Toxins–Useful Biochemical Tools for Leukocyte Research

Abstract
:
1. Introduction
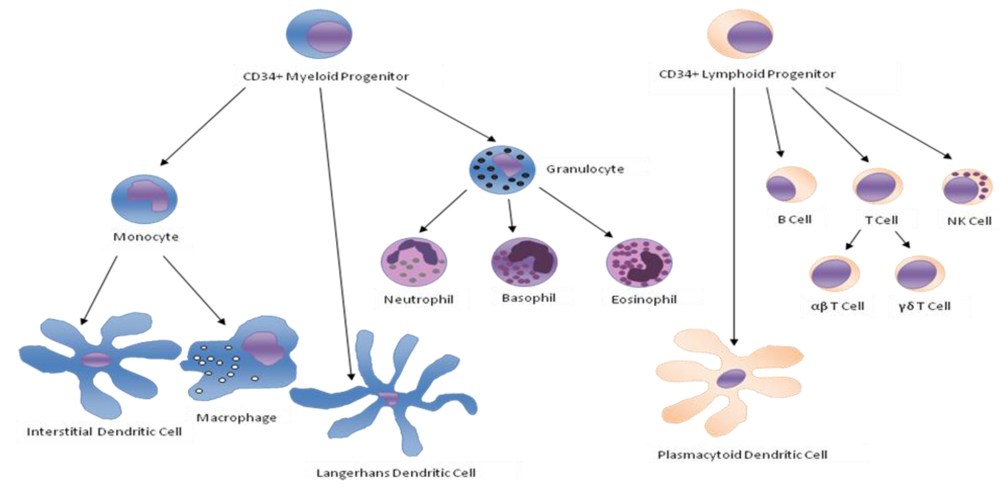

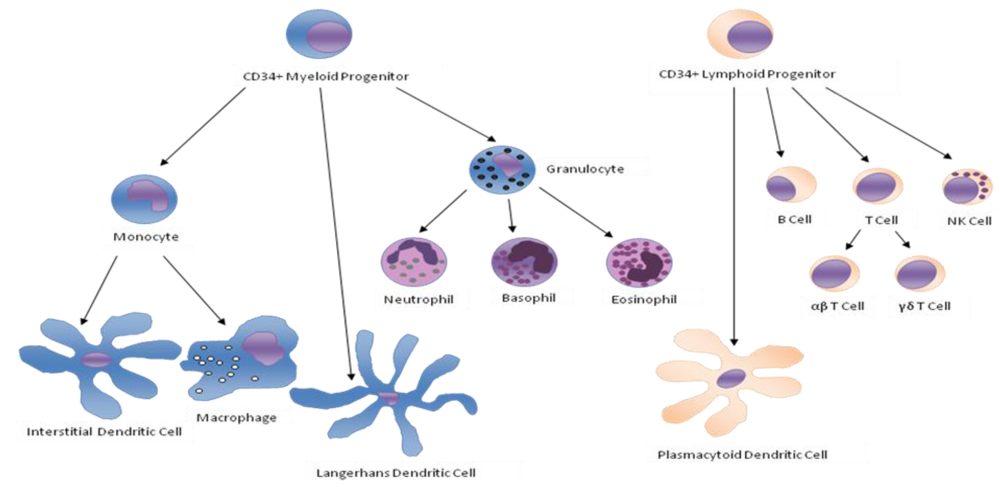
{kind=link}
{kind=link}
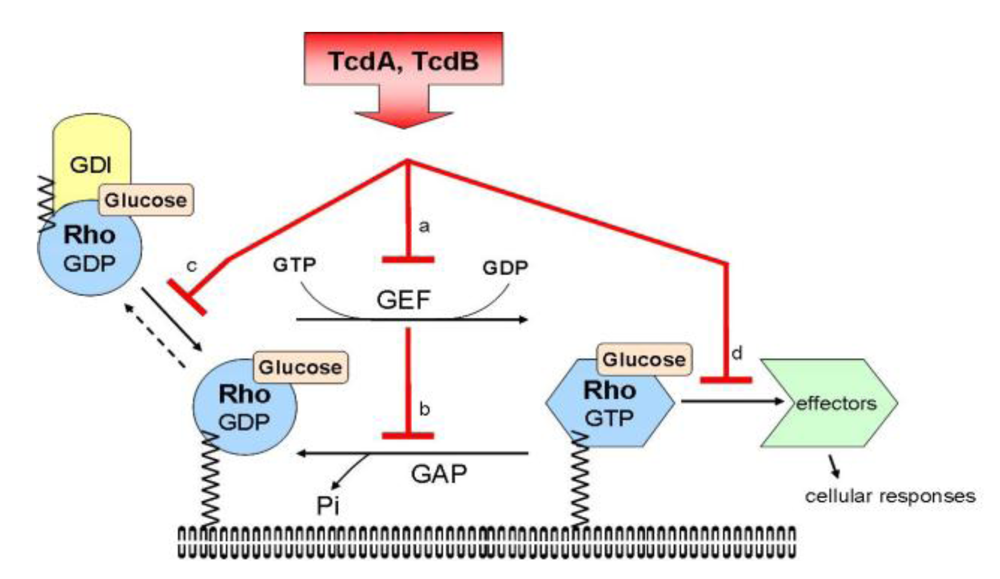{kind=link}
| Cell Type | Main functions |
|---|---|
| Monocytes | Monocytes ingest antibodies or complement opsonized material by phagocytosis [1]. They are able to produce reactive oxygen species. Once activated, monocytes can present antigens to T cells and secrete cytokines such as IL-1, TNF-α, IL-6 and INF-α/β. They can express cell-adhesion molecules and migrate to inflammation sites by chemotaxis. |
| Neutrophils | Neutrophils can express cell-adhesion molecules that allow diapedesis [2,3,4] and they can move to a site of infection or inflammation through a process called chemotaxis [5]. They can directly attack micro-organisms by phagocytosis [6] and are able to produce reactive oxygen species [7], release soluble anti-microbials and granule proteins (degranulation), and generate neutrophil extracellular traps (NETs) [8]. |
| B Cells | B cells produce antibodies, which assist in the destruction of microbes by binding to them and making them easier targets for phagocytes and activation of the complement system, and eventually develop into memory B cells after activation by antigen interaction. |
| αβ T Cells | T cells play a central role in cell-mediated immunity. There are several subgroups: |
| T cells that express the CD4 protein on their surface are called T-helper cells because they assist B cells and cytotoxic T cells. T-helper cells recognize peptide antigens associated with MHC-class II on the surface of antigen presenting cells (APCs). Once activated, they divide rapidly and secrete cytokines that regulate the immune response. | |
| T cells that express the CD8 protein on their surface are called T-cytotoxic cells (CTLs) and can destroy virally infected cells and tumor cells with toxic granules, and are also implicated in transplant rejection. | |
| Memory T cells are a subset of antigen-specific T cells that persist long-term after an infection has resolved. They quickly expand to large numbers of effector T cells upon re-exposure to their cognate antigen, thus providing the immune system with "memory" against past infections. | |
| Regulatory T cells, formerly known as suppressor T cells, are crucial for the maintenance of immunological tolerance. Their major role is to regulate T cell-mediated immunity toward the end of an immune reaction and to suppress auto-reactive T cells that escaped the process of negative selection in the thymus. | |
| γδ T Cells | This small group of T cells possesses a distinct T cell receptor (TCR) on their surface. It seems that these cells are able to recognize whole proteins rather than requiring peptides to be presented by MHC molecules on APCs. |
| γδ T cells utilize a variety of different mechanisms to regulate the inflammatory response. They secret cytokines and mediate inflammatory balance by inducing apoptosis in opposing cell populations (cytotoxicity). | |
| NK Cells | NK cells play a major role in the rejection of tumors and cells infected by viruses. The cells kill by releasing small cytoplasmic granules of proteins called perforins and granzymes. They can also kill target cells by mean of antibody dependent cell mediated cytotoxicity (ADCC). |
| Dendritic Cells | Upon activation, dendritic cells phagocyte and process antigens, migrate to lymph nodesto present the antigens to antigen-specific T cells and secrete various cytokines like IL-12 [9] and INF-γ [10] and chemokines that initiate and/or enhance many T and B lymphocyte responses. |
2. Pertussis Toxin
| Cell Type | Cell functions | Pertussis toxin |
|---|---|---|
| Monocytes | Phagocytosis | +/- [27,28,44] |
| Cytokine production | +/- [20,23,31] | |
| Chemotaxis | - [21,22,24,30,33] | |
| Migration | + [29,34] | |
| Neutrophils | Cytotoxicity | - [25] |
| Phagocytosis | - [44,45] | |
| Oxygen reactive species | - [39,40,41,42,45] | |
| Degranulation | - [36] | |
| Migration | - [37,43] | |
| Chemotaxis | - [35,38,39,45] | |
| B Cells | Proliferation | +/- [49,51,55] |
| Activation | - [50,53,54,56,57] | |
| Antibody production | - [51] | |
| Migration | - [46,47,48,52] | |
| αβ T Cells | Proliferation | + [32,60] |
| Cytokine production | +/- [60,65,71,72] | |
| Migration | - [61,64,69,70] | |
| Chemotaxis | - [58,62,67,69] | |
| Th1 selection | + [66] | |
| Th2 differentiation | + [59] | |
| γδ T Cells | Proliferation | + [68] |
| NK Cells | Cytotoxicity | - [81] |
| Chemotaxis | - [73,74,75,76,77,78,79,80,82,83,84] | |
| Dendritic Cells | Antigen presentation | - [89] |
| Cytokine production | - [26,88] | |
| Migration | - [85,86,89] | |
| Chemotaxis | - [63,87,90] |
3. Cholera Toxin
| Cell Type | Cell functions | Cholera toxin |
|---|---|---|
| Monocytes | Phagocytosis | -/ne [44,112,113] |
| Cytokine production | +/- [93,94,96,97,98,99,100,101,102,103,109,110,111] | |
| Oxygen reactive species | ne[108] | |
| Chemotaxis | ne [105,106,107] | |
| Migration | ne[104] | |
| Neutrophils | Phagocytosis | -/ne [112,119,124,125] |
| Oxygen reactive species | -/ne [116,117,120,121,123] | |
| Degranulation | ne [122,123,126] | |
| Migration | -/ne [114,115,126] | |
| Chemotaxis | - [118,127,128,129] | |
| B Cells | Proliferation | +/- [131,135,148,149,150,173,179] |
| Immunoglobulin production | + [130,131,134,135,136,138,142,143,144,147,151,152,155,171] | |
| Memory cells | + [133,141,146,153] | |
| αβ T Cells | Proliferation | +/- [131,136,157,162,164,165,167,169,172,175,177,178,179,197] |
| Activation | +/- [140,143,160,171,172,173,174,176] | |
| Cytotoxicity | + [174] | |
| Cytokine production | +/- [72,78,96,101,154,156,160,161,162,163,164,172] | |
| Chemotaxis | - [183] | |
| Th1 selection | - [95,101,159,161,165,166] | |
| Th2 differentiation | + [95,101,144,145,156,158,168,170] | |
| Tolerance or memory | + [132,146] | |
| NK Cells | Proliferation | - [139] |
| Activation | - [181,185] | |
| Cytotoxicity | - [81,182,184,186,187] | |
| Migration | - [180] | |
| Chemotaxis | - [83,84,181] | |
| Dendritic Cells | Activation | + [194,195,196,200] |
| Antigen presentation | + [191,197] | |
| Cytokine production | +/- [93,101,161,189,190,192,194,196,199] | |
| Migration | + [137,188,193,199] |
4. Toxin A and B
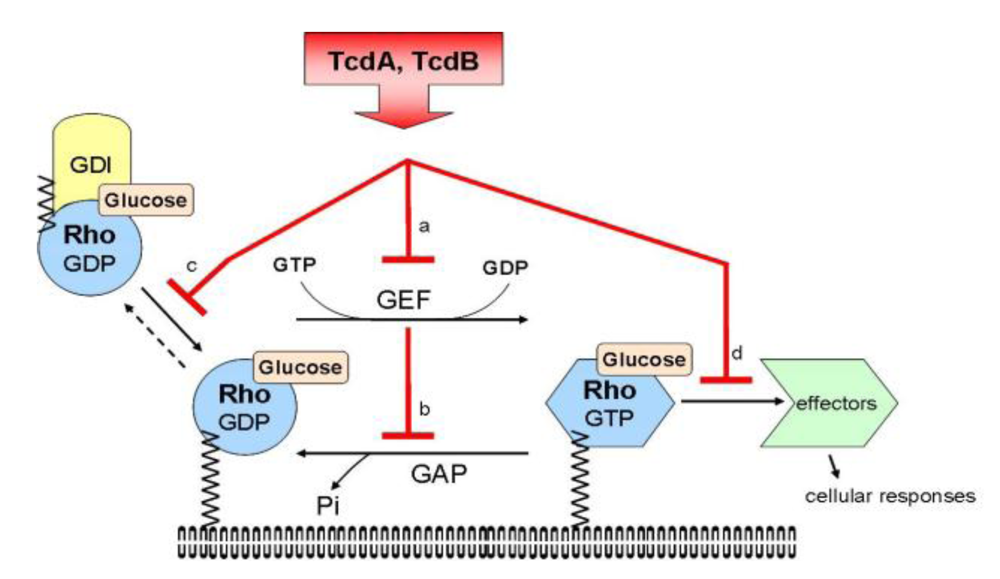
| Cell Type | Cell functions | Clostridium difficile toxins A and B |
|---|---|---|
| Monocytes | Phagocytosis | ne [205,214] |
| Cytokine production | + [206,208,209,210,212,213,214,215] | |
| Migration | - [207,211] | |
| Neutrophils | Phagocytosis | -/ne [216,222] |
| Oxygen reactive species | - [218,219] | |
| Migration | + [209,212,213,219] | |
| Chemotaxis | - [220] | |
| αβ T Cells | Proliferation | - [221] |
| NK Cells | Cytotoxicity | + [226] |
| Dendritic Cells | Maturation | + [217] |
| Phagocytosis | - [224,225] | |
| Migration | - [223] |
5. Conclusions
References and Notes
- Geske, F.J.; Monks, J.; Lehman, L.; Fadok, V.A. The role of the macrophage in apoptosis: hunter, gatherer, and regulator. Int. J. Hematol. 2002, 76, 16–26. [Google Scholar] [PubMed]
- Lawrence, M.B.; Springer, T.A. Leukocytes roll on a selectin at physiologic flow rates: distinction from and prerequisite for adhesion through integrins. Cell 1991, 65, 859–873. [Google Scholar] [PubMed]
- Zimmerman, G.A.; Prescott, S.M.; McIntyre, T.M. Endothelial cell interactions with granulocytes: tethering and signaling molecules. Immunol. Today 1992, 13, 93–100. [Google Scholar] [PubMed]
- Huber, A.R.; Kunkel, S.L.; Todd, R.F.; Weiss, S.J. Regulation of transendothelial neutrophil migration by endogenous interleukin-8. Science 1991, 254, 99–102. [Google Scholar] [PubMed]
- Zigmond, S.H. Ability of polymorphonuclear leukocytes to orient in gradients of chemotactic factors. J. Cell Biol. 1977, 75, 606–616. [Google Scholar] [PubMed]
- Hirschhorn, R.; Weissman, G. Isolation and properties of human leukocyte lyosomes in vitro. Proc. Soc. Exp. Biol. Med. 1965, 119, 36–38. [Google Scholar] [PubMed]
- Klebanoff, S.J. Myeloperoxidase-halide-hydrogen peroxide antibacterial system. J. Bacteriol. 1968, 95, 2131–2138. [Google Scholar] [PubMed]
- Volker, B.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps kill Bacteria”. Science 2004, 303, 1532–1535. [Google Scholar] [PubMed]
- Reis e Sousa, C.; Hieny, S.; Scharton-Kersten, T.; Jankovic, D.; Charest, H.; Germain, R.N.; Sher, A. In vivo microbial stimulation induces rapid CD40 ligand-independent production of interleukin 12 by dendritic cells and their redistribution to T cell areas. J. Exp. Med. 1997, 186, 1819–1829. [Google Scholar] [PubMed]
- Siega, F.P.; Kadowaki, N.; Shodell, M.; Fitzgerald-Bocarsly, P.A.; Shah, K.; Ho, S.; Antonenko, S.; Liu, Y.J. The nature of the principal type 1 interferon-producing cells in human blood. Science 1999, 284, 1835–1837. [Google Scholar] [PubMed]
- Downes, G.B.; Gautam, N. The G protein subunit gene families. Genomics 1999, 62, 544–552. [Google Scholar] [PubMed]
- Oka, Y.; Saraiva, L.R.; Kwan, Y.Y.; Korsching, S.I. The fifth class of Galpha proteins. Proc. Natl. Acad. Sci. USA 2009, 5, 1484–1489. [Google Scholar]
- Bonacci, T.M.; Ghosh, M.; Malik, S.; Smrcka, A.V. Regulatory interactions between the amino terminus of G-protein beta gamma subunits and the catalytic domain of phospholipase Cbeta2. J. Biol. Chem. 2005, 280, 10174–10181. [Google Scholar] [PubMed]
- Scheid, M.P.; Marignani, P.A.; Woodgett, J.R. Multiple phosphoinositide 3-kinase-dependent steps in activation of protein kinase B. Mol. Cell Biol. 2002, 22, 6247–6260. [Google Scholar] [PubMed]
- Sarbassov, D.D.; Guertin, D.A.; Ali, S.M.; Sabatini, D.M. Phosphorylation and regulation of Akt/PKB by the rictor-mTOR complex. Science 2005, 307, 1098–1101. [Google Scholar] [PubMed]
- Datta, S.R.; Dudek, H.; Tao, X.; Masters, S.; Fu, H.; Gotoh, Y.; Greenberg, M.E. Akt phosphorylation of BAD couples survival signals to the cell-intrinsic death machinery. Cell 1997, 91, 231–234. [Google Scholar] [PubMed]
- Bustelo, X.R.; Sauzeau, V.; Berenjeno, I.M. GTP-binding proteins of the Rho/Rac family: regulation, effectors and functions in vivo. Bioessays 2007, 4, 356–370. [Google Scholar]
- Stein, P.E.; Boodhoo, A.; Armstrong, G.D.; Cockle, S.A.; Klein, M.H.; Read, R.J. The crystal structure of pertussis toxin. Structure 1994, 1, 45–57. [Google Scholar]
- Plaut, R.D.; Carbonetti, N.H. Retrograde transport of pertussis toxin in the mammalian cell. Cell Microbiol. 2008, 5, 1130–1139. [Google Scholar]
- Schepetkin, I.A.; Kirpotina, L.N.; Tian, J.; Khlebnikov, A.I.; Ye, R.D.; Quinn, M.T. Identification of novel formyl peptide receptor-like 1 agonists that induce macrophage tumor necrosis factor alpha production. Mol. Pharmacol. 2008, 2, 392–402. [Google Scholar]
- Park, K.S.; Lee, H.Y.; Lee, S.Y.; Kim, M.K.; Kim, S.D.; Kim, J.M.; Yun, J.; Im, D.S.; Bae, Y.S. Lysophosphatidylethanolamine stimulates chemotactic migration and cellular invasion in SK-OV3 human ovarian cancer cells: involvement of pertussis toxin-sensitive G-protein coupled receptor. FEBS Lett. 2007, 581, 4411–4416. [Google Scholar] [PubMed]
- Leach, K.; Charlton, S.J.; Strange, P.G. Analysis of second messenger pathways stimulated by different chemokines acting at the chemokine receptor CCR5. Biochem. Pharmacol. 2007, 74, 881–890. [Google Scholar] [PubMed]
- Fan, H.; Williams, D.L.; Breuel, K.F.; Zingarelli, B.; Teti, G.; Tempel, G.E.; Halushka, P.V.; Cook, J.A. Gi proteins regulate lipopolysaccharide and Staphylococcus aureus induced cytokine production but not (1-3)-beta-D-glucan induced cytokine suppression. Front. Biosci. 2006, 11, 2264–2274. [Google Scholar] [PubMed]
- Musso, T.; Cappello, P.; Stornello, S.; Ravarino, D.; Caorsi, C.; Otero, K.; Novelli, F.; Badolato, R.; Giovarelli, M. IL-10 enhances CCL2 release and chemotaxis induced by CCL16 in human monocytes. Int. J. Immunopathol. Pharmacol. 2005, 18, 339–349. [Google Scholar] [PubMed]
- Bellner, L.; Thorén, F.; Nygren, E.; Liljeqvist, J.A.; Karlsson, A.; Eriksson, K. A proinflammatory peptide from herpes simplex virus type 2 glycoprotein G affects neutrophil, monocyte, and NK cell functions. J. Immunol. 2005, 174, 2235–2241. [Google Scholar] [PubMed]
- Idzko, M.; Panther, E.; Sorichter, S.; Herouy, Y.; Berod, L.; Geissler, M.; Mockenhaupt, M.; Elsner, P.; Girolomoni, G.; Norgauer, J. Characterization of the biological activities of uridine diphosphate in human dendritic cells: Influence on chemotaxis and CXCL8 release. J. Cell. Physiol. 2004, 201, 286–293. [Google Scholar] [PubMed]
- Schaeffer, L.M.; Weiss, A.A. Pertussis toxin and lipopolysaccharide influence phagocytosis of Bordetella pertussis by human monocytes. Infect. Immun. 2001, 69, 7635–7641. [Google Scholar] [PubMed]
- Boschwitz, J.S.; Batanghari, J.W.; Kedem, H.; Relman, D.A. Bordetella pertussis infection of human monocytes inhibits antigen-dependent CD4 T cell proliferation. J. Infect. Dis. 1997, 176, 678–686. [Google Scholar] [PubMed]
- Wong, W.S.; Luk, J.M. Signaling mechanisms of pertussis toxin-induced myelomonocytic cell adhesion: role of tyrosine phosphorylation. Biochem. Biophys. Res. Commun. 1997, 236, 479–482. [Google Scholar] [PubMed]
- Schratzberger, P.; Wöll, E.; Reinisch, N.; Kähler, C.M.; Wiedermann, C.J. Secretoneurin-induced in vitro chemotaxis of human monocytes is inhibited by pertussis toxin and an inhibitor of protein kinase C. Neurosci. Lett. 1996, 214, 208–210. [Google Scholar] [PubMed]
- Torre, D.; Pugliese, A.; Tambini, R.; Speranza, F.; Zeroli, C. Production and release of tumor necrosis factor alfa, interleukin-1B and interleukin-6 by human mononuclear leukocytes stimulated with pertussis toxin. New Microbiol. 1993, 16, 309–314. [Google Scholar] [PubMed]
- Grenier-Brossette, N.; Bourget, I.; Breittmayer, J.P.; Ferrua, B.; Fehlmann, M.; Cousin, J.L. Pertussis toxin-induced mitogenesis in human T lymphocytes. Immunopharmacology 1991, 21, 109–119. [Google Scholar] [PubMed]
- Verghese, M.; Uhing, R.J.; Snyderman, R. A pertussis/choleratoxin-sensitive N protein may mediate chemoattractant receptor signal transduction. Biochem. Biophys. Res. Commun. 1986, 138, 887–894. [Google Scholar] [PubMed]
- Meade, B.D.; Kind, P.D.; Manclark, C.R. Lymphocytosis-promoting factor of Bordetella pertussis alters mononuclear phagocyte circulation and response to inflammation. Infect. Immun. 1984, 46, 733–739. [Google Scholar] [PubMed]
- Mariano-Oliveira, A.; De Freitas, M.S.; Monteiro, R.Q.; Barja-Fidalgo, C. Prothrombin fragments containing kringle domains induce migration and activation of human neutrophils. Int J. Biochem. Cell. Biol. 2008, 40, 517–529. [Google Scholar] [PubMed]
- Frasch, S.C.; Zemski-Berry, K.; Murphy, R.C.; Borregaard, N.; Henson, P.M.; Bratton, D.L. Lysophospholipids of different classes mobilize neutrophil secretory vesicles and induce redundant signaling through G2A. J. Immunol. 2007, 178, 6540–6548. [Google Scholar] [PubMed]
- Lokuta, M.A.; Nuzzi, P.A.; Huttenlocher, A. Calpain regulates neutrophil chemotaxis. Proc. Natl. Acad. Sci. USA 2003, 100, 4006–4011. [Google Scholar]
- Chakraborty, A.; Hentzen, E.R.; Seo, S.M.; Smith, C.W. Granulocyte colony-stimulating factor promotes adhesion of neutrophils. Am. J. Physiol. Cell. Physiol. 2003, 284, C103–C110. [Google Scholar] [PubMed]
- Berger, M.; Budhu, S.; Lu, E.; Li, Y.; Loike, D.; Silverstein, S.C.; Loike, J.D. Different G(i)-coupled chemoattractant receptors signal qualitatively different functions in human neutrophils. J. Leukoc. Biol. 2002, 71, 798–806. [Google Scholar] [PubMed]
- Lärfars, G.; Gyllenhammar, H. Stimulus-dependent transduction mechanisms for nitric oxide release in human polymorphonuclear neutrophil leukocytes. J. Lab. Clin. Med. 1998, 132, 54–60. [Google Scholar] [PubMed]
- Rossi, M.A.; Di Mauro, C.; Dianzani, M.U. Action of lipid peroxidation products on phosphoinositide specific phospholipase C. Mol. Aspects Med. 1993, 14, 273–279. [Google Scholar] [PubMed]
- Balazovich, K.J.; Almeida, H.I.; Boxer, L.A. Recombinant human G-CSF and GM-CSF prime human neutrophils for superoxide production through different signal transduction mechanisms. J. Lab. Clin. Med. 1991, 118, 576–584. [Google Scholar] [PubMed]
- Bengtsson, T. Correlation between chemotactic peptide-induced changes in chlorotetracycline fluorescence and F-actin content in human neutrophils: a role for membrane-associated calcium in the regulation of actin polymerization? Exp. Cell Res. 1990, 191, 57–63. [Google Scholar]
- Brown, E.J.; Newell, A.M.; Gresham, H.D. Molecular regulation of phagocyte function. Evidence for involvement of a guanosine triphosphate-binding protein in opsonin-mediated phagocytosis by monocytes. J. Immunol. 1987, 139, 3777–3782. [Google Scholar] [PubMed]
- Lad, P.M.; Olson, C.V.; Smiley, P.A. Association of the N-formyl-Met-Leu-Phe receptor in human neutrophils with a GTP-binding protein sensitive to pertussis toxin. Proc. Natl. Acad. Sci. USA 1985, 82, 869–873. [Google Scholar]
- Rieken, S.; Herroeder, S.; Sassmann, A.; Wallenwein, B.; Moers, A.; Offermanns, S.; Wettschureck, N. Lysophospholipids control integrin-dependent adhesion in splenic B cells through G(i) and G(12)/G(13) family G-proteins but not through G(q)/G(11). J. Biol. Chem. 2006, 281, 36985–36992. [Google Scholar] [PubMed]
- Itakura, A.; Szczepanik, M.; Campos, R.A.; Paliwal, V.; Majewska, M.; Matsuda, H.; Takatsu, K.; Askenase, P.W. An hour after immunization peritoneal B-1 cells are activated to migrate to lymphoid organs where within 1 day they produce IgM antibodies that initiate elicitation of contact sensitivity. J. Immunol. 2005, 175, 7170–7178. [Google Scholar] [PubMed]
- Lo, C.G.; Lu, T.T.; Cyster, J.G. Integrin-dependence of lymphocyte entry into the splenic white pulp. J. Exp. Med. 2003, 197, 353–361. [Google Scholar] [PubMed]
- Deehan, M.R.; Klaus, G.G.; Holman, M.J.; Harnett, W.; Harnett, M.M. MAP-kinase: a second site of G-protein regulation of B-cell activation via the antigen receptors. Immunology 1998, 95, 169–177. [Google Scholar] [PubMed]
- Rosskopf, D.; Daelman, W.; Busch, S.; Schurks, M.; Hartung, K.; Kribben, A.; Michel, M.C.; Siffert, W. Growth factor-like action of lysophosphatidic acid on human B lymphoblasts. Am. J. Physiol. 1998, 274, C1573–C1582. [Google Scholar] [PubMed]
- Huang, R.; Cioffi, J.; Kimberly, R.; Edberg, J.; Mayer, L. B cell differentiation factor-induced human B cell maturation: stimulation of intracellular calcium release. Cell. Immunol. 1995, 164, 227–233. [Google Scholar] [PubMed]
- Cyster, J.G.; Goodnow, C.C. Pertussis toxin inhibits migration of B and T lymphocytes into splenic white pulp cords. J. Exp. Med. 1995, 182, 581–586. [Google Scholar] [PubMed]
- Huang, R.; Cioffi, J.; Berg, K.; London, R.; Cidon, M.; Maayani, S.; Mayer, L. B cell differentiation factor-induced B cell maturation: regulation via reduction in cAMP. Cell. Immunol. 1995, 162, 49–55. [Google Scholar] [PubMed]
- Melamed, I.; Wang, G.; Roifman, C.M. Antigen receptor-mediated protein tyrosine kinase activity is regulated by a pertussis toxin-sensitive G protein. J. Immunol. 1992, 149, 169–174. [Google Scholar] [PubMed]
- Kolb, J.P.; Genot, E.; Petit-Koskas, E.; Paul-Eugene, N.; Dugas, B. Effect of bacterial toxins on human B cell activation. I. Mitogenic activity of pertussis toxin. Eur. J. Immunol. 1990, 20, 969–976. [Google Scholar] [PubMed]
- Dziarski, R. Correlation between ribosylation of pertussis toxin substrates and inhibition of peptidoglycan-, muramyl dipeptide- and lipopolysaccharide-induced mitogenic stimulation in B lymphocytes. Eur. J. Immunol. 1989, 1, 125–130. [Google Scholar]
- Jakway, J.P.; DeFranco, A.L. Pertussis toxin inhibition of B cell and macrophage responses to bacterial lipopolysaccharide. Science 1986, 234, 743–746. [Google Scholar] [PubMed]
- Schneider, O.D.; Weiss, A.A.; Miller, W.E. Pertussis toxin signals through the TCR to initiate cross-desensitization of the chemokine receptor CXCR4. J. Immunol. 2009, 182, 5730–5739. [Google Scholar] [PubMed]
- Rossi-Paccani, S.; Benagiano, M.; Capitani, N.; Zornetta, I.; Ladant, D.; Montecucco, C.; D'Elios, M.M.; Baldari, C.T. The adenylate cyclase toxins of Bacillus anthracis and Bordetella pertussis promote Th2 cell development by shaping T cell antigen receptor signaling. PLoS Pathog. 2009, 5, e1000325. [Google Scholar] [CrossRef] [PubMed]
- Schneider, O.D.; Weiss, A.A.; Miller, W.E. Pertussis toxin utilizes proximal components of the T-cell receptor complex to initiate signal transduction events in T cells. Infect. Immun. 2007, 75, 4040–4049. [Google Scholar] [PubMed]
- Matsuyuki, H.; Maeda, Y.; Yano, K.; Sugahara, K.; Chiba, K.; Kohno, T.; Igarashi, Y. Involvement of sphingosine 1-phosphate (S1P) receptor type 1 and type 4 in migratory response of mouse T cells toward S1P. Cell. Mol. Immunol. 2006, 3, 429–437. [Google Scholar] [PubMed]
- Smit, M.J.; Verdijk, P.; van der Raaij-Helmer, E.M.; Navis, M.; Hensbergen, P.J.; Leurs, R.; Tensen, C.P. CXCR3-mediated chemotaxis of human T cells is regulated by a Gi- and phospholipase C-dependent pathway and not via activation of MEK/p44/p42 MAPK nor Akt/PI-3 kinase. Blood 2003, 102, 1959–1965. [Google Scholar] [PubMed]
- Panther, E.; Idzko, M.; Corinti, S.; Ferrari, D.; Herouy, Y.; Mockenhaupt, M.; Dichmann, S.; Gebicke-Haerter, P.; Di Virgilio, F.; Girolomoni, G.; Norgauer, J. The influence of lysophosphatidic acid on the functions of human dendritic cells. J. Immunol. 2002, 169, 4129–4135. [Google Scholar] [PubMed]
- Adamson, P.; Wilbourn, B.; Etienne-Manneville, S.; Calder, V.; Beraud, E.; Milligan, G.; Couraud, P.O.; Greenwood, J. Lymphocyte trafficking through the blood-brain barrier is dependent on endothelial cell heterotrimeric G-protein signaling. FASEB J. 2002, 16, 1185–1194. [Google Scholar] [PubMed]
- Nanki, T.; Lipsky, P.E. Stimulation of T-Cell activation by CXCL12/stromal cell derived factor-1 involves a G-protein mediated signaling pathway. Cell. Immunol. 2001, 214, 145–154. [Google Scholar] [PubMed]
- He, J.; Gurunathan, S.; Iwasaki, A.; Ash-Shaheed, B.; Kelsall, B.L. Primary role for Gi protein signaling in the regulation of interleukin 12 production and the induction of T helper cell type 1 responses. J. Exp. Med. 2000, 191, 1605–1610. [Google Scholar] [PubMed]
- Sotsios, Y.; Whittaker, G.C.; Westwick, J.; Ward, S.G. The CXC chemokine stromal cell-derived factor activates a Gi-coupled phosphoinositide 3-kinase in T lymphocytes. J. Immunol. 1999, 163, 5954–5963. [Google Scholar] [PubMed]
- Zorzeto, T.Q.; Hihashi, H.G.; da Silva, M.T.; Carniel Ede, F.; Dias, W.O.; Ramalho, V.D.; Mazzola, T.N.; Lima, S.C.; Morcillo, A.M.; Stephano, M.A.; Antonio, M.A.; Zanolli, M.d.L.; Raw, I.; Vilela, M.M. Immunogenicity of a whole cell pertussis vaccine with lipopolysaccharide content in infants. Clin. Vaccine Immunol. 2009, 16, 544–550. [Google Scholar] [PubMed]
- del Pozo, M.A.; Sánchez-Mateos, P.; Nieto, M.; Sánchez-Madrid, F. Chemokines regulate cellular polarization and adhesion receptor redistribution during lymphocyte interaction with endothelium and extracellular matrix. Involvement of cAMP signaling pathway. J. Cell. Biol. 1995, 131, 495–508. [Google Scholar] [CrossRef] [PubMed]
- Chaffin, K.E.; Perlmutter, R.M. A pertussis toxin-sensitive process controls thymocyte emigration. Eur. J. Immunol. 1991, 144, 2565–2573. [Google Scholar]
- Stanley, J.B.; Gorczynski, R.M.; Delovitch, T.L.; Mills, G.B. IL-2 secretion is pertussis toxin sensitive in a T lymphocyte hybridoma. J. Immunol. 1989, 142, 3546–3552. [Google Scholar] [PubMed]
- Gilmore, W.; Weiner, L.P. The effects of pertussis toxin and cholera toxin on mitogen-induced interleukin-2 production: evidence for G protein involvement in signal transduction. Cell. Immunol. 1988, 113, 235–250. [Google Scholar] [PubMed]
- Kim, S.D.; Kim, J.M.; Jo, S.H.; Lee, H.Y.; Lee, S.Y.; Shim, J.W.; Seo, S.K.; Yun, J.; Bae, Y.S. Functional expression of formyl peptide receptor family in human NK cells. J. Immunol. 2009, 183, 5511–5517. [Google Scholar] [CrossRef] [PubMed]
- Starnes, T.; Rasila, K.K.; Robertson, M.J.; Brahmi, Z.; Dahl, R.; Christopherson, K.; Hromas, R. The chemokine CXCL14 (BRAK) stimulates activated NK cell migration: implications for the downregulation of CXCL14 in malignancy. Exp. Hematol. 2006, 34, 1101–1105. [Google Scholar] [PubMed]
- Dagan-Berger, M.; Feniger-Barish, R.; Avniel, S.; Wald, H.; Galun, E.; Grabovsky, V.; Alon, R.; Nagler, A.; Ben-Baruch, A.; Peled, A. Role of CXCR3 carboxyl terminus and third intracellular loop in receptor-mediated migration, adhesion and internalization in response to CXCL11. Blood 2006, 107, 3821–3831. [Google Scholar] [PubMed]
- Jin, Y.; Knudsen, E.; Wang, L.; Maghazachi, A.A. Lysophosphatidic acid induces human natural killer cell chemotaxis and intracellular calcium mobilization. Eur. J. Immunol. 2003, 33, 2083–2089. [Google Scholar] [PubMed]
- Kveberg, L.; Bryceson, Y.; Inngjerdingen, M.; Rolstad, B.; Maghazachi, A.A. Sphingosine 1 phosphate induces the chemotaxis of human natural killer cells. Role for heterotrimeric G proteins and phosphoinositide 3 kinases. Eur. J. Immunol. 2002, 32, 1856–1864. [Google Scholar] [CrossRef] [PubMed]
- Inngjerdingen, M.; Damaj, B.; Maghazachi, A.A. Human NK cells express CC chemokine receptors 4 and 8 and respond to thymus and activation-regulated chemokine, macrophage-derived chemokine, and I-309. J. Immunol. 2000, 164, 4048–4054. [Google Scholar] [PubMed]
- Imai, T.; Hieshima, K.; Haskell, C.; Baba, M.; Nagira, M.; Nishimura, M.; Kakizaki, M.; Takagi, S.; Nomiyama, H.; Schall, T.J.; Yoshie, O. Identification and molecular characterization of fractalkine receptor CX3CR1, which mediates both leukocyte migration and adhesion. Cell 1997, 91, 521–530. [Google Scholar] [PubMed]
- Maghazachi, A.A.; Skalhegg, B.S.; Rolstad, B.; Al-Aoukaty, A. Interferon-inducible protein-10 and lymphotactin induce the chemotaxis and mobilization of intracellular calcium in natural killer cells through pertussis toxin-sensitive and -insensitive heterotrimeric G-proteins. FASEB J. 1997, 11, 765–774. [Google Scholar] [PubMed]
- Maghazachi, A.A.; Al-Aoukaty, A.; Naper, C.; Torgersen, K.M.; Rolstad, B. Preferential involvement of Go and Gz proteins in mediating rat natural killer cell lysis of allogeneic and tumor target cells. J. Immunol. 1996, 157, 5308–5314. [Google Scholar] [PubMed]
- Allavena, P.; Bianchi, G.; Zhou, D.; van Damme, J.; Jílek, P.; Sozzani, S.; Mantovani, A. Induction of natural killer cell migration by monocyte chemotactic protein-1; -2 and -3. Eur. J. Immunol. 1994, 24, 3233–3236. [Google Scholar] [PubMed]
- Maghazachi, A.A.; al-Aoukaty, A.; Schall, T.J. C-C chemokines induce the chemotaxis of NK and IL-2-activated NK cells. Role for G proteins. J. Immunol. 1994, 153, 4969–4977. [Google Scholar] [PubMed]
- Sebok, K.; Woodside, D.; al-Aoukaty, A.; Ho, A.D.; Gluck, S.; Maghazachi, A.A. IL-8 induces the locomotion of human IL-2-activated natural killer cells. Involvement of a guanine nucleotide binding (Go) protein. J. Immunol. 1993, 150, 1524–1534. [Google Scholar] [PubMed]
- Salogni, L.; Musso, T.; Bosisio, D.; Mirolo, M.; Jala, V.R.; Haribabu, B.; Locati, M.; Sozzani, S. Activin A induces dendritic cell migration through the polarized release of CXC chemokine ligands 12 and 14. Blood 2009, 113, 5848–5856. [Google Scholar] [PubMed]
- Lambert, H.; Hitziger, N.; Dellacasa, I.; Svensson, M.; Barragan, A. Induction of dendritic cell migration upon Toxoplasma gondii infection potentiates parasite dissemination. Cell. Microbiol. 2006, 8, 1611–1623. [Google Scholar] [PubMed]
- Shin, E.H.; Lee, H.Y.; Bae, Y.S. Leukotriene B4 stimulates human monocyte-derived dendritic cell chemotaxis. Biochem. Biophys. Res. Commun. 2006, 348, 606–611. [Google Scholar] [PubMed]
- Oz-Arslan, D.; Rüscher, W.; Myrtek, D.; Ziemer, M.; Jin, Y.; Damaj, B.B.; Sorichter, S.; Idzko, M.; Norgauer, J.; Maghazachi, A.A. IL-6 and IL-8 release is mediated via multiple signaling pathways after stimulating dendritic cells with lysophospholipids. J. Leukoc. Biol. 2006, 80, 287–297. [Google Scholar] [PubMed]
- Idzko, M.; Dichmann, S.; Ferrari, D.; Di Virgilio, F.; la Sala, A.; Girolomoni, G.; Panther, E.; Norgauer, J. Nucleotides induce chemotaxis and actin polymerization in immature but not mature human dendritic cells via activation of pertussis toxin-sensitive P2y receptors. Blood 2002, 100, 925–932. [Google Scholar] [PubMed]
- Idzko, M.; Panther, E.; Corinti, S.; Morelli, A.; Ferrari, D.; Herouy, Y.; Dichmann, S.; Mockenhaupt, M.; Gebicke-Haerter, P.; Di Virgilio, F.; Girolomoni, G.; Norgauer, J. Sphingosine 1-phosphate induces chemotaxis of immature and modulates cytokine-release in mature human dendritic cells for emergence of Th2 immune responses. FASEB J. 2002, 16, 625–627. [Google Scholar] [PubMed]
- Merritt, E.A.; Sarfaty, S.; van den Akker, F.; L'Hoir, C.; Martial, J.A.; Hol, W.G. Crystal structure of cholera toxin B-pentamer bound to receptor GM1 pentasaccharide. Protein Sci. 1994, 3, 166–175. [Google Scholar] [PubMed]
- Fujinaga, Y. Transport of bacterial toxins into target cells: pathways followed by cholera toxin and botulinum progenitor toxin. J. Biochem. 2006, 2, 155–160. [Google Scholar]
- la Sala, A.; He, J.; Laricchia-Robbio, L.; Gorini, S.; Iwasaki, A.; Braun, M.; Yap, G.; Sher, A.; Ozato, K.; Kelsall, B. Cholera toxin inhibits IL-12 production and CD8alpha+ dendritic cell differentiation by cAMP-mediated inhibition of IRF8 function. J. Exp. Med. 2009, 206, 1227–1235. [Google Scholar] [CrossRef] [PubMed]
- Domingos, M.O.; Andrade, R.G.; Barbaro, K.C.; Borges, M.M.; Lewis, D.J.; New, R.R. Influence of the A and B subunits of cholera toxin (CT) and Escherichia coli toxin (LT) on TNF-alpha release from macrophages. Toxicon 2009, 53, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Royaee, A.R.; Mendis, C.; Das, R.; Jett, M.; Yang, D.C. Cholera toxin induced gene expression alterations. Mol. Immunol. 2006, 43, 702–709. [Google Scholar] [PubMed]
- Royaee, A.R.; Hammamieh, R.; Mendis, C.; Das, R.; Jett, M.; Yang, D.C.H. Induction of immunomodulator transcriptional responses by cholera toxin. Mol. Immunol. 2006, 43, 1020–1028. [Google Scholar] [PubMed]
- Burkart, V.; Kim, Y.E.; Hartmann, B.; Ghiea, I.; Syldath, U.; Kauer, M.; Fingberg, W.; Hanifi-Moghaddam, P.; Müller, S.; Kolb, H. Cholera toxin B pretreatment of macrophages and monocytes diminishes their proinflammatory responsiveness to lipopolysaccharide. J. Immunol. 2002, 168, 1730–1737. [Google Scholar] [PubMed]
- Seldon, P.M.; Giembycz, M.A. Suppression of granulocyte/macrophage colony-stimulating factor release from human monocytes by cyclic AMP-elevating drugs: role of interleukin-10. Br. J. Pharmacol. 2001, 134, 58–67. [Google Scholar] [PubMed]
- Burkart, V.; Kim, Y.; Kauer, M.; Kolb, H. Induction of tolerance in macrophages by cholera toxin B chain. Pathobiology 1999, 67, 314–317. [Google Scholar] [PubMed]
- Yan, Z.; Yang, D.C.; Jett, M. Cholera toxin induces tumor necrosis factor alpha production in human monocytes. Mol. Cell. Biol. Res. Commun. 1999, 2, 124–130. [Google Scholar] [PubMed]
- Braun, M.C.; He, J.; Wu, C.Y.; Kelsall, B.L. Cholera toxin suppresses interleukin (IL)-12 production and IL-12 receptor beta1 and beta2 chain expression. J. Exp. Med. 1999, 189, 541–552. [Google Scholar] [PubMed]
- López-Zabalza, M.J.; Martínez-Lausín, S.; Bengoechea-Alonso, M.T.; López-Moratalla, N.; González, A.; Santiago, E. Signaling pathway triggered by a short immunomodulating peptide on human monocytes. Arch. Biochem. Biophys. 1997, 338, 136–142. [Google Scholar] [PubMed]
- Mattsson, E.; Van Dijk, H.; Van Kessel, K.; Verhoef, J.; Fleer, A.; Rollof, J. Intracellular pathways involved in tumor necrosis factor-alpha release by human monocytes on stimulation with lipopolysaccharide or staphylococcal peptidoglycan are partly similar. J. Infect. Dis. 1996, 173, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Luini, W.; Bernasconi, S.; Diomede, L.; Salmona, M.; Mantovani, A.; Sozzani, S. Phosphatidic acid and lysophosphatidic acid induce haptotactic migration of human monocytes. J. Biol. Chem. 1995, 270, 25549–25556. [Google Scholar] [PubMed]
- Badolato, R.; Johnston, J.A.; Wang, J.M.; McVicar, D.; Xu, L.L.; Oppenheim, J.J.; Kelvin, D.J. Serum amyloid A induces calcium mobilization and chemotaxis of human monocytes by activating a pertussis toxin-sensitive signaling pathway. J. Immunol. 1995, 155, 4004–4010. [Google Scholar] [PubMed]
- Mansfield, P.J.; Suchard, S.J. Thrombospondin promotes chemotaxis and haptotaxis of human peripheral blood monocytes. J. Immunol. 1994, 153, 4219–4229. [Google Scholar] [PubMed]
- Sozzani, S.; Zhou, D.; Locati, M.; Rieppi, M.; Proost, P.; Magazin, M.; Vita, N.; van Damme, J.; Mantovani, A. Receptors and transduction pathways for monocyte chemotactic protein-2 and monocyte chemotactic protein-3. Similarities and differences with MCP-1. J. Immunol. 1994, 152, 3615–3622. [Google Scholar] [PubMed]
- Puri, J.; Taplits, M.; Alava, M.; Bonvini, E.; Hoffman, T. Inhibition of release of arachidonic acid, superoxide and IL-1 from human monocytes by monoclonal anti-HLA class II antibodies: effects at proximal and distal points of inositol phospholipid hydrolysis pathway. Inflammation 1992, 16, 31–44. [Google Scholar] [PubMed]
- Sung, S.S.; Walters, J.A. Increased cyclic AMP levels enhance IL-1 alpha and IL-1 beta mRNA expression and protein production in human myelomonocytic cell lines and monocytes. J. Clin. Invest. 1991, 88, 1915–1923. [Google Scholar] [PubMed]
- Vellenga, E.; van der Vinne, B.; De Wolf, J.T.; Halie, M.R. Simultaneous expression and regulation of G-CSF and IL-6 mRNA in adherent human monocytes and fibroblasts. Br. J. Haematol. 1991, 78, 14–18. [Google Scholar] [PubMed]
- Kassis, S.; Lee, J.C.; Hanna, N. Effects of prostaglandins and cAMP levels on monocyte IL-1 production. Agents Actions 1989, 27, 274–276. [Google Scholar] [PubMed]
- Yang, K.D.; Augustine, N.H.; Shaio, M.F.; Bohnsack, J.F.; Hill, H.R. Effects of fibronectin on actin organization and respiratory burst activity in neutrophils, monocytes, and macrophages. J. Cell. Physiol. 1994, 158, 347–353. [Google Scholar] [CrossRef] [PubMed]
- O'Dorisio, M.S.; Vandenbark, G.R.; LoBuglio, A.F. Human monocyte killing of Staphylococcus aureus: modulation by agonists of cyclic adenosine 3',5'-monophosphate and cyclic guanosine 3',5'-monophosphate. Infect. Immun. 1979, 26, 604–610. [Google Scholar] [PubMed]
- Viana, C.F.; Melo, D.H.; Carneiro-Filho, B.A.; Michelin, M.A.; Brito, G.A.; Cunha, F.Q.; Lima, AA.; Ribeiro, R.A. Pro-inflammatory effects of cholera toxin: role of tumor necrosis factor alpha. Toxicon 2002, 40, 1487–1494. [Google Scholar] [PubMed]
- Garcia, J.G.; Verin, A.D.; Herenyiova, M.; English, D. Adherent neutrophils activate endothelial myosin light chain kinase: role in transendothelial migration. J. Appl. Physiol. 1998, 84, 1817–1821. [Google Scholar] [PubMed]
- Tithof, P.K.; Watts, S.; Ganey, P.E. Protein tyrosine kinase involvement in the production of superoxide anion by neutrophils exposed to Aroclor 1242, a mixture of polychlorinated biphenyls. Biochem. Pharmacol. 1997, 53, 1833–1842. [Google Scholar] [PubMed]
- Carreras, M.C.; Riobó, N.A.; Pargament, G.A.; Boveris, A.; Poderoso, J.J. Effects of respiratory burst inhibitors on nitric oxide production by human neutrophils. Free Radic. Res. 1997, 26, 325–334. [Google Scholar] [PubMed]
- Suchard, S.J.; Mansfield, P.J. Neutrophil thrombospondin receptors are linked to GTP-binding proteins. J. Cell. Physiol. 1996, 168, 217–227. [Google Scholar] [PubMed]
- Jin, G.F.; Guo, Y.S.; Ball, C.; Houston, C.W. Insulin-like growth factors enhance phagocytosis by human neutrophils in vitro. Regul. Pept. 1993, 49, 125–131. [Google Scholar] [PubMed]
- Braquet, P.; Hosford, D.; Koltz, P.; Guilbaud, J.; Paubert-Braquet, M. Effect of platelet-activating factor on tumor necrosis factor-induced superoxide generation from human neutrophils. Possible involvement of G proteins. Lipids 1991, 26, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Paubert-Braquet, M.; Hosford, D.; Klotz, P.; Guilbaud, J.; Braquet, P. Tumor necrosis factor alpha 'primes' the platelet-activating factor-induced superoxide production by human neutrophils: possible involvement of G proteins. J. Lipid Mediat. 1990, 2, S1–S4. [Google Scholar] [PubMed]
- Gabler, W.L.; Creamer, H.R.; Bullock, W.W. Fluoride activation of neutrophils: similarities to formylmethionyl-leucyl-phenylalanine. Inflammation 1989, 13, 47–58. [Google Scholar] [PubMed]
- Blackburn, W.D., Jr.; Heck, L.W. Neutrophil activation by surface bound IgG: pertussis toxin insensitive activation. Biochem. Biophys. Res. Commun. 1988, 152, 136–142. [Google Scholar] [CrossRef] [PubMed]
- Gresham, H.D.; Clement, L.T.; Volanakis, J.E.; Brown, E.J. Cholera toxin and pertussis toxin regulate the Fc receptor-mediated phagocytic response of human neutrophils in a manner analogous to regulation by monoclonal antibody 1C2. J. Immunol. 1987, 139, 4159–4166. [Google Scholar] [PubMed]
- Lad, P.M.; Olson, C.V.; Grewal, I.S. A step sensitive to pertussis toxin and phorbol ester in human neutrophils regulates chemotaxis and capping but not phagocytosis. FEBS Lett. 1986, 200, 91–96. [Google Scholar] [PubMed]
- Shefcyk, J.; Yassin, R.; Volpi, M.; Molski, T.F.; Naccache, P.H.; Munoz, J.J.; Becker, E.L.; Feinstein, M.B.; Sha'afi, R.I. Pertussis but not cholera toxin inhibits the stimulated increase in actin association with the cytoskeleton in rabbit neutrophils: role of the "G proteins" in stimulus-response coupling. Biochem. Biophys. Res. Commun. 1985, 126, 1174–1181. [Google Scholar] [PubMed]
- Roch-Arveiller, M.; Boquet, P.; Bradshaw, D.; Giroud, J.P. Inhibition by cholera toxin of rat polymorphonuclear leukocyte chemotaxis demonstrated in vitro and in vivo. Infect. Immun. 1979, 25, 187–190. [Google Scholar] [PubMed]
- Issekutz, A.C.; Ng, M.; Biggar, W.D. Effect of cyclic adenosine 3',5'-monophosphate antagonists on endotoxin-induced inhibition of human neutrophil chemotaxis. Infect. Immun. 1979, 24, 434–440. [Google Scholar] [PubMed]
- Rivkin, I.; Rosenblatt, J.; Becker, E.L. The role of cyclic AMP in the chemotactic responsiveness and spontaneous motility of rabbit peritoneal neutrophils. The inhibition of neutrophil movement and the elevation of cyclic AMP levels by catecholamines, prostaglandins, theophylline and cholera toxin. J. Immunol. 1975, 115, 1126–1134. [Google Scholar] [PubMed]
- Smits, H.H.; Gloudemans, A.K.; van Nimwegen, M.; Willart, M.A.; Soullié, T.; Muskens, F.; de Jong, E.C.; Boon, L.; Pilette, C.; Johansen, F.E.; Hoogsteden, H.C.; Hammad, H.; Lambrecht, B.N. Cholera toxin B suppresses allergic inflammation through induction of secretory IgA. Mucosal. Immunol. 2009, 2, 331–339. [Google Scholar] [PubMed]
- Negri, D.R.; Pinto, D.; Vendetti, S.; Patrizio, M.; Sanchez, M.; Riccomi, A.; Ruggiero, P.; Del Giudice, G.; De Magistris, M.T. Cholera toxin and Escherichia coli heat-labile enterotoxin, but not their nontoxic counterparts, improve the antigen-presenting cell function of human B lymphocytes. Infect. Immun. 2009, 77, 1924–1935. [Google Scholar] [PubMed]
- Sun, J.B.; Flach, C.F.; Czerkinsky, C.; Holmgren, J. B lymphocytes promote expansion of regulatory T cells in oral tolerance: powerful induction by antigen coupled to cholera toxin B subunit. J. Immunol. 2008, 181, 8278–8287. [Google Scholar] [PubMed]
- Jayasekera, C.R.; Harris, J.B.; Bhuiyan, S.; Chowdhury, F.; Khan, A.I.; Faruque, A.S.; Larocque, R.C.; Ryan, E.T.; Ahmed, R.; Qadri, F.; Calderwood, S.B. Cholera toxin-specific memory B cell responses are induced in patients with dehydrating diarrhea caused by Vibrio cholerae O1. J. Infect. Dis. 2008, 198, 1055–1061. [Google Scholar] [PubMed]
- Kataoka, K.; Fujihashi, K.; Sekine, S.; Fukuiwa, T.; Kobayashi, R.; Suzuki, H.; Nagata, H.; Takatsu, K.; Shizukuishi, S.; McGhee, J.R.; Fujihashi, K. Nasal cholera toxin elicits IL-5 and IL-5 receptor alpha-chain expressing B-1a B cells for innate mucosal IgA antibody responses. J. Immunol. 2007, 178, 6058–6065. [Google Scholar] [PubMed]
- Arce, S.; Nawar, H.F.; Muehlinghaus, G.; Russell, M.W.; Connell, T.D. In vitro induction of immunoglobulin A (IgA)- and IgM-secreting plasma blasts by cholera toxin depends on T-cell help and is mediated by CD154 up-regulation and inhibition of gamma interferon synthesis. Infect. Immun. 2007, 75, 1413–1423. [Google Scholar] [PubMed]
- Arce, S.; Nawar, H.F.; Russell, M.W.; Connell, T.D. Differential binding of Escherichia coli enterotoxins LT-IIa and LT-IIb and of cholera toxin elicits differences in apoptosis, proliferation, and activation of lymphoid cells. Infect. Immun. 2005, 73, 2718–2727. [Google Scholar] [PubMed]
- Shreedhar, V.K.; Kelsall, B.L.; Neutra, M.R. Cholera toxin induces migration of dendritic cells from the subepithelial dome region to T- and B-cell areas of Peyer's patches. Infect. Immun. 2003, 71, 504–509. [Google Scholar] [PubMed]
- Fan, J.L.; Peterson, J.W.; Prabhakar, B.S. Adjuvant effects of cholera toxin b subunit on immune response to recombinant thyrotropin receptor in mice. J. Autoimmun. 2000, 14, 43–52. [Google Scholar] [PubMed]
- Miller, J.S.; Cervenka, T.; Lund, J.; Okazaki, I.J.; Moss, J. Purine metabolites suppress proliferation of human NK cells through a lineage-specific purine receptor. J. Immunol. 1999, 162, 7376–7382. [Google Scholar] [PubMed]
- Iijima, H.; Takahashi, I.; Hiroi, T.; Shimaoka, M.; Kawano, S.; Nagano, K.; Hori, M.; Kiyono, H. Orally administered cholera toxin prevents murine intestinal T cells from staphylococcal enterotoxin B-induced anergy. Gastroenterology 1998, 115, 1197–1204. [Google Scholar] [PubMed]
- Benedetti, R.; Lev, P.; Massouh, E.; Fló, J. Long-term antibodies after an oral immunization with cholera toxin are synthesized in the bone marrow and may play a role in the regulation of memory B-cell maintenance at systemic and mucosal sites. Res. Immunol. 1998, 149, 107–118. [Google Scholar] [PubMed]
- Kim, P.H.; Eckmann, L.; Lee, W.J.; Han, W.; Kagnoff, M.F. Cholera toxin and cholera toxin B subunit induce IgA switching through the action of TGF-beta 1. J. Immunol. 1998, 160, 1198–1203. [Google Scholar] [PubMed]
- Sverremark, E.; Fernandez, C. Immunogenicity of bacterial carbohydrates: cholera toxin modulates the immune response against dextran B512. Immunology 1997, 92, 153–159. [Google Scholar] [PubMed]
- Xu-Amano, J.; Jackson, R.J.; Fujihashi, K.; Kiyono, H.; Staats, H.F.; McGhee, J.R. Helper Th1 and Th2 cell responses following mucosal or systemic immunization with cholera toxin. Vaccine 1994, 12, 903–911. [Google Scholar] [PubMed]
- Xu-Amano, J.; Kiyono, H.; Jackson, R.J.; Staats, H.F.; Fujihashi, K.; Burrows, P.D.; Elson, C.O.; Pillai, S.; McGhee, J.R. Helper T cell subsets for immunoglobulin A responses: oral immunization with tetanus toxoid and cholera toxin as adjuvant selectively induces Th2 cells in mucosa associated tissues. J. Exp. Med. 1993, 178, 1309–1320. [Google Scholar] [CrossRef] [PubMed]
- Vajdy, M.; Lycke, N. Stimulation of antigen-specific T- and B-cell memory in local as well as systemic lymphoid tissues following oral immunization with cholera toxin adjuvant. Immunology 1993, 80, 197–203. [Google Scholar] [PubMed]
- Nilssen, D.E.; Friman, V.; Theman, K.; Björkander, J.; Kilander, A.; Holmgren, J.; Hanson, L.A.; Brandtzaeg, P. B-cell activation in duodenal mucosa after oral cholera vaccination in IgA deficient subjects with or without IgG subclass deficiency. Scand. J. Immunol. 1993, 38, 201–208. [Google Scholar] [PubMed]
- Garrone, P.; Banchereau, J. Agonistic and antagonistic effects of cholera toxin on human B lymphocyte proliferation. Mol. Immunol. 1993, 30, 627–635. [Google Scholar] [PubMed]
- Anastassiou, E.D.; Yamada, H.; Boumpas, D.T.; Tsokos, G.C.; Thyphronitis, G.; Balow, J.; Mond, J.J. Cholera toxin promotes the proliferation of anti-mu antibody-prestimulated human B cells. Cell. Immunol. 1992, 140, 237–247. [Google Scholar] [PubMed]
- Dugas, B.; Paul-Eugène, N.; Génot, E.; Mencia-Huerta, J.M.; Braquet, P.; Kolb, J.P. Effect of bacterial toxins on human B cell activation. II. Mitogenic activity of the B subunit of cholera toxin. Eur. J. Immunol. 1991, 21, 495–500. [Google Scholar] [PubMed]
- Lycke, N.; Severinson, E.; Strober, W. Cholera toxin acts synergistically with IL-4 to promote IgG1 switch differentiation. J. Immunol. 1990, 145, 3316–3324. [Google Scholar] [PubMed]
- Chen, K.S.; Strober, W. Cholera holotoxin and its B subunit enhance Peyer's patch B cell responses induced by orally administered influenza virus: disproportionate cholera toxin enhancement of the IgA B cell response. Eur. J. Immunol. 1990, 20, 433–436. [Google Scholar] [PubMed]
- Lycke, N.; Hellström, U.; Holmgren, J. Circulating cholera antitoxin memory cells in the blood one year after oral cholera vaccination in humans. Scand. J. Immunol. 1987, 26, 207–211. [Google Scholar] [PubMed]
- Lee, J.B.; Jang, J.E.; Song, M.K.; Chang, J. Intranasal delivery of cholera toxin induces th17-dominated T-cell response to bystander antigens. PLoS One 2009, 4, e5190. [Google Scholar] [PubMed]
- Nyström-Asklin, J.; Adamsson, J.; Harandi, A.M. The adjuvant effect of CpG oligodeoxynucleotide linked to the non-toxic B subunit of cholera toxin for induction of immunity against H. pylori in mice. Scand. J. Immunol. 2008, 67, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.F.; Wang, C.C.; Wu, T.C.; Chu, C.H.; Peng, H.J. Effect of sublingual administration with a native or denatured protein allergen and adjuvant CpG oligodeoxynucleotides or cholera toxin on systemic T(H)2 immune responses and mucosal immunity in mice. Ann. Allergy Asthma. Immunol. 2007, 99, 443–452. [Google Scholar] [PubMed]
- Vendetti, S.; Riccomi, A.; Sacchi, A.; Sciaraffia, E.; Gatta, L.; Pioli, C.; De Magistris, M.T. Inhibition of T cell proliferation by cholera toxin involves the modulation of costimulatory molecules CTLA-4 and CD28. Immunol. Lett. 2008, 115, 59–69. [Google Scholar] [PubMed]
- Lysaght, J.; Jarnicki, A.G.; Mills, K.H. Reciprocal effects of Th1 and Treg cell inducing pathogen-associated immunomodulatory molecules on anti-tumor immunity. Cancer Immunol. Immunother. 2007, 56, 1367–1379. [Google Scholar] [PubMed]
- Su, S.B.; Silver, P.B.; Wang, P.; Chan, C.C.; Caspi, R.R. Cholera toxin prevents Th1-mediated autoimmune disease by inducing immune deviation. J. Immunol. 2004, 173, 755–761. [Google Scholar] [PubMed]
- Wang, M.; Bregenholt, S.; Petersen, J.S. The cholera toxin B subunit directly costimulates antigen-primed CD4+ T cells ex vivo. Scand. J. Immunol. 2003, 58, 342–349. [Google Scholar] [PubMed]
- Lavelle, E.C.; McNeela, E.; Armstrong, M.E.; Leavy, O.; Higgins, S.C.; Mills, K.H. Cholera toxin promotes the induction of regulatory T cells specific for bystander antigens by modulating dendritic cell activation. J. Immunol. 2003, 171, 2384–2392. [Google Scholar] [PubMed]
- Staples, K.J.; Bergmann, M.; Tomita, K.; Houslay, M.D.; McPhee, I.; Barnes, P.J.; Giembycz, M.A.; Newton, R. Adenosine 3',5'-cyclic monophosphate (cAMP)-dependent inhibition of IL-5 from human T lymphocytes is not mediated by the cAMP-dependent protein kinase A. J. Immunol. 2001, 167, 2074–2080. [Google Scholar] [PubMed]
- Pacheco, S.E.; Gibbs, R.A.; Ansari-Lari, A.; Rogers, P. Intranasal immunization with HIV reverse transcriptase: effect of dose in the induction of helper T cell type 1 and 2 immunity. AIDS Res. Hum. Retroviruses 2000, 16, 2009–2017. [Google Scholar] [PubMed]
- Eriksson, K.; Nordström, I.; Czerkinsky, C.; Holmgren, J. Differential effect of cholera toxin on CD45RA+ and CD45RO+ T cells: specific inhibition of cytokine production but not proliferation of human naive T cells. Clin. Exp. Immunol. 2000, 121, 283–288. [Google Scholar] [PubMed]
- Yamamoto, M.; Kiyono, H.; Yamamoto, S.; Batanero, E.; Kweon, M.N.; Otake, S.; Azuma, M.; Takeda, Y.; McGhee, J.R. Direct effects on antigen-presenting cells and T lymphocytes explain the adjuvanticity of a nontoxic cholera toxin mutant. J. Immunol. 1999, 162, 7015–7021. [Google Scholar] [PubMed]
- McSorley, S.J.; Rask, C.; Pichot, R.; Julia, V.; Czerkinsky, C.; Glaichenhaus, N. Selective tolerization of Th1-like cells after nasal administration of a cholera toxoid-LACK conjugate. Eur. J. Immunol. 1998, 28, 424–432. [Google Scholar] [PubMed]
- Sobel, D.O.; Yankelevich, B.; Goyal, D.; Nelson, D.; Mazumder, A. The B-subunit of cholera toxin induces immunoregulatory cells and prevents diabetes in the NOD mouse. Diabetes 1998, 47, 186–191. [Google Scholar] [PubMed]
- Marinaro, M.; Boyaka, P.N.; Finkelman, F.D.; Kiyono, H.; Jackson, R.J.; Jirillo, E.; McGhee, J.R. Oral but not parenteral interleukin (IL)-12 redirects T helper 2 (Th2)-type responses to an oral vaccine without altering mucosal IgA responses. J. Exp. Med. 1997, 185, 415–427. [Google Scholar] [PubMed]
- Yankelevich, B.; Soldatenkov, V.A.; Hodgson, J.; Polotsky, A.J.; Creswell, K.; Mazumder, A. Differential induction of programmed cell death in CD8+ and CD4+ T cells by the B subunit of cholera toxin. Cell. Immunol. 1996, 168, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Marinaro, M.; Staats, H.F.; Hiroi, T.; Jackson, R.J.; Coste, M.; Boyaka, P.N.; Okahashi, N.; Yamamoto, M.; Kiyono, H.; Bluethmann, H.; Fujihashi, K.; McGhee, J.R. Mucosal adjuvant effect of cholera toxin in mice results from induction of T helper 2 (Th2) cells and IL-4. J. Immunol. 1995, 155, 4621–4629. [Google Scholar] [PubMed]
- Bowen, J.C.; Nair, S.K.; Reddy, R.; Rouse, B.T. Cholera toxin acts as a potent adjuvant for the induction of cytotoxic T-lymphocyte responses with non-replicating antigens. Immunology 1994, 81, 338–342. [Google Scholar] [PubMed]
- Hörnquist, E.; Lycke, N. Cholera toxin adjuvant greatly promotes antigen priming of T cells. Eur. J. Immunol. 1993, 23, 2136–2143. [Google Scholar] [PubMed]
- Woogen, S.D.; Turo, K.; Dieleman, L.A.; Beagley, K.W.; Elson, C.O. Inhibition of murine T cell activation by cholera toxin B subunit is not mediated through the phosphatidylinositol second messenger system. J. Immunol. 1993, 150, 3274–3283. [Google Scholar] [PubMed]
- Valitutti, S.; Dessing, M.; Lanzavecchia, A. Role of cAMP in regulating cytotoxic T lymphocyte adhesion and motility. Eur. J. Immunol. 1993, 23, 790–795. [Google Scholar] [PubMed]
- Maghazachi, A.A. Cholera toxin inhibits interleukin-2-induced, but enhances pertussis toxin-induced T-cell proliferation: regulation by cyclic nucleotides. Immunology 1992, 75, 103–107. [Google Scholar] [PubMed]
- Gupta, S.; Shimizu, M.; Ohira, K.; Vayuvegula, B. T cell activation via the T cell receptor: a comparison between WT31 (defining alpha/beta TcR)-induced and anti-CD3-induced activation of human T lymphocytes. Cell. Immunol. 1991, 132, 26–44. [Google Scholar] [PubMed]
- Anderson, D.L.; Tsoukas, C.D. Cholera toxin inhibits resting human T cell activation via a cAMP-independent pathway. J. Immunol. 1989, 143, 3647–3652. [Google Scholar] [PubMed]
- Bjorndahl, J.M.; Sung, S.S.; Hansen, J.A.; Fu, S.M. Human T cell activation: differential response to anti-CD28 as compared to anti-CD3 monoclonal antibodies. Eur. J. Immunol. 1989, 19, 881–887. [Google Scholar] [PubMed]
- Lycke, N.; Bromander, A.K.; Ekman, L.; Karlsson, U.; Holmgren, J. Cellular basis of immunomodulation by cholera toxin in vitro with possible association to the adjuvant function in vivo. J. Immunol. 1989, 142, 20–27. [Google Scholar] [PubMed]
- Poggi, A.; Spada, F.; Costa, P.; Tomasello, E.; Revello, V.; Pella, N.; Zocchi, M.R.; Moretta, L. Dissection of lymphocyte function-associated antigen 1-dependent adhesion and signal transduction in human natural killer cells shown by the use of cholera or pertussis toxin. Eur. J. Immunol. 1996, 26, 967–975. [Google Scholar] [PubMed]
- Maghazachi, A.A.; al-Aoukaty, A. Guanine nucleotide binding proteins mediate the chemotactic signal of transforming growth factor-beta 1 in rat IL-2 activated natural killer cells. Int. Immunol. 1993, 5, 825–832. [Google Scholar] [PubMed]
- Watanabe, M.; Kiyohara, T.; Murata, K.; Yodoi, J.; Itakura, H. Cholera toxin inhibits lethal hit stage of natural killer cell-mediated cytotoxicity. Microbiol. Immunol. 1993, 37, 317–323. [Google Scholar] [PubMed]
- Maghazachi, A.A. Tumor necrosis factor-alpha is chemokinetic for lymphokine-activated killer cells: regulation by cyclic adenosine monophosphate. J. Leukoc. Biol. 1991, 49, 302–308. [Google Scholar] [PubMed]
- Whalen, M.M.; Bankhurst, A.D. Effects of beta-adrenergic receptor activation, cholera toxin and forskolin on human natural killer cell function. Biochem. J. 1990, 272, 327–331. [Google Scholar] [PubMed]
- Hussain, A.; Himeno, K.; Mayumi, H.; Kawamura, I.; Tsuru, S.; Nomoto, K. Immunomodulatory effects of cholera toxin in mice. Nat. Immun. Cell. Growth Regul. 1989, 8, 231–244. [Google Scholar] [PubMed]
- Fuyama, S.; Naiki, M.; Sendo, F. The mechanism of cholera toxin-induced suppression of natural killer cytotoxicity. Gann. 1982, 73, 798–804. [Google Scholar] [PubMed]
- Fuyama, S.; Sendo, F.; Watabe, S.; Seiji, K.; Arai, S. Inhibition of mouse natural killer activity by cholera toxin. Gann. 1981, 72, 141–144. [Google Scholar] [PubMed]
- Anosova, N.G.; Chabot, S.; Shreedhar, V.; Borawski, J.A.; Dickinson, B.L.; Neutra, M.R. Cholera toxin, E. coli heat-labile toxin, and non-toxic derivatives induce dendritic cell migration into the follicle-associated epithelium of Peyer's patches. Mucosal Immunol. 2008, 1, 59–67. [Google Scholar] [PubMed]
- Feng, B.S.; Zheng, P.Y.; Chen, X.; Liao, X.Q.; Yang, P.C. Investigation of the role of cholera toxin in assisting the initiation of the antigen-specific Th2 response. Immunol. Invest. 2008, 37, 782–797. [Google Scholar] [PubMed]
- Krishnamoorthy, N.; Oriss, T.B.; Paglia, M.; Fei, M.; Yarlagadda, M.; Vanhaesebroeck, B.; Ray, A.; Ray, P. Activation of c-Kit in dendritic cells regulates T helper cell differentiation and allergic asthma. Nat. Med. 2008, 14, 565–573. [Google Scholar] [PubMed]
- Chandy, A.G.; Nurkkala, M.; Josefsson, A.; Eriksson, K. Therapeutic dendritic cell vaccination with Ag coupled to cholera toxin in combination with intratumoural CpG injection leads to complete tumour eradication in mice bearing HPV 16 expressing tumours. Vaccine 2007, 25, 6037–6046. [Google Scholar] [PubMed]
- Bimczok, D.; Rau, H.; Wundrack, N.; Naumann, M.; Rothkötter, H.J.; McCullough, K.; Summerfield, A. Cholera toxin promotes the generation of semi-mature porcine monocyte-derived dendritic cells that are unable to stimulate T cells. Vet. Res. 2007, 38, 597–612. [Google Scholar] [CrossRef] [PubMed]
- Wendland, M.; Czeloth, N.; Mach, N.; Malissen, B.; Kremmer, E.; Pabst, O.; Förster, R. CCR9 is a homing receptor for plasmacytoid dendritic cells to the small intestine. Proc. Natl. Acad. Sci. USA 2007, 104, 6347–6352. [Google Scholar]
- Isomura, I.; Yasuda, Y.; Tsujimura, K.; Takahashi, T.; Tochikubo, K.; Morita, A. Recombinant cholera toxin B subunit activates dendritic cells and enhances antitumor immunity. Microbiol. Immunol. 2005, 49, 79–87. [Google Scholar] [PubMed]
- Bagley, K.C.; Abdelwahab, S.F.; Tuskan, R.G.; Lewis, G.K. Calcium signaling through phospholipase C activates dendritic cells to mature and is necessary for the activation and maturation of dendritic cells induced by diverse agonists. Clin. Diagn. Lab. Immunol. 2004, 11, 77–82. [Google Scholar] [PubMed]
- Bagley, K.C.; Abdelwahab, S.F.; Tuskan, R.G.; Fouts, T.R.; Lewis, G.K. Cholera toxin and heat-labile enterotoxin activate human monocyte-derived dendritic cells and dominantly inhibit cytokine production through a cyclic AMP-dependent pathway. Infect. Immun. 2002, 70, 5533–5539. [Google Scholar] [PubMed]
- George-Chandy, A.; Eriksson, K.; Lebens, M.; Nordström, I.; Schön, E.; Holmgren, J. Cholera toxin B subunit as a carrier molecule promotes antigen presentation and increases CD40 and CD86 expression on antigen-presenting cells. Infect. Immun. 2001, 69, 5716–5725. [Google Scholar] [PubMed]
- Porgador, A.; Staats, H.F.; Itoh, Y.; Kelsall, B.L. Intranasal immunization with cytotoxic T-lymphocyte epitope peptide and mucosal adjuvant cholera toxin: selective augmentation of peptide-presenting dendritic cells in nasal mucosa-associated lymphoid tissue. Infect. Immun. 1998, 66, 5876–5881. [Google Scholar] [PubMed]
- Anjuère, F.; Luci, C.; Lebens, M.; Rousseau, D.; Hervouet, C.; Milon, G.; Holmgren, J.; Ardavin, C.; Czerkinsky, C. In vivo adjuvant-induced mobilization and maturation of gut dendritic cells after oral administration of cholera toxin. J. Immunol. 2004, 173, 5103–5111. [Google Scholar] [PubMed]
- Gagliardi, M.C.; De Magistris, MT. Maturation of human dendritic cells induced by the adjuvant cholera toxin: role of cAMP on chemokine receptor expression. Vaccine 2003, 21, 856–861. [Google Scholar] [PubMed]
- Jank, T.; Giesemann, T.; Aktories, K. Rho-glucosylating Clostridium difficile toxins A and B: new insights into structure and function. Glycobiology 2007, 17, 15–22. [Google Scholar]
- Sandvig, K.; Spilsberg, B.; Lauvrak, S.U.; Torgersen, M.L.; Iversen, T.G.; van Deurs, B. Pathways followed by protein toxins into cells. Int. J. Med. Microbiol. 2004, 293, 483–490. [Google Scholar] [PubMed]
- Schirmer, J.; Aktories, K. Large clostridial cytotoxins: cellular biology of Rho/Ras-glucosylating toxins. Biochim. Biophys. Acta 2004, 1673, 66–74. [Google Scholar]
- Genth, H.; Dreger, S.C.; Huelsenbeck, J.; Just, I. Clostridium difficile toxins: more than mere inhibitors of Rho proteins. Int. J. Biochem. Cell. Biol. 2008, 40, 592–597. [Google Scholar] [PubMed]
- Van Hamme, E.; Dewerchin, H.L.; Cornelissen, E.; Verhasselt, B.; Nauwynck, H.J. Clathrin- and caveolae-independent entry of feline infectious peritonitis virus in monocytes depends on dynamin. J. Gen.Virol. 2008, 89, 2147–2156. [Google Scholar] [PubMed]
- Ausiello, C.M.; Cerquetti, M.; Fedele, G.; Spensieri, F.; Palazzo, R.; Nasso, M.; Frezza, S.; Mastrantonio, P. Surface layer proteins from Clostridium difficile induce inflammatory and regulatory cytokines in human monocytes and dendritic cells. Microbes Infect. 2006, 8, 2640–2646. [Google Scholar] [PubMed]
- Harokopakis, E.; Albzreh, M.H.; Martin, M.H.; Hajishengallis, G. TLR2 transmodulates monocyte adhesion and transmigration via Rac1- and PI3K-mediated inside-out signaling in response to Porphyromonas gingivalis fimbriae. J. Immunol. 2006, 176, 7645–7656. [Google Scholar] [PubMed]
- Methe, H.; Kim, J.O.; Kofler, S.; Nabauer, M.; Weis, M. Statins decrease Toll-like receptor 4 expression and downstream signaling in human CD14+ monocytes. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 1439–1445. [Google Scholar] [PubMed]
- Warny, M.; Keates, A.C.; Keates, S.; Castagliuolo, I.; Zacks, J.K.; Aboudola, S.; Qamar, A.; Pothoulakis, C.; LaMont, J.T.; Kelly, C.P. p38 MAP kinase activation by Clostridium difficile toxin A mediates monocyte necrosis, IL-8 production, and enteritis. J. Clin. Invest. 2000, 105, 1147–1156. [Google Scholar] [PubMed]
- Jefferson, K.K.; Smith, M.F., Jr.; Bobak, D.A. Roles of intracellular calcium and NF-kappa B in the Clostridium difficile toxin A-induced up-regulation and secretion of IL-8 from human monocytes. J. Immunol. 1999, 163, 5183–5191. [Google Scholar] [PubMed]
- Hmama, Z.; Knutson, K.L.; Herrera-Velit, P.; Nandan, D.; Reiner, N.E. Monocyte adherence induced by lipopolysaccharide involves CD14, LFA-1, and cytohesin-1. Regulation by Rho and phosphatidylinositol 3-kinase. J. Biol. Chem. 1999, 274, 1050–1057. [Google Scholar] [PubMed]
- Linevsky, J.K.; Pothoulakis, C.; Keates, S.; Warny, M.; Keates, A.C.; Lamont, J.T.; Kelly, C.P. IL-8 release and neutrophil activation by Clostridium difficile toxin-exposed human monocytes. Am. J. Physiol. 1997, 273, 1333–1340. [Google Scholar]
- Souza, M.H.; Melo-Filho, A.A.; Rocha, M.F.; Lyerly, D.M.; Cunha, F.Q.; Lima, A.A.; Ribeiro, R.A. The involvement of macrophage-derived tumour necrosis factor and lipoxygenase products on the neutrophil recruitment induced by Clostridium difficile toxin B. Immunology 1997, 91, 281–288. [Google Scholar] [PubMed]
- Siffert, J.C.; Baldacini, O.; Kuhry, J.G.; Wachsmann, D.; Benabdelmoumene, S.; Faradji, A.; Monteil, H.; Poindron, P. Effects of Clostridium difficile toxin B on human monocytes and macrophages: possible relationship with cytoskeletal rearrangement. Infect. Immun. 1993, 61, 1082–1090. [Google Scholar] [PubMed]
- Flegel, W.A.; Müller, F.; Däubener, W.; Fischer, H.G.; Hadding, U.; Northoff, H. Cytokine response by human monocytes to Clostridium difficile toxin A and toxin B. Infect. Immun. 1991, 59, 3659–3666. [Google Scholar] [PubMed]
- Kang, J.H.; Lee, G.S.; Jeung, E.B.; Yang, M.P. Trans-10, cis-12 conjugated linoleic acid modulates phagocytic responses of canine peripheral blood polymorphonuclear neutrophilic leukocytes exposed to Clostridium difficile toxin B. Vet. Immunol. Immunopathol. 2009, 130, 178–186. [Google Scholar] [PubMed]
- Lee, J.Y.; Kim, H.; Cha, M.Y.; Park, H.G.; Kim, Y.J.; Kim, I.Y.; Kim, J.M. Clostridium difficile toxin A promotes dendritic cell maturation and chemokine CXCL2 expression through p38, IKK, and the NF-kappaB signaling pathway. J. Mol. Med. 2009, 87, 169–180. [Google Scholar] [PubMed]
- El Bekay, R.; Alba, G.; Reyes, M.E.; Chacón, P.; Vega, A.; Martín-Nieto, J.; Jiménez, J.; Ramos, E.; Oliván, J.; Pintado, E.; Sobrino, F. Rac2 GTPase activation by angiotensin II is modulated by Ca2+/calcineurin and mitogen-activated protein kinases in human neutrophils. J. Mol. Endocrinol. 2007, 39, 351–363. [Google Scholar] [PubMed]
- Brito, G.A.; Sullivan, G.W.; Ciesla, W.P., Jr.; Carper, H.T.; Mandell, G.L.; Guerrant, R.L. Clostridium difficile toxin A alters in vitro-adherent neutrophil morphology and function. J. Infect. Dis. 2002, 185, 1297–1306. [Google Scholar] [PubMed]
- Servant, G.; Weiner, O.D.; Herzmark, P.; Balla, T.; Sedat, J.W.; Bourne, H.R. Polarization of chemoattractant receptor signaling during neutrophil chemotaxis. Science 2000, 287, 1037–1040. [Google Scholar] [PubMed]
- Däubener, W.; Leiser, E.; von Eichel-Streiber, C.; Hadding, U. Clostridium difficile toxins A and B inhibit human immune response in vitro. Infect. Immun. 1988, 56, 1107–1112. [Google Scholar] [PubMed]
- Dailey, D.C.; Kaiser, A.; Schloemer, R.H. Factors influencing the phagocytosis of Clostridium difficile by human polymorphonuclear leukocy tes. Infect. Immun. 1987, 55, 1541–1546. [Google Scholar] [PubMed]
- Yanagawa, Y.; Onoé, K. CCL19 induces rapid dendritic extension of murine dendritic cells. Blood 2002, 100, 1948–1956. [Google Scholar] [PubMed]
- Garrett, W.S.; Chen, L.M.; Kroschewski, R.; Ebersold, M.; Turley, S.; Trombetta, S.; Galán, J.E.; Mellman, I. Developmental control of endocytosis in dendritic cells by Cdc42. Cell 2000, 102, 325–334. [Google Scholar] [PubMed]
- West, M.A.; Prescott, A.R.; Eskelinen, E.L.; Ridley, A.J.; Watts, C. Rac is required for constitutive macropinocytosis by dendritic cells but does not control its downregulation. Curr. Biol. 2000, 10, 839–848. [Google Scholar] [PubMed]
- Malorni, W.; Paradisi, S.; Dupuis, M.L.; Fiorentini, C.; Ramoni, C. Enhancement of cell-mediated cytotoxicity by Clostridium difficile toxin A: an in vitro study. Toxicon 1991, 29, 417–428. [Google Scholar] [PubMed]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cubillos, S.; Norgauer, J.; Lehmann, K. Toxins–Useful Biochemical Tools for Leukocyte Research. Toxins 2010, 2, 428-452. https://doi.org/10.3390/toxins2040428
Cubillos S, Norgauer J, Lehmann K. Toxins–Useful Biochemical Tools for Leukocyte Research. Toxins. 2010; 2(4):428-452. https://doi.org/10.3390/toxins2040428
Chicago/Turabian StyleCubillos, Susana, Johannes Norgauer, and Katja Lehmann. 2010. "Toxins–Useful Biochemical Tools for Leukocyte Research" Toxins 2, no. 4: 428-452. https://doi.org/10.3390/toxins2040428