Food-Derived Antioxidant Polysaccharides and Their Pharmacological Potential in Neurodegenerative Diseases
by

Haifeng Li
1,2,†, 
Fei Ding
1,†,
Lingyun Xiao
2,
Ruona Shi
1,
Hongyu Wang
1,
Wenjing Han
1 and
Zebo Huang
1,* 1
Center for Bioresources & Drug Discovery and School of Biosciences & Biopharmaceutics, Guangdong Pharmaceutical University, Guangzhou 510006, China
2
School of Pharmaceutical Sciences, Wuhan University, Wuhan 430071, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Nutrients 2017, 9(7), 778; https://doi.org/10.3390/nu9070778
Submission received: 31 May 2017
/
Revised: 10 July 2017
/
Accepted: 13 July 2017
/
Published: 19 July 2017
(This article belongs to the Special Issue Antioxidants in Health and Disease)
Abstract
:Oxidative stress is known to impair architecture and function of cells, which may lead to various chronic diseases, and therefore therapeutic and nutritional interventions to reduce oxidative damages represent a viable strategy in the amelioration of oxidative stress-related disorders, including neurodegenerative diseases. Over the past decade, a variety of natural polysaccharides from functional and medicinal foods have attracted great interest due to their antioxidant functions such as scavenging free radicals and reducing oxidative damages. Interestingly, these antioxidant polysaccharides are also found to attenuate neuronal damages and alleviate cognitive and motor decline in a range of neurodegenerative models. It has recently been established that the neuroprotective mechanisms of polysaccharides are related to oxidative stress-related pathways, including mitochondrial function, antioxidant defense system and pathogenic protein aggregation. Here, we first summarize the current status of antioxidant function of food-derived polysaccharides and then attempt to appraise their anti-neurodegeneration activities.

1. Introduction
Oxygen is essential for normal life of aerobic organisms. Due to its high redox potential, oxygen is inevitably involved in the production of reactive oxygen species (ROS) such as superoxide anion, hydroxyl radical and hydrogen peroxide. ROS are known to play an important role in a variety of cellular functions including signal transduction and regulation of enzyme activity [1,2]. Excessive ROS, on the other hand, can also interact with biological molecules and generate by-products such as peroxides and aldehydes, which can cause damages to architecture and function of cells [3,4]. Under normal circumstances, cells have a set of antioxidant defense system, including enzymatic antioxidants such as superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx) and non-enzymatic antioxidants such as glutathione and vitamins, to combat excessive ROS [5,6]. However, when a detrimental stress compromises the antioxidant defense system, a fraction of ROS may escape the intrinsic clearance machinery and induce a state of oxidative stress, leading to cell dysfunction [7,8].
Growing evidence has demonstrated that oxidative stress is implicated in the development and progression of many chronic diseases such as neurodegenerative diseases (NDD) and diabetes [9,10,11]. NDD, including Alzheimer’s disease (AD), Parkinson’s disease (PD) and Huntington’s disease (HD), are a group of chronic disorders pathologically characterized by selective and progressive loss of neurons [12]. Clinical evidence has shown that NDD patients display an oxidative stress-related manifestation, including increases of ROS level, lipid peroxidation and protein oxidation [13,14]. Recent studies have revealed that ROS-induced peroxidation products, such as the lipid peroxidation product malondialdehyde (MDA) and the protein oxidation product carbonyl groups, can damage other cellular components and exacerbate neuronal dysfunction, further demonstrating the detrimental consequence of oxidative stress in neurodegeneration [4,15,16]. Therefore, strategies to reduce oxidative damages are shown to be beneficial to alleviate neurodegenerative symptoms. For example, intake of foods rich in antioxidant ingredients has shown potentials to prevent oxidative stress-related conditions, including NDD [17,18,19]. Among the reported ingredients, polysaccharides, an important type of natural polymers consisting of monosaccharide units that contain multiple free hydroxyl groups, are shown to have both in vitro and in vivo antioxidant activities [20,21]. Here, we first review the antioxidant effects of food-derived polysaccharides and then focus on their protective function against neurodegeneration.
2. Reduction of Oxidative Stress by Food-Derived Polysaccharides
During the last decade, a large body of evidence has shown that polysaccharides and glycoconjugates from a variety of natural sources, including bacteria, fungi, algae, plants and animals, have antioxidant potentials [20,21,22,23]. In particular, polysaccharides isolated from functional and medicinal foods as well as from common foods have drawn great attention in antioxidant studies. Here, we attempt to summarize recent studies of antioxidant polysaccharides from food resources, including vegetables, fruits, cereals, beans, mushrooms, tea, milk products and meat (Table 1) [24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73].
2.1. Reduction of Free Radical and Peroxidation Product Levels
Many food-derived polysaccharides are reported to have potent reducing power and free radical scavenging ability in vitro. For example, we have previously isolated a polysaccharide from Nostoc commune, a widespread microalga with a long history as food and medicine, and found that the polysaccharide is capable of scavenging both superoxide anion and hydroxyl radicals in vitro [66]. The antioxidant capability of polysaccharides is shown to be related with their functional groups such as hydroxyl, amino, carbonyl and carboxyl groups, e.g., the scavenging capacity of chitosan against superoxide radicals is correlated with its number of hydroxyl and amino groups [74]. The polysaccharide fractions from Zizyphus jujuba with higher uronic acid content exhibit stronger free radical scavenging activities than other polysaccharide fractions from the same species containing no uronic acid [75]. These functional groups in polysaccharides can donate hydrogen to electron-deficient free radicals to generate alkoxyl products, which accelerate intramolecular hydrogen abstraction and further induce spirocyclization reaction to prevent radical chain reaction [22,76]. Interestingly, free radicals are usually generated via transition metal ions in in vitro antioxidant assays. In Fenton reaction, for instance, ferrous ion is used to catalyze superoxide or hydrogen peroxide to generate hydroxyl radicals [77]. Therefore, the direct scavenging effect of polysaccharides against free radicals may also be through chelating ions. For example, the polysaccharide fraction GAPS-1 isolated from Aloe barbadensis has a higher chelating ability against ferrous ion and meanwhile exhibits stronger scavenging effect against hydroxyl radicals as compared to SAPS-1, another polysaccharide fraction isolated from the same species [51]. Moreover, monosaccharide composition and substitution groups of polysaccharides are reported to play important roles in their chelating capacity, e.g., the chelating ability of the polysaccharides from Zizyphus jujuba, a well-known traditional food, against ferrous ion is positively correlated with their galacturonic acid contents [50].
In addition to scavenge free radicals in vitro, antioxidant polysaccharides are also shown to reduce the levels of ROS and associated peroxidation products in cellular and animal models under oxidative stress. For instance, a polysaccharide from the common fungus Auricularia auricular is capable of increasing the survival rate and reducing the ROS level in hydrogen peroxide-stressed Caenorhabditis elegans [59], while the wheat bran-derived feruloyl oligosaccharides can reduce MDA content and suppress protein carbonyl formation in human erythrocytes exposed to 2,2′-Azobis(2-amidinopropane) dihydrochloride, a potent free radical generator [34]. It is well established that peroxidation products can modify cellular components, leading to cell damages. For instance, MDA interacts with proteins and DNA to generate covalent adducts with mutagenic and carcinogenic effects [3], while protein carbonyl groups can cause rapid degradation of proteins [78]. Therefore, reduction of peroxidation product contents may contribute to the protective effects of feruloyl oligosaccharides against oxidative stress.
Mitochondria are the main source of ROS and energy production in cells. However, mitochondrial dysfunction, including mitochondrial membrane potential (MMP) decline, respiratory chain malfunction and calcium dysregulation, can accelerate ROS generation and reduce ATP generation, leading to oxidative damage and energy deficiency [79,80]. In a vicious cycle, excessive ROS further impair mitochondrial components such as membrane lipids and DNA, resulting in a secondary mitochondrial dysfunction that amplifies oxidative stress [81,82]. Therefore, restoring mitochondrial function is a beneficial strategy to reduce oxidative impairment. Interestingly, recent reports have revealed that the antioxidant function of food-derived polysaccharides is associated with the alleviation of mitochondrial dysfunction. For example, we have recently shown that the polysaccharide DiPS from Dictyophora indusiata, an edible mushroom traditionally used for inflammatory and neural diseases, can reduce paraquat-mediated increase of ROS level through elevating MMP in C. elegans [58]. A polysaccharide from Ganoderma lucidum, a well-known mushroom traditionally used to delay ageing and enhance immune function, is able to attenuate isoproterenol-induced cardiotoxicity via increasing MMP and mitochondrial complex activity in rats [56]. In addition to mitochondria, several other biochemical pathways such as NADPH oxidase also contribute to ROS production [9]. Interestingly, a recent study has found that a polysaccharide from Sophora subprosrate, a medicinal food used for inflammatory disorders, can reduce superoxide anion in porcine circovirus type 2-infected murine macrophage RAW264.7 cells by inhibiting the expression of NADPH oxidase, which is a major enzyme responsible for generating superoxide anion in phagocytes [43]. Together, these findings demonstrate that antioxidant polysaccharides can inhibit cellular ROS generation through multiple pathways.
2.2. Improvement of the Antioxidant Defense System
A number of studies have revealed that food-derived polysaccharides can reduce oxidative stress and associated damages through modulation of antioxidant enzymes in experimental models. For example, we have recently found that the acidic polysaccharide EbPS-A1 from Epimedium brevicornum, a functional food used for a variety of medical conditions including neurological disorders, can increase oxidative survival and reduce ROS level and MDA content of both wild-type and polyglutamine (polyQ) C. elegans under paraquat-induced oxidative stress. The protective effect of EbPS-A1 against paraquat toxicity is shown to be related with increasing SOD and CAT activities [38]. Interestingly, the polysaccharides isolated from the tonic food Chuanminshen violaceum are also shown to up-regulate the mRNA expression levels of SOD isoforms and CAT and enhance the activities of these antioxidant enzymes in mice injected with d-galactose [39], an ageing-promoting agent that induces cognitive and motor performance deterioration similar to AD symptoms via oxidative stress and mitochondrial dysfunction [83].
In addition to their effect on antioxidant enzymes, several food-derived polysaccharides are also reported to have modulatory function on non-enzyme components of the cellular antioxidant system. For example, a polysaccharide from Anoectochilus roxburghii, a medicinal food used to treat a variety of chronic diseases such as hepatitis and diabetes, is shown to attenuate oxidative stress by increasing glutathione level as well as antioxidant enzyme activities in the hepatic tissue of mice injected with carbon tetrachloride, an organic chemical that can induce hepatotoxicity through increased oxidative stress [84]. Interestingly, A. roxburghii polysaccharide is also shown to reduce the mRNA levels of inflammation-related genes including tumor necrosis factor alpha (TNF-α) and interleukin-6 (IL-6) [52]. Oxidative stress is known to increase the expression of TNF-α, a key cytokine that promotes inflammation, while elevated TNF-α level can activate NADPH oxidase, ultimately leading to ROS overproduction [85,86].
2.3. Regulation of Oxidative Stress-Related Signaling
A number of signaling pathways, such as those involving nuclear factor erythroid 2-related factor 2/antioxidant response element (Nrf2/ARE), mitogen-activated protein kinases (MAPKs), phosphoinositide 3 kinase/Akt (PI3K/Akt) and insulin/insulin-like growth factor-1 signaling (IIS), are known to be associated with cellular responses to multiple stresses including oxidative stress [87,88,89]. For instance, Nrf2, a basic region leucine-zipper transcription factor, plays an important role in cellular antioxidant response. When Nrf2 is activated, it translocates into nucleus and binds to ARE, leading to up-regulation of genes involved in cellular antioxidant and anti-inflammatory defense as well as mitochondrial protection [87]. Interestingly, some food-derived polysaccharides are recently reported to exert their antioxidant activity via Nrf2/ARE pathway in cellular and animal models. For instance, a polysaccharide from Lycium barbarum, a medicinal food traditionally used to retard ageing and improve neuronal function, is shown to attenuate ultraviolet B-induced cell viability decrease and ROS level increase in human keratinocytes HaCaT cells by promoting the nuclear translocation of Nrf2 and the expression of Nrf2-dependent ARE target genes [90]. This protective effect of L. barbarum polysaccharide can be neutralized by siRNA-mediated Nrf2 silencing, indicating an involvement of Nrf2/ARE pathway in the antioxidant effect of the polysaccharide [90]. Intriguingly, however, the above-mentioned polysaccharide DiPS is shown to increase oxidative survival through promoting nuclear translocation of transcription factor DAF-16/FOXO transcription factor but not SKN-1 (worm homologue of Nrf2) in wild-type C. elegans under paraquat exposure, demonstrating the antioxidant activity of the polysaccharide is associated with IIS, an evolutionarily conserved pathway that regulates organismal metabolism and lifespan, as DAF-16 is a key regulator in IIS [58].
Several signaling pathways related to cell death and survival are also involved in the antioxidant effect of food-derived polysaccharides. For example, hydrogen peroxide can induce apoptosis of rat pheochromocytoma PC12 cells via activation of p38 MAPK, while a polysaccharide from the fruiting bodies of the edible mushroom Morchella importuna increases the viability of hydrogen peroxide-exposed PC12 cells by inhibiting p38 MAPK phosphorylation [91]. In addition, hydrogen peroxide can inhibit the activation of PI3K/Akt signaling in human neuroblastoma SH-SY5Y cells, while sulfated polysaccharides prepared from fucoidan are able to increase the phosphorylation of PI3K/Akt and inhibit cell apoptosis [92]. Interestingly, the PI3K inhibitor LY294002 can partially prevent the beneficial role of the polysaccharide, demonstrating that modulation of PI3K/Akt pathway contributes to the protective effect of the sulfated polysaccharides against hydrogen peroxide cytotoxicity [92].
Recent studies provide clear evidence for the protective effects of food-derived polysaccharides against oxidative stress. Many polysaccharides exhibit potent reducing power, total antioxidant capacity and scavenging ability against free radicals in vitro. Moreover, some polysaccharides can decrease ROS and peroxidation product levels, improve antioxidant defense system and regulate stress-related signaling events to attenuate oxidative damage in cellular and animal models exposed to a variety of external stimuli, such as hydrogen peroxide, paraquat, ultraviolet radiation and virus. Together, these findings suggest a potential of these dietary polysaccharides to maintain health and prevent oxidative stress-related disorders.
3. Alleviation of Neurodegeneration by Food-Derived Antioxidant Polysaccharides
It is known that oxidative stress and chronic inflammation are two intertwined pathological events in NDD [85]. Excessive ROS can modulate inflammatory signaling to up-regulate the expression of pro-inflammatory factors such as cytokines, which act as potent stimuli in brain inflammation [93,94]. In turn, elevated inflammatory stress further provokes ROS generation via multiple pathways such as nuclear factor κB (NF-κB) signaling [85]. On the other hand, abnormal protein aggregation is known to be a common pathological hallmark of late-onset NDD. These protein aggregates, including amyloid-β peptide (Aβ) aggregates in AD and polyQ aggregates in HD, can induce neuronal damages through induction of oxidative stress, inflammation and mitochondrial dysfunction [95,96,97]. Oxidative stress can also promote the aggregation of pathogenic proteins as ROS modified-proteins tend to form aggregates [98]. In addition, a variety of chemical interventions, including excitatory amino acids such as glutamate, N-methyl-d-aspartate (NMDA) and kainic acid; neurotoxins such as 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) and 6-hydroxydopamine (6-OHDA); and ageing-promoting agents such as d-galactose, are also shown to induce neurodegenerative symptoms via oxidative and inflammatory stresses [83,99,100]. As oxidative stress plays a pivotal role in neurodegeneration, antioxidant strategies, including food-derived antioxidant polysaccharides, are shown to attenuate neuronal damage and improve cognitive and motor functions in a range of neurodegenerative models (Table 2) [101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142].
3.1. Effects on Alzheimer’s Disease
AD is characterized by amyloid plaques and neurofibrillary tangles in the brain, which lead to progressive memory loss and cognitive decline [143]. As global population ages, AD has become a major public health concern. Among therapeutic and nutritional interventions, growing evidence has shown that adequate intake of antioxidants may be helpful to reduce neuronal damages and alleviate AD symptoms [144,145]. For example, dietary intake of α-tocopherol or combined tocopherols shows beneficial effects to alleviate age-related cognitive decline and lower AD risk [146,147].
Antioxidant polysaccharides from various food sources are also found to inhibit Aβ-mediated neurotoxicity in experimental models (Table 2). The polysaccharide PS-WNP from the medicinal food Polygonatum sibiricum, for instance, is shown to significantly attenuate Aβ-induced apoptosis of PC12 cells by alleviating mitochondrial dysfunction, regulating apoptosis-related protein Bax and Bcl-2 levels, inhibiting apoptotic executor caspase-3 activation and enhancing Akt phosphorylation [105]. In rats injected with Aβ40 aggregates, fucoidan is shown to attenuate learning and memory deficits by elevating SOD and GPx activities and decreasing MDA content, Bax/Bcl-2 ratio and caspase-3 activity in hippocampal tissue [104]. Using transgenic C. elegans models that overexpress Aβ proteins, the D. indusiata polysaccharide DiPS is shown to alleviate chemosensory behavior dysfunction, which is associated with reduction of ROS level and MDA content, increase of SOD activity and alleviation of mitochondrial dysfunction [58]. Antioxidant polysaccharides are also shown to modulate pathogenic protein aggregation, e.g., L. barbarum polysaccharides can reduce Aβ42 protein level in hippocampal tissue and improve the performance of learning and memory in APP/PS1 mice [109]. Intriguingly, L. barbarum polysaccharide is also shown to inhibit the apoptosis and reduce cleaved-tau protein level, the main component of neurofibrillary tangles in AD patients, in rat primary cortical cells exposed to homocysteine, a sulfur-containing amino acid associated with several NDD [128]. Moreover, several studies have uncovered that the regulatory effect of polysaccharides on protein aggregation is through the interaction with aggregation-prone proteins, and this effect is influenced by the chemical structure of polysaccharides. For example, four glycosaminoglycans from different animal tissues are shown to inhibit the neurotoxicity of serum amyloid P component and its interaction with Aβ, and the inhibitory efficacy is correlated with the uronic acid content in glycosaminoglycans [148]. In addition, the well-known glycosaminoglycan heparin is reported to bind with Aβ and promote amyloid fibrillogenesis, while low molecular weight heparin can prevent Aβ aggregation by blocking β-sheet formation and inhibiting fibril formation, suggesting that the molecular weight of polysaccharides may also affect their interaction with proteins [149,150]. Together, these studies demonstrate that the neuroprotective effects of food-derived polysaccharides in AD-like models correlate with their modulation of oxidative and related stresses.
3.2. Effects on Parkinson’s Disease
PD is a chronic and progressive NDD characterized by selective loss of dopaminergic neurons in the substantia nigra pars compacta and abnormal accumulation of Lewy bodies in these neurons [151]. The major clinical symptoms of PD include motor symptoms such as tremor and bradykinesia, and neuropsychiatric symptoms such as cognitive decline and anxiety [152]. Current clinic therapy for PD only concentrates on symptomatic management as the available therapeutics do not prevent disease progression [153].
Recent studies have shown that several food-derived antioxidant polysaccharides are capable of inhibiting the neurotoxicity mediated by MPTP and 6-OHDA, which can selectively induce dopaminergic neuron death and cause PD-like motor deficits in experimental models (Table 2). For instance, the polysaccharides from the seaweed Saccharina japonica and from the sea cucumber Stichopus japonicus can increase 6-OHDA-induced reduction of cell viability in SH-SY5Y cells and murine embryonic stem MES 23.5 cells, respectively [119,142]. The S. japonicus polysaccharides are shown to increase SOD activity, regulate the level of apoptosis-related proteins, inhibit NF-κB and p38 MAPK activation and activate PI3K/Akt pathway, indicating the involvement of antioxidant, anti-apoptotic and anti-inflammatory signaling pathways in its neuroprotective effect [119]. Using MPTP-injected mouse models, low molecular weight fucoidan DF and its two fractions DF1 and DF2 are shown to ameliorate dopaminergic neuron injury and prevent dopamine depletion in the substantia nigra through enhancing antioxidant enzyme activities and inhibiting neuronal apoptosis [154]. Interestingly, DF1 exerts better neuroprotective activity than DF and DF2 in general, and their monosaccharide compositions are different: DF1 is a hetero-polysaccharide with low content of fucose and high content of uronic acid and other monosaccharides, while DF2 mainly consists of fucose and galactose, suggesting that chemical composition may play an important role in the neuroprotective activity of fucoidan [154]. In addition, a polysaccharide from the edible microalga Chlorella pyrenoidosa is recently shown to reduce bradykinesia, inhibit the loss of striatal dopamine and its metabolites, and increase tyrosine hydroxylase in MPTP-injected mice [117]. The polysaccharide can also elevate the levels of small intestinal secretory immunoglobulin A, a protein that is crucial for the immune function of mucous membranes, in mice serum [117], and has been previously shown to enhance immune function [155]. As immune system dysfunction is known to contribute to PD development and progression [156], immune-related therapies may be a useful strategy to reduce disease risks and retard disease progression [157].
3.3. Effects on Huntington’s Disease
HD is an autosomal-dominant neurodegenerative disorder that is clinically manifested by a variety of motor, cognitive and psychiatric deficits [158]. This disease is caused by an abnormal expanded CAG trinucleotide repeat in the huntingtin gene on the short arm of chromosome 4. In normal individuals, the average number of CAG repeats in the huntingtin gene is 17–20; when the number of repeats exceeds 36, the risk of developing HD is significantly increased [159]. The prevalence of HD varies geographically, with the highest rates in Europe (~10–15 per 100,000 individuals) and lower rates in Asia and Africa [160]. Similar with AD and PD, currently there is no efficient treatment for HD.
Among various pharmacological interventions, natural antioxidants such as epigallocatechin gallate and salidroside have been found to alleviate HD-like symptoms in transgenic cellular and animal models [161,162]. Interestingly, several recent studies have uncovered that food-derived antioxidant polysaccharides also have beneficial effects in HD-like animal models (Table 2). For example, the E. brevicornum polysaccharide EbPS-A1 can alleviate polyQ-mediated chemosensory dysfunction in transgenic C. elegans model HA759 [38], which expresses a polyQ tract of 150 glutamine repeats in amphid sensilla (ASH) neurons, leading to progressive ASH death and chemotactic behavior deficit [163]. EbPS-A1 also reduces ROS level, inhibits lipid peroxidation and enhances antioxidant enzyme activities in HA759 nematodes, indicating that the antioxidant activity of the polysaccharide contributes to its protective effect against polyQ neurotoxicity [38]. Other studies suggest that some antioxidant polysaccharides exert their neuroprotective effects by targeting polyQ aggregate itself, e.g., L. barbarum polysaccharide not only increases the viability of HEK293 cells that express mutant-huntingtin containing 160 glutamine repeats but also improves motor behavior and lifespan in HD-related transgenic mice [122]. The neuroprotective effect of L. barbarum polysaccharide against mutant-huntingtin toxicity in both cellular and mouse models are shown to be associated with reducing mutant-huntingtin levels and activating AKT [122]. These studies provide an important insight into the therapeutic potential of food-derived antioxidant polysaccharides in HD.
3.4. Effects on Other Neurodegenerative Symptoms
Several recent studies have shown that food-derived antioxidant polysaccharides are capable of inhibiting excitatory amino acid-mediated neurotoxicity, which is implicated in many NDD [99,164]. For instance, L. barbarum polysaccharide can increase cell viability and suppress JNK activation in glutamate-exposed rat primary cortical neurons [129], suggesting an involvement of MAPK pathway in the neuroprotective effect of the polysaccharide. Another example is G. lucidum polysaccharide, which is shown to alleviate epileptic symptoms and up-regulate the expression of calcium/calmodulin-dependent protein kinase II, a kinase that plays an important role in calcium transfer in neurons, in kainic acid-injected rats [123]. As calcium overload mediates excitatory amino acid-induced neurotoxicity [164], prevention of calcium transporting may contribute to this neuroprotective effect of G. lucidum polysaccharide. In addition, other chemicals can also induce cognitive impairment and behavior deficit through increase of oxidative and inflammatory stresses, and the polysaccharides isolated form mushrooms, medicinal herbs and algae are reported to attenuate neurodegenerative symptoms induced by these toxic chemicals. For instance, a polysaccharide from Pleurotus ostreatus can decrease escape latency in Morris water maze test and increase passive avoidance latency in step-down test in rats under d-galactose and aluminum chloride challenge [125]. P. ostreatus polysaccharide also reduces MDA level and elevates SOD, GPx and CAT activities [125], indicating that the behavior-improving capability of the polysaccharide correlates with reduction of oxidative stress.
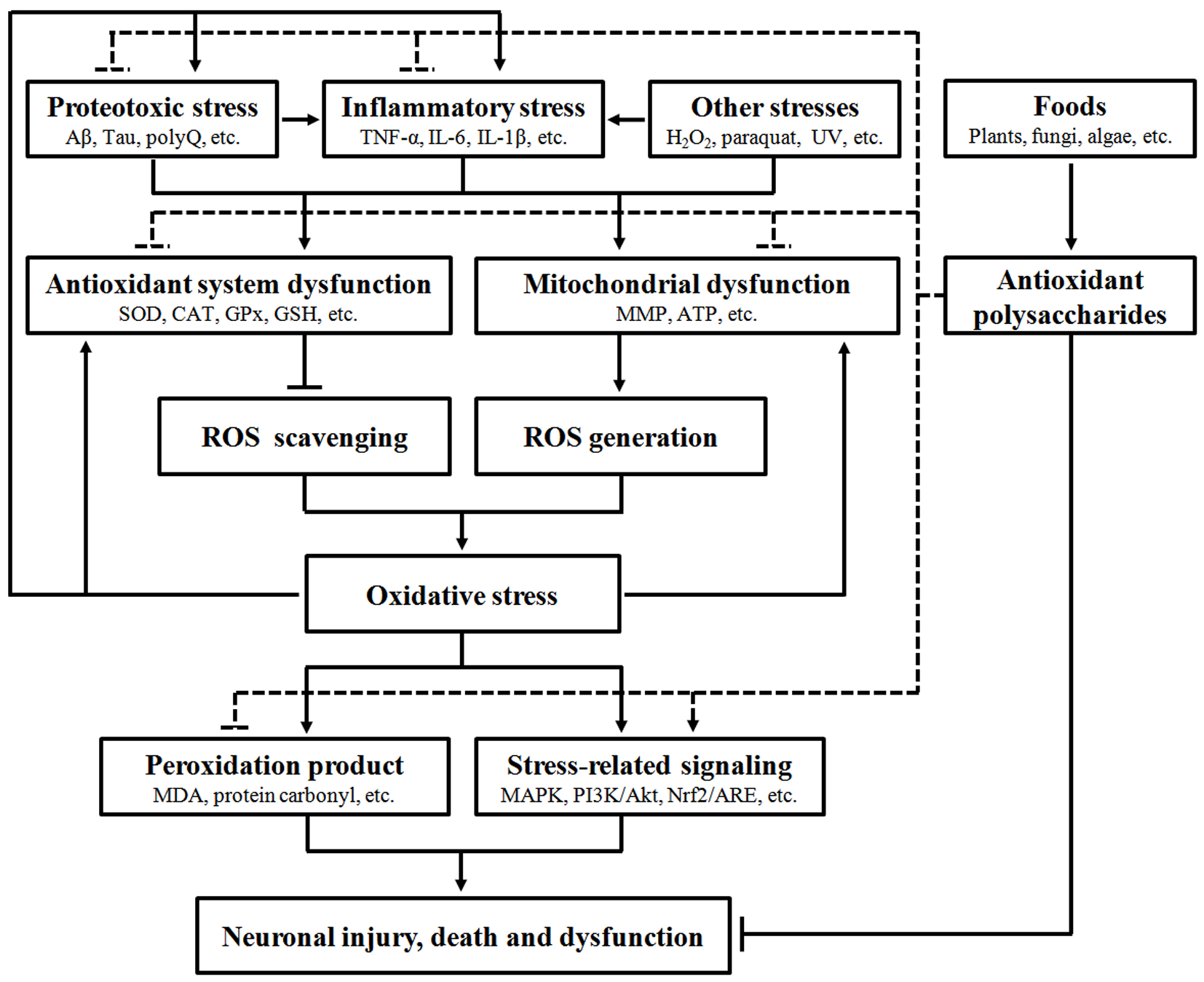
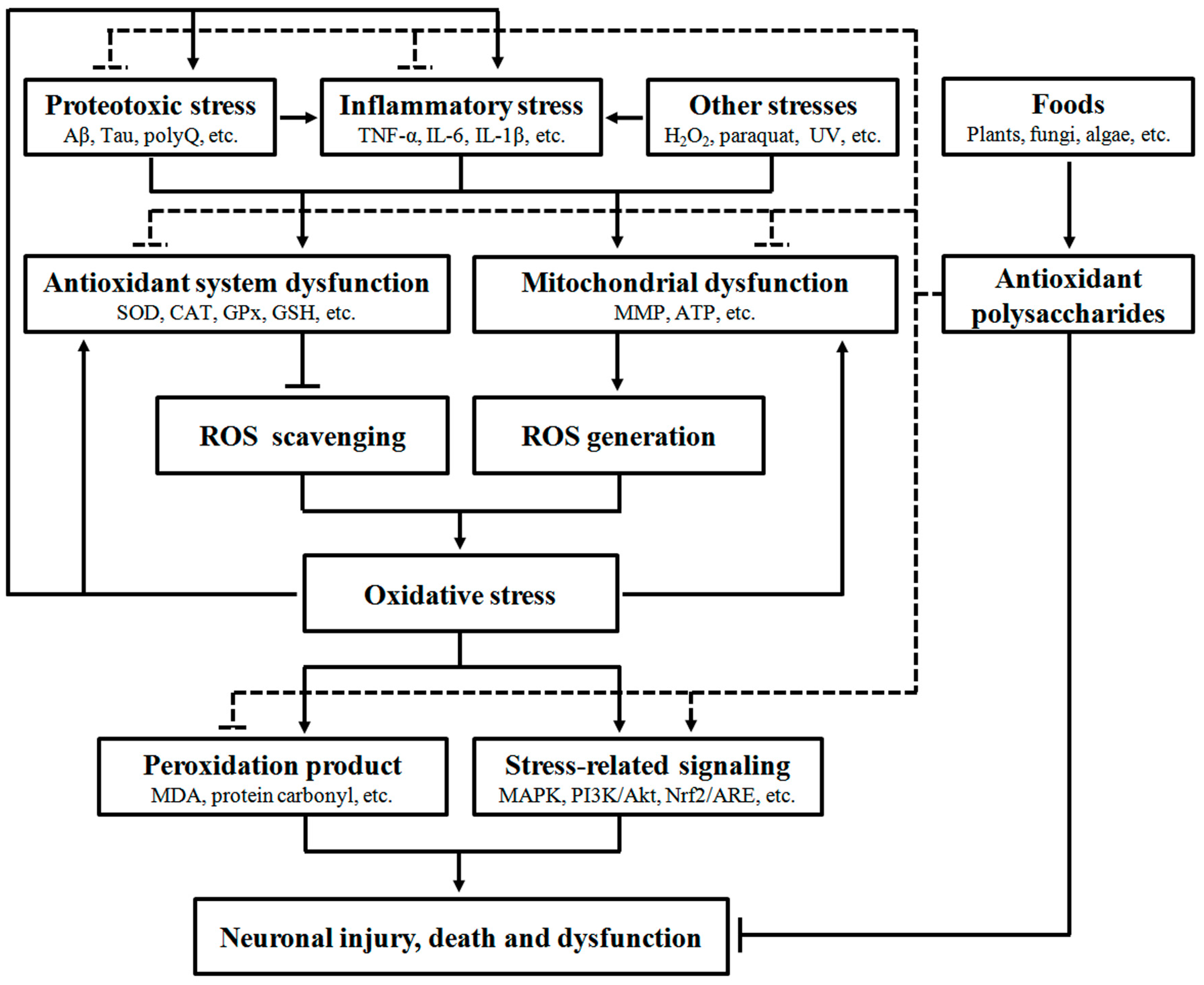
A large body of evidence has confirmed that oxidative stress can interact with many other stresses to induce neurodegeneration, indicating its significant role in NDD development. Food-derived antioxidant polysaccharides are recently shown to alleviate neuronal injury, death and dysfunction through modulation of multiple oxidative stress-related pathways, including antioxidant defense system, mitochondrial function, peroxidation products, protein aggregation, inflammation and stress-related signaling (Figure 1), demonstrating their pharmacological potentials in NDD.
4. Conclusions
Food-derived polysaccharides have been shown to scavenge free radicals in vitro and reduce oxidative damages in cellular and animal models, and their in vivo antioxidant capacities are related with regulation of peroxidation products, antioxidant defense system and stress-related signaling. As oxidative stress is closely associated with neurodegeneration, some antioxidant polysaccharides are also tested for their anti-NDD activity and found to attenuate neuronal damages and dysfunction in a number of neurodegenerative models. The neuroprotective effects of polysaccharide are associated with alleviation of multiple stresses, including oxidative, inflammatory and proteotoxic stresses (Figure 1). Therefore, consumption of foods rich in antioxidant polysaccharides may not only reduce oxidative damage but also provide protection against oxidative stress-related disorders. It is noted that most recent studies focus on the antioxidant polysaccharides from terrestrial plants and fungi, and relatively less attention is paid to marine organisms although they represent a rich resource of bioactive polysaccharides. In addition, many food-derived antioxidant polysaccharides are shown to have potent immunomodulatory effects, and therefore it would be interesting to explore the involvement of immunomodulation in the neuroprotective effect of antioxidant polysaccharides.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (Grant 81403081), the National High-Tech R & D Program of China (863 Program; Grant 2014AA022001), the International Science & Technology Cooperation Program of China (Grant 2015DFA30280), the Special Funds of the Central Finance to Support the Development of Local Universities and Colleges, and Guangdong Province Department of Education (Grants 2015KGJHZ022 and 2016KQNCX084).
Author Contributions
Zebo Huang and Haifeng Li conceived and designed the review. Haifeng Li, Fei Ding, Lingyun Xiao, Ruona Shi, Hongyu Wang, and Wenjing Han collected the literature, analyzed the data and made figures and tables. Haifeng Li, Fei Ding, and Zebo Huang wrote and revised the manuscript. All authors have read and approved the final version.
Conflicts Interest
The authors declare that there is no conflict of interests.
References
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108, 652–659. [Google Scholar] [CrossRef]
- Reczek, C.R.; Chandel, N.S. ROS-dependent signal transduction. Curr. Opin. Cell Biol. 2015, 33, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Dalle-Donne, I.; Aldini, G.; Carini, M.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation, cellular dysfunction, and disease progression. J. Cell. Mol. Med. 2006, 10, 389–406. [Google Scholar] [CrossRef] [PubMed]
- Rahal, A.; Kumar, A.; Singh, V.; Yadav, B.; Tiwari, R.; Chakraborty, S.; Dhama, K. Oxidative stress, prooxidants, and antioxidants: The interplay. BioMed Res. Int. 2014, 2014, 761264. [Google Scholar] [CrossRef] [PubMed]
- Al-Dalaen, S.M.; Al-Qtaitat, A.I. Review article: Oxidative stress versus antioxidants. J. Biosci. Bioeng. 2014, 2, 60–71. [Google Scholar] [CrossRef]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ. J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Patel, A.K.; Shah, N.; Chaudhary, A.K.; Jha, U.K.; Yadav, U.C.; Gupta, P.K.; Pakuwal, U. Oxidative stress and antioxidants in disease and cancer: A review. Asian Pac. J. Cancer Prev. 2014, 15, 4405–4409. [Google Scholar] [CrossRef] [PubMed]
- Rahman, T.; Hosen, I.; Towhidul Islam, M.M.; Uddin Shekhar, H. Oxidative stress and human health. Adv. Biosci. Biotechnol. 2012, 3, 997–1019. [Google Scholar] [CrossRef]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; de Bittencourt, P.I., Jr. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Gammon, K. Neurodegenerative disease: Brain windfall. Nature 2014, 515, 299–300. [Google Scholar] [CrossRef] [PubMed]
- Nikam, S.; Nikam, P.; Ahaley, S.K.; Sontakke, A.V. Oxidative stress in Parkinson’s disease. Indian J. Clin. Biochem. 2009, 24, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhong, C. Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 2014, 30, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Melo, A.; Monteiro, L.; Lima, R.M.; Oliveira, D.M.; Cerqueira, M.D.; El-Bachá, R.S. Oxidative stress in neurodegenerative diseases: Mechanisms and therapeutic perspectives. Oxid. Med. Cell. Longev. 2011, 2011, 467180. [Google Scholar] [CrossRef] [PubMed]
- Sultana, R.; Perluigi, M.; Allan Butterfield, D. Lipid peroxidation triggers neurodegeneration: A redox proteomics view into the Alzheimer disease brain. Free Radic. Biol. Med. 2013, 62, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F. Nutraceuticals, functional foods and dietary supplements in health and disease. J. Food Drug Anal. 2012, 20, 226–230. [Google Scholar]
- Herrera, E.; Jiménez, R.; Aruoma, O.I.; Hercberg, S.; Sánchez-García, I.; Fraga, C. Aspects of antioxidant foods and supplements in health and disease. Nutr. Rev. 2009, 67, S140–144. [Google Scholar] [CrossRef] [PubMed]
- Virmani, A.; Pinto, L.; Binienda, Z.; Ali, S. Food, nutrigenomics, and neurodegeneration--neuroprotection by what you eat! Mol. Neurobiol. 2013, 48, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, Y.M.; Qi, Z.M.; Wang, S.Y.; Liu, S.X.; Li, X.; Wang, H.J.; Xia, X.C. An overview on natural polysaccharides with antioxidant properties. Curr. Med. Chem. 2013, 20, 2899–2913. [Google Scholar] [CrossRef] [PubMed]
- Mei, X.; Yi, C.; Huang, G. The antioxidant activities of polysaccharides and their derivatives. Curr. Drug Targets. in press. [CrossRef]
- Wang, J.; Hu, S.; Nie, S.; Yu, Q.; Xie, M. Reviews on mechanisms of in vitro antioxidant activity of polysaccharides. Oxid. Med. Cell. Longev. 2016, 2016, 5692852. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Xie, J.H.; Nie, S.P.; Xie, M.Y. Review on cell models to evaluate the potential antioxidant activity of polysaccharides. Food Funct. 2017, 8, 915–926. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhu, P.; Zhao, S.; Nie, C.; Wang, N.; Du, X.; Zhou, Y. Characterization, antioxidant activity and immunomodulatory activity of polysaccharides from the swollen culms of Zizania latifolia. Int. J. Biol. Macromol. 2017, 95, 809–817. [Google Scholar] [CrossRef] [PubMed]
- Ghazala, I.; Sila, A.; Frikha, F.; Driss, D.; Ellouz-Chaabouni, S.; Haddar, A. Antioxidant and antimicrobial properties of water soluble polysaccharide extracted from carrot peels by-products. J. Food Sci. Technol. 2015, 52, 6953–6965. [Google Scholar] [CrossRef]
- Nara, K.; Yamaguchi, A.; Maeda, N.; Koga, H. Antioxidative activity of water soluble polysaccharide in pumpkin fruits (Cucurbita maxima Duchesne). Biosci. Biotechnol. Biochem. 2009, 73, 1416–1418. [Google Scholar] [CrossRef] [PubMed]
- Jeddou, K.B.; Chaari, F.; Maktouf, S.; Nouri-Ellouz, O.; Helbert, C.B.; Ghorbel, R.E. Structural, functional, and antioxidant properties of water-soluble polysaccharides from potatoes peels. Food Chem. 2016, 205, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Wei, X.; Zhang, X.; Cheng, F.; Shuai, X.; Zhang, L.; Kang, L. Protective effect of Potentilla anserine polysaccharide (PAP) on hydrogen peroxide induced apoptosis in murine splenic lymphocytes. Carbohydr. Polym. 2010, 79, 356–361. [Google Scholar] [CrossRef]
- Kim, S.Y.; Kim, E.A.; Kim, Y.S.; Yu, S.K.; Choi, C.; Lee, J.S.; Kim, Y.T.; Nah, J.W.; Jeon, Y.J. Protective effects of polysaccharides from Psidium guajava leaves against oxidative stresses. Int. J. Biol. Macromol. 2016, 91, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Dou, J.; Guo, Y.; Xue, Z.; Chen, W.; Li, J.; Meng, Y. Purification and antioxidative activity of polysaccharides from cold-extracting apple peel. Sci. Technol. Food Ind. 2014, 1, 111–115. [Google Scholar]
- Zhang, Y.; Lu, X.; Fu, Z.; Wang, Z.; Zhang, J. Sulphated modification of a polysaccharide obtained from fresh persimmon (Diospyros kaki L.) fruit and antioxidant activities of the sulphated derivatives. Food Chem. 2011, 127, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Song, S.; Guo, J.; Luo, S.; Zhang, J. Protective effects of polysaccharide from seed watermelon on the oxidative damage of PC12 cells induced by H2O2. Nat. Prod. Res. Dev. 2015, 2, 338–343. [Google Scholar]
- Hefnawy, H.T.; El-Shourbagy, G.A. Chemical analysis and antioxidant activity of polysaccharide extracted from rice bran. World J. Dairy Food Sci. 2014, 9, 95–104. [Google Scholar] [CrossRef]
- Wang, J.; Sun, B.; Cao, Y.; Tian, Y. Protection of wheat bran feruloyl oligosaccharides against free radical-induced oxidative damage in normal human erythrocytes. Food Chem. Toxicol. 2009, 47, 1591–1599. [Google Scholar] [CrossRef] [PubMed]
- Mateos-Aparicio, I.; Mateos-Peinado, C.; Jiménez-Escrig, A.; Rupérez, P. Multifunctional antioxidant activity of polysaccharide fractions from the soybean by product okara. Carbohydr. Polym. 2010, 82, 245–250. [Google Scholar] [CrossRef]
- Ye, Z.; Wang, W.; Yuan, Q.; Ye, H.; Sun, Y.; Zhang, H.; Zeng, X. Box-Behnken design for extraction optimization, characterization and in vitro antioxidant activity of Cicer arietinum L. hull polysaccharides. Carbohydr. Polym. 2016, 147, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Wang, Y.; Li, X.; Yu, P. Purification and structural characterization of Chinese yam polysaccharide and its activities. Carbohydr. Polym. 2015, 117, 1021–1027. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.; Zhang, J.; Li, H.; Wang, Q.; Xiao, L.; Weng, H.; Zhou, X.; Ma, C.; Ma, F.; Hu, M.; et al. Epimedium polysaccharide alleviates polyglutamine-induced neurotoxicity in Caenorhabditis elegans by reducing oxidative stress. Rejuvenation Res. 2017, 20, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Feng, H.; Yu, Y.; Sun, M.; Liu, Y.; Li, T.; Sun, X.; Liu, S.; Sun, M. Antioxidant activities of the polysaccharides of Chuanminshen violaceum. Carbohydr. Polym. 2017, 157, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Sui, Z.; Li, L.; Liu, B.; Gu, T.; Zhao, Z.; Liu, C.; Shi, C.; Yang, R. Optimum conditions for Radix Rehmanniae polysaccharides by RSM and its antioxidant and immunity activity in UVB mice. Carbohydr. Polym. 2013, 92, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Qi, B.; Ji, Q.; Wen, Y.; Liu, L.; Guo, X.; Hou, G.; Wang, G.; Zhong, J. Lycium barbarum polysaccharides protect human lens epithelial cells against oxidative stress-induced apoptosis and senescence. PLoS ONE 2014, 9, e110275. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Li, H.; Fang, Z.; Lin, J.; Wang, S.; Xiao, L.; Yang, F.; Liu, X.; Zhang, J.; Huang, Z.; et al. Polysaccharides from Angelica sinensis alleviate neuronal cell injury caused by oxidative stress. Neural. Regen. Res. 2014, 9, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.J.; Yang, J.; Luo, W.J.; Wei, Y.Y.; Shuai, X.H.; Hu, T.J. Inhibitory effect of Sophora subprosrate polysaccharide on mitochondria oxidative stress induced by PCV-2 infection in RAW264.7 cells. Int. J. Biol. Macromol. 2017, 95, 608–617. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Liu, Q.; Wang, W.; Li, X.; Zhang, J. A polysaccharide isolated from Cynomorium songaricum Rupr. protects PC12 cells against H2O2-induced injury. Int. J. Biol. Macromol. 2016, 87, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Qu, Z.; Fu, L.; Dong, P.; Zhang, X. Physicochemical properties and antioxidant capacity of 3 polysaccharides from green tea, oolong tea, and black tea. J. Food Sci. 2009, 74, C469–474. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, Z.; Wei, X. Antioxidant activities potential of tea polysaccharide fractions obtained by ultra filtration. Int. J. Biol. Macromol. 2012, 50, 558–564. [Google Scholar] [CrossRef] [PubMed]
- Shang, X.; Chao, Y.; Zhang, Y.; Lu, C.; Xu, C.; Niu, W. Immunomodulatory and antioxidant effects of polysaccharides from Gynostemma pentaphyllum Makino in immunosuppressed mice. Molecules 2016, 21, 1085. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; He, L.; Wang, Y.; Cheng, J. Optimization extraction, preliminary characterization and antioxidant activities of polysaccharides from Semen Juglandis. Molecules 2016, 21, 1335. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, X.; Wang, L.; Lu, F.; Zhang, C.; Zheng, Y.; Xie, H. Physicochemical properties and antioxidant activity of Ginkgo biloba L. nut. Mod. Food Sci. Technol. 2013, 10, 2395–2400. [Google Scholar]
- Chang, S.C.; Hsu, B.Y.; Chen, B.H. Structural characterization of polysaccharides from Zizyphus jujuba and evaluation of antioxidant activity. Int. J. Biol. Macromol. 2010, 47, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, C.; Xu, Z.; Wang, Y. Isolation, chemical characterization and antioxidant activities of two polysaccharides from the gel and the skin of Aloe barbadensis Miller irrigated with sea water. Process Biochem. 2007, 42, 961–970. [Google Scholar] [CrossRef]
- Zeng, B.; Su, M.; Chen, Q.; Chang, Q.; Wang, W.; Li, H. Protective effect of a polysaccharide from Anoectochilus roxburghii against carbon tetrachloride-induced acute liver injury in mice. J. Ethnopharmacol. 2017, 200, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, Q.; Li, H.; Guo, L. Neuroprotective and antioxidative effect of cactus polysaccharides in vivo and in vitro. Cell. Mol. Neurobiol. 2009, 29, 1211–1221. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Ning, Y. Antioxidant and antitumor activities of the polysaccharide from seed cake of Camellia oleifera Abel. Int. J. Biol. Macromol. 2012, 51, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Park, C.M.; Cho, C.W.; Song, Y.S. TOP 1 and 2, polysaccharides from Taraxacum officinale, inhibit NFκB-mediated inflammation and accelerate Nrf2-induced antioxidative potential through the modulation of PI3K-Akt signaling pathway in RAW 264.7 cells. Food Chem. Toxicol. 2014, 66, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Sudheesh, N.P.; Ajith, T.A.; Janardhanan, K.K. Ganoderma lucidum ameliorate mitochondrial damage in isoproterenol-induced myocardial infarction in rats by enhancing the activities of TCA cycle enzymes and respiratory chain complexes. Int. J. Cardiol. 2013, 165, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Kozarski, M.; Klaus, A.; Niksˇic´, M.; Vrvic´, M.; Todorovic´, N.; Jakovljevic´, D.; Griensven, L. Antioxidative activities and chemical characterization of polysaccharide extracts from the widely used mushrooms Ganoderma applanatum, Ganoderma lucidum, Lentinus edodes and Trametes versicolor. J. Food Compos. Anal. 2012, 26, 144–153. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, R.; Li, H.; Xiang, Y.; Xiao, L.; Hu, M.; Ma, F.; Ma, C.; Huang, Z. Antioxidant and neuroprotective effects of Dictyophora indusiata polysaccharide in Caenorhabditis elegans. J. Ethnopharmacol. 2016, 192, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhang, Y.; Jiang, K. Antioxidant activity in vitro and in vivo of the polysaccharides from different varieties of Auricularia auricula. Food Funct. 2016, 7, 3868–3879. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Gao, Q.; Ma, C.; Ge, Y.; You, Li.; Liu, R.; Fu, X.; Liu, D. Effect of polysaccharides from Tremella fuciformis on UV-induced photoaging. J. Funct. Foods 2016, 20, 400–410. [Google Scholar] [CrossRef]
- Zhang, Q.; Yu, P.; Li, Z.; Zhang, H.; Xu, Z.; Li, P. Antioxidant activities of sulfated polysaccharide fractions from Porphyra haitanesis. J. Appl. Phycol. 2003, 15, 305–310. [Google Scholar] [CrossRef]
- Cui, C.; Lu, J.; Sun-Waterhouse, D.; Mu, L.; Sun, W.; Zhao, M.; Zhao, H. Polysaccharides from Laminaria japonica: Structural characteristics and antioxidant activity. Lwt-Food Sci. Technol. 2016, 73, 602–608. [Google Scholar] [CrossRef]
- Rupérez, P.; Ahrazem, O.; Leal, J.A. Potential antioxidant capacity of sulfated polysaccharides from the edible marine brown seaweed Fucus vesiculosus. J. Agric. Food Chem. 2002, 50, 840–845. [Google Scholar] [CrossRef] [PubMed]
- Qi, H.; Zhang, Q.; Zhao, T.; Hu, R.; Zhang, K.; Li, Z. In vitro antioxidant activity of acetylated and benzoylated derivatives of polysaccharide extracted from Ulva pertusa (Chlorophyta). Bioorg. Med. Chem. Lett. 2006, 16, 2441–2445. [Google Scholar] [CrossRef] [PubMed]
- Ku, M.J.; Lee, M.S.; Moon, H.J.; Lee, Y.H. Antioxidation effects of polysaccharide fucoidan extracted from seaweeds in skin photoaging. FASEB J. 2008, 22 647.1. [Google Scholar]
- Li, H.; Xu, J.; Liu, Y.; Ai, S.; Qin, F.; Li, Z.; Zhang, H.; Huang, Z. Antioxidant and moisture-retention activities of the polysaccharide from Nostoc commune. Carbohydr. Polym. 2011, 83, 1821–1827. [Google Scholar] [CrossRef]
- Morifuji, M.; Kitade, M.; Fukasawa, T.; Yamaji, T.; Ichihashi, M. Exopolysaccharides isolated from milk fermented with lactic acid bacteria prevent ultraviolet-induced skin damage in hairless mice. Int. J. Mol. Sci. 2017, 18, 146. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, M.J.; Isaacs, M.; Matsuhiro, B.; Mendoza, L.; Zúñiga, E.A. Characterization of a neutral polysaccharide with antioxidant capacity from red wine. Carbohydr. Res. 2009, 344, 1095–1101. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Shang, N.; Li, P. In vitro and in vivo antioxidant activity of exopolysaccharide fractions from Bifidobacteriumanimalis RH. Anaerobe 2011, 17, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, R.; Shah, N.; Tao, X.; Xiong, Y.; Wei, H. Antioxidant and antibacterial activities of exopolysaccharides from Bifidobacterium bifidum WBIN03 and Lactobacillus plantarum R315. J. Dairy Sci. 2014, 97, 7334–7343. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liang, H.; Guo, W.; Peng, Z.; Chen, J.; Zhang, Q. Isolation, identification, and antioxidant activity of polysaccharides from the shell of abalone (Haliotis discus hannai Ino). Genet. Mol. Res. 2014, 13, 4883–4892. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Pan, J.; Wan, P.; Chen, D.; Long, S.; Sun, H. Ultrasonic-assisted production of antioxidative polysaccharides from Crassostrea hongkongensis. Prep. Biochem. Biotechnol. 2014, 44, 708–724. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Guo, T.; Guo, Y.; Zhang, J.; Li, Y.; Feng, W.; Jiao, B. Characterization and protection on acute liver injury of a polysaccharide MP-I from Mytilus coruscus. Glycobiology 2008, 18, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Xing, R.; Liu, S.; Yu, H.; Wang, P.; Li, C.; Li, P. The synthesis and antioxidant activity of the Schiff bases of chitosan and carboxymethyl chitosan. Bioorg. Med. Chem. Lett. 2005, 15, 4600–4603. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Fan, L.; Ai, L.; Shan, L. Antioxidant activities of polysaccharides from the fruiting bodies of Zizyphus Jujuba cv. Jinsixiaozao. Carbohydr. Polym. 2011, 84, 390–394. [Google Scholar] [CrossRef]
- Francisco, C.G.; Herrera, A.J.; Suárez, E. Intramolecular hydrogen abstraction reaction promoted by alkoxy radicals in carbohydrates. Synthesis of chiral 2,7-dioxabicyclo[2.2.1]heptane and 6,8-dioxabicyclo[3.2.1]octane ring systems. J. Org. Chem. 2002, 67, 7439–7445. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, R.; Tatsumi, Y.; Asano, M.; Kato, K.; Ueno, Y. Effect of metal salts and fructose on the autoxidation of methyl linoleate in emulsions. Agric. Biol. Chem. 1988, 52, 849–850. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Rossi, R.; Giustarini, D.; Milzani, A.; Colombo, R. Protein carbonyl groups as biomarkers of oxidative stress. Clin. Chim. Acta 2003, 329, 23–38. [Google Scholar] [CrossRef]
- Lin, M.T.; Beal, M.F. Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature 2006, 443, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.H.; Dar, K.B.; Anees, S.; Zargar, M.A.; Masood, A.; Sofi, M.A.; Ganie, S.A. Oxidative stress, mitochondrial dysfunction and neurodegenerative diseases; a mechanistic insight. Biomed. Pharmacother. 2015, 74, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Gong, X. Upstream reactive oxidative species (ROS) signals in exogenous oxidative stress-induced mitochondrial dysfunction. Cell Biol. Int. 2009, 33, 658–664. [Google Scholar] [CrossRef] [PubMed]
- Federico, A.; Cardaioli, E.; Da Pozzo, P.; Formichi, P.; Gallus, G.N.; Radi, E. Mitochondria, oxidative stress and neurodegeneration. J. Neurol. Sci. 2012, 322, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Prakash, A.; Dogra, S. Naringin alleviates cognitive impairment, mitochondrial dysfunction and oxidative stress induced by d-galactose in mice. Food Chem. Toxicol. 2010, 48, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Hafez, M.M.; Al-Shabanah, O.A.; Al-Harbi, N.O.; Al-Harbi, M.M.; Al-Rejaie, S.S.; Alsurayea, S.M.; Sayed-Ahmed, M.M. Association between paraoxonases gene expression and oxidative stress in hepatotoxicity induced by CCl4. Oxid. Med. Cell. Longev. 2014, 2014, 893212. [Google Scholar] [CrossRef] [PubMed]
- Fischer, R.; Maier, O. Interrelation of oxidative stress and inflammation in neurodegenerative disease: Role of TNF. Oxid. Med. Cell. Longev. 2015, 2015, 610813. [Google Scholar] [CrossRef] [PubMed]
- Brandes, R.P.; Weissmann, N.; Schröder, K. Nox family NADPH oxidases: Molecular mechanisms of activation. Free Radic. Biol. Med. 2014, 76, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.; Dillin, A. The insulin paradox: Aging, proteotoxicity and neurodegeneration. Nat. Rev. Neurosci. 2008, 9, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, Z.; Peng, L.; Jiang, N.; Liu, Q.; Zhang, E.; Liang, B.; Li, R.; Zhu, H. Lycium barbarum polysaccharide protects human keratinocytes against UVB-induced photo-damage. Free Radic. Res. 2017, 51, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Xiong, C.; Li, Q.; Chen, C.; Chen, Z.; Huang, W. Neuroprotective effect of crude polysaccharide isolated from the fruiting bodies of Morchella importuna against H2O2-induced PC12 cell cytotoxicity by reducing oxidative stress. Biomed. Pharmacother. 2016, 83, 569–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, H.; Zhang, X.; Li, X.; Geng, L.; Zhang, H.; Zhang, Q. Sulfated hetero-polysaccharides protect SH-SY5Y cells from H2O2-induced apoptosis by affecting the PI3K/Akt signaling pathway. Mar. Drugs 2017, 15, 110. [Google Scholar] [CrossRef] [PubMed]
- Gloire, G.; Legrand-Poels, S.; Piette, J. NF-kappaB activation by reactive oxygen species: Fifteen years later. Biochem. Pharmacol. 2006, 72, 1493–1505. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, H.L.; Yang, C.M. Role of redox signaling in neuroinflammation and neurodegenerative diseases. BioMed Res. Int. 2013, 2013, 484613. [Google Scholar] [CrossRef] [PubMed]
- Mayo, K.J.; Cyr, D.M. Protein aggregation and neurodegeneration. Methods 2011, 53, 185–186. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sami, N.; Kashav, T.; Islam, A.; Ahmad, F.; Hassan, M.I. Protein aggregation and neurodegenerative diseases: From theory to therapy. Eur. J. Med. Chem. 2016, 124, 1105–1120. [Google Scholar] [CrossRef] [PubMed]
- Readnower, R.D.; Sauerbeck, A.D.; Sullivan, P.G. Mitochondria, amyloid β, and Alzheimer’s Disease. Int. J. Alzheimers Dis. 2011, 2011, 104545. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.H.; Connor, T.; Stiles, M.; Kama, J.; Lu, Z.; Dorsey, K.; Liebermann, G.; Sapp, E.; Cherny, R.A.; Banks, M.; et al. Cysteine oxidation within N-terminal mutant huntingtin promotes oligomerization and delays clearance of soluble protein. J. Biol. Chem. 2011, 286, 18320–18330. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Prabhakar, M.; Kumar, P.; Deshmukh, R.; Sharma, P.L. Excitotoxicity: Bridge to various triggers in neurodegenerative disorders. Eur. J. Pharmacol. 2013, 698, 6–18. [Google Scholar] [CrossRef] [PubMed]
- Bové, J.; Perier, C. Neurotoxin-based models of Parkinson’s disease. Neuroscience 2012, 211, 51–76. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Mao, J.; Ding, K.; Zhou, Y.; Zeng, X.; Yang, W.; Wang, P.; Zhao, C.; Yao, J.; Xia, P.; et al. Polysaccharides from Ganoderma lucidum promote cognitive function and neural progenitor proliferation in mouse model of Alzheimer’s disease. Stem Cell Rep. 2017, 8, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jiang, L.; Li, X. κ-carrageenan-derived pentasaccharide attenuates Aβ25-35-induced apoptosis in SH-SY5Y cells via suppression of the JNK signaling pathway. Mol. Med. Rep. 2017, 15, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective effects of fucoidan on Aβ25–35 and d-Gal-induced neurotoxicity in PC12 cells and d-Gal-induced cognitive dysfunction in mice. Mar. Drugs 2017, 15, 77. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, C.; Yin, J.; Shen, J.; Wang, H.; Wu, Y.; Jin, H. Fucoidan, a sulfated polysaccharide from brown algae, improves cognitive impairment induced by infusion of Aβ peptide in rats. Environ. Toxicol. Pharmacol. 2012, 33, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Cao, Y.; Chen, L.; Wang, J.; Tian, Q.; Wang, N.; Liu, Z.; Li, J.; Wang, N.; Wang, X.; et al. A polysaccharide from Polygonatum sibiricum attenuates amyloid-β-induced neurotoxicity in PC12 cells. Carbohydr. Polym. 2015, 117, 879–886. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Liao, W.; Fang, J.; Liu, Q.; Hu, M.; Ding, K. A glucan isolated from flowers of Lonicera japonica Thunb. Inhibits aggregation and neurotoxicity of Aβ42. Carbohydr. Polym. 2014, 110, 142–147. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Li, J.; Xia, W.; Geng, M. A marine-derived acidic oligosaccharide sugar chain specifically inhibits neuronal cell injury mediated by beta-amyloid-induced astrocyte activation in vitro. Neurol. Res. 2007, 29, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Lin, C.M.; Chiang, B.H. Protective effects of Angelica sinensis extract on amyloid β-peptide-induced neurotoxicity. Phytomedicine 2008, 15, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Du, X.; Xu, Y.; Dang, L.; Xiang, L.; Zhang, J. The effects of Gouqi extracts on Morris maze learning in the APP/PS1 double transgenic mouse model of Alzheimer’s disease. Exp. Ther. Med. 2013, 5, 1528–1530. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Lai, C.; Ho, Y.; Zee, S.; So, K.; Yuen, W.; Chang, R. Characterization of the effects of anti-aging medicine Fructus lycii on β-amyloid peptide neurotoxicity. Int. J. Mol. Med. 2007, 20, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.; Yu, M.; Yuen, W.; So, K.; Zee, S.; Chang, R. Antagonizing beta-amyloid peptide neurotoxicity of the anti-aging fungus Ganoderma lucidum. Brain Res. 2008, 1190, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Chakrabortee, S.; Liu, Y.; Zhang, L.; Matthews, H.; Zhang, H.; Pan, N.; Cheng, C.; Guan, S.; Guo, D.; Huang, Z.; et al. Macromolecular and small-molecule modulation of intracellular Aβ42 aggregation and associated toxicity. Biochem. J. 2012, 442, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Jia, D.; Rao, C.; Xue, S.; Lei, J. Purification, characterization and neuroprotective effects of a polysaccharide from Gynostemma pentaphyllum. Carbohydr. Polym. 2015, 122, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Liu, M.; Cao, J.; Yao, M.; Lu, Y.; Li, J.; Zhu, X.; Yang, Z.; Wen, A. Protective effects of Lycium barbarum polysaccharide on 6-OHDA-induced apoptosis in PC12 cells through the ROS-NO pathway. Molecules 2014, 20, 293–308. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.; Yang, X. Protective effects of Gynostemma pentaphyllum polysaccharides on PC12 cells impaired by MPP(+). Int. J. Biol. Macromol. 2014, 69, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Lu, J.; Zhang, J.; Xie, J. Protective effects of a polysaccharide from Spirulina platensis on dopaminergic neurons in an MPTP-induced Parkinson’s disease model in C57BL/6J mice. Neural. Regen. Res. 2015, 10, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Wang, H.; Liu, Y.; Lin, S.; Chou, H.; Sheen, L. Immunomodulatory activities of polysaccharides from Chlorella pyrenoidosa in a mouse model of Parkinson’s disease. J. Funct. Foods 2014, 11, 103–113. [Google Scholar] [CrossRef]
- Souza, R.B.; Frota, A.F.; Sousa, R.S.; Cezario, N.A.; Santos, T.B.; Souza, L.M.; Coura, C.O.; Monteiro, V.S.; Cristino Filho, G.; Vasconcelos, S.M.; et al. Neuroprotective effects of sulphated agaran from marine alga Gracilaria cornea in rat 6-hydroxydopamine Parkinson’s disease model: behavioural, neurochemical and transcriptional alterations. Basic Clin. Pharmacol. Toxicol. 2017, 120, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Cui, N.; Wang, P.; Song, S.; Liang, H.; Ji, A. Neuroprotective effect of sulfated polysaccharide isolated from sea cucumber Stichopus japonicus on 6-OHDA-induced death in SH-SY5Y through inhibition of MAPK and NF-κB and activation of PI3K/Akt signaling pathways. Biochem. Biophys. Res. Commun. 2016, 470, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Kuo, H.; Lu, C.; Shen, C.; Tung, S.; Hsieh, M.; Lee, K.; Lee, L.; Chen, C.; Teng, C.; Huang, W.; et al. Hericium erinaceus mycelium and its isolated erinacine A protection from MPTP-induced neurotoxicity through the ER stress, triggering an apoptosis cascade. J. Transl. Med. 2016, 14, 78. [Google Scholar] [CrossRef] [PubMed]
- Meenakshi, S.; Umayaparvathi, S.; Saravanan, R.; Manivasagam, T.; Balasubramanian, T. Neuroprotective effect of fucoidan from Turbinaria decurrens in MPTP intoxicated Parkinsonic mice. Int. J. Biol. Macromol. 2016, 86, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Peng, T.; Yang, S.; Wang, W.; Zhang, Y.; Li, H. Lycium barbarum polysaccharide attenuates the cytotoxicity of mutant huntingtin and increases the activity of AKT. Int. J. Dev. Neurosci. 2016, 52, 66–74. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Wang, S.; Sun, C.; Mao, H.; Di, W.; Ma, X.; Liu, L.; Liu, J.; Wang, F.; Kelly, P.; et al. Investigation into anti-epileptic effect and mechanisms of Ganoderma lucidum polysaccharides in in vivo and in vitro models. Proc. Nutr. Soc. 2015, 74, E65. [Google Scholar] [CrossRef]
- Zhang, J.; An, S.; Hu, W.; Teng, M.; Wang, X.; Qu, Y.; Liu, Y.; Yuan, Y.; Wang, D. The neuroprotective properties of Hericium erinaceus in glutamate-damaged differentiated PC12 cells and an Alzheimer’s disease mouse model. Int. J. Mol. Sci. 2016, 17, 1810. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yang, X.; Jin, G.; Yang, X.; Zhang, Y. Polysaccharides from Pleurotus ostreatus alleviate cognitive impairment in a rat model of Alzheimer’s disease. Int. J. Biol. Macromol. 2016, 92, 935–941. [Google Scholar] [CrossRef] [PubMed]
- Hu, P.; Li, Z.; Chen, M.; Sun, Z.; Ling, Y.; Jiang, J.; Huang, C. Structural elucidation and protective role of a polysaccharide from Sargassum fusiforme on ameliorating learning and memory deficiencies in mice. Carbohydr. Polym. 2016, 139, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; He, D.; Zhang, Y.; Yang, S.; Chen, L.; Wang, S.; Zou, H.; Liao, Z.; Zhang, X.; Wu, M. Sargassum fusiforme polysaccharides activate antioxidant defense by promoting Nrf2-dependent cytoprotection and ameliorate stress insult during aging. Food Funct. 2016, 7, 4576–4588. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.; Yu, M.; Yang, X.; So, K.; Yuen, W.; Chang, R. Neuroprotective effects of polysaccharides from wolfberry, the fruits of Lycium barbarum, against homocysteine-induced toxicity in rat cortical neurons. J. Alzheimers Dis. 2010, 19, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.; Yu, M.; Yik, S.; So, K.; Yuen, W.; Chang, R. Polysaccharides from wolfberry antagonizes glutamate excitotoxicity in rat cortical neurons. Cell. Mol. Neurobiol. 2009, 29, 1233–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haider, A.; Inam, W.; Khan, S.A.; Hifza; Mahmood, W.; Abbas, G. β-glucan attenuated scopolamine induced cognitive impairment via hippocampal acetylcholinesterase inhibition in rats. Brain Res. 2016, 1644, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Yu, J.; Zhao, L.; Ma, N.; Fang, Y.; Pei, F.; Mariga, A.; Hu, Q. Polysaccharides from Flammulina velutipes improve scopolamine-induced impairment of learning and memory of rats. J. Funct. Foods 2015, 18, 411–422. [Google Scholar] [CrossRef]
- Chen, W.; Cheng, X.; Chen, J.; Yi, X.; Nie, D.; Sun, X.; Qin, J.; Tian, M.; Jin, G.; Zhang, X. Lycium barbarum polysaccharides prevent memory and neurogenesis impairments in scopolamine-treated rats. PLoS ONE 2014, 9, e88076. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; He, B. Treatment of d-galactose induced mouse aging with Lycium barbarum polysaccharides and its mechanism study. Afr. J. Tradit. Complement. Altern. Med. 2013, 10, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, J.; Wang, L.; Mao, D. Effects of polygonatum sibiricum polysaccharide on learning and memory in a scopolamine-induced mouse model of dementia. Neural Regen. Res. 2008, 1, 33–36. [Google Scholar]
- Xu, T.; Shen, X.; Yu, H.; Sun, L.; Lin, W.; Zhang, C. Water-soluble ginseng oligosaccharides protect against scopolamine-induced cognitive impairment by functioning as an antineuroinflammatory agent. J. Ginseng Res. 2016, 40, 211–219. [Google Scholar] [CrossRef] [PubMed]
- You, R.; Wang, K.; Liu, J.; Liu, M.; Luo, L.; Zhang, Y. A comparison study between different molecular weight polysaccharides derived from Lentinus edodes and their antioxidant activities in vivo. Pharm. Biol. 2011, 49, 1298–1305. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.; Zhang, Y.; Li, J.; Xia, J.; Chen, X.; Jing, P.; Song, X.; Wang, L.; Wang, Y. Angelica Sinensis polysaccharide prevents hematopoietic stem cells senescence in d-galactose-induced aging mouse model. Stem. Cells Int. 2017, 2017, 3508907. [Google Scholar] [CrossRef] [PubMed]
- Ding, Q.; Yang, D.; Zhang, W.; Lu, Y.; Zhang, M.; Wang, L.; Li, X.; Zhou, L.; Wu, Q.; Pan, W.; et al. Antioxidant and anti-aging activities of the polysaccharide TLH-3 from Tricholoma lobayense. Int. J. Biol. Macromol. 2016, 85, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Guo, L.; Ren, Y.; Wang, B.; Li, R.; Qi, Y.; Yu, H.; Chang, N.; Li, M.; Peng, H. Anti-apoptosis effect of polysaccharide isolated from the seeds of Cuscuta chinensis Lam on cardiomyocytes in aging rats. Mol. Biol. Rep. 2014, 41, 6117–6124. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Nie, S.; Peng, X.; Liu, X.; Li, C.; Chen, Y.; Li, J.; Song, W.; Xie, M. Ganoderma atrum polysaccharide improves age-related oxidative stress and immune impairment in mice. J. Agric. Food Chem. 2012, 60, 1413–1418. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, Z.; Zhang, Z.; Wang, X. Purified Auricularia auricular-judae polysaccharide (AAP I-a) prevents oxidative stress in an ageing mouse model. Carbohydr. Polym. 2011, 84, 638–648. [Google Scholar] [CrossRef]
- Jin, W.; Wang, J.; Jiang, H.; Song, N.; Zhang, W.; Zhang, Q. The neuroprotective activities of heteropolysaccharides extracted from Saccharina japonica. Carbohydr. Polym. 2013, 97, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Tsai, L.H.; Madabhushi, R. Alzheimer’s disease: A protective factor for the ageing brain. Nature 2014, 507, 439–440. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, X. Antioxidant therapies for Alzheimer’s disease. Oxid. Med. Cell. Longev. 2012, 2012, 472932. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhao, B. Natural antioxidants in prevention and management of Alzheimer’s disease. Front. Biosci. (Elite Ed.) 2012, 4, 794–808. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Evans, D.A.; Tangney, C.C.; Bienias, J.L.; Wilson, R.S.; Aggarwal, N.T.; Scherr, P.A. Relation of the tocopherol forms to incident Alzheimer disease and to cognitive change. Am. J. Clin. Nutr. 2005, 81, 508–514. [Google Scholar] [PubMed]
- Mangialasche, F.; Kivipelto, M.; Mecocci, P.; Rizzuto, D.; Palmer, K.; Winblad, B.; Fratiglioni, L. High plasma levels of vitamin E forms and reduced Alzheimer’s disease risk in advanced age. J. Alzheimers Dis. 2010, 20, 1029–1037. [Google Scholar] [CrossRef] [PubMed]
- Urbányi, Z.; Forrai, E.; Sárvári, M.; Likó, I.; Illés, J.; Pázmány, T. Glycosaminoglycans inhibit neurodegenerative effects of serum amyloid P component in vitro. Neurochem. Int. 2005, 46, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Jin, L. The structure-activity relationship of glycosaminoglycans and their analogues with β-amyloid peptide. Protein Pept. Lett. 2016, 23, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Ariga, T.; Miyatake, T.; Yu, R.K. Role of proteoglycans and glycosaminoglycans in the pathogenesis of Alzheimer’s disease and related disorders: Amyloidogenesis and therapeutic strategies—A review. J. Neurosci. Res. 2010, 88, 2303–2315. [Google Scholar] [CrossRef] [PubMed]
- Abeliovich, A.; Gitler, A.D. Defects in trafficking bridge Parkinson’s disease pathology and genetics. Nature 2016, 539, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, H.H. 2015 Update on Parkinson disease. Clevel. Clin. J. Med. 2015, 82, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Wood, L.D. Clinical review and treatment of select adverse effects of dopamine receptor agonists in Parkinson’s disease. Drugs Aging 2010, 27, 295–310. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, H.; Jin, W.; Zhang, H.; Zhang, Q. Structure-activity relationship of sulfated hetero/galactofucan polysaccharides on dopaminergic neuron. Int. J. Biol. Macromol. 2016, 82, 878–883. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Shi, Y.; Sheng, J.; Hu, Q. In vivo immunomodulatory activity of polysaccharides derived from Chlorella pyrenoidosa. Eur. Food Res. Technol. 2006, 224, 225–228. [Google Scholar] [CrossRef]
- Panaro, M.A.; Cianciulli, A. Current opinions and perspectives on the role of immune system in the pathogenesis of Parkinson’s disease. Curr. Pharm. Des. 2012, 18, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Kanemaru, K. Immunotherapy targeting misfolded proteins in neurodegenerative disease. Brain Nerve 2013, 65, 469–474. [Google Scholar] [PubMed]
- Walker, F.O. Huntington’s disease. Lancet 2007, 369, 218–228. [Google Scholar] [CrossRef]
- Berry-Kravis, E. Huntington’s disease: Genetics. Encycl. Mov. Disord. 2010, 31–36. [Google Scholar] [CrossRef]
- Dayalu, P.; Albin, R.L. Huntington disease: Pathogenesis and treatment. Neurol. Clin. 2015, 33, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Ehrnhoefer, D.E.; Duennwald, M.; Markovic, P.; Wacker, J.L.; Engemann, S.; Roark, M.; Legleiter, J.; Marsh, J.L.; Thompson, L.M.; Lindquist, S.; et al. Green tea (−)-epigallocatechin-gallate modulates early events in huntingtin misfolding and reduces toxicity in Huntington’s disease models. Hum. Mol. Genet. 2006, 15, 2743–2751. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Li, H.; Zhang, J.; Yang, F.; Huang, A.; Deng, J.; Liang, M.; Ma, F.; Hu, M.; Huang, Z. Salidroside protects Caenorhabditis elegans neurons from polyglutamine-mediated toxicity by reducing oxidative stress. Molecules 2014, 19, 7757–7769. [Google Scholar] [CrossRef] [PubMed]
- Faber, P.W.; Voisine, C.; King, D.C.; Bates, E.A.; Hart, A.C. Glutamine/proline-rich PQE-1 proteins protect Caenorhabditis elegans neurons from huntingtin polyglutamine neurotoxicity. PNAS 2002, 99, 17131–17136. [Google Scholar] [CrossRef] [PubMed]
- Ezza, H.S.A.; Khadrawyb, Y.A. Glutamate excitotoxicity and neurodegeneration. J. Mol. Genet. Med. 2014, 8, 4. [Google Scholar] [CrossRef]
Figure 1.
Pharmacological intervention of neurodegeneration by food-derived antioxidant polysaccharides. A number of extrinsic and intrinsic stresses such as proteotoxic stress, inflammatory stress and chemical interruption can stimulate oxidative stress through impairing the function of antioxidant system and mitochondria. Increase of oxidative stress can promote pathogenic protein aggregation and inflammation, eventually leading to neuronal injury, death and dysfunction via multiple biochemical pathways (solid line). However, food-derived antioxidant polysaccharides can exert beneficial effects to suppress neurodegeneration via attenuating oxidative, inflammatory and proteotoxic stresses and regulating stress-related signaling (dashed line).
Figure 1.
Pharmacological intervention of neurodegeneration by food-derived antioxidant polysaccharides. A number of extrinsic and intrinsic stresses such as proteotoxic stress, inflammatory stress and chemical interruption can stimulate oxidative stress through impairing the function of antioxidant system and mitochondria. Increase of oxidative stress can promote pathogenic protein aggregation and inflammation, eventually leading to neuronal injury, death and dysfunction via multiple biochemical pathways (solid line). However, food-derived antioxidant polysaccharides can exert beneficial effects to suppress neurodegeneration via attenuating oxidative, inflammatory and proteotoxic stresses and regulating stress-related signaling (dashed line).


{kind=link}
{kind=link}
Table 1.
The antioxidant activities and mechanisms of food-derived polysaccharides.
| Source | Polysaccharide | Test Model | Protective Effect | Potential Mechanism | Ref. |
|---|---|---|---|---|---|
| Vegetables | |||||
| Zizania latifolia | ZLPs-W | In vitro assays | Scavenging activity against DPPH and ·OH | [24] | |
| Daucus carota | CWSP | In vitro assays | Scavenging activity against DPPH, reducing power, prevention of β-carotene bleaching | Ferrous chelating ability | [25] |
| Cucurbita maxima Duchesne | WSP | In vitro assays | Scavenging activity against DPPH, inhibition of ascorbic acid oxidation | SOD-like activity | [26] |
| Solanum tuberosum | PPPWs | In vitro assays | Scavenging activity against DPPH and ABTS, reducing power, total antioxidant capacity | [27] | |
| Potentilla anserine | PAP | H2O2-exposed murine splenic lymphocytes | Apoptosis rate↓ | [28] | |
| Psidium guajava | PS-PGL | In vitro assays; H2O2-exposed Vero cells and zebrafish | Scavenging activity against DPPH, ·OH and alkyl radicals in vitro; Cell viability↑, DNA fragmentation↓, nuclear condensation and morphological disruption↓ in Vero cells; Survival↑, heart-beating rate↓, cell death↓ in zebrafish embryos | ROS level↓ in Vero cells; ROS level↓, MDA content↓ in zebrafish embryos | [29] |
| Fruits | |||||
| Malus pumila | APPS | In vitro assays | Scavenging activity against DPPH, O2−· and ·OH, reducing power | [30] | |
| Diospyros kaki L. | PFP | In vitro assays | Scavenging activity against DPPH, O2−· and ·OH, reducing power | [31] | |
| Seed watermelon | SWP | H2O2-exposed PC12 cells | Cell viability↑, LDH release↓ | ROS level↓, 8-OHdG content↓, caspase-3 and caspase-9 activities↓, MMP↑ | [32] |
| Cereals and Beans | |||||
| Rice bran | RBP2 | In vitro assays | Scavenging activity against DPPH, O2−·, ·OH and ABTS, reducing power | Ferrous chelating ability | [33] |
| Wheat bran | Feruloyl oligosaccharides | AAPH-exposed human erythrocytes | Erythrocyte hemolysis↓ | GSH level↓, MDA content↓, PCG level↓ | [34] |
| Glycine max (L.) Merr. | MSF | In vitro assays | Scavenging activity against ABTS, reducing power | [35] | |
| Cicer arietinum L. hull | CHPS | In vitro assays; H2O2-exposed PC12 cells | Scavenging activity against ABTS, DPPH O2−; reducing power in vitro; Cell viability↑ | [36] | |
| Herbs | |||||
| Dioscorea opposita | Yam polysaccharide | In vitro assays | Scavenging activity against O2−· and ·OH | [37] | |
| Epimedium brevicornum Maxim. | EbPS-A1 | In vitro assays; PQ-exposed C. elegans | Scavenging activity against DPPH and ·OH in vitro; Survival rate↑ in C. elegans | ROS level↓, MDA content↓, SOD and CAT activities↑ in C. elegans | [38] |
| Chuanminshen violaceum | CVPS | In vitro assays; d-Gal-treated ICR mice | Scavenging activity against DPPH, O2−· and ·OH in vitro; Body weights and spleen indices↑ in mice | Activities and mRNA levels of Mn-SOD, Cu/Zn-SOD, GPx and CAT↑, MDA content↓ in mouse liver, heart and brain | [39] |
| Radix Rehmanniae | RRPs | UV-irradiated mice | GSH level↑, SOD, CAT and GPx activities↑, MDA content↓, IL-2, IL-4 and IL-10 levels↑ | [40] | |
| Lycium barbarum | LBPs | H2O2-exposed SRA01/04 cells | Cell viability↑, apoptotic rate↓, ratio of ageing cells↓, G0/G1 cell cycle phase arrest↓ | ROS level↓, MMP↑, Bcl-2 protein level↑, Bax protein level↓, MDA content↓, SOD activity↑, GSH level↑ | [41] |
| Angelica sinensis | ASP | H2O2-exposed PC12 cells; SD rats with middle cerebral artery occlusion | Cell viability↑, apoptosis rate↓ in PC12 cells; Number of microvessels in rat brain↑ | ROS level↓, MMP↑ in PC12 cells; SOD and GPx activities↑ in rat cortex | [42] |
| Sophora subprosrate | SSP | PCV-2 infection RAW264.7 cells | Activities of Total-SOD, Cu/Zn-SOD and Mn-SOD↑, mRNA levels of Mn-SOD↑ and NOX2↓, NOX2 protein level↓, MMP↑ | [43] | |
| Cynomorium songaricum Rupr. | CSP | H2O2-exposed PC12 cells | Cell viability↑, ratio of sub G1and S phase↓, ratio of G2/M phase↑, apoptosis rate↓, LDH release↓ | ROS level↓,MDA content↓, 8-OHdG content↓, SOD and GPx activities↑, capase-3 and capase-9 activities↓ | [44] |
| Tea | |||||
| Black tea | BTPS | In vitro assays | Scavenging activity against DPPH and ·OH | [45] | |
| Green tea | TPS1 | In vitro assays | Scavenging activity against DPPH, O2−· and ·OH, ferrous chelating ability, reducing power, total antioxidant capacity, inhibition of lipid hydroperoxide | [46] | |
| Gynostemma pentaphyllum Makino | GPMMP | Cyclophosphamide-treated C57BL/6 mice | Spleen and thymus indices↑, CD4+ T lymphocyte counts↑, total antioxidant capacity↑ | CAT, SOD and GPx activities↑, MDA content↓, GSH level↑, IL-2 level in sera and spleen↑ | [47] |
| Nuts | |||||
| Juglans regia L. | SJP | In vitro assays | Scavenging activity against DPPH, ·OH and ABTS, reducing power | [48] | |
| Ginkgo biloba L. | GNP | In vitro assays; Hyperlipemia mice | Scavenging activity against DPPH, O2−· and ·OH in vitro | CAT, SOD and GPx activities↑, MDA content↓ in mouse serum and liver | [49] |
| Other Plants | |||||
| Zizyphus jujuba Mill | ZJPa | In vitro assays | Scavenging activity against O2−· and ·OH | Ferrous chelating ability | [50] |
| Aloe barbadensis Miller | GAPS-1 and SAPS-1 | In vitro assays | Scavenging activity against O2−·, ·OH and H2O2, reducing power, MDA content↓ | Ferrous chelating ability | [51] |
| Anoectochilus roxburghii | ARPT | CCl4-treated Kunming mice | Hepatocyte necrosis↓, serum alanine transaminase and aspartate transaminase activities↓ | MDA level↓, SOD, CAT and GPx activities↑, GSH level↓, mRNA levels of TNF-α, IL-6 and Bax↓, protein levels of TNF-α, IL-6, NF-κB and cleaved-caspase 3↓ in liver | [52] |
| Opuntia dillenii Haw | CP | H2O2-exposed PC12 cells | Cell viability↑, LDH release↓, apoptosis rate↓ | ROS level↓, ratio of Bax/Bcl-2 mRNA level↑ | [53] |
| Camellia oleifera Abel | SCP1 | In vitro assays; PQ-exposed C. elegans | Scavenging activity against O2−· and ·OH in vitro; Survival rate↑ in C. elegans | Ferric chelating ability in vitro; SOD, CAT and GPx activities↑, MDA content↓ in C. elegans | [54] |
| Taraxacum officinale | TOP2 | LPS or t-BHP-exposed RAW 264.7 cells | NO production↓ in LPS-exposed cells; Cell viability↑ in t-BHP-exposed cells | Protein levels of TNF-α, p-IκBα, p-p65, p-Akt, iNOS and heme oxygenase 1↓ | [55] |
| Mushrooms | |||||
| Ganoderma lucidum | G. lucidum polysaccharide | Isoproterenol-treated albino rats | Creatinine kinase and LDH activities↓ in serum, cardiac muscle fibers with mild hyalinization | ROS level↓, MDA content↓, SOD and GPx activities↑, GSH level↑, activities of Krebs cycle dehydrogenases and mitochondrial complexes↑, MMP↑ | [56] |
| Lentinus edodes, Ganoderma applanatum, Trametes versicolor | Mushroom polysaccharides | In vitro assays | Scavenging activity against DPPH, reducing power, inhibition of linoleic acid peroxidation | Ferric chelating ability | [57] |
| Dictyophora indusiata | DiPS | PQ-exposed C. elegans | Survival rate↑ | ROS level↓, SOD activity↑, MDA content↓, MMP↑, ATP content↑, DAF-16 activation↑ | [58] |
| Other Fungi | |||||
| Auricularia auricula | AAP1 | In vitro assays; PQ or H2O2-exposed C. elegans | Scavenging activity against DPPH, O2−· and ·OH, reducing power in vitro; Survival rate↑ in C. elegans | Ferric chelating ability in vitro; ROS level↓, SOD and CAT activities↑ in C. elegans | [59] |
| Tremella fuciformis | TP | UV-irradiated SD rats | Water and collagen content↑, glycosaminoglycan↓, endogenous collagen breakdown↓, ratio of type I/III collagen↑ in rat skin | SOD, GPx and CAT activities↑ | [60] |
| Algae | |||||
| Porphyra haitanesis | P. haitanesis polysaccharide | In vitro assays; H2O2-exposed rat erythrocytes and liver microsome | Scavenging activity against O2−· and ·OH in vitro; Erythrocyte hemolysis↓; lipid peroxidation of rat liver microsome↓ | [61] | |
| Laminaria japonica | LJPA-P3 | In vitro assays | Oxygen radical absorbance capacity, scavenging activity against ABTS | [62] | |
| Fucus vesiculosus | F. vesiculosus polysaccharide | In vitro assays | Ferric reducing antioxidant power | [63] | |
| Ulva pertusa | U. pertusa polysaccharide | In vitro assays | Scavenging activity against O2−· and ·OH, reducing power | Ferric chelating ability | [64] |
| Brown seaweed | Fucoidan | UV-irradiated HS68 cells | ROS level↓, MDA content↓, GSH level↑ | [65] | |
| Nostoc commune | Nostoc polysaccharide | In vitro assays; PQ-exposed C. elegans | Scavenging activity against O2−· and ·OH in vitro; Survival rate↑ in C. elegans | SOD, CAT and GPx activities↑, MDA content↓ in C. elegans | [66] |
| Milkproducts | |||||
| Milk fermented with lactic acid bacteria | Exopolysaccharides | UV-irradiated hairless mice | Erythema formation, dryness and epidermal proliferation, cyclobutane pyrimidine dimers↓ in mouse skin | mRNA levels of xeroderma pigmentosum complementation group A↑, ratio of mRNA levels of IL10/IL12α and IL10/IFN-γ↓ in mouse skin | [67] |
| Wine | |||||
| Red wine | PS-SI | In vitro assays | Scavenging activity against ·OH, oxygen radical absorbance capacity | [68] | |
| Probiotics | |||||
| Bifidobacterium animalis RH | EPS | In vitro assays; d-Gal-treated Kunming mice | Inhibition of linoleic acid peroxidation, total antioxidant capacity, scavenging activity against DPPH, O2−· and ·OH in vitro | Total antioxidant capacity, SOD, CAT and GPx activities↑, MDA content↓ in serum, GST activity and MDA content↓ in liver, MAO activity and lipofuscin level↓ in brain | [69] |
| Bifidobacterium bifidum WBIN03, Lactobacillus plantarum R31 | B-EPS and L-EPS | In vitro assays; H2O2-exposed rat erythrocytes | Scavenging activity against DPPH, O2−· and ·OH, inhibition of lipid peroxidation in vitro; Erythrocyte hemolysis↓ | [70] | |
| Meat | |||||
| Haliotis discus hannai Ino | ASP-1 | In vitro assays | Scavenging activity against O2−· | [71] | |
| Crassostrea hongkongensis | CHPs | In vitro assays | Scavenging activity against DPPH, ·OH and ABTS, inhibition of linoleic acid peroxidation | [72] | |
| Mytilus coruscus | MP-I | CCl4-treated Kunming mice | Serum alanine transaminase and aspartate transaminase levels↓, necrosis of liver cells↓, immigration of inflammatory cells↓ | MDA content↓, SOD activity↑ in liver | [73] |
AAPH, 2,2′-Azobis(2-amidinopropane) dihydrochloride; ABTS, 2,2′-Azino-bis(3-ethylbenzothiazoline-6-sulfonic acid); CAT, catalase; CCl4, carbon tetrachloride; Cu/Zn-SOD, copper-zinc superoxide dismutase; d-Gal, d-galactose; DPPH, 2,2-diphenyl-1-picrylhydrazyl radical; GPx, glutathione peroxidase; GSH, glutathione; GST, glutathione S-transferase; H2O2, hydrogen peroxide; HS68 cells, human foreskin fibroblast line; IFN-γ, interferon-γ; IκBα, NF-κB inhibitor α; ILs, interleukins; iNOS, inducible nitric oxide synthase; LDH, lactate dehydrogenases; LPS, lipopolysaccharide; MAO, monoamine oxidase; MDA, malondialdehyde; MMP, mitochondrial membrane potential; Mn-SOD, manganese superoxide dismutase; NF-κB, nuclear factor-κB; NO, nitric oxide; NOX2, cytochrome b-245β chain; O2−·, superoxide anion; ·OH, hydroxyl radical; RAW 264.7 cells, murine macrophage cell line; PC12 cells, rat pheochromocytoma cell line; PCG, protein carbonyl group; PCV-2, porcine circovirus type 2; PQ, paraquat; ROS, reactive oxygen species; SOD, superoxide dismutase; SRA01/04 cells, SV40 T-antigen-transformed human lens epithelial cell line; t-BHP, tert-Butyl hydroperoxide; TNF-α, tumor necrosis factor α; UV, ultraviolet; 8-OHdG, 8-hydroxy-2’-deoxyguanosine.
Table 2.
Protective effects and mechanisms of food-derived antioxidant polysaccharides in neurodegeneration models.
Table 2.
Protective effects and mechanisms of food-derived antioxidant polysaccharides in neurodegeneration models.
| Source | Polysaccharide | Test Model | Protective Effect | Potential Mechanism | Ref. |
|---|---|---|---|---|---|
| Ganoderma lucidum | GLP | APP/PS1 transgenic mice | Learning and memory in MWM↑, neural progenitor cell proliferation↑ | Aβ deposits↓, protein levels of p-FGFR1, p-ERK and p-Akt↑ | [101] |
| Marine red algae | KCP | Aβ(25–35)-exposed SH-SY5Y cells | Cell viability↑, apoptosis rate↓ | Protein level of cleavage caspase 3↓, JNK signaling activation↓ | [102] |
| Undaria pinnatifida sporophylls | Fucoidan | Aβ(25–35) and d-Gal-exposed PC12 cells; d-Gal treated ICR mice | Cell viability↑, apoptosis rate↓ in PC12 cells; Learning and memory in MWM↑ | Protein levels of cleaved caspase-3, caspase-8 and caspase-9↓, cytochrome c release↓, SOD activity↑, GSH level↑ in PC12 cells; Aβ deposits in hippocampus↓, SOD activity and GSH level↑ in serum, Ach content↑, ChAT activity↑ and AChE activity↓ in brain | [103] |
| Laminaria japonica Aresch. | Fucoidan | Aβ40-treated SD rats | Learning and memory in MWM, single-trial passive avoidance and eight-arm radial maze task↑ | Ach content↑, ChAT activity↑, AChE activity↓, SOD and GPx activities↑, MDA content↓, Bax/Bcl-2 protein level ratio↓, cleaved caspase-3 protein level↓ in hippocampus | [104] |
| Polygonatum sibiricum | PS-WNP | Aβ(25–35)-exposed PC12 cells | Cell viability↑, apoptosis rate↓ | Bax/Bcl-2 protein level ratio↓, MMP↑, cytochrome c release↓, cleaved caspase-3 protein level↓, caspase-3 activity↓, p-Akt protein level↑ | [105] |
| Lonicera japonica Thunb. | LJW0F2 | Aβ42-exposed SH-SY5Y cells | Cell viability↑ | Aβ42 aggregates↓ | [106] |
| Echlonia Kurome Okam. | AOSC | Aβ(25–35)-exposed SH-SY5Y cells | Cell viability↑, apoptosis rate↓, activation of astrocytes↓, cell redox activity↑ | ROS level↓, TNF-α and IL-6 level↓, calcium influx in astrocytes↓ | [107] |
| Angelica sinensis | AS | Aβ(25–35)-exposed Neuro 2A cells | Cell viability↑ | ROS level↓, GSH level↑, MMP↑, mitochondria mass↑, TBARS content↓, autophagosomes or residual bodies↓ | [108] |
| Lycium barbarum | L. barbarum polysaccharide | APP/PS1 transgenic mice | Learning and memory in MWM↑ | Aβ deposits in hippocampus↓ | [109] |
| Lycium barbarum | LBP-III | Aβ(25–35)-exposed rat primary cortical neurons | Maintain neurite fasciculation and neuron integrity | Caspase-3 and caspase-2 activities↓, p-PKR protein level↓ | [110] |
| Ganoderma lucidum | GLA | Aβ(25–35)- or Aβ42-exposed rat primary cortical neurons | Apoptosis rate↓, synaptophysin immunoreactivity↑ | DEVD-cleavage activity↓, protein levels of p-JNK, p-c-Jun, and p-p38↓ | [111] |
| Rubia cordifolia L. | PS5 | T-REx293 cells | Cell viability↑ | Aβ42-EGFP aggregates↓ | [112] |
| Dictyophora indusiata | DiPS | C. elegans CL2355 | Survival rate↑, chemotaxis index↑ | ROS level↓ | [58] |
| Gynostemma pentaphyllum Makino | GPP1 | Aβ(25–35)-exposed PC12 cells | Cell viability↑, LDH release↓, DNA fragmentation↓ | ROS level↓, MDA content↓, SOD activity↑, GSH level↑, Calcium overload↓, MMP↑, Bcl-2 protein level↑, protein levels of Bax, cytochrome c and cleaved caspase-3↓ | [113] |
| Lycium barbarum L. | LBP | 6-OHDA-exposed PC12 cells | Cell viability↑, nuclear morphology changes↓, apoptosis rate↓ | ROS and NO levels↓, calcium overload↓, protein-bound 3-nitrotyrosine level↓, protein levels of nNOS, iNOS and cleaved caspase-3↓ | [114] |
| Gynostemma pentaphyllum Makino | GP | MPP+-exposed PC12 cells | Cell viability↑, LDH release↓, apoptosis rate↓ | Cytochrome c release↓, caspase-3 and caspase-9 activities↓, Bax/Bcl-2 protein level ratio↓, protein levels of cleaved caspase-3 and poly (ADP-ribose) polymerase↓ | [115] |
| Spirulina platensis | PSP | MPTP-treated C57BL/6J mice | Number of TH-immunoreactive neurons and DAT binding ratio in the substantia nigra pars compacta↑ | TH and DAT mRNA levels in substantia nigra↑, SOD and GPx activity↑ in serum and midbrain | [116] |
| Chlorella pyrenoidosa | CPS | MPTP-treated C57BL/6J mice | Body weight↑, movement in pole test and gait test↑ | Contents of DA, DOPAC and HVA↑, ratio of DOPAC and HVA to DA↓, TH mRNA level↑, striatal Emr1 mRNA level↓, TNF-α, IL-1β and IL-6 levels in serum↓, d-amino acid oxidase and secretory immunoglobulin A levels↑ | [117] |
| Gracilaria cornea J. Agardh | SA-Gc | 6-OHDA-treated Wistar rats | Locomotor performance in OFT, rotarod and apomorphine-induced rotation test↑, weight gain↑ | DA and DOPAC content↑, NO2/NO3 and GSH level↑ in brain, p65, iNOS and IL1β mRNA levels↓, BDNF mRNA level↑ | [118] |
| Stichopus japonicus | SJP | 6-OHDA-exposed SH-SY5Y cells | Cell viability↑, apoptosis rate↓, LDH release↓ | SOD activity↑, ROS level↓, NO release↓, MDA content↓, MMP↑, cytochrome c release↓, percentage of cells in S phase↑, Bax/Bcl-2 protein level ratio↓, protein levels of Cyclin D3, p-p53, p-p38, p-JNK1/2, p-p65, iNOS and p-IκB↓, cleaved caspase-9/caspase-9 and cleaved caspase-3/caspase-3 protein level ratio↓, p-Akt and IκB protein levels↑ | [119] |
| Hericium erinaceus | EA | MPTP-treated C57BL/6 mice | Apoptosis rate↓, number of normal neurons↑, motor function in RT↑ | Nitro-tyrosine and 4-HNE level↓, dopamine, NGF, and GSH level↑, protein levels of Fas, p-JNK1/2, p-p38, DNA damage inducible transcript 3, NF-κB and p65↓ | [120] |
| Epimedium brevicornum Maxim. | EbPS-A1 | C. elegans HA759 | Avoidance index↑ | ROS level↓, MDA content↓, SOD and CAT activities↑ | [38] |
| Turbinaria decurrens | TD fucoidan | MPTP-treated C57BL/6 mice | Motor performance in OFT, Narrow beam walking and RT↑, nigral TH immunoreactivity↑ | DA, DOPAC, and HVA content↑, TBARS level↓, GSH level↑, SOD and CAT activities↓, GPx activity↑, TH and DAT protein levels↑ | [121] |
| Lycium barbarum | LBP | HEK293-160Q cells; HD-related transgenic mice | Cell viability↑ in HEK293 cells; Survival rate↑, weight gain↑, motor performance in RT↑ in mice | Soluble and aggregated huntingtin levels↓, caspase-3 activity↓, p-Akt/Akt and p-GSK3β/ GSK3β protein levels↑ in HEK293 cells; Mutant huntingtin level↓, p-Akt/Akt and p-GSK3β/ GSK3β protein levels↑ in mouse brain | [122] |
| Ganoderma lucidum | GLP | Kainic acid-treated Wistar rats | Frequency of epilepsy↓ | CaMK II level↑, ERK1/2 level↓, calcium turnover↓, Caveolin-1 positive cells↑, NF-κB positive cells↓ | [123] |
| Hericium erinaceus | HE | l-Glu-exposed PC12 cells; AlCl₃ and d-Gal-treated Balb/c mice | Differentiation rate↑, cell viability↑, apoptosis rate↓ in PC12 cells; learning, memory and locomotor in MWM, Autonomic activities and RT↑ | β-tubulin III protein level↑, MMP↑, calcium overload↓, ROS level↓ in PC12 cells; Ach and ChAT contents in mouse serum and hypothalamus↑ | [124] |
| Pleurotus ostreatus | POP | d-Gal and AlCl3-treated Wistar rats | Learning and memory in MWM and SDT↑, hippocampal impairment↓ | AchE activity↓, in hippocampus, MDA content↓, SOD, GPx and CAT activities↑ in hippocampus, liver and serum, protein levels of APP, Aβ, BACE1 and p-tau↓, Protein phosphatase 2 protein level↑ | [125] |
| Sargassum fusiforme | SFPS65A | SCO-, ethanol- and sodium nitrite-treated ICR mice | Learning and memory in SDT↑ | [126] | |
| Sargassum fusiforme | SFPS | d-Gal-treated ICR mice | CAT and SOD activities↑, MDA content in hearts and MAO in brains↓, protein levels of Nrf2, Bcl-2, p21 and JNK1/2↑, mRNA levels of Nrf2, Cu/Zn-SOD, Mn-SOD, glutamate cysteine ligase and GPX1↑, voltage dependent anion channel 1 protein level↓ | [127] | |
| Lycium barbarum | LBA | Homocysteine-exposed cortical neurons | Cell viability↑, apoptosis rate↓ | LDH release and caspase-3 activity↓, p-tau-1 protein level↑, cleaved-tau protein level↓, p-ERK1/2 and p-JNK protein levels↓ | [128] |
| Lycium barbarum | LBA | l-Glu- or NMDA-exposed cortical neurons | Cell viability↑, maintained their integrity and fasciculation of neurites | LDH release and caspase-3 activity↓, p-JNK-1/JNK protein level ratio↓ | [129] |
| Saccharomyces cerevisiae | β-glucan | SCO-treated SD rats | Learning, memory, and locomotor in MWM and PTT↑ | AChE activity↓ | [130] |
| Flammulina velutipes | FVP | SCO-treated Wistar rats | Learning and memory in MWM and PTT↑ | SOD and GPx activities↑, TBARS level↓, Ach, 5-HT, DA and NE content↑, ChAT activity↑, AChE activity↓, connexin 36 and p-CaMK II protein level↑ in hippocampus and cerebral cortex | [131] |
| Lycium barbarum | LBPs | SCO-treated SD rats | Learning and memory in MWM, NOR and OLR↑, cell proliferation and neuroblast differentiation in dentate gyrus↑ | SOD and GPX activities↑, MDA content↓, Bax/Bcl-2 protein level ratio↓ in hippocampus | [132] |
| Lycium barbarum | LBP | d-Gal-treated Kunming mice | Weight gain↑, learning and memory in Jumping test↑, thymus and spleen indices↑ | Lipid peroxidation, lipofuscin and MAO-B contents↓ in brain | [133] |
| Polygonatum sibiricum | PSP | SCO-treated Kunming mice | Learning and memory in SDT and Memory test↑ | SOD and GPx activities↑, MDA content↓ | [134] |
| Panax ginseng | WGOS | SCO-treated ICR mice | Learning and memory in MWM and NOR↑ | mRNA levels of GFAP, IL-1β and IL-6↓ in hippocampus, number of GFAP-positive cells↓ in hippocampal subregions | [135] |
| Lentinus edodes | LT2 | d-Gal-treated Kunming mice | Erythrocyte membrane fluidity↑ | SOD and GPx activities↑ in liver, heart and brain | [136] |
| Angelica sinensis | ASP | d-Gal-treated C57BL/6J mice | Percentage of ageing cells↓ | Advanced glycation end-product level in serum↓, ROS level↓, TAOC content↑, 8-OHDG content↓, 4-HNE level↓, protein levels of H2A histone family member X, p16, p21, p53, β-catenin, p-GSK-3β and transcription factor 4↓, mRNA levels of p16, p21 and β-catenin↓, GSK-3β protein level↑ | [137] |
| Tricholoma lobayense | TLH-3 | t-BHP-exposed HELF cells; d-Gal-treated Kunming mice | Cell viability↑, percentage of ageing cells↓, ratio of G0/G1phase↓, nucleic morphological changes↓ in HELF cells | ROS level↓, in HELF cells; SOD and CAT activities↑, MDA content↓, in mouse liver and serum | [138] |
| Cuscuta chinensis Lam | PCCL | d-Gal-treated SD rats | Apoptosis rate of cardiomyoctyes↓ | Calcium overload↓, Bax/Bcl-2 protein level ratio↓, caspase-3 activity↓, cytochrome c release↓ | [139] |
| Ganoderma atrum | PSG-1 | d-Gal-treated Kunming mice | Weight gain↑, lymphocyte proliferation↑ | MDA content↓, SOD, CAT and GPx activities↑, GSH level↑, GSSG level↓ in liver, brain and spleen | [140] |
| Auricularia auricula-judae | APP 1-a | d-Gal-treated Kunming mice | Spleen and thymus indexes↑ | MDA content↓, SOD and GPx activities↑ in liver, serum and heart | [141] |
| Saccharina japonica | DJ0.5 | 6-OHDA-exposed MES 23.5 cells and SH-SY5Y cells | Cell viability↑ | [142] |
Aβ, amyloid-β peptide; Ach, acetylcholine; AChE, acetylcholinesterase; APP, amyloid precursor protein; BACE1, β-secretase 1; BDNF, brain-derived neurotrophic factor; CaMK II, calmodulin-dependent protein kinase II; CAT, catalase; ChAT, choline acetyltransferase; CL2355, a nematode that pan-neuronally expresses Aβ42; Cu/Zn-SOD, copper-zinc superoxide dismutase; d-Gal, d-galactose; DA, dopamine; DAT, dopamine transporter; DOPAC, 3,4-Dihydroxyphenylacetic acid; FGFR1, fibroblast growth factor receptor 1; GFAP, glial fibrillary acid protein; GPx, glutathione peroxidase; GSH, glutathione; GSK-3β, glycogen synthase kinase-3β; GSSG, glutathione disulfide; HA759, a nematode that expresses HtnQ150 in ASH neurons; HEK293 cells, human embryonic kidney cell line; HELF cells, human embryonic lung fibroblast line; HVA, homovanillic acid; IκB, NF-κB inhibitor; iNOS, inducible nitric oxide synthase; LDH, lactate dehydrogenases; l-Glu, l-glutamate; MAO, monoamine oxidase; MES 23.5 cells, rodent mesencephalic neuronal cell line; MDA, malondialdehyde; MMP, mitochondrial membrane potential; MPTP, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine; MPP+, 1-methyl-4-phenylpyridinium; MWM, Morris water maze; NE, norepinephrine; Neuro 2A cells, murine neuroblastoma cell line; NMDA, N-methyl-d-aspartate; nNOS, neuronal nitric oxide synthase; NO, nitric oxide; NOR, novel object recognition; Nrf2, nuclear factor erythroid 2-related factor 2; OLR, object location recognition; OFT, open field test; PC12 cells, rat pheochromocytoma cell line; PS1, presenilin-1; PTT, probe trial test; ROS, reactive oxygen species; RT, Rotarod test; SCO, scopolamine; SDT, step-down test; SH-SY5Y, human neuroblastoma cell line; SOD, superoxide dismutase; TAOC, total antioxidant capacity; TBARS, thiobarbituric acid reactive substances; TH, tyrosine hydroxylase; TNF-α, tumor necrosis factor α; T-REx293, human embryonic kidney cell line transiently transfected with Aβ42-EGFP; t-BHP, tert-butylhydroperoxide; 4-HNE, 4-hydroxynonenal; 5-HT, 5-hydroxytryptamine; 6-OHDA, 6-hydroxydopamine; 8-OHDG, 8-hydroxydeoxyguanosine.
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, H.; Ding, F.; Xiao, L.; Shi, R.; Wang, H.; Han, W.; Huang, Z. Food-Derived Antioxidant Polysaccharides and Their Pharmacological Potential in Neurodegenerative Diseases. Nutrients 2017, 9, 778. https://doi.org/10.3390/nu9070778
AMA Style
Li H, Ding F, Xiao L, Shi R, Wang H, Han W, Huang Z. Food-Derived Antioxidant Polysaccharides and Their Pharmacological Potential in Neurodegenerative Diseases. Nutrients. 2017; 9(7):778. https://doi.org/10.3390/nu9070778
Chicago/Turabian StyleLi, Haifeng, Fei Ding, Lingyun Xiao, Ruona Shi, Hongyu Wang, Wenjing Han, and Zebo Huang. 2017. "Food-Derived Antioxidant Polysaccharides and Their Pharmacological Potential in Neurodegenerative Diseases" Nutrients 9, no. 7: 778. https://doi.org/10.3390/nu9070778
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.