1. Introduction
Members of the
Chlamydiaceae family can cause infections in all animal kingdoms [
1]. After the recent reclassification of the
Chlamydiae phylum, the
Chlamydiaceae family has been divided into two genus,
Chlamydia and
Chlamydophila [
2]. The two genus contain nine species and four of them,
Chlamydia trachomatis,
Chlamydophila pneumoniae,
Chlamydophila psittaci and
Chlamydophila abortus, can cause infections in human [
3]. They are Gram-negative bacteria with intracellular growth. Among them,
C. abortus is identified as the Serovar 1 of
Chlamydia psittaci [
2], which infects epithelial mucosal cells responsible for chlamydioses in a number of animal species.
Infections with
C. abortus cause abortions in sheep and goats. This bacteria, when transmitted to humans, can equally be responsible for abortions [
4,
5]. It has been difficult to generate a vaccine against
C. abortus infection, since this intracellular obligate bacteria is difficult to obtain in large quantities.
The POMP91B protein belongs to a group of four proteins, named Polymorphic Outer Membrane Proteins (POMPs), of approximately 90 kDa, present in the outer membrane of
C. abortus [
6]. Although they are present in small amounts in the bacteria, these proteins are powerfully immunogenic, as confirmed by serum analysis after abortion in sheep [
7], or after injection of live bacteria to mice [
8]. POMP91B possesses an
N-terminal extracellular exposed region [
7], responsible for the strong response in sheep which supported its use in an ELISA detection test [
9,
10]. The structure of this
N-terminal part is composed of a Beta-barrel [
11].
Pathogen Associated Molecular Patterns (PAMPs) are expressed only by micro-organisms and are recognized by eukaryotic cells through the Pattern Recognition Receptors (PRRs) of the innate immune system, such as the Toll-like receptors (TLRs) [
12]. The interaction of PAMPs with their corresponding TLRs helps to identify the nature of the PAMP and to guide the adequate adaptive immune response [
12]. Muramyl dipeptides, a major element of Freund's complete adjuvant, bacterial DNA, and flagellin are three PAMPs recognized by TLR2, TLR9, and TLR5, respectively. Freund’s adjuvant is usually used with high efficiency to amplify the immune response in experimental animals [
13]. DNA technology has been used successfully in the vaccination of animal models against infection with viruses, bacteria, and parasites as well as in anti-tumor therapy, allergies and treatment of autoimmune diseases [
14].
Salmonella flagellin was also used with partial success as an adjuvant and a carrier for a synthetic peptide representing a neutralizing epitope of influenza virus [
15], and to produce antibodies against cholera toxin epitope [
16]. The antibody response, systemic (IgG) or secreted (IgA), against flagellin was dependent on the route of immunization [
17] and finally, immunization of mice with a fusion protein containing a flagellin-enhanced green protein was capable of stimulating antigen presenting cells and thus develop a specific T-cell response [
18]. We previously used this strategy and obtained a good Th1 response against two different bacterial antigens [
19,
20].
The extracellular localization of the
N-part of the POMP91B protein [
7], as well as the other described POMPs proteins led to the belief that they might play an important immunological role either in diagnosis or in protection. In the present report, we examined the immune response against the
N-terminal region of the POMP91B protein using different PAMPs as adjuvants and vectors for immunization and compared it with the responses obtained with P27 antigen of tuberculosis. A DNA plasmid encoding this region of POMP91B protein (DNA immunization) was prepared and its immunogenicity was compared with that of the recombinant protein (classical immunization) and with that of a recombinant fusion protein carried by the flagellar filament of bacteria
E. coli (flagellin immunization).
Here, we show that the administration of the recombinant POMP91B protein induces the strongest Th2-like response, as indicated by IL-4 and antibody production. In comparison with two other antigens, immunization with the bacteria expressing the partial POMB91B protein into their flagella is not able to induce a strong Th1‑like response, as indicated by the weak cell proliferation and IFN-γ production. The DNA immunization was unable to induce an antigen-specific immune response.
Regarding the other antigens, the N-terminal part of the C. abortus POMP91B stimulates a strong Th2 response profile when the immunizations are done with the recombinant protein which does not require Freund’s adjuvant to present a good IL-4 and antibody response. In addition, this antigen is weaker in inducing a Th1 response, regardless of the immunization method used.
3. Discussion
Two conclusions may be drawn from our results; first the difference between the two immune responses (i.e., Th1 vs. Th2) is determined by the nature of antigen, and the second concerns the difficulty in obtaining a cellular immune response against the POMP91B antigen of the obligateintracellular growth pathogen Chlamydophila abortus.
Using different PAMPs including bacterial flagellin which is recognized by TLR5, Freund’s adjuvant which contains muramyl dipeptides and lipoarabinomannans, the ligand of the TLR2, and the CpG DNA recognized by the TLR9, we studied the immune response against the
N-terminal part of the POMP91B protein of
Chlamydophila abortus,which was found to be strongly immunogenic [
7]. Then we compared them with those obtained in the same conditions with the p27 PPE antigen, a cell surface protein from
M. tuberculosis [
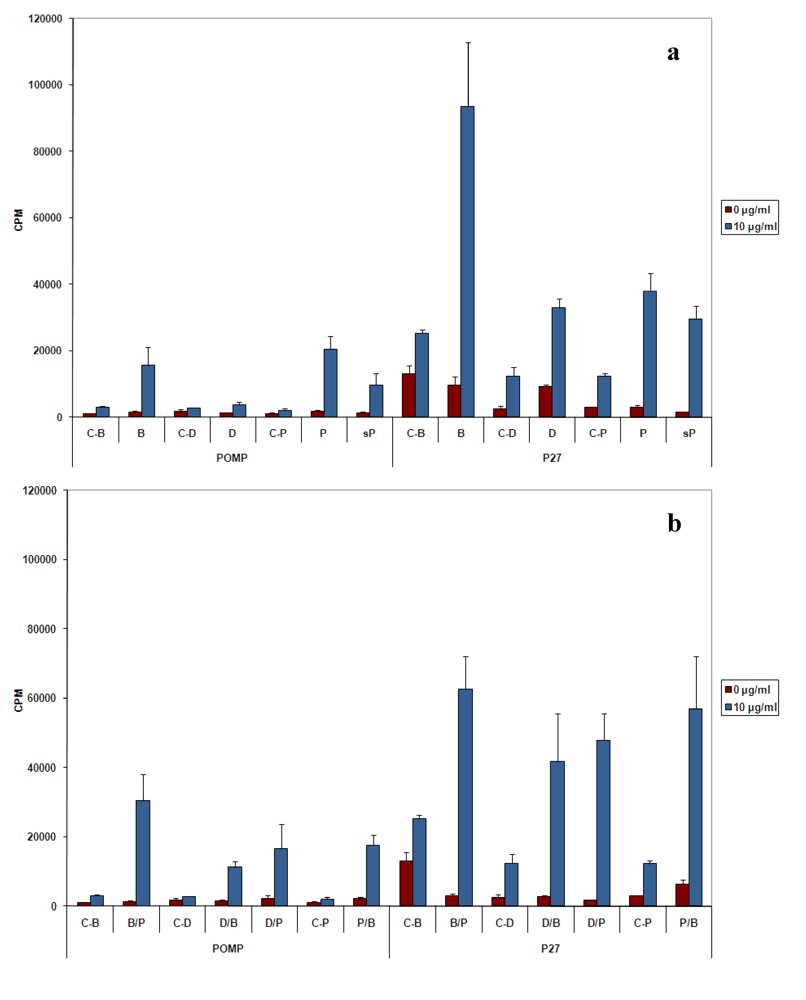
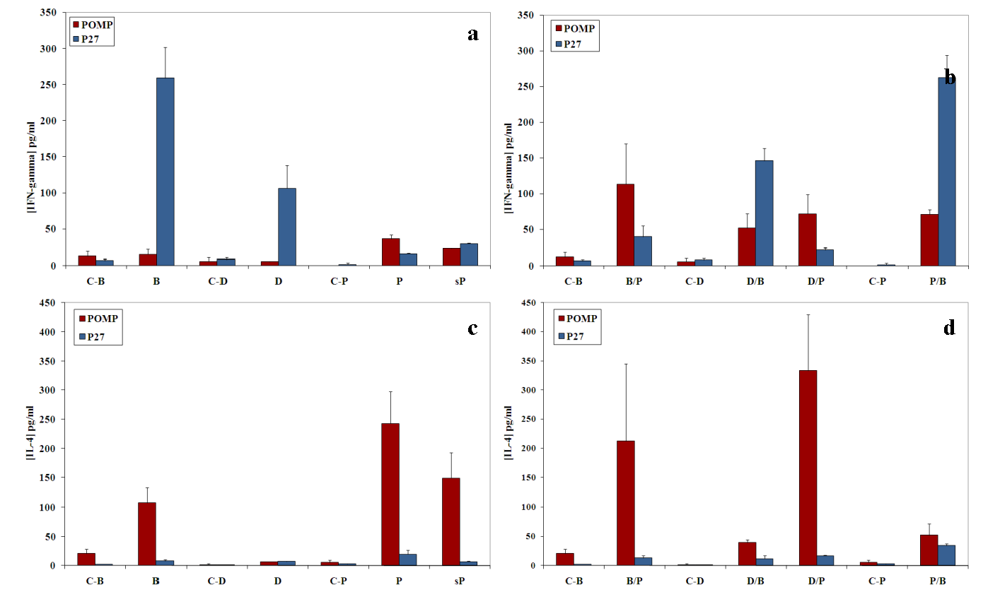
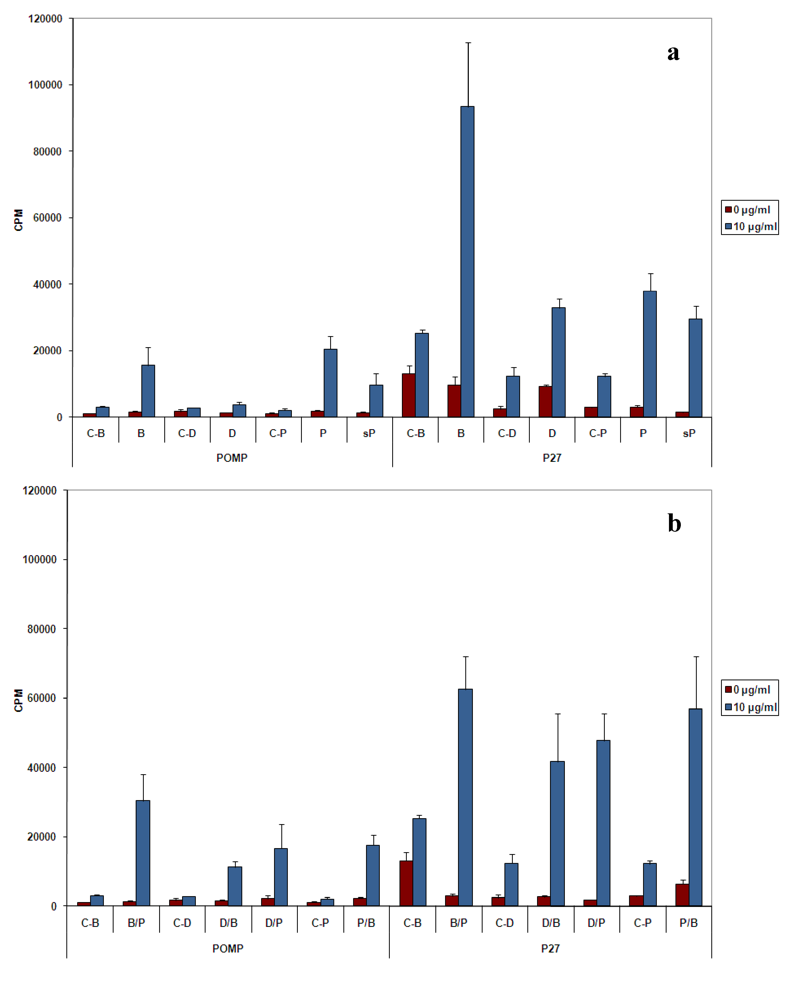
20]. Unlike the p27 PPE antigen that induced a strong proliferative response accompanied by high production of IFN‑γ and low amount of IL-4, the POMP91B antigen induced in general a Th2-like response, with weak proliferation of cells, low production of IFN-γ and high levels of IL-4. This is particularly true when immunizations are practised with the recombinant protein (
Figure 2 and
Figure 3). The results indicated that this antigen may be involved in the establishment of the host humoral immune response against
C. abortus. However, variations were observed in the intensity of the responses depending on the method of immunization used. While with p27 the strongest proliferative response of splenic cells in our experimental conditions was obtained utilizing the flagellin immunization, POMP91B antigen was not able to induce such a strong response and a better response was only obtained by immunization with the recombinant POMP91B antigen with incomplete Freund’s adjuvant after priming with flagellin-modified bacteria. In addition, the highest amount of IFN‑γ production was found in cells derived from mice immunized using the same combination. It is interesting to note that for POMP91B, as for p27, the group that showed the best cell proliferation was also producing the highest amount of IFN-γ. IFN‑γ is one of the cytokine known to be important in promoting a Th1 response and enhancing cytotoxicity during the priming and maturation of lymphocytes [
22].
In our experiments, the proliferation response induced by DNA immunization was the lowest and the production of IFN‑γ was not significantly different from the control. The pcDNA3 plasmid used as the immunization vector contains two GACpGTT motifs, which are known to induce Th1 cytokines in mice [
23,
24]. However, a synergistic effect was observed between DNA vaccination and flagellin expression vector [
25].
The results with POMP91B are also different from our previous observation concerning the immune response to
Clostridium tyrobutyricum flagellin, in which a good cellular and antibody response were obtained using the flagellin as carrier and adjuvant [
19]. The absence of IFN-γ production using the POMP91B antigen and the dominance of Th2 like response were also observed with another antigen, the Major Outer Membrane Protein (MOMP) of
C. trachomatis using another immunization system in which murine dentritic cells were pulsed with the antigen [
26]. These observations showing that surface proteins of
Chlamydia genus do not seem to induce a CD4+ Type1 response could be the reason for the failure to produce the needed vaccine to prevent infections. It is evident that IFN-γ is an important effector cytokine, particularly in the clearance of infections caused by human strains [
27,
28].
Despite the absence of a strong cellular response with POMP91B antigen, the role of flagellin in the induction of cellular responses could be clearly demonstrated by the use of combined immunizations. Cells isolated from mice primed by the antigen presented on the modified flagellin and boosted with protein in IFA released the highest amount of IFN‑γ [
Figure 3(b)].
In conclusion, we found that the administration of recombinant bacteria carrying the P27 PPE
M. tuberculosis antigen resulted in the induction of Th1 response. This kind of response may play a key role in protection against intracellular pathogens. Unfortunately, this immunization did not give such a good response with the POMP91B
C. abortus antigen; however, both are cell surface exposed bacterial antigens. These results suggest that the nature of the antigen could influence the Th1-Th2 balance of the immune response. Only the three-dimensional structure of the extracellular
N‑terminal part of POMP91B was determined and it represented a compact beta-helical domain with six rungs [
11]. Combined immunization can be useful to obtain a global response (antibody and cytokine production). Indeed using POMP91B antigen, the best global response was observed in the group primed by flagellin-modified bacteria that was then boosted by the recombinant protein in incomplete Freund’s adjuvant [
Figure 2(b),
Figure 3(b),
Figure 3(d) and
Figure 4(d)].
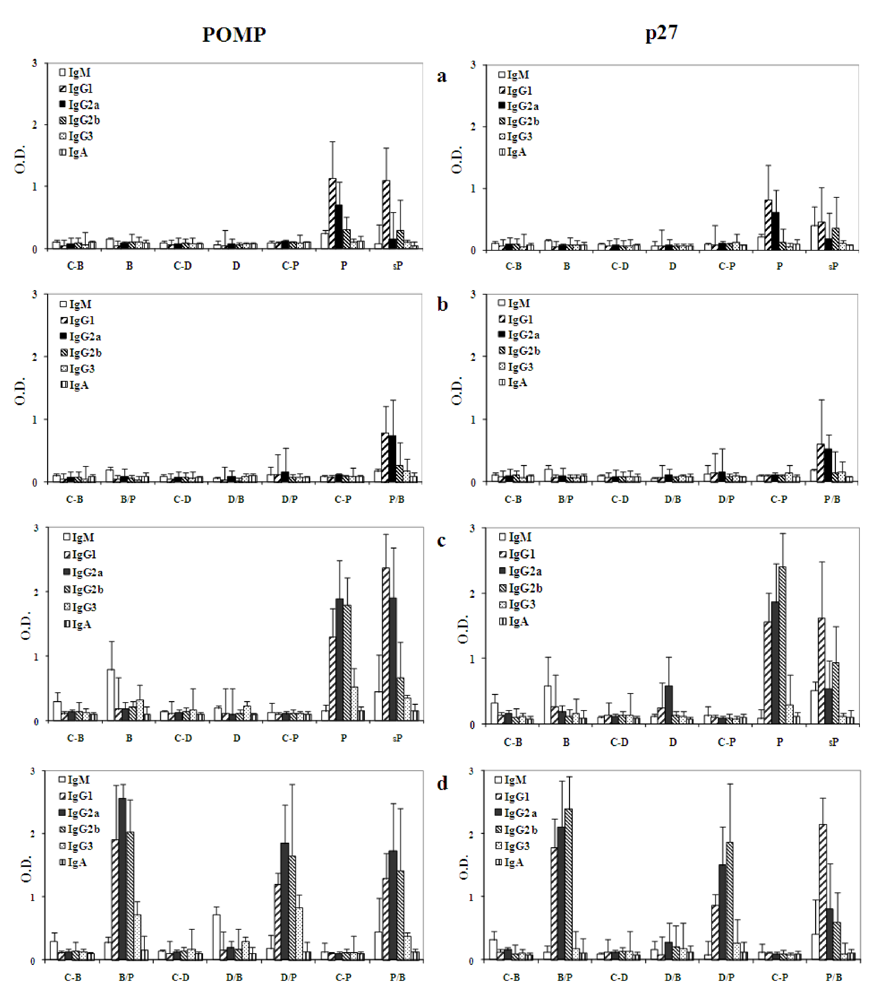
Concerning the humoral response to POMP91B in recombinant protein form with Freund’s adjuvant, the fact that the major antibody isotype primed is IgG2a, is quite surprising since this isotype is promoted by a Th1-like response. However, when the POMP91B antigen is injected in saline (sP) without any adjuvant, the IgG1 isotype of the Th2-like response is the one showing the strongest intensity. Here again, the study of the antibody responses confirmed our previous observations showing that the flagellin and the DNA system of immunization were unable to induce a good antibody response. In contrast, the optimal response resulted from the classical method using Freund’s adjuvant. For the flagellin immunization, the major isotype found in the sera was the IgM. The interaction of flagellin with the immune cells in our experiments failed to lead to an isotype switch.
4. Experimental Section
4.1. Bacterial strains, plasmid and antigens preparation
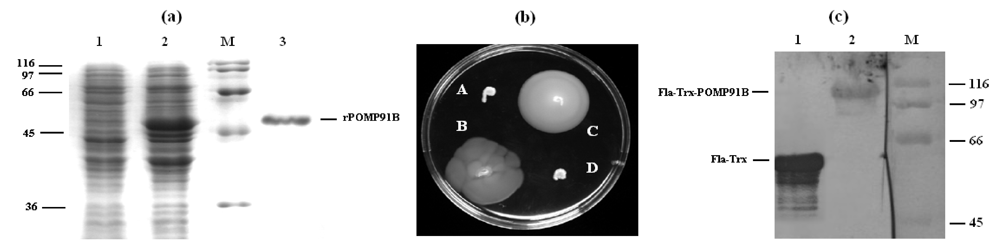
Escherichia coli strains DH5a (Invitrogen, San Diego, CA, USA) and BL21 (DE3) (Novagen, San Diego, CA, USA) were used for cloning and overexpression of the recombinant protein, respectively. For the production of recombinant protein, the 5’part of the pomp91B gene was amplified by PCR from an original template plasmid provided by Dr. Olivier Grépinet (INRA Tours, Nouzilly, France) using oligonucleotide pair 5’-GGG AAT TCC ATA TGA AAC ATC CAG TCT ACT GGT T-3’ and 5’-CGC GGA TCC GGA CGG TGT TTG TAA CGT A-3’. The oligonucleotide primers (Eurogentec, Seraing, Belgium) contained a NdeI and a BamHI site (underlined) respectively. PCR products were purified, digested by the two restriction enzymes NdeI and BamHI and ligated during 16 h at 4 °C with T4 ligase (Eurogentec) into the expression vector pET15b (Novagen) pre-digested with NdeI and BamHI. This plasmid encodes six histidines tagged to the N-terminus of the expressed recombinant protein to be purified. Expression of the recombinant protein was induced by adding 1 mM isopropyl-b-D-thiogalactopyranoside (IPTG) to BL21(DE3) bacteria (OD600 = 0.6) carrying the recombinant pET-15b-pomp91b plasmid in 2 × TY medium at 28 °C for 3 to 4 hours. The bacteria were harvested by centrifugation, resuspended and lysed by incubation for 1 hour in lysis buffer (100 mM Na2HPO4, 10 mM Tris-HCl, 8 M urea, pH 8.0). After centrifugation (10,000 g, 30 min), the supernatant was added to a Ni-NTA resin (Qiagen, Hilden, Germany). The mixture was incubated for 1 h at room temperature. After two washes with the same buffer, recombinant proteins were eluted with urea 8 M at pH of 5.9, then 4.5. The purified recombinant proteins were renatured by dialysis against PBS pH 7.4 at 4 °C, quantified by a Bradford assay, filtered, and stored in the same buffer at ‑20 °C.
For DNA immunization, the gene encoding the N-part of the POMP91B protein was cloned in the eukaryotic expression vector pcDNA3 (Invitrogen). Plasmid DNA was amplified in E. coli DH5a, purified by Qiagen purification kit, dissolved in phosphate-buffered saline (PBS), and stored at ‑20 °C until use.
For immunizations with the bacteria carrying the POMP91B protein in their flagellin FliC protein, a plasmid (pFliTrx) coding for
fliC gene and a specific
E. coli flagellin-deficient strain were used as described [
21]. The plasmid pFliTrx, carries the incomplete
E. coli fliC coding sequence in which the coding region for amino-acid residues 244-351 of the
E. coli flagellin has been replaced, in frame, with the entire coding sequence for the
E. coli thioredoxin (
trxA) gene. The partial
pomp91b gene was cloned in the
trxA gene at the unique
RsrII restriction site. The two primers used (5’-GGG GG
G GTC CGA AAC ATC CAG TCT ACT GGT-3’ and 5’-GGG GG
G GAC CGG ACG GTG TTT GTA ACG TA-3’) have an
AvaII restriction site (underlined) allowing a directional cloning. Plasmid constructs were then used to transform the non-motile
E. coli GI809 strain [
21], which carries a specific 512 bp deletion within the flagellin gene (
fliC, GenBank accession #M14358). Transformed motile bacteria were assessed for motility by swarming assay and by western blotting using specific antibodies, before preparation of immunization solutions.
4.2. Swarming assays
Non-motile E. coli GI809 cells were transformed by a flagellin containing pFliTrx plasmid and placed in a Petri dish containing IMC semi-gelosed medium with an agar specific for motility (Difco, Detroit, MI, USA) at 0.3%. The incubation was realized at 28 °C and the motility was observed at least 15 h after the start of the culture.
4.3. SDS-PAGE and western blot analysis
Purified proteins and bacterial extracts were heated 5 min at 100 °C in the lysis buffer (50 mM Tris, pH 8.0, 150 mM NaCl, 4% SDS, 5% b-mercaptoethanol) and electrophoresed through a 12.5% polyacrylamide gel, transferred to nitrocellulose membranes, then blotted with anti-POMP91B or anti-thioredoxin antibodies as described in Le Moigne
et al. [
19].
4.4. Animals and immunization
Adult female BALB/c mice (Janvier, Le Genest, France) were divided in groups of six mice and immunized either with 50 mg of plasmid DNA carrying the antigen coding sequence, in 100 mL PBS, three times with one month interval in the quadriceps muscle (DNA immunization), with 25 mg of antigen emulsified in complete Freund's adjuvant (CFA, containing the H37Ra, ATCC 25177
M. tuberculosis strain) for the first immunization or incomplete Freund's adjuvant (IFA) for the two following immunizations, by a subcutaneous route (Classical immunization), with 50 mg of antigen in PBS (Soluble antigen) or with 10
8 modified
E. coli bacteria which express the antigen into the flagellin at the surface of the bacteria given intraperitonneally (Flagellin immunization). Control mice for DNA immunization were immunized with empty pcDNA3 plasmid, whereas control mice for classical immunization and soluble antigen immunization were immunized with PBS incorporated in Freund's adjuvant or PBS alone, respectively. For the flagellin immunization, control mice were immunized with non modified
E. coli (bacteria which express normal flagellin). Combined immunizations between the different methods were applied as in
Table 1.
Table 1.
Group’s name in function of the different immunization practiced *.
Table 1.
Group’s name in function of the different immunization practiced *.
| | Method of immunization |
|---|
| | Classical Immunization. | DNA Immunization | Flagellin Immunization |
| First immunization | Soluble protein | Protein + CFA | Eukaryotic plasmid | Modified bacteria |
| Following Immunizations | Soluble protein | Protein + IFA | Bacteria | Protein + IFA | Bacteria | Plasmid | Protein + IFA | Bacteria |
| Group’s name | sP | P/P | P/B | D/P | D/B | D/D | B/P | B/B |
Blood samples were collected one week after the last immunization from the retro‑orbital plexus and sera were stored at −20 °C until use. All animal experiments were done in accordance with institutional and national ethical guidelines.
4.5. Enzyme-linked immunosorbent assay (ELISA)
Plates coated overnight at 4 °C with 1 µg/mL of recombinant proteins in 100 µL of carbonate-bicarbonate buffer (0.1 M, pH 9.6), were washed with phosphate-buffered saline-Tween 20 (PBS-T) (0.05%; v/v) and blocked for one hour at 37 °C with PBS-T containing 0.5% gelatin (PBS-T-G). Then serum dilutions (1/500) in the same buffer were added. After 1 h of incubation at 37 °C and four washes, goat anti-mouse IgM, IgG1, IgG2a, IgG2b, IgG3 and IgA alkaline phosphatase-conjugate (Southern biotechnology, Birmingham, AL, USA) were added and the plates were incubated for another 1 h at 37 °C. After four washes, 100 µL of 1 mg/mL of p-nitrophenylphosphate (Sigma, Saint Quentin Fallavier, France) in diethanolamine buffer (pH 9.8) was added, and then plates were read at 405 nm with a Titertek Multiscan instrument (Skatron, Oslo, Norway).
4.6. Proliferation assay
To check the effect of antigen boosting in our system, only half of the mice were boosted intraperitonneally with 25 mg of soluble antigen in PBS six months after the first immunization. One week later, splenic cells (5 × 105 cells/well) from antigen-boosted or non-boosted mice were suspended in RPMI-10% (FCS) and plated in 96-well flat bottomed plates containing serial dilution of antigen. Two days later, 0.5 µCi of [3H] thymidine was added to each well and incubated for an additional 18 hours, then cells were harvested and thymidine incorporation was measured using a liquid scintillation spectrophotometer Packard 1600 Tricarb (Packard, Downers Grove, IL, USA).
4.7. Cytokine production and quantification by ELISA
Splenic cells (5 × 105 cells) from boosted, non-boosted and control mice were cultured in RPMI-10% fetal calf serum (FCS) at 37 °C in 5% CO2 in 96-well plates containing different concentrations: 0, 1.1, 3.3 and 10 µg of recombinant proteins per mL. Supernatants were harvested after 48 h and one week of incubation and stored at ‑20 °C until use. IL-4 and IFN‑γ contents were measured with a commercial ELISA kit (Bender MedSystems, Vienna, Austria) according to the manufacturer’s protocol.


{kind=link}
{kind=link}
{kind=link}
{kind=link}