A Pilot Study of Nuclear Instability in Archived Renal and Upper Urinary Tract Tumours with Putative Ochratoxin Aetiology

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
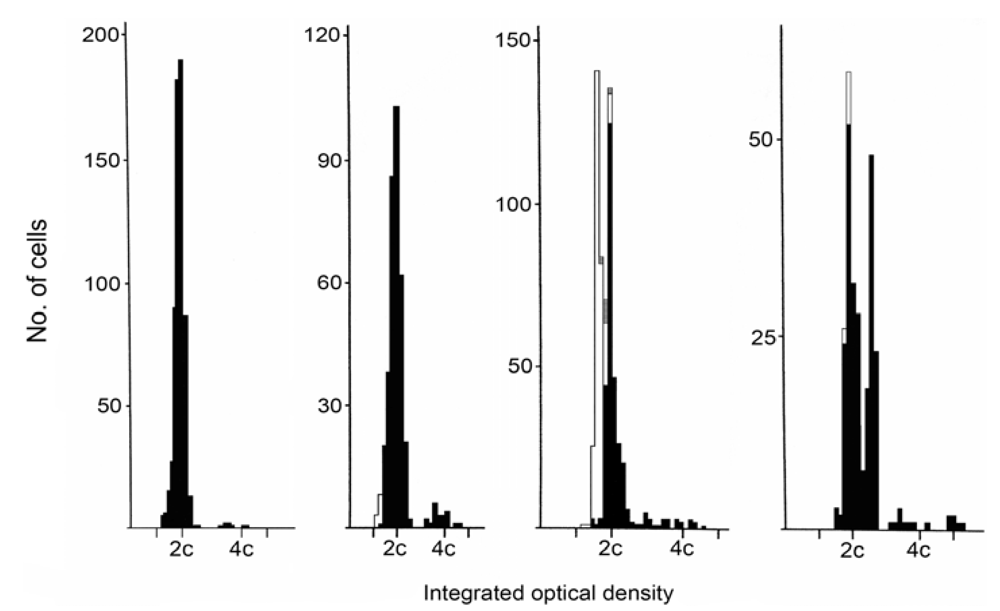
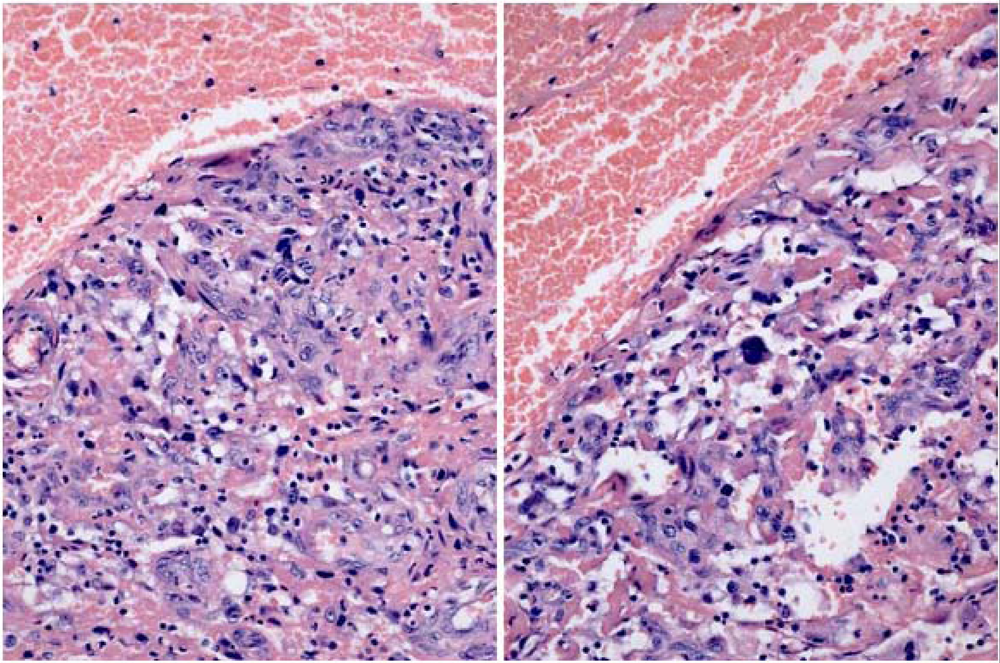
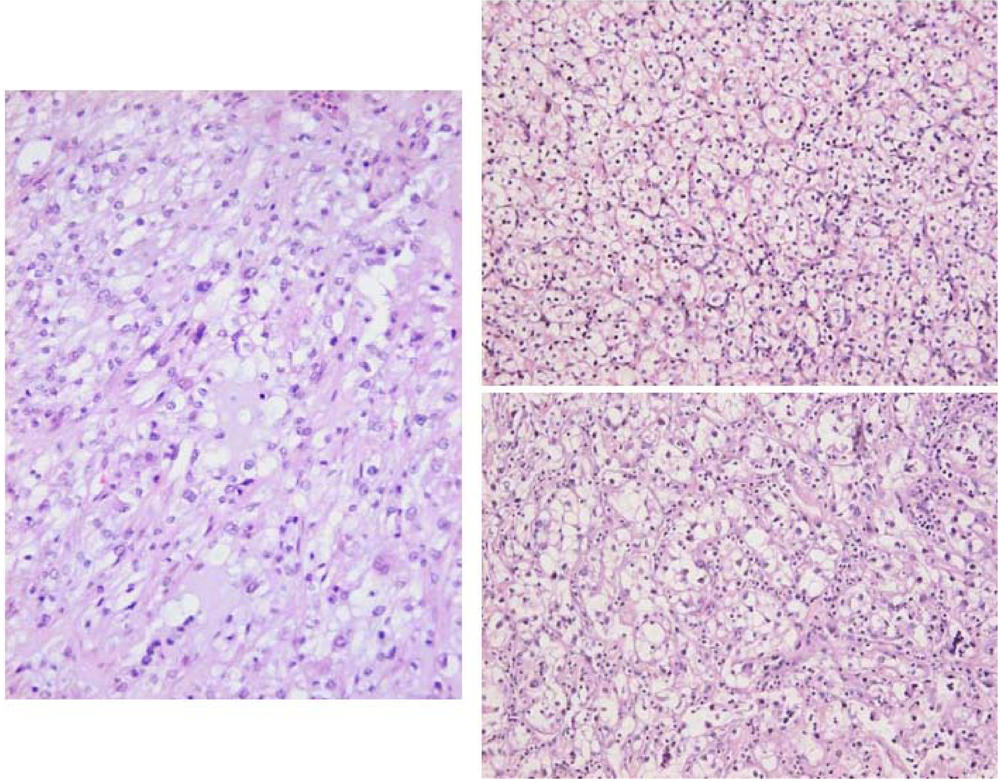
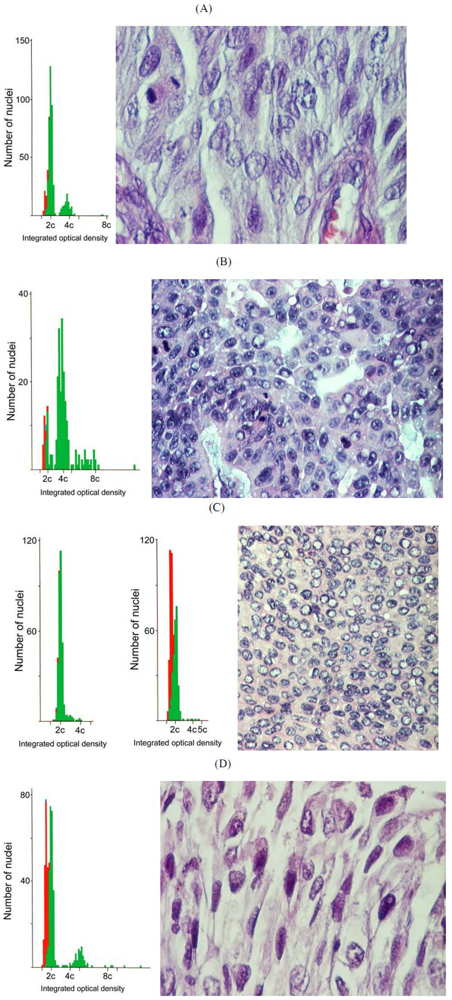
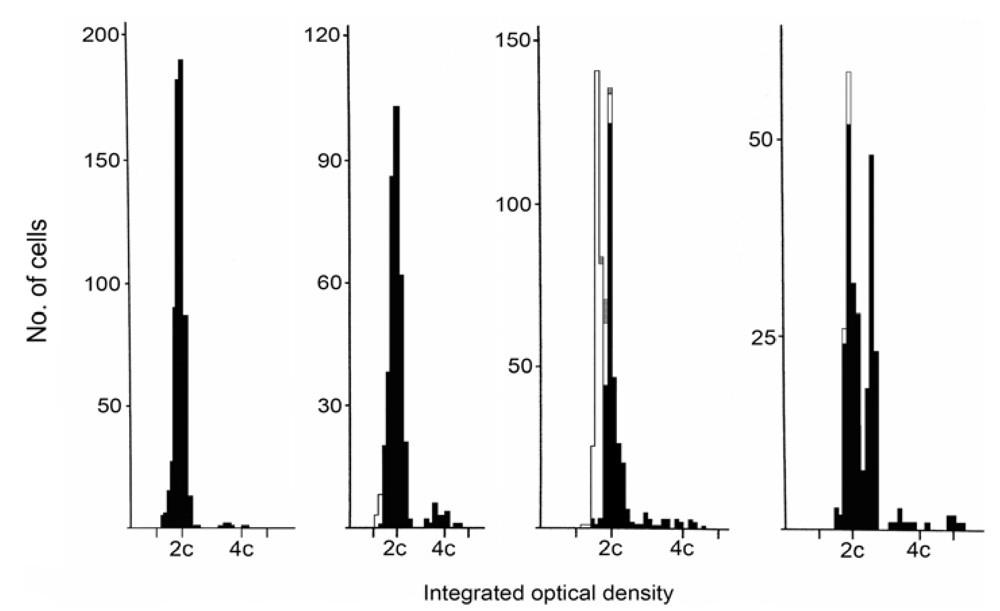
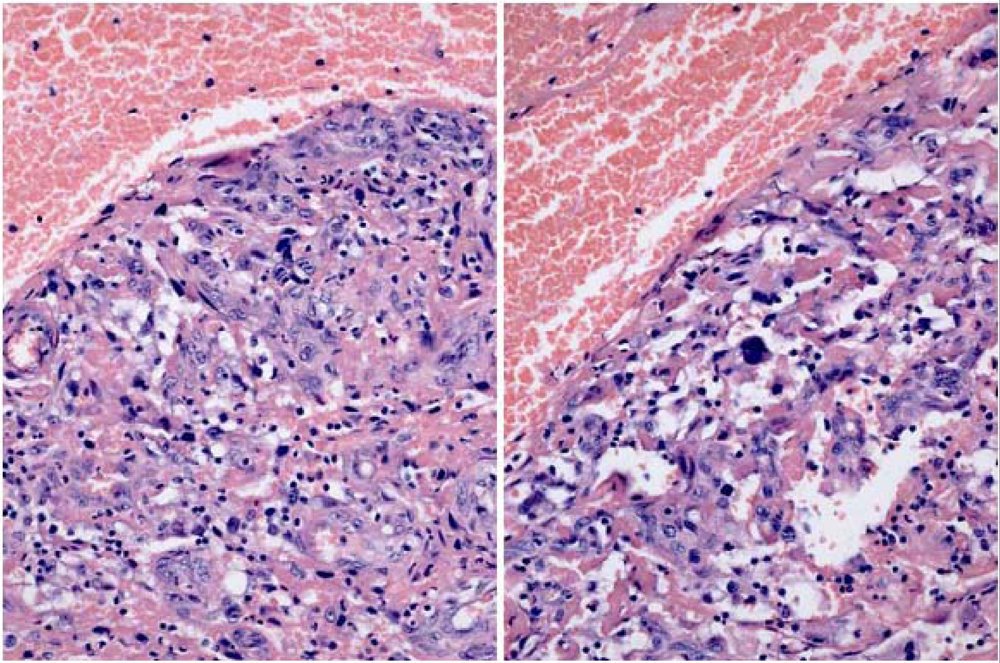
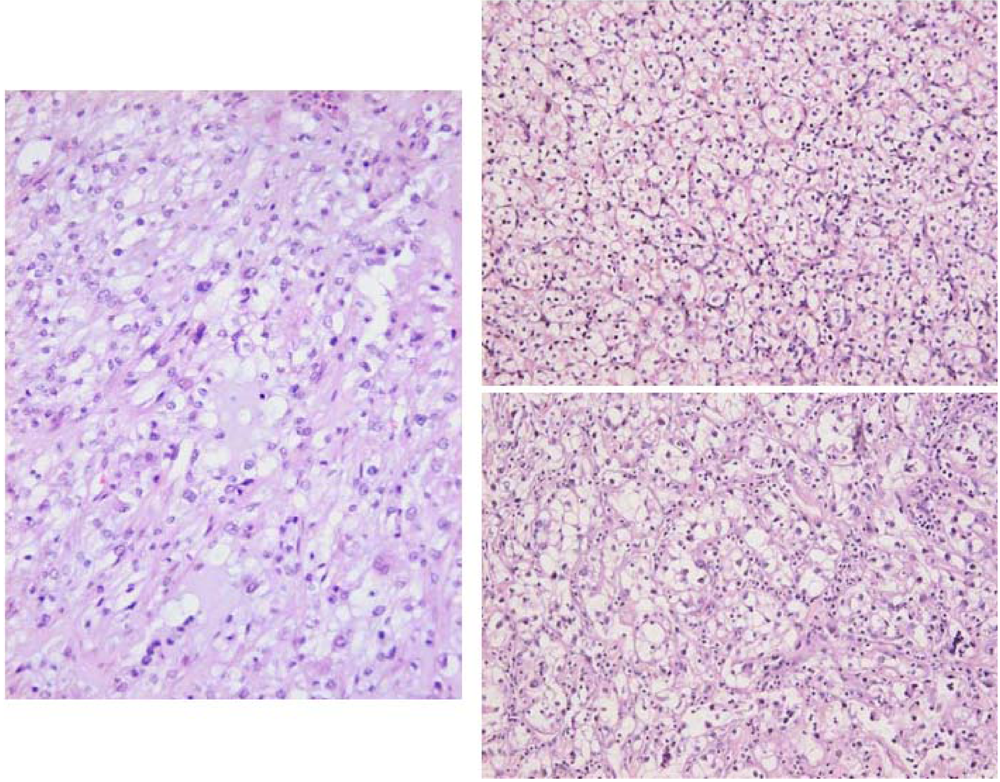
2. Results
3. Discussion
3.1. The renal cancer cases
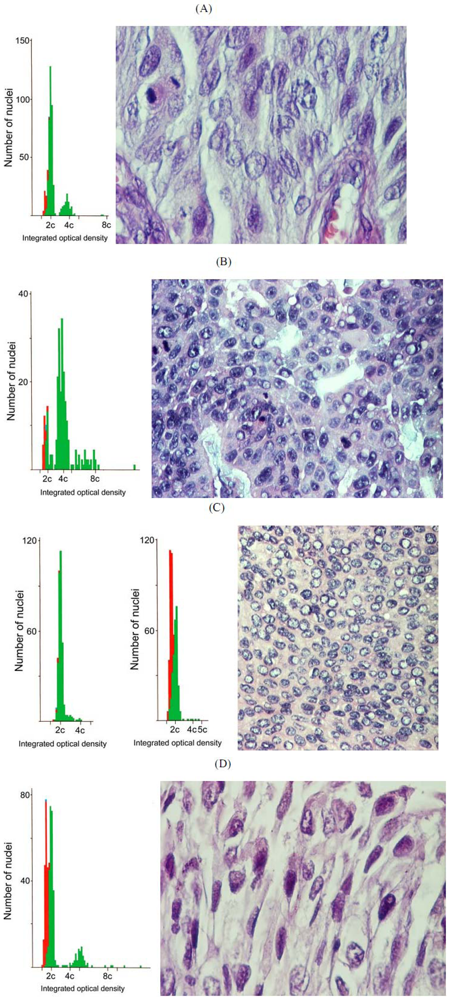
3.2. Balkan endemic nephropathy: The ochratoxin A hypothesis
4. Conclusions
5. Experimental Section
5.1. Ploidy analysis
5.2. Tissues relating to renal cell carcinoma

5.3. Romanian transitional cell tumours (TCC) from female BEN patients (2002-2004):
Acknowledgements
References
- van der Merwe, K.J.; Steyn, P.S.; Fourie, L.; Scott, D.B.; Theron, J.J. Ochratoxin A, a toxicmetabolite produced by Aspergillus ochraceus Wilh. Nature 1965, 205, 1112–1113. [Google Scholar]
- van der Merwe, K.J.; Steyn, P.S.; Fourie, L. Mycotoxins II. The constitution of ochratoxins A, B, and C, metabolites of Aspergillus ochraceus Wilh. J. Chem. Soc. Perkin 1 1965, 7083–7088. [Google Scholar] [PubMed]
- Krogh, P. Causal associations of mycotoxic nephropathy. Acta. Pathol. Microbiol. Scand. Sect A 1978, Suppl.269, 1–28. [Google Scholar]
- Krogh, P. Porcine Nephropathy Associated with Ochratoxin A. In Mycotoxins and Animal Foods; Smith, J.E., Henderson, R.S., Eds.; CRC Press: Boca Raton,FL,USA, 1991; pp. 627–645. [Google Scholar]
- Tanchev, Y.; Evstatiev, Z.; Dorossiev, D.; Pencheva, J.; Zvetkov, G. Studies on the nephritides in the District of Vratza (in Bulgarian). Sovrem. Med. 1956, 7, 14–29. [Google Scholar]
- Tanchev, Y.; Dorossiev, D. The first clinical description of Balkan endemic nephropathy (1956) and its validity 35 years later. In Mycotoxins, Endemic Nephropathy and Urinary Tract Tumours; International Agency for Research on Cancer: Lyon, France, 1991; Volume 115, pp. 21–28. [Google Scholar]
- WHO. Memoranda. The endemic nephropathy of South-Eastern Europe. WHO: Geneva, Switzerland, 1965; pp. 431–448. [Google Scholar]
- CIBA Foundation Study Group No 30. The Balkan Nephropathy. ; Wolstenholme, G.E.W., Knight, J., Eds.; J. and A. Churchill: London, 1967; p. 123. [Google Scholar]
- Austwick, P.K.C. Balkan Nephropathy. Proc. R. Soc. Med. 1975, 68, 219–221. [Google Scholar]
- Krogh, P. Mycotoxic porcine nephropathy- a possible model for Balkan (endemic) nephropathy. In Endemic nephropathy. In Endemic nephropathy, Proceedings of the second International Symposium on Endemic Nephropathy, 9–12 November 1974; Bulgarian Academy of Sciences: Sofia, Bulgaria; pp. 266–277.
- Krogh, P.; Hald, B.; Pedersen, E.J. Occurrence of ochratoxin A and citrinin in cereals associated with mycotoxic porcine nephropathy. Acta Pathol. Microbiol. Scand. Sect. A 1973, 81, 689–695. [Google Scholar]
- Krogh, P.; Axelsen, N.H.; Elling, F.; Gryd-Hansen, N.; Hald, B.; Hyldgaard-Jensen, J. ; Larsen, A.E.; Madsen, A.; Mortensen, H.P.; Moller, T; Petersen, O.K.; Ravnskov, U.; Rostgaard, M.; Aalund, O. Experimental porcine nephropathy: Changes of renal function and structure induced by ochratoxin A-contaminated feed. Acta. Pathol. Microbiol. Scand. Sect. A Suppl. 1974, 246, 1–21. [Google Scholar]
- MacGeorge, K.M.; Mantle, P.G. Nephrotoxic fungi in a Yugoslavian community in which Balkan nephropathy is hyperendemic. Mycol. Res. 1991, 95, 660–664. [Google Scholar] [CrossRef]
- Mantle, P.G.; McHugh, K.M. Nephrotoxic fungi in foods from nephropathy households in Bulgaria. Mycol. Res. 1993, 97, 205–212. [Google Scholar] [CrossRef]
- Cvetnic, Z.; Pepeljnjak, S. Ochratoxinogenicity of Aspergillus ochraceus strains from nephropathic and non-nephropathic areas in Yugoslavia. Mycopathologia 1990, 110, 93–99. [Google Scholar] [CrossRef]
- Stoev, S.D.; Hald, B.; Mantle, P.G. Porcine nephropathy in Bulgaria: A progressive syndrome of complex or uncertain (mycotoxin) aetiology. Vet. Rec. 1998, 142, 190–194. [Google Scholar] [CrossRef]
- Boorman, G.A. Toxicology and Carcinogenesis studies of ochratoxin A. In Toxicology and Carcinogenesis studies of ochratoxin A. NIH Publication No 89–2813 Research Triangle Park, NC, National Toxicology Programme; 1989. [Google Scholar]
- Lock, E.A.; Hard, G.C. Chemically induced renal tubule tumours in the laboratory rat and mouse: Review of the NCI/NTP database and categorization of renal carcinogens based on mechanistic information. Crit. Rev. Toxicol. 2004, 34, 211–299. [Google Scholar]
- Chernozemsky, I.N.; Stoyanov, I.S.; Petkova-Bocharova, T.K.; Nikolov, I.G.; Draganov, I.V.; Stoichev, I.; Tanchev, Y.; Naidenov, D.; Kalcheva, N.D. Geographic correlation between the occurrence of endemic nephropathy and urinary tract tumours in Vratza district. Int.J. Cancer 1977, 19, 1–11. [Google Scholar]
- Mantle, P.; Kulinskaya, E.; Nestler, S. Renal tumourigenesis in male rats in response to chronic dietary ochratoxin A. Food Add. Contam. 2005, 1, 58–64. [Google Scholar]
- Brown, A.L.; Odell, E.W.; Mantle, P.G. DNA ploidy distribution in renal tumours induced in male rats by dietary ochratoxin A. Exp. Toxicol. Pathol. 2007, 59, 85–95. [Google Scholar] [CrossRef]
- Diwakar, N.; Sperandio, M.; Sherrif, M.; Brown, A.; Odell, E.W. Heterogeneity, histological features and DNA ploidy in oral carcinoma by image-based analysis. Oral Oncol. 2005, 41, 416–422. [Google Scholar] [CrossRef]
- Hedley, D.W. DNA analysis from paraffin-embedded blocks. Methods Cell Biol. 1994, 41, 231–240. [Google Scholar] [CrossRef]
- Sherwood, B.; Burke, D. Metastatic Renal Cell Carcinoma: Novel therapies and health economics. Urology News 2009, 13, 7–10. [Google Scholar]
- Pinto, A.E.; Monteiro, P.; Silva, G.; Ayres, J.V.; Soares, J. Prognostic biomarkers in renal cell carcinoma; relevance of DNA ploidy in predicting disease-related survival. Int. J. Biol. Markers 2005, 20, 249–256. [Google Scholar] [PubMed]
- Li, G.; Cottier, M.; Sabido, O.; Gentil-Perret, A.; Lambert, C.; Passebosc-Faure, K.; Genin, C.; Tostain, J. The in vivo DNA aneuploidisation during expansion of conventional renal cell carcinoma. In Vivo 2002, 16, 341–344. [Google Scholar]
- Li, G.; Cottier, M.; Sabido, O.; Gentil-Perret, A.; Lambert, C.; Genin, C.; Tostain, J. Different DNA ploidy patterns for the differentiation of common subtypes of renal tumours. Cell. Oncol. 2005, 27, 51–56. [Google Scholar]
- Lang, H.; Lindner, V.; Letourneux, H.; Martin, M.; Saussine, C.; Jacqmin, D. Prognostic value of microscopic venous invasion in renal cell carcinoma: long-term follow-up. Eur. Urol. 2004, 46, 331–335. [Google Scholar] [CrossRef]
- Allen,, D.E. The Epidemiology of Renal Cell Carcinoma: Rates and Risk Factors. In The Effective Management of Renal Cell Carcinoma; Aitchison, M., Oliver, R.T.D., Miles, A., Eds.; Aesculapius Medical Press: London,UK, 2004; pp. 3–18. [Google Scholar]
- Marin-Kuan, M.; Nestler, S.; Verguet, C.; Bezencon, C.; Piguet, D.; Mansourian, R.; Holzwarth, J.; Grigorov, M.; Delatour, T.; Mantle, P.; Cavin, C.; Schilter, B. A toxicogenomics approach to identifying new plausible epigenetic mechanisms of ochratoxin A carcinogenicity in rat. Toxicol. Sci. 2006, 89, 120–134. [Google Scholar]
- Cavin, C.; Delatour, T.; Marin-Kuan, M.; Holzhauser, D.; Higgins, L.; Bezencon, C.; Guignard, G.; Junod, S.; Richoz-Payot, J.; Gremaud, E.; Hayes, J.D.; Nestler, S.; Mantle, P.; Schilter, B. Reduction in antioxidant defenses may contribute to ochratoxin a toxicity and carcinogenicity. Toxicol. Sci. 2007, 96, 30–39. [Google Scholar]
- Mally, A.; Pepe, G.; Ravoori, S.; Fiore, M.; Gupta, R.; Dekant, W.; Mosesso, P. Ochratoxin A causes DNA-damage and cytogenetic effects, but no DNA-adducts in rats. Chem. Res. Toxicol. 2005, 18, 1253–1261. [Google Scholar] [CrossRef] [Green Version]
- Mally, A.; Dekant, W. Mycotoxins and the kidney; Modes of action for renal tumor formation by ochratoxin A in rodents. Mol. Nutr. Food Res. 2009, 53, 467–478. [Google Scholar] [CrossRef] [PubMed]
- EFSA Opinion of the scientific panel on contaminants in the food chain on a request from the Commission related to ochratoxin A in food. EFSA J. 2006, 365, 1–56.
- IARC Ochratoxin A. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans 56; IARC Press: Lyon, France, 1993; pp. 489–521. [Google Scholar]
- Mantle, P.G. A reappraisal of ochratoxin A as a factor in human renal disease. In Occurrence and Significance of Mycotoxins; Scudamore., K., Ed.; Central Science Laboratory: Slough,UK, 1993; pp. 101–108. [Google Scholar]
- Radić, B.; Fuchs, R.; Peraica, M.; Lucić, A. Ochratoxin A in human sera in the area with endemic nephropathy in Croatia. Toxicol. Lett. 1997, 91, 105–109. [Google Scholar] [CrossRef]
- Pfohl-leszkowicz, A.; Grosse, Y.; Castegnaro, M.; Nikolov, I.G.; Chernozemsky, I.N.; Bartsch, H.; Betbeder, A.M.; Creppy, E.E.; Dirheimer, G. Ochratoxin A-related DNA adducts in urinary tract tumours of Bulgarian subjects. IARC Sci. Publ. 1993, 124, 141–148. [Google Scholar]
- Pfohl-Leszkowicz, A.; Tozlovanu, M.; Manderville, R.; Peraica, M.; Castegnaro, M.; Stefanovic, V. New molecular and field evidences for the implication of mycotoxins but not aristolochic acid in human nephropathy and urinary tract tumor. Mol. Nutr. Food Res. 2007, 51, 1131–1146. [Google Scholar]
- Turney, B.; Muneer, A. Upper tract urothelial tumours. Urology News 2007, 11, 20–21. [Google Scholar]
- Vrabcheva, T.; Usleber, E.; Dietrich, R.; Martlbauer, E. Co-occurrence of ochratoxin A and citrinin in cereals from Bulgarian villages with a history of Balkan Endemic Nephropathy. J. Agric. FoodChem. 2000, 48, 2483–2488. [Google Scholar]
- Scott, P.M. Biomarkers of human exposure to ochratoxin A. Food Add. Contam. 2005, 1, 99–107. [Google Scholar]
- Vrabcheva, T.; Petkova-Bocharova, T.; Grosso, F.; Nikolov, I.; Chernozemsky, I.N.; Castegnaro, M.; Dragacci, S. Analysis of ochratoxin A in foods consumed by inhabitants from an area with Balkan endemic nephropathy: A 1 month follow-up study. J. Agric. Food Chem. 2004, 52, 2404–2410. [Google Scholar]
- Abouzeid, M.M.; Horvath, A.D.; Podlesny, P.M.; Regina, D.P.; Metodiev, V.D.; Kamenova-Tozeva, R.M.; Niagolova, N.D.; Stein, A.D.; Petropoulos, E.A.; Ganev, V.S. Ochratoxin A concentrations in food and feed from a region with Balkan endemic nephropathy. Food Add. Contam. 2002, 19, 755–764. [Google Scholar] [CrossRef]
- Pitt, J.I. The Genus Penicillium and Its Teleomorphic States Eupenicillium and Talaromyces; Academic Press: London,UK, 1979. [Google Scholar]
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mantle, P.G.; Amerasinghe, C.; Brown, A.L.; Herman, D.; Horn, T.; Krogh, T.; Odell, E.W.; Rosenbaum, T.; Tatu, C.A. A Pilot Study of Nuclear Instability in Archived Renal and Upper Urinary Tract Tumours with Putative Ochratoxin Aetiology. Toxins 2010, 2, 326-340. https://doi.org/10.3390/toxins2030326
Mantle PG, Amerasinghe C, Brown AL, Herman D, Horn T, Krogh T, Odell EW, Rosenbaum T, Tatu CA. A Pilot Study of Nuclear Instability in Archived Renal and Upper Urinary Tract Tumours with Putative Ochratoxin Aetiology. Toxins. 2010; 2(3):326-340. https://doi.org/10.3390/toxins2030326
Chicago/Turabian StyleMantle, Peter G., Cyrille Amerasinghe, Amy L. Brown, Diana Herman, Thomas Horn, Thoger Krogh, Edward W. Odell, Tomas Rosenbaum, and Calin A. Tatu. 2010. "A Pilot Study of Nuclear Instability in Archived Renal and Upper Urinary Tract Tumours with Putative Ochratoxin Aetiology" Toxins 2, no. 3: 326-340. https://doi.org/10.3390/toxins2030326