]
- Hunter, S.E.; Clarke, I.N.; Kelly, D.C.; Titball, R.W. Cloning and nucleotide sequencing of the Clostridium perfringens epsilon-toxin gene and its expression in Escherichia coli. Infect. Immun. 1992, 60, 102–110. [Google Scholar]
- Oyston, P.C.; Payne, D.W.; Havard, H.L.; Williamson, E.D.; Titball, R.W. Production of a non-toxic site-directed mutant of Clostridium perfringens epsilon toxin which induces protective immunity in mice. Microbiology 1998, 144, 333–341. [Google Scholar] [CrossRef]
- Kaushik, H.; Deshmukh, S.; Mathur, D.D.; Tiwar, A.; Garg, L.C. Recombinant expression of in silico identified B-cell epitope of epsilon toxin of Clostridium perfringens in translational fusion with a carrier protein. Bioinformation 2013, 9, 617–621. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Stiles, B.G.; Barth, G.; Barth, H.; Popoff, M.R. Clostridium perfringens Epsilon Toxin: A Malevolent Molecule for Animals and Man? Toxins 2013, 5, 2138-2160. https://doi.org/10.3390/toxins5112138
Stiles BG, Barth G, Barth H, Popoff MR. Clostridium perfringens Epsilon Toxin: A Malevolent Molecule for Animals and Man? Toxins. 2013; 5(11):2138-2160. https://doi.org/10.3390/toxins5112138
Chicago/Turabian StyleStiles, Bradley G., Gillian Barth, Holger Barth, and Michel R. Popoff. 2013. "Clostridium perfringens Epsilon Toxin: A Malevolent Molecule for Animals and Man?" Toxins 5, no. 11: 2138-2160. https://doi.org/10.3390/toxins5112138