How Should Staphylococcal Food Poisoning Outbreaks Be Characterized?

Abstract
:1. Coagulase-Positive Staphylococci and Staphylococcal Enterotoxins
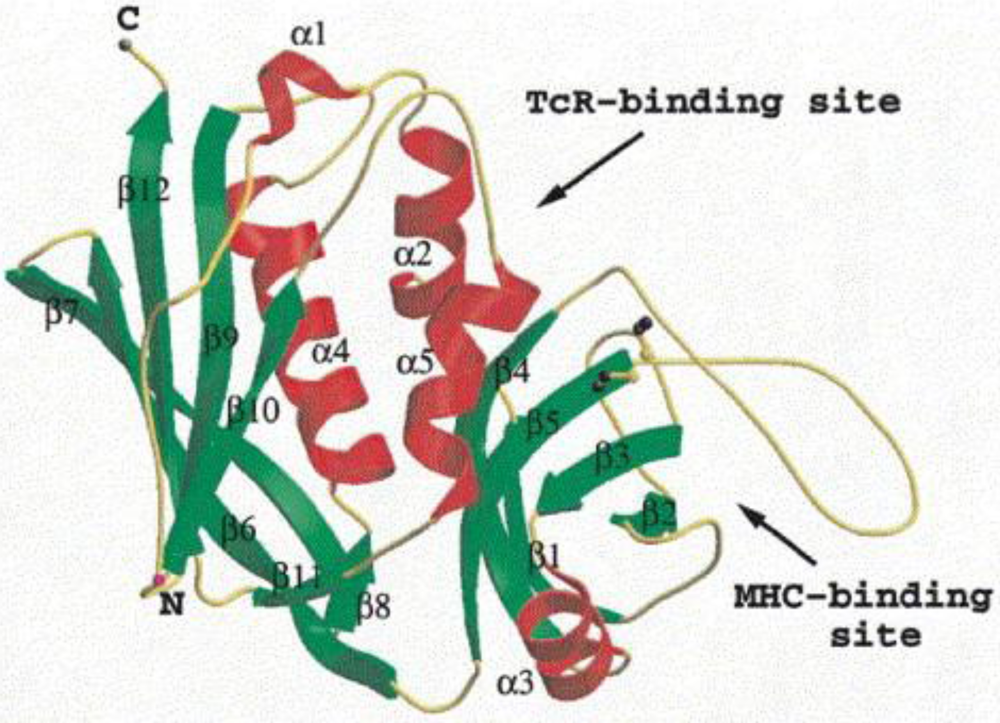
1.1. Coagulase-Positive Staphylococci

{kind=link}
| Species | Main sources | Ref. |
|---|---|---|
| S. aureus subsp. aureus | humans, animals | [1] |
| S. aureus subsp. anaerobius | sheep | [2] |
| S. intermedius | dog, horse, mink, pigeon | [3] |
| S. hyicus | pig, chicken | [4] |
| S. delphini | dolphin | [5] |
| S. schleiferi subsp. coagulans | dog (external ear) | [6] |
| S. lutrae | otter | [7] |
1.2. Staphylococcal Enterotoxins
| Toxin type | Molecular weight (Da) | Genetic basis of SE | Superantigenic action | Emetic action |
|---|---|---|---|---|
| SEA | 27,100 | Prophage | + | + |
| SEB | 28,336 | Chromosome, plasmid, pathogenicity island | + | + |
| SEC1-2-3 | ≈27,500 | Plasmid | + | + |
| SED | 26,360 | Plasmid (pIB485) | + | + |
| SEE | 26,425 | Prophage | + | + |
| SEG | 27,043 | enterotoxin gene cluster (egc), chromosome | + | + |
| SEH | 25,210 | Transposon | + | + |
| SEI | 24,928 | egc, chromosome | + | + |
| SElJ | 28,565 | Plasmid (pIB485) | + | nk |
| SEK | 25,539 | Pathogenicity island | + | nk |
| SElL | 24,593 | Pathogenicity island | + | − |
| SElM | 24,842 | egc, chromosome | + | nk |
| SElN | 26,067 | egc, chromosome | + | nk |
| SElO | 26,777 | egc, chromosome | + | nk |
| SElP | 26,608 | Prophage (Sa3n) | + | nk |
| SElQ | 25,076 | Pathogenicity island | + | − |
| SER | 27,049 | Plasmid (pIB485) | + | + |
| SES | 26,217 | Plasmid (pIB485) | + | + |
| SET | 22,614 | Plasmid (pIB485) | + | + |
| SElU | 27,192 | egc, chromosome | + | nk |
| SElU2 | 26,672 | egc, chromosome | + | nk |
| SElV | 24,997 | egc, chromosome | + | nk |
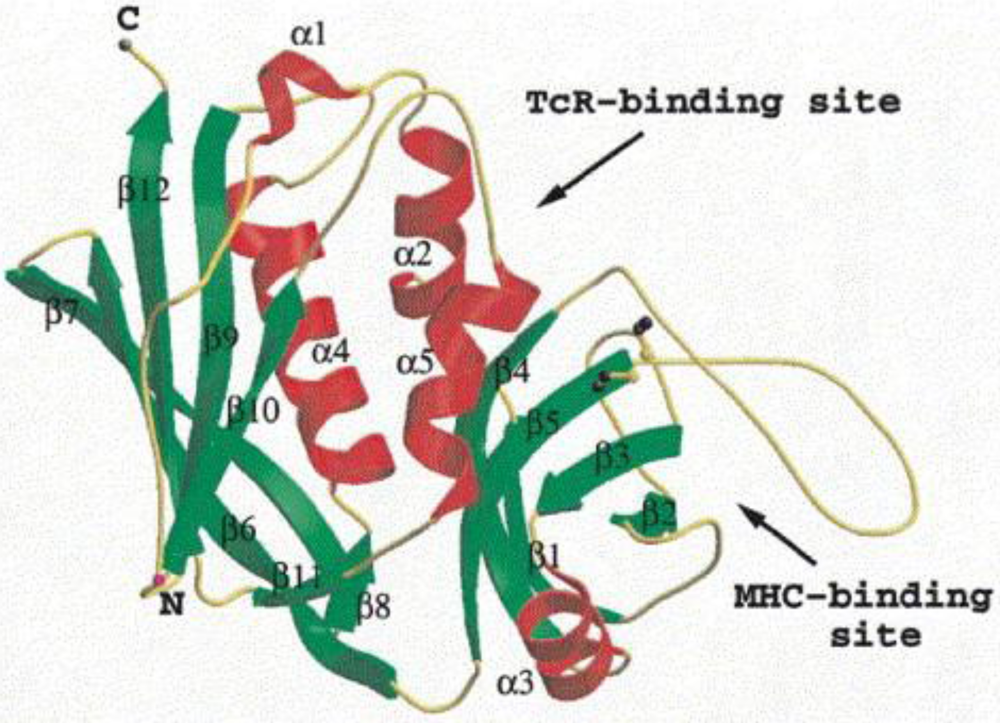
2. SFPOs: Definition and Required Conditions for Their Occurrence (European Data)
3. Analytical Tools Used in SFPO Characterization: Pros and Cons
3.1. Bioassays
3.2. Molecular Tools
3.3. Immunological Tools
3.4. Chromatographic Methods for the Detection and Quantification of SEs
4. An Integrated Approach to Improve SFPO Characterization
5. Conclusions
References
- Rosenbach, F.J. Microorganismen bei den Wund-Infections-Krankheiten des Menschen; Bergmann, J.F., Ed.; Wiesbaden, Germany, 1884; pp. 1–122. [Google Scholar]
- De la Fuente, R.; Suarez, G.; Schleifer, K.H. Staphylococcus aureus subsp. Anaerobius subsp. nov., the causal agent of abscess disease of sheep. Int. J. Syst. Bacteriol. 1985, 35, 99–102. [Google Scholar] [CrossRef]
- Hajek, V. Staphylococcus intermedius, a New Species Isolated from Animals. Int. J. Syst. Bacteriol. 1976, 26, 401–408. [Google Scholar]
- Devriese, L.A.; Hajek, K.; Oeding, P.; Meyer, S.A.; Schleifer, K.H. Staphylococcus hyicus (Sompolinsky 1953) comb. nov. and Staphylococcus hyicus subsp. chromogenes subsp. nov. Int. J. Syst. Bacteriol. 1978, 28, 482–490. [Google Scholar] [CrossRef]
- Varaldo, P.E.; Kilpper-Balz, R.; Biavasco, F.; Satta, G.; Scheifer, K.H. Staphylococcus delphini sp. nov., a coagulase-positive species isolated from dolphins. Int. J. Syst. Bacteriol. 1988, 38, 436–439. [Google Scholar] [CrossRef]
- Igimi, S.; Takahashi, E.; Mitsouka, T. Staphylococcus schleiferi subsp. coagulans subsp. nov., isolated from the external auditory meatus of dogs with external ear otitis. Int. J. Syst. Bacteriol. 1990, 40, 409–411. [Google Scholar] [CrossRef] [PubMed]
- Foster, G.; Ross, H.M.; Hutson, R.A.; Collins, M.D. Staphylococcus lutrae sp. nov., a new coagulase-positive species isolated from otters. Int. J. Syst. Bacteriol. 1997, 47, 724–726. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar]
- Orwin, P.M.; Leung, D.Y.; Donahue, H.L.; Novick, R.P.; Schlievert, P.M. Biochemical and Biological Properties of Staphylococcal Enterotoxin K. Infect. Immun. 2001, 69, 360–366. [Google Scholar]
- Jarraud, S.; Peyrat, M.A.; Lim, A.; Tristan, A.; Bes, M.; Mougel, C.; Etienne, J.; Vandenesch, F.; Bonneville, M.; Lina, G. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol. 2001, 166, 669–677. [Google Scholar]
- Omoe, K.; Imanishi, K.; Hu, D.L.; Kato, H.; Fugane, Y.; Abe, Y.; Hamaoka, S.; Watanabe, Y.; Nakane, A.; Uchiyama, T.; Shinagawa, K. Characterization of novel staphylococcal enterotoxin-like toxin type P. Infect. Immun. 2005, 73, 5540–5546. [Google Scholar]
- Orwin, P.M.; Leung, D.Y.M.; Tripp, T.J.; Bohach, G.A.; Earhart, C.A.; Ohlendorf, D.H.; Schlievert, P.M. Characterization of a novel staphylococcal enterotoxin-like superantigen, a member of the group V subfamily of pyrogenic toxins. Biochemistry 2002, 41, 14033–14040. [Google Scholar]
- Omoe, K.; Hu, D.L.; Takahashi-Omoe, H.; Nakane, A.; Shinagawa, K. Identification and characterization of a new staphylococcal enterotoxin-related putative toxin encoded by two kinds of plasmids. Infect. Immun. 2003, 71, 6088–6094. [Google Scholar]
- Ono, H.K.; Omoe, K.; Imanishi, K.; Iwakabe, Y.; Hu, D.L.; Kato, H.; Saito, N.; Nakane, A.; Uchiyama, T.; Shinagawa, K. Identification and characterization of two novel staphylococcal enterotoxins types S and T. Infect. Immun. 2008, 76, 4999–5005. [Google Scholar]
- Letertre, C.; Perelle, S.; Dilasser, F.; Fach, P. Identification of a new putative enterotoxin SEU encoded by the egc cluster of Staphylococcus aureus. J. Appl. Microbiol. 2003, 95, 38–43. [Google Scholar]
- Thomas, D.Y.; Jarraud, S.; Lemercier, B.; Cozon, G.; Echasserieau, K.; Etienne, J.; Gougeon, M.L.; Lina, G.; Vandenesch, F. Staphylococcal enterotoxin-like toxins U2 and V, two new staphylococcal superantigens arising from recombination within the enterotoxin gene cluster. Infect. Immun. 2006, 74, 4724–4734. [Google Scholar]
- Betley, M.J.; Mekalanos, J.J. Staphylococcal enterotoxin A is encoded by a phage. Science 1985, 229, 185–187. [Google Scholar]
- Bayles, K.W.; Iandolo, J.J. Genetic and molecular analysis of gene encoding staphylococcal enterotoxin D. J. Bacteriol. 1989, 171, 4799–4806. [Google Scholar]
- Yarwood, J.M; McCormick, J.K.; Paustian, M.L.; Orwin, P.M.; Kapur, V.; Schlievert, P.M. Characterization and expression analysis of Staphylococcus aureus pathogenicity island 3. Implications for the evolution of staphylococcal pathogenicity islands. Biol. Chem. 2002, 277, 13138–13147. [Google Scholar] [CrossRef]
- Papageorgiou, A.; Acharya, K. Microbial superantigens: from structure to function. Trends Microbiol. 2000, 8, 369–375. [Google Scholar]
- Dinges, M.M.; Orwin, P.M.; Schlievert, P.M. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev. 2000, 13, 16–34. [Google Scholar]
- Papageorgiou, A.C.; Tranter, H.S.; Acharya, H.K. Crystal structure of microbial superantigen staphylococcal enterotoxin B at 1.5 Å resolution: Implications for superantigen recognition by MHC class II molecules and T-cell receptors. J. Mol. Biol. 1998, 277, 61–79. [Google Scholar] [CrossRef] [PubMed]
- Jablonski, L.; Bohach, G.A. Staphylococcus aureus. In Food microbiology fundamentals and frontiers; Doyle, M.P., Beuchat, L.R., Montville, T.J., Eds.; America society of Microbiology Press: Washington, DC, USA, 2001; pp. 411–434. [Google Scholar]
- Asao, T.; Kumeda, Y.; Kawai, T.; Shibata, T.; Oda, H.; Haruki, K.; Nakazawa, H.; Kozaki, S. An extensive outbreak of staphylococcal food poisoning due to low-fat milk in Japan: Estimation of enterotoxin A in the incriminated milk and powdered skim milk. Epidemiol. Infect. 2003, 130, 33–40. [Google Scholar]
- Pereira, M.L.; Do Carmo, L.; Dos Santos, E.J.; Pereira, J.L.; Bergdoll, M.S. Enterotoxin H in staphylococcal food poisoning. J. Food Prot. 1996, 59, 559–561. [Google Scholar]
- The Community Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and food-borne outbreaks in the European Union in 2008. EFSA J. 2010, 1496. Available online: http://www.efsa.europa.eu/en/scdocs/doc/s1496.pdf (accessed on 6 August 2010).
- Bryan, F.L.; Guzewich, J.J.; Todd, E.C.D. Surveillance of foodborne disease II. Summary and presentation of descriptive data and epidemiologic patterns; their value and limitations. J. Food Prot. 1997, 60, 567–578. [Google Scholar]
- Surgalla, M.; Bergdoll, M.S.; Dack, G.M. Some observations on the assay of staphylococcal enterotoxin by the monkey feeding test. J. Lab. Clin. Med. 1953, 41, 782–788. [Google Scholar]
- Bergdoll, M.S. Staphylococcal intoxications. In Food-borne Infections and Intoxications; Riemann, H., Bryan, F.L., Eds.; Academic press: New York, NY, USA, 1979; pp. 443–494. [Google Scholar]
- Ikeda, T.; Tamate, N.; Yamaguchi, K.; Makino, S. Mass outbreak of food poisoning disease caused by small amounts of staphylococcal enterotoxins A and H. Appl. Environ. Microbiol. 2005, 71, 2793–2795. [Google Scholar]
- Hawryluk, T.; Hirshfield, I. A super antigen bioassay to detect staphylococcal enterotoxin A. J. Food Prot. 2002, 65, 1183–1187. [Google Scholar]
- Lee, Y.D.; Moon, B.Y.; Park, J.H.; Chang, H.I.; Kim, W.J. Expression of enterotoxin genes in Staphylococcus aureus isolates based on mRNA analysis. J. Microbiol. Biotechnol. 2007, 17, 461–467. [Google Scholar]
- Akineden, O.; Hassan, A.A.; Schneider, E.; Usleber, E. Enterotoxigenic properties of Staphylococcus aureus isolated from goats' milk cheese. Int. J. Food Microbiol. 2008, 124, 211–216. [Google Scholar]
- Derzelle, S.; Dilasser, F.; Duquenne, M.; Deperrois, V. Differential temporal expression of the staphylococcal enterotoxins genes during cell growth. Food Microbiol. 2009, 26, 896–904. [Google Scholar]
- Duquenne, M.; Fleurot, I.; Aigle, M.; Darrigo, C.; Borezée-Dupont, E.; Derzelle, S.; Bouix, M.; Deperrois-Lafarge, V.; Delacroix-Buchet, A. Tool for Quantification of Staphylococcal Enterotoxin Gene Expression in Cheese. Appl. Environ. Microbiol. 2010, 76, 1367–1374. [Google Scholar]
- Schlievert, P.M.; Case, L.C. Molecular analysis of staphylococcal superantigens. Methods Mol. Biol. 2007, 391, 113–126. [Google Scholar]
- Park, C.E.; Akhtar, M.; Rayman, M.K. Nonspecific reactions of a commercial enzyme-linked immunosorbent assay kit (TECRA) for detection of staphylococcal enterotoxins in foods. Appl. Environ. Microbiol. 1992, 58, 2509–2512. [Google Scholar] [PubMed]
- Wieneke, A.A. Comparison of four kits for the detection of staphylococcal enterotoxin in foods from outbreaks of food poisoning. Int. J. Food Microbiol. 1991, 14, 305–312. [Google Scholar]
- Macaluso, L.; Lapeyre, C.; Dragacci, S. Determination of influential factors during sample preparation for staphylococcal enterotoxin detection in dairy products. Analusis 1998, 26, 300–304. [Google Scholar]
- Meyrand, A.; Atrache, V.; Bavai, C.; Montet, M.P.; Vernozy-Rozand, C. Evaluation of an alternative extraction procedure for enterotoxin determination in dairy products. Lett. Appl. Microbiol. 1999, 28, 411–415. [Google Scholar]
- Lapeyre, C.; Maire, T.; Messio, S.; Dragacci, S. Enzyme immunoassay of staphylococcal enterotoxins in dairy products with cleanup and concentration by immunoaffinity column. J. AOAC Int. 2001, 84, 1587–1592. [Google Scholar]
- European Community. Commission Regulation. No. 1441/2007 of 5 December 2007. Off. J. Eur. Union. 2007, L322, 12–29.
- Bennett, R.W. Staphylococcal enterotoxin and its rapid identification in foods by enzyme-linked immunosorbent assay-based methodology. J. Food Prot. 2005, 68, 1264–1270. [Google Scholar]
- Hennekinne, J.A.; Ostyn, A.; Guillier, F.; Gohier, M.; Messio, S.; Dragacci, S.; Krys, S.; Lombard, B. Interlaboratory validation of the Vidas SET2 detection kit for an use in official controls of staphylococcal enterotoxins detection in milk products especially low-fat cheeses. J. AOAC Int. 2007, 90, 756–764. [Google Scholar]
- Lapeyre, C.; Janin, F.; Kaveri, S.V. Indirect double sandwich ELISA using monoclonal antibodies for detection of staphylococcal enterotoxins A, B, C1 and D in food samples. Food Microbiol. 1988, 5, 25–31. [Google Scholar]
- Mamone, G.; Picariello, S.; Caira, F.; Addeo, P. Analysis of food proteins and peptides by mass spectrometry based techniques. J. Chromatogr. A 2009, 1216, 7130–7142. [Google Scholar]
- Norrgran, J.; Williams, T.L.; Woolfitt, A.R.; Solano, M.I.; Pirkle, J.L.; Barr, J.R. Optimization of digestion parameters for protein quantification. Anal. Biochem. 2009, 393, 48–55. [Google Scholar]
- Oeljeklaus, S.; Meyer, H.E.; Warscheid, B. New dimensions in the study of protein complexes using quantitative mass spectrometry. FEBS Lett. 2009, 583, 1674–1683. [Google Scholar]
- Bernardo, K.; Pakulat, N.; Macht, M.; Krut, O.; Seifert, H.; Fleer, S.; Hunger, F.; Kronke, M. Identification and discrimination of Staphylococcus aureus strains using matrix-assisted laser desorption/ionization-time of flight mass spectrometry. Proteomics 2002, 2, 747–753. [Google Scholar]
- Callahan, J.H.; Shefcheck, K.J.; Williams, T.L.; Musser, S.M. Detection, confirmation, and quantification of staphylococcal enterotoxin B in food matrixes using liquid chromatography-mass spectrometry. Anal. Chem. 2006, 78, 1789–1800. [Google Scholar]
- Brun, V.; Dupuis, A.; Adrait, A.; Marcellin, M.; Thomas, D.; Court, M.; Vandenesch, F.; Garin, J. Isotope-labeled protein standards: toward absolute quantitative proteomics. Mol. Cell Proteomics 2007, 6, 2139–2149. [Google Scholar]
- Dupuis, A.; Hennekinne, J.A.; Garin, J.; Brun, V. Protein Standard Absolute Quantification (PSAQ) for improved investigation of staphylococcal food poisoning outbreaks. Proteomics 2008, 8, 4633–4636. [Google Scholar]
- Kerouanton, A.; Hennekinne, J.A.; Letertre, C.; Petit, L.; Chesneau, O.; Brisabois, A.; De Buyser, M.L. Characterization of Staphylococcus aureus strains associated with food poisoning outbreaks in France. Int. J. Food Microbiol. 2007, 115, 369–375. [Google Scholar]
- Ostyn, A.; De Buyser, M.L.; Guillier, F.; Groult, J.; Félix, B.; Salah, S.; Delmas, G.; Hennekinne, J.A. First evidence of a food poisoning outbreak due to staphylococcal enterotoxin type E, France, 2009. EuroSurveillance. 2010, 15, p. 19528. Available online: http://www.eurosurveillance.org/ViewArticle.aspx?ArticleId=19528 (accessed on 13 July 2010).
- Hennekinne, J.A.; Brun, V.; De Buyser, M.L.; Dupuis, A.; Ostyn, A.; Dragacci, S. Innovative contribution of mass spectrometry to characterise staphylococcal enterotoxins involved in food outbreaks. Appl. Environ. Microbiol. 2009, 75, 882–884. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hennekinne, J.-A.; Ostyn, A.; Guillier, F.; Herbin, S.; Prufer, A.-L.; Dragacci, S. How Should Staphylococcal Food Poisoning Outbreaks Be Characterized? Toxins 2010, 2, 2106-2116. https://doi.org/10.3390/toxins2082106
Hennekinne J-A, Ostyn A, Guillier F, Herbin S, Prufer A-L, Dragacci S. How Should Staphylococcal Food Poisoning Outbreaks Be Characterized? Toxins. 2010; 2(8):2106-2116. https://doi.org/10.3390/toxins2082106
Chicago/Turabian StyleHennekinne, Jacques-Antoine, Annick Ostyn, Florence Guillier, Sabine Herbin, Anne-Laure Prufer, and Sylviane Dragacci. 2010. "How Should Staphylococcal Food Poisoning Outbreaks Be Characterized?" Toxins 2, no. 8: 2106-2116. https://doi.org/10.3390/toxins2082106