Effects of Ochratoxin A on Livestock Production




Abstract
:1. Introduction

{kind=link}
{kind=link}
| Products Intended for Animal Feed | Guidance Valuein mg/kg |
|---|---|
| Feed materials | |
| Cereals and cereal products | 0.25 |
| Complementary and complete feedingstuffs | |
| Complementary and complete feedingstuffs for pigs | 0.05 |
| Complementary and complete feedingstuffs for poultry | 0.1 |
2. Occurrence of OTA in Feeds
3. Effects of OTA on Livestock Performance and Health
3.1. Swine
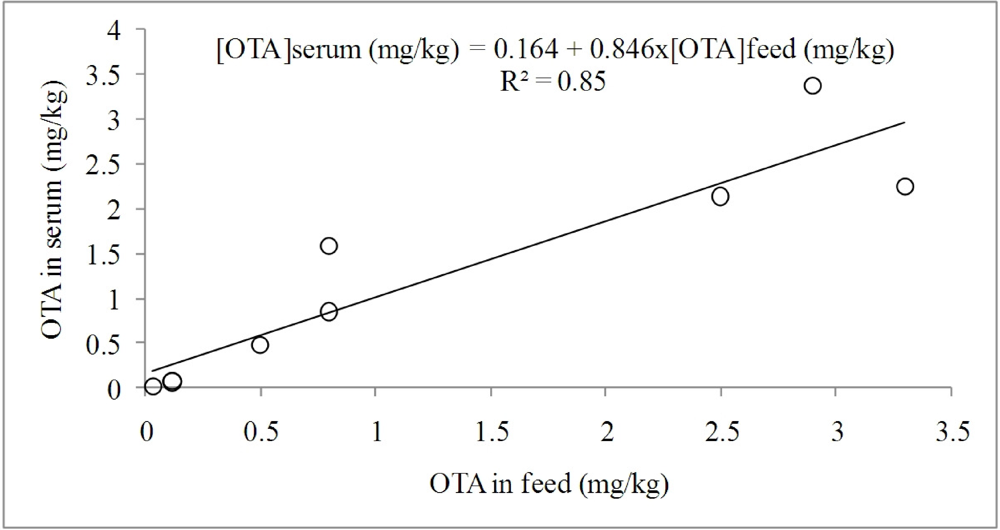
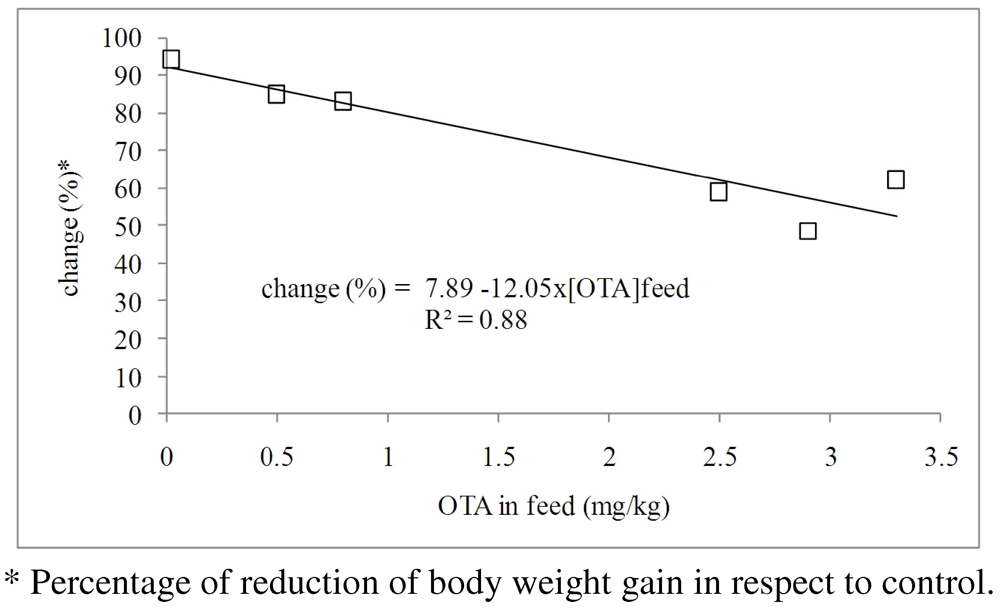
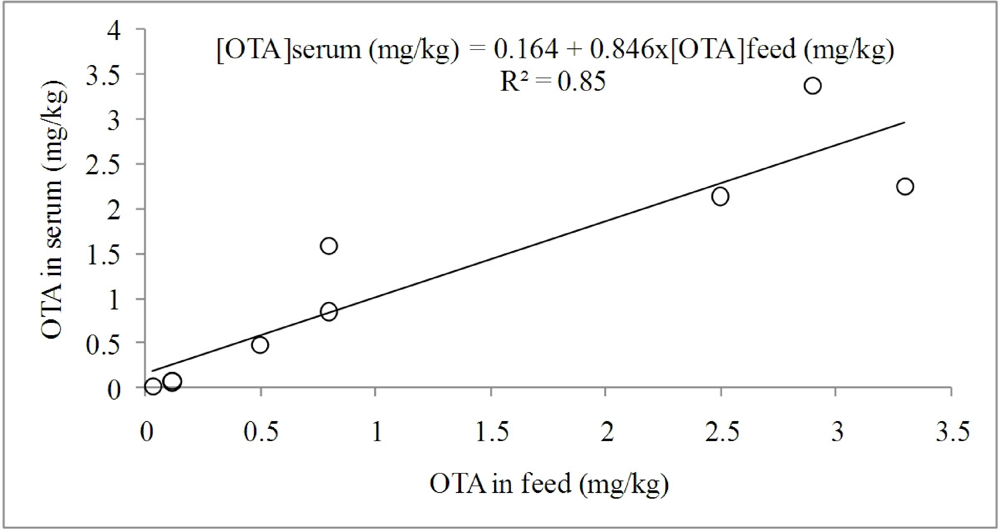
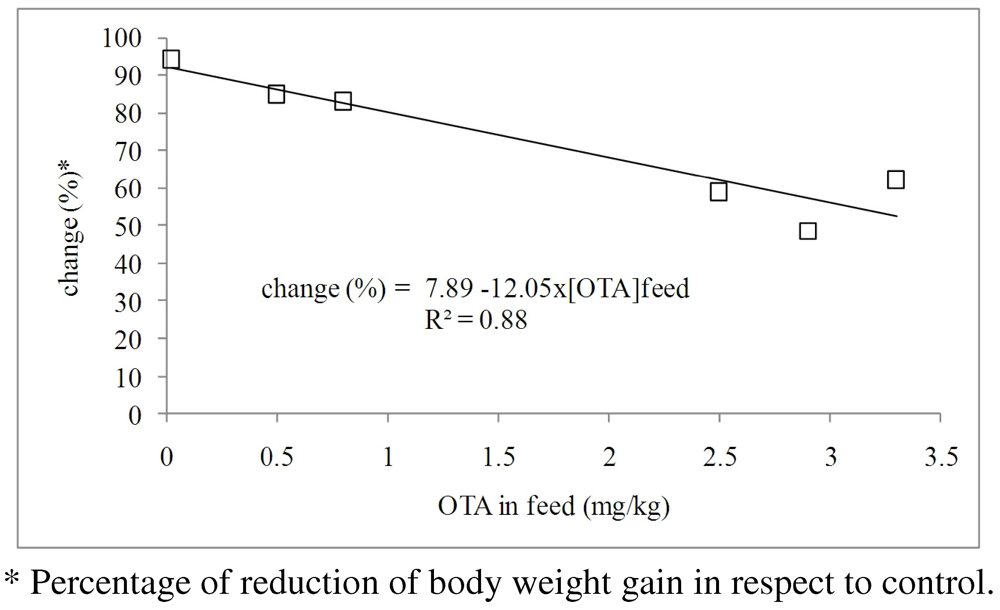
3.1.1. Interaction of OTA with Other Mycotoxins
3.1.2. OTA Binders Supplements
3.2. Poultry
| Breed | Initial age | OTA in feed (mg/kg) | Exposition time (weeks) | Change in BW, as % of control | Reference |
|---|---|---|---|---|---|
| commercial broilers | 1-day-old | 0.5 | 4 | −0.5 | [65] |
| 1 | −2.4 | ||||
| 2 | −14.5 | ||||
| 4 | −39.8 | ||||
| 8 | −74.2 | ||||
| Comb White Leghorn | 7-day-old | 2.0 | 1 | −11.0 | [66] |
| 2 | −12.0 | ||||
| 4.0 | 1 | −28 | |||
| 2 | −31 | ||||
| Leghorn | 10-day-old | 2.5 | 2 | −27.5 | [67] |
| 5 | −28.0 | ||||
| Petersen × Hubbard | 1-day-old | 2.5 | 1 | −2.2 | [68] |
| 2 | −19.3 | ||||
| 3 | −22.7 | ||||
| commercial broilers | 1-day-old | 2.0 | 5 | −24.5 | [69] |
| Plymouth Rock | 2-week- old | 1 | 2 | −0.8 | [59] |
| 4 | −15.8 | ||||
| 6 | −18.5 | ||||
| 10 | −27.2 | ||||
| 5 | 2 | −31.5 | |||
| 4 | −44.4 | ||||
| 6 | −60.9 | ||||
| 10 | −45.3 | ||||
| Ross | 1-day-old | 0.4 | 1 | −26.5 | [70] |
| 2 | −29.6 | ||||
| 3 | −41.1 | ||||
| 4 | −45.8 | ||||
| 5 | −48.7 | ||||
| 0.8 | 1 | −42.5 | |||
| 2 | −47.7 | ||||
| 3 | −57.7 | ||||
| 4 | −63.7 | ||||
| 5 | −66.5 |
| Breed | OTA in feed (mg/kg) | Exposition time | Change in number of eggs, in % respect to control * | Change in weight of eggs, in % respect to control * | Reference |
|---|---|---|---|---|---|
| Shaver 288 | 1.7 | 2 weeks | ns | ns | [72] |
| 3.1 | −51.8 | ns | |||
| 3 | (25 °C) | −24 | ns | ||
| 3 | (33 °C) | −52.4 | −3 | ||
| Hisex Brown | 2 | 3 weeks | −6.9 | −5.6 | [108] |
| White leghorn | 1 | 60 days | −21.6 | [64] | |
| Plymouth Rock | 1 | 10 days | −10.8 | −3.5 | [80] |
| 5 | −49.2 | −19 |
3.2.1. Interaction of OTA with Other Mycotoxins
3.2.2. OTA Binders Supplements
3.3. Cattle
3.4. Sheep and Goat
4. OTA Presence in Meat, Eggs and Milk
4.1. Swine
| Tissue | No. samples | LOD (ppt) | LOQ (ppt) | Incidence of positives (%) | Mean of positives (ppt) | Range (ppt) | Country | Reference |
|---|---|---|---|---|---|---|---|---|
| kidney | 300 | 0.05–0.2 | 0.15–0.5 | 13.4 | 0.81 | >0.17–1.4 | France | [101] |
| kidney | 710 | 0.11–0.2 | 0.5 | 25.9 | 0.5–5 | France | ||
| kidney | 300 | 0.02 | 0.06 | 94.7 | 0.5 | 0.02–15 | Denmark | [102] |
| muscle fillet | 300 | 0.03 | 0.09 | 76 | 0.12 | 0.03–2.9 | Denmark | |
| muscle for ham | 22 | 0.01 | 0.03 | 9 | 0.05 | 0.04–0.06 | Italy | [103] |
| kidney | 54 | 0.01 | 0.52 | 100 | 0.29 | Italy | [104] | |
| muscle | 54 | 0.01 | 77.8 | 0.024 | Italy | |||
| kidney | 90 | 0.01 | 33.3 | 1.26 | 0.17–52.5 | Serbia | [105] | |
| liver | 90 | 0.01 | 26.6 | 0.63 | 0.22–14.5 | Serbia | ||
| kidney | 60 | 0.5 | 1 | 70 | 3.97 | 1.3–22.0 | Serbia | [106] |
| liver | 60 | 0.5 | 1 | 65 | 3.2 | 1.2–19.5 | Serbia |
4.2. Poultry
4.3. Ruminants
5. Conclusions
Acknowledgements
References
- Magan, N.; Aldred, D. Conditions of formation of ochratoxin A in drying, transport and in different commodities. Food Add. Contam. 2005, 1, 10–16. [Google Scholar]
- Ringot, D.; Chango, A.; Schneider, Y.J.; Larondelle, Y. Toxicokinetics and toxicodynamics of ochratoxin A, an uptade. Chem. Biol. Interact. 2006, 159, 18–46. [Google Scholar]
- Palma, N.; Cinelli, S.; Sapora, O.; Wilson, S.H.; Dogliotti, E. Ochratoxin A-induced mutagenesis in mammalian cells is consistent with the production of oxidative stress. Chem. Res. Toxicol. 2007, 20, 1031–1037. [Google Scholar]
- Pfohl-Leszkowicz, A.; Manderville, R.A. Ochratoxin A: An overview on toxicity and carcinogenicity in animals and humans. Mol. Nutr. Food Res. 2007, 51, 61–99. [Google Scholar]
- Bruinink, A.; Rasonyi, T.; Sidler, C. Differences in neurotoxic effects of ochratoxin A, ochracin and ochratoxin-alpha in vitro. Nat. Toxins 1998, 6, 173–177. [Google Scholar]
- Galtier, P.; Alvinerie, M.; Charpenteau, J.L. The pharmacokinetic profiles of ochratoxin A in pigs, rabbits and chickens. Food Cosmet. Toxicol. 1981, 19, 735–738. [Google Scholar]
- Sreemannarayana, O.; Frohlich, A.A.; Vitti, T.G.; Marquardt, R.R.; Abramson, D. Studies of the tolerance and disposition of Ochratoxin A in young calves. J. Anim. Sci. 1988, 66, 1703–1711. [Google Scholar]
- European Commission. Commission Recommendation of 17 August 2006, on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Official J. Eur. Union 2006, L 229, 7–9. [Google Scholar]
- Potentially Hazardous Contaminants in Animal Feed and Feed Ingredients. Available online: http://www.fda.gov/AnimalVeterinary/SafetyHealth/AnimalFeedSafetySystemAFSS/ucm054996.htm (accessed on 26 February 2010).
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Harwig, J.; Munro, I.C. Mycotoxins of possible importance in diseases of Canadian farm animals. Can. Vet. J. 1975, 16, 125–141. [Google Scholar]
- Scudamore, K.A. Prevention of ochratoxin A in commodities and likely effects of processing fractionation and animal feeds. Food Addit. Contam. A 2005, 22, 17–25. [Google Scholar]
- Cairns-Fuller, V.; Aldred, D.; Magan, N. Water, temperature and gas composition interactions affect growth and ochratoxin A production by isolates of Penicillium verrucosum on wheat grain. J. Appl. Microb. 2005, 99, 1215–1221. [Google Scholar] [CrossRef]
- Shotwell, O.L.; Hesseltine, C.W.; Goulden, M.L. Ochratoxin A: Occurrence as natural contaminant of a corn sample. Appl. Microb. 1969, 17, 765–766. [Google Scholar]
- Buckle, A.E. The occurrence of mycotoxins in cereals and animal feedstuffs. Vet. Res. Com. 1983, 7, 171–186. [Google Scholar]
- Scudamore, K.A.; Hetmanski, M.T.; Chan, H.K.; Collins, S. Occurrence of mycotoxins in raw ingredients used for animal feeding stuffs in the UK in 1992. Food Add. Contam. 1997, 14, 157–173. [Google Scholar]
- Rafai, P.; Bata, A.; Jakab, L.; Vanyi, A. Evaluation of mycotoxin-contaminated cereals for their use in animal feeds in Hungary. Food Addit. Contam. 2000, 17, 799–808. [Google Scholar]
- Fazekas, B.; Tar, A.K.; Zomborszky-Kovács, M. Ochratoxin a contamination of cereal grains and coffee in Hungary in the year 2001. Acta Vet. Hung. 2002, 50, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Czerwiecki, L.; Czajkowska, D.; Witkowska-Gwiazdowska, A. On ochratoxin A and fungal flora in Polish cereals from conventional and ecological farms—Part 1: Occurrence of ochratoxin A and fungi in cereals in 1997. Food Addit. Contam. A 2002, 19, 470–477. [Google Scholar]
- Czerwiecki, L.; Czajkowska, D.; Witkowska-Gwiazdowska, A. On ochratoxin A and fungal flora in Polish cereals from conventional and ecological farms. Part 2: Occurrence of ochratoxin A and fungi in cereals in 1998. Food Addit. Contam. A 2002, 19, 1051–1057. [Google Scholar] [CrossRef]
- Garalevičienė, D.; Pettersson, H.; Agnedal, M. Occurrence of trichothecenes, zearalenone and ochratoxin A in cereals and mixed feed from central Lithuania. Mycotoxin Res. 2002, 18, 77–89. [Google Scholar]
- Baliukoniene, V.; Bakutis, B.; Stankevicius, H. Mycological and mycotoxicological evaluation of grain. Ann. Agric. Environ. Med. 2003, 10, 223–227. [Google Scholar]
- Rosa, C.A.R.; Ribeiro, J.M.M.; Fraga, M.J.; Gatti, M.; Cavaglieri, L.R.; Magnoli, C.E.; Dalcero, A.M.; Lopes, C.W.G. Mycoflora of poultry feeds and ochratoxin-producing ability of isolated Aspergillus and Penicillium species. Vet. Microb. 2006, 113, 89–96. [Google Scholar]
- Fraga, M.E.; Curvello, F.; Gatti, M.J.; Cavaglieri, L.R.; Dalcero, A.M.; Rosa, C.A.R. Potential aflatoxin and ochratoxin A production by Aspergillus species in poultry feed processing. Vet. Res. Com. 2007, 31, 343–353. [Google Scholar]
- Rosa, C.A.R.; Keller, K.M.; Keller, L.A.M.; González Pereyra, M.L.; Pereyra, C.M.; Dalcero, A.M.; Cavaglieri, L.R.; Lopes, C.W.G. Mycological survey and ochratoxin A natural contamination of swine feedstuffs in Rio de Janeiro State, Brazil. Toxicon 2009, 53, 283–288. [Google Scholar]
- Binder, E.M.; Tan, L.M.; Chin, L.J.; Handl, J.; Richard, J. Worldwide occurrence of mycotoxins in commodities, feeds and feed ingredients. Anim. Feed Sci. Tech. 2007, 137, 265–282. [Google Scholar] [CrossRef]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in maize, grass and wheat silage for dairy cattle in the Netherlands. Food Addit. Contam. B 2008, 1, 41–50. [Google Scholar]
- Driehuis, F.; Spanjer, M.C.; Scholten, J.M.; Te Giffel, M.C. Occurrence of mycotoxins in feedstuffs of dairy cows and estimation of total dietary intakes. J. Dairy Sci. 2008, 91, 4261–4271. [Google Scholar]
- Lund, F.; Frisvad, J.C. Penicillium verrucosum in wheat and barley indicates presence of ochratoxin A. J. Appl. Microb. 2003, 95, 1117–1123. [Google Scholar]
- Lindblad, M.; Johnsson, P.; Jonsson, N.; Lindqvist, R.; Olsen, M. Predicting noncompliant levels of ochratoxin A in cereal grain from Penicillium verrucosum counts. J. Appl. Microb. 2004, 97, 609–616. [Google Scholar]
- JECFA (Joint FAO/WHO Expert Committee on Food Additives). Safety Evaluation of Certain Mycotoxins in Food. WHO Food Additives Series 59. 2008. Available online: http://whqlibdoc.who.int/publications/2008/9789241660594_eng.pdf (accessed on 25 February 2010).
- Duarte, S.C.; Pena, A.; Lino, C.M. A review on ochratoxin A occurrence and effects of processing of cereal and cereal derived food products. Food Microb. 2010, 27, 187–198. [Google Scholar]
- Larsen, S. On chronic degeneration of the kidneys caused by mouldy rye. Maanedsskr. Dyrl. 1928, 40, 259–284. [Google Scholar]
- Elling, F.; Moller, T. Mycotoxic nephropathy in pigs. Bull. World Health Organ. 1973, 49, 411–418. [Google Scholar]
- Szczech, G.M.; Carlton, W.W.; Tuite, J.; Caldwell, R. Ochratoxin A toxicosis in swine. Vet. Pathol. 1973, 10, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Rutqvist, L.; Bjorklund, N.E.; Hult, K.; Hokby, E.; Carlsson, B. Ochratoxin A as the cause of spontaneous nephropathy in fattening pigs. Appl. Environ. Microbiol. 1978, 36, 920–925. [Google Scholar]
- Hult, K.; Hökby, E.; Gatenbeck, S.; Rutqvist, L. Ochratoxin A in blood from slaughter pigs in Sweden: use in evaluation of toxin content of consumed feed. Appl. Environ. Microbiol. 1980, 39, 828–830. [Google Scholar]
- Krogh, P. Epidemiology of mycotoxic porcine nephropathy. Nord. Vet. Med. 1976, 28, 452–458. [Google Scholar]
- Krogh, P.; Axelsen, N.H.; Elling, F.; Gyrd-Hansen, N.; Hald, B.; Hyldgaard-Jensen, J.; Larsen, A.E.; Madsen, A.; Mortensen, H.P.; Moller, T.; Petersen, O.K.; Ravnskov, U.; Rostgaard, M.; Aalund, O. Experimental porcine nephropathy. Changes of renal function and structure induced by ochratoxin A- contaminated feed. Acta Pathol. Microbiol. Scand. 1974, 246, 1–21. [Google Scholar]
- Lippoldl, C.C.; Stothers, S.C.; Frohlich, A.A.; Boila, R.J.; Marquardt, R.R. Effects of periodic feeding of diets containing ochratoxin A on the performance and clinical chemistry of pigs from 15 to 50 kg body weight. Can. J. Anim. Sci. 1992, 72, 135–146. [Google Scholar]
- Stoev, S.D.; Paskalev, M.; MacDonald, S.; Mantle, P.G. Experimental one year ochratoxin A toxicosis in pigs. Exp. Toxicol. Path. 2002, 53, 481–487. [Google Scholar]
- Malagutti, L.; Zannotti, M.; Scampini, A.; Sciaraffia, F. Effects of Ochratoxin A on heavy pig production. Anim. Res. 2005, 54, 179–184. [Google Scholar]
- Jarczyk, A.; Bancewicz, E.; Jędryczko, R. An attempt at inactivation of ochratoxin A in pigs’ feed with two feed-added adsorbents. Anim. Sci. Pap. Rep. 2008, 4, 269–276. [Google Scholar]
- Aoudia, N.; Callu, P.; Grosjean, F.; Larondelle, Y. Effectiveness of mycotoxin sequestration activity of micronized wheat fibres on distribution of ochratoxin A in plasma, liver and kidney of piglets fed a naturally contaminated diet. Food Chem. Toxicol. 2009, 47, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Hult, K.; Hokby, E.; Hagglund, U.; Gatenbeck, S.; Rutquist, L.; Sellyey, G. Ochratoxin A inpig blood: Method of analysis and use of a tool for feed studies. Appl. Environ. Microbiol. 1979, 38, 772–776. [Google Scholar]
- Frohlich, A.A.; Marquardt, R.R.; Ominski, K.H. Ochratoxin A as a contaminant in the human food chain: A Canadian perspective. In Mycotoxins, Endemic Nephropathy and Urinary Tract Tumours; Castegnaro, M., Plestina, R., Dirheimer, G., Chernozemsky, I.N., Bartsch, H., Eds.; International Agency for Research on Cancer: Lyon, France, 1991; pp. 139–144. [Google Scholar]
- Battacone, G.; Carboni, G.A.; Nicolussi, P.; Ligios, C.; Pulina, G. Use of a glucomannan polymer to reduce the effects of mycotoxin-contaminated diets in finishing pigs. Ital. J. Anim. Sci. 2007, 6, 673–675, (Suppl. 1).. [Google Scholar]
- Tapia, M.O.; Seawright, A.A. Experimental ochratoxicosis A in pigs. Aust. Vet. J. 1984, 61, 219–226. [Google Scholar]
- Biró, K.; Barna-Vetró, I.; Pécsi, T.; Szabó, E.; Winkler, G.; Fink-Gremmels, J.; Solti, L. Evaluation of spermatological parameters in ochratoxin A—Challenged boars. Theriogenology 2003, 60, 199–207. [Google Scholar]
- Van Egmond, H.P.; Speijers, G.J.A. Survey of data on the incidence and levels of ochratoxin A in food and animal feed worldwide. J. Nat. Toxins 1994, 3, 25–144. [Google Scholar]
- Speijers, G.J.A.; Speijers, M.H.M. Combined toxic effects of mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Avantaggiato, G.; Havenaar, R.; Visconti, A. Assessment of the multi-mycotoxin-binding efficacy of a carbon/aluminosilicate-based product in an in vitro gastrointestinal model. J. Agric. Food Chem. 2007, 55, 4810–4819. [Google Scholar] [CrossRef] [PubMed]
- Huwig, A.; Freimund, S.; Käppeli, O.; Dutler, H. Mycotoxin detoxication of animal feed by different adsorbents. Toxicol. Lett. 2001, 122, 179–188. [Google Scholar]
- Van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxic metabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar]
- Burditt, S.J.; Hagler, W.M., Jr.; Hamilton, P.B. Feed refusal during ochratoxicosis in turkeys. Poult. Sci. 1984, 63, 2172–2174. [Google Scholar]
- Marquardt, R.R.; Frohlich, A.A. A review of recent advances in understanding ochratoxicosis. J. Anim. Sci. 1992, 70, 3968–3988. [Google Scholar] [PubMed]
- Hamilton, P.B.; Huff, W.E.; Harris, J.R.; Wyatt, R.D. Natural occurrences of ochratoxicosis in poultry. Poult. Sci. 1982, 51, 1832–1841. [Google Scholar]
- Stoev, S.D.; Djuvinov, D.; Mirtcheva, T.; Pavlov, D.; Mantle, P. Studies on some feed additives giving partial protection against ochratoxin A toxicity in chicks. Toxicol. Lett. 2002, 135, 33–50. [Google Scholar]
- Santin, E.; Paulillo, A.C.; Maiorka, P.C.; Alessi, A.C.; Krabbe, E.L.; Maiorka, A. The effects of ochratoxin/aluminosilicate interaction on the tissues and humoral immune response of broilers. Avian Path. 2002, 31, 73–79. [Google Scholar]
- Gupta, S.; Jindal, N.; Khokhar, R.S.; Asrani, R.K.; Ledoux, D.R.; Rottinghaus, G.E. Individual and combined effects of ochratoxin A and Salmonella enterica serovar Gallinarum infection on pathological changes in broiler chickens. Avian Path. 2008, 37, 265–272. [Google Scholar]
- Kumar, A.; Jindal, N.; Shukla, C.L.; Asrani, R.K.; Ledoux, D.R.; Rottinghaus, G.E. Pathological changes in broiler chickens fed ochratoxin A and inoculated with Escherichia coli. Avian Path. 2004, 33, 413–417. [Google Scholar]
- Bozzo, G.; Ceci, E.; Bonerba, E.; Desantis, S.; Tantillo, G. Ochratoxin A in laying hens: High-performance liquid chromatography detection and cytological and histological analysis of target tissues. J. Appl. Poult. Res. 2008, 17, 151–156. [Google Scholar]
- Sawale, G.K.; Gosh, R.C.; Ravikanth, K.; Maini, S.; Rekhe, D.S. Experimental Mycotoxicosis in Layer Induced by Ochratoxin A and its Amelioration with Herbomineral Toxin Binder ‘Toxiroak’. Int. J. Poult. Sci. 2009, 8, 798–803. [Google Scholar]
- Huff, W.E.; Wyatt, R.D.; Hamilton, P.B. Nephrotoxicity of dietary ochratoxin a in broiler chickens. Appl. Microb. 1975, 30, 48–51. [Google Scholar]
- Rotter, R.G.; Frohlich, A.A.; Marquardt, R.R. Influence of dietary charcoal on ochratoxin A toxicity in Leghorn chicks. Can. J. Vet. Res. 1989, 53, 449–453. [Google Scholar]
- Hoehler, D.; Marquardt, R.R.; Frohlich, A.A. Lipid peroxidation as one mode of action in ochratoxin A toxicity in rats and chicks. Can. J. Anim. Sci. 1997, 77, 287–292. [Google Scholar]
- Gentles, A.; Smith, E.E.; Kubena, L.F.; Duffus, E.; Johnson, P.; Thompson, J.; Harvey, R.B.; Edrington, T.S. Toxicological evaluations of cyclopiazonic acid and ochratoxin A in broilers. Poult. Sci. 1999, 78, 1380–1384. [Google Scholar]
- Raju, M.V.L.N.; Devegowda, G. Influence of esterified-glucomannan on performance and organ morphology, serum biochemistry and haematology in broilers exposed to individual and combined mycotoxicosis (aflatoxin, ochratoxin and T-2 toxin). Brit. Poult. Sci. 2000, 41, 640–650. [Google Scholar] [CrossRef]
- Elaroussi, M.A.; Mohamed, F.R.; El Barkouky, E.M.; Atta, A.M.; Abdou, A.M.; Hatab, M.H. Experimental ochratoxicosis in broiler chickens. Avian Path. 2006, 35, 263–269. [Google Scholar]
- Verma, J.; Johri, T.S.; Swain, B.K. Effect of aflatoxin, ochratoxin and their combination on protein and energy utilisation in white leghorn laying hens. J. Sci. Food Agric. 2007, 87, 760–764. [Google Scholar]
- Haazele, F.M.; Guenter, W.; Marquardt, R.R.; Frohlich, A.A. Beneficial effects of dietary ascorbic acid supplement on hens subjected to ochratoxin A toxicosis under normal and high ambient temperatures. Can. J. Anim. Sci. 1993, 73, 149–157. [Google Scholar]
- Prior, M.G.; Sisodia, C.S.; O'Neil, J.B.; Hrudka, F. Effect of ochratoxin A on fertility and embryo viability of Japanese quail (Coturnix coturnix japonica). Can. J. Anim. Sci. 1979, 59, 605–609. [Google Scholar]
- Prior, M.G.; O'Neil, J.B.; Sisodia, C.S. Effects of ochratoxin A on production characteristics and hatchability of Japanese quail. Can. J. Anim. Sci. 1978, 58, 29–33. [Google Scholar]
- Scudamore, K.A.; Patel, S. Survey for ochratoxin A, zearalenone and fumonisins in maize imported into the United Kingdom. Food Addit. Contam. 2000, 17, 407–416. [Google Scholar]
- Kubena, L.F.; Harvey, R.B.; Huff, W.E.; Corrier, D.E.; Philips, T.D.; Rottinghaus, G.E. Influence of ochratoxin A and T-2 toxin singly and in combination on broiler chickens. Poult. Sci. 1989, 68, 867–872. [Google Scholar]
- Garcia, A.R.; Avila, E.; Rosiles, R.; Petrone, V.M. Evaluation of two mycotoxin binders to reduce toxicity of broiler diets containing ochratoxin A and T-2 toxin contaminated grain. Avian Dis. 2003, 47, 691–699. [Google Scholar]
- Wang, G.H.; Xue, C.Y.; Chen, F.; Ma, Y.L.; Zhang, X.B.; Bi, Y.Z.; Cao, Y.C. Effects of combinations of ochratoxin A and T-2 toxin on immune function of yellow-feathered broiler chickens. Poult. Sci. 2009, 88, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Sakhare, P.S.; Harne, S.D.; Kalorey, D.R.; Warke, S.R.; Bhandarker, A.G.; Kurkure, N.V. Effect of toxiroak polyherbal feed supplement during induced aflatoxicosis, ochratoxicosis and combined mycotoxicosis in broilers. Vet. Arch. 2007, 77, 129–146. [Google Scholar]
- Stoev, S.D. Studies on some feed additives and materials giving partial protection against the suppressive effect of ochratoxin A on egg production of laying hens. Res. Vet. Sci. 2010. [Google Scholar]
- Garalevičienė, D. Effect of antioxidant preparation “oxynil” on health status and productivity of laying hens fed naturally moulded feed. Vet. Zootech. 2003, 24, 22–29. [Google Scholar]
- Diaz, D.E.; Smith, T.K. Mycotoxin sequestering agents: practical tools for the neutralisation of mycotoxins. In The Mycotoxin Blue Book; Diaz, D., Ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 323–339. [Google Scholar]
- Hult, K.; Teiling, A.; Gatenbeck, S. Degradation of ochratoxin A by a ruminant. Appl. Environ. Microbiol. 1976, 32, 443–444. [Google Scholar] [PubMed]
- Ribelin, W.E.; Fukushima, K.; Still, P.E. The toxicity of ochratoxin to ruminants. Can. J. Comp. Med. 1978, 42, 172–176. [Google Scholar]
- Müller, H.M.; Müller, K.; Steingass, H. Effect of feeding regime on the metabolism of Ochratoxin A during the in vitro incubation in buffered rumen fluid from cows. Arch. Anim. Nutr. 2001, 54, 265–279. [Google Scholar]
- Müller, H.M.; Lerch, C.; Müller, K.; Eggert, W. Kinetic profiles of ochratoxin A and ochratoxin α during in vitro incubation in buffered forestomach and abomasal contents from cows. Nat. Toxins 1998, 6, 251–258. [Google Scholar]
- Özpinar, H.; Augonyte, G.; Drochner, W. Inactivation of ochratoxin in ruminal fluid with variation of pH-value and fermentation parameters in an in vitro system. Environ. Toxicol. Pharmacol. 1999, 7, 1–9. [Google Scholar]
- Jouany, J.; Diaz, D. Effects of mycotoxins in ruminants. In The Mycotoxin Blue Book; Diaz, D., Ed.; Nottingham University Press: Nottingham, UK, 2005; pp. 295–321. [Google Scholar]
- Xiao, H.; Marquardt, R.R.; Frohlich, A.A.; Phillips, G.D.; Vitti, T.G. Effect of a hay and a grain diet on the rate of hydrolysis of ochratoxin A in the rumen of sheep. J. Anim. Sci. 1991, 69, 3706–3714. [Google Scholar]
- Xiao, H.; Marquardt, R.R.; Frohlich, A.A.; Phillips, G.D.; Vitti, T.G. Effect of a hay and a grain diet on the bioavailability of ochratoxin A in the rumen of sheep. J. Anim. Sci. 1991, 69, 3715–3723. [Google Scholar]
- Hohler, D.; Sudekum, K.H.; Wolffram, S.; Frohlich, A.A.; Marquardt, R.R. Metabolism and excretion of ochratoxin A fed to sheep. J. Anim. Sci. 1999, 77, 1217–1223. [Google Scholar]
- Blank, R.; Rolfs, J.P.; Sűdekum, K.H.; Frohlich, A.A.; Marquardt, R.R.; Wolffram, S. Effects of chronic ingestion of ochratoxin A on blood levels and excretion of the mycotoxin in sheep. J. Agric. Food Chem. 2003, 51, 6899–6905. [Google Scholar]
- Blank, R.; Wolffram, S. Effects of live yeast cell supplementation to high concentrate diets on the toxicokinetics of ochratoxin A in sheep. Food Addit. Contam. A 2009, 26, 119–126. [Google Scholar]
- Nip, W.K.; Chu, F.S. Fate of ochratoxin A in goats. J. Environ. Sci. Health B 1979, 14, 319–333. [Google Scholar]
- Skaug, M.A.; Helland, I.; Solvoll, K.; Saugstad, O.D. Presence of ochratoxin A in human milk in relation to dietary intake. Food Addit. Contam. A 2001, 18, 321–327. [Google Scholar] [Green Version]
- Jørgensen, K. Occurrence of ochratoxin A in commodities and processed food—A review of EU occurrence data. Food Addit. Contam. A 2005, 22, 26–30. [Google Scholar]
- Krogh, P.; Elling, F.; Hald, B.; Larsen, A.E.; Lillehoj, E.B.; Madsen, A.; Mortensen, H.P. Time-dependent disappearance of ochratoxin A residues in tissues of bacon pigs. Toxicology 1976, 6, 235–242. [Google Scholar]
- Dall’Asta, C.; Galaverna, G.; Bertuzzi, T.; Moseriti, A.; Pietri, A.; Dossena, A.; Marchelli, R. Occurrence of ochratoxin A in raw ham muscle, salami and dry-cured ham from pigs fed with contaminated diet. Food Chem. 2010, 120, 978–983. [Google Scholar]
- Food and Agriculture Organization. Worldwide regulations for mycotoxins in food and feed in 2003. In Fao Food and Nutrition Paper 81; Food and Agriculture Organization of the United Nations: Rome, Italy, 2004. [Google Scholar]
- Ministere de l’Agriculture et de la Pêche. Note de service DGAL/SDQA/N2008-8329. In Bulletin officiel; 2008; p. n. 51. [Google Scholar]
- Dragacci, S.; Grosso, F.; Bire, R.; Fremy, J.M.; Coulon, S. A french monitoring programme for determining ochratoxin A occurrence in pig kidneys. Nat. Toxins 1999, 7, 167–173. [Google Scholar]
- Jørgensen, K.; Petersen, A. Content of ochratoxin A in paired kidney and meat samples from healthy Danish slaughter pigs. Food Addit. Contam. A 2002, 19, 562–567. [Google Scholar]
- Pietri, A.; Bertuzzi, T.; Gualla, A.; Piva, G. Occurrence of ochratoxin a in raw ham muscles and in pork products from Northern Italy. Ital. J. Food Sci. 2006, 18, 99–106. [Google Scholar]
- Matrella, R.; Monaci, L.; Milillo, M.A.; Palmisano, F.; Tantillo, M.G. Ochratoxin A determination in paired kidneys and muscle samples from swines slaughtered in southern Italy. Food Control 2006, 17, 114–117. [Google Scholar]
- Milićević, D.; Jurić, V.; Stefanović, S.; Jovanović, M.; Janković, S. Survey of slaughtered pigs for occurrence of ochratoxin a and porcine nephropathy in Serbia. Int. J. Mol. Sci. 2008, 9, 2169–2183. [Google Scholar]
- Milićević, D.R.; Jurić, V.B.; Vuković, D.Z.; Mandić, M.M.; Baltić, T.M. Residue of ochratoxin A in swine tissues—risk assessment. Arch. Oncol. 2009, 17, 56–60. [Google Scholar]
- Ferrufino-Guardia, E.V.; Tangni, E.K.; Larondelle, Y.; Ponchaut, S. Transfer of ochratoxin A during lactation: Exposure of suckling via the milk of rabbit does fed a naturally-contaminated feed. Food Addit. Contam. A 2000, 17, 167–175. [Google Scholar]
- Denli, M.; Blandon, J.C.; Guynot, M.E.; Salado, S.; Perez, J.F. Efficacy of a new ochratoxin-binding agent (OcraTox) to counteract the deleterious effects of ochratoxin A in laying hens. Poult. Sci. 2008, 87, 2266–2272. [Google Scholar]
- Krogh, P.; Elling, F.; Hald, B.; Jylling, B.; Petersen, V.E.; Skadhauge, E.; Svendsen, C.K. Experimental avian nephropathy. Acta Pathol. Microbiol. Scand. A. 1976, 84, 215–221. [Google Scholar]
- Juszkiewicz, T.; Piskorska-Pliszczynska, J.; Wisniewska, H. Ochratoxin A in laying hens: Tissue deposition and passage into eggs. In Mycotoxins and Phycotoxins, 5th International IUPAC Symposium on Mycotoxins and Phytotoxins, Technical University, Vienna, Austria, 1982; pp. 122–125.
- Shreeve, B.J.; Patterson, D.S.P.; Roberts, B.A. The ‘carry-over’ of aflatoxin, ochratoxin and zearalenone from naturally contaminated feed to tissues, urine and milk of dairy cows. Food Cosmet. Toxicol. 1979, 17, 151–152. [Google Scholar]
- Valenta, H.; Goll, M. Determination of ochratoxin A in regional samples of cow's milk from Germany. Food Addit. Contam. A 1996, 13, 669–676. [Google Scholar]
- Breitholtz-Emanuelsson, A.; Olsen, M.; Oskarsson, A.; Palminger, I.; Hult, K. Ochratoxin A in cow’s milk and in human milk with corresponding human blood samples. J. AOAC Int. 1993, 76, 842–846. [Google Scholar]
- Skaug, M.A. Analysis of Norwegian milk and infant formulas for ochratoxin A. Food Addit. Contam. 1999, 16, 75–78. [Google Scholar] [Green Version]
- Boudra, H.; Barnouin, J.; Dragacci, S.; Morgavi, D.P. Aflatoxin M1 and ochratoxin A in raw bulk milk from French dairy herds. J. Dairy Sci. 2007, 90, 3197–3201. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Battacone, G.; Nudda, A.; Pulina, G. Effects of Ochratoxin A on Livestock Production. Toxins 2010, 2, 1796-1824. https://doi.org/10.3390/toxins2071796
Battacone G, Nudda A, Pulina G. Effects of Ochratoxin A on Livestock Production. Toxins. 2010; 2(7):1796-1824. https://doi.org/10.3390/toxins2071796
Chicago/Turabian StyleBattacone, Gianni, Anna Nudda, and Giuseppe Pulina. 2010. "Effects of Ochratoxin A on Livestock Production" Toxins 2, no. 7: 1796-1824. https://doi.org/10.3390/toxins2071796