Nitrates and Glucosinolates as Strong Determinants of the Nutritional Quality in Rocket Leafy Salads



Abstract
:1. Introduction
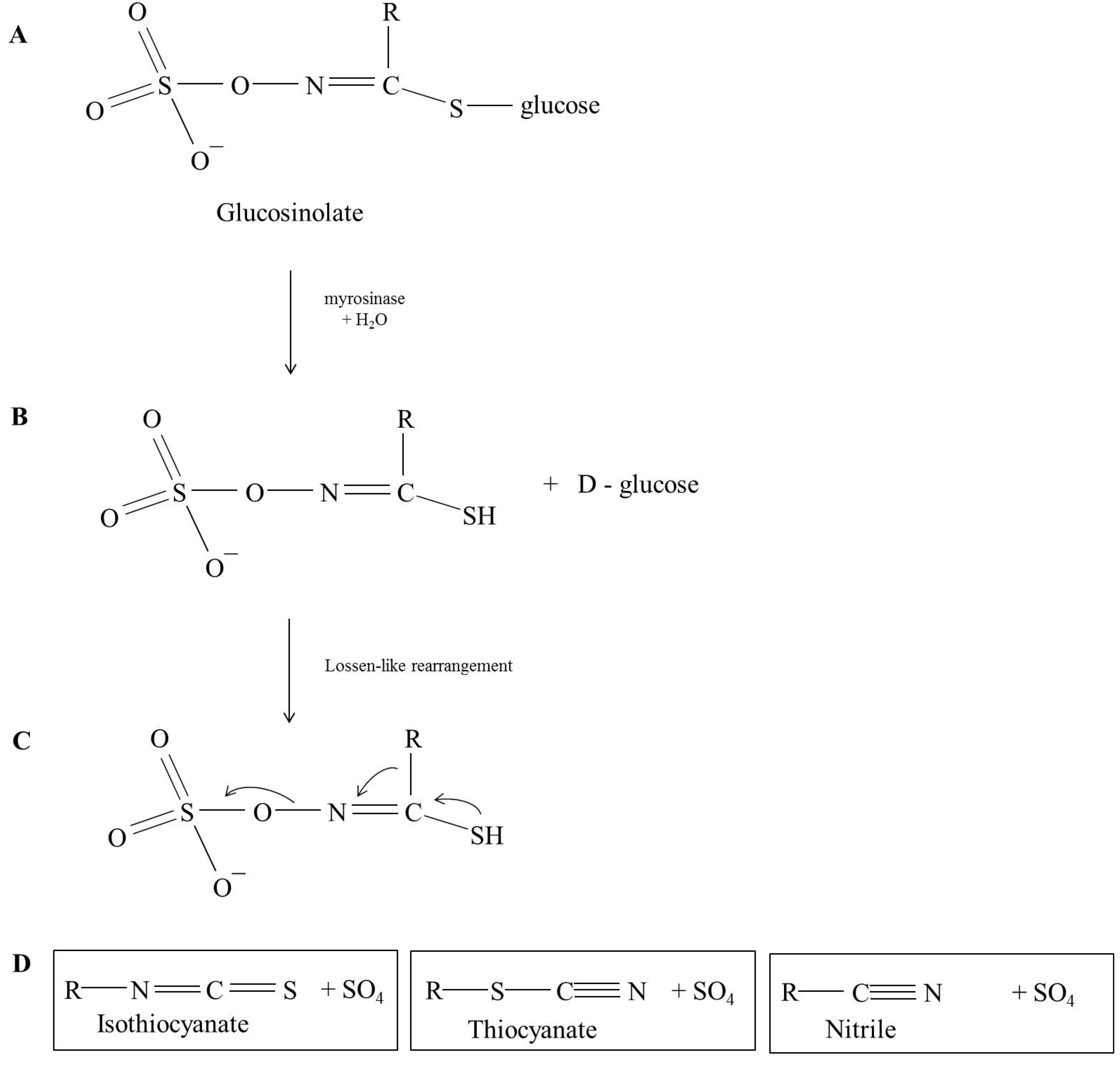
2. The Glucosinolates
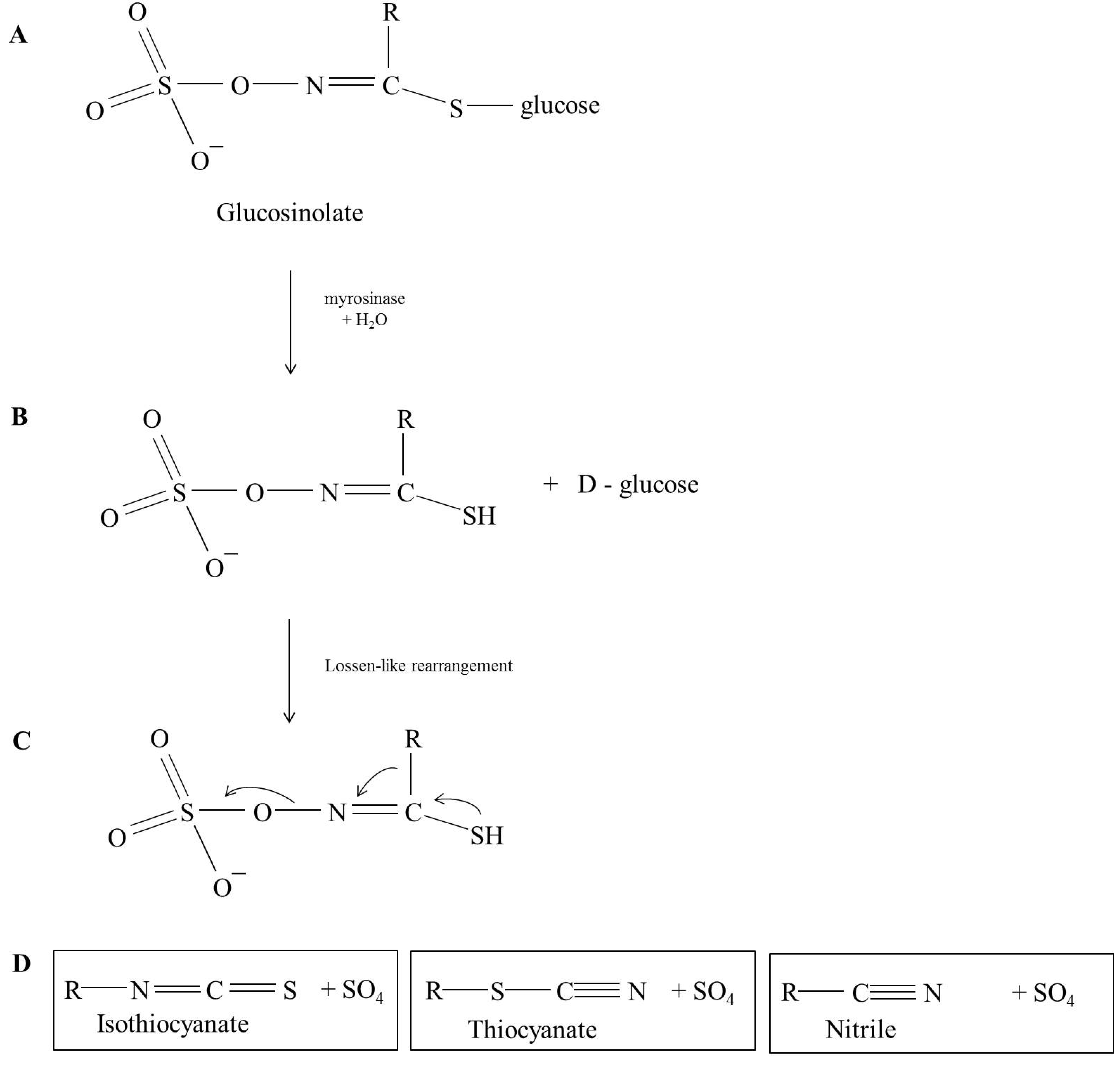
Glucosinolate Profile and Their Breakdown Products in Rocket

{kind=link}
{kind=link}
| Chemical name | Common name | Species | Reference |
|---|---|---|---|
| 2-Propenyl | Sinigrin | D. tenuifolia, E. sativa | [21] |
| 3-Butenyl | Gluconapin | E. sativa | [22] |
| Ethyl | Glucolepidin | D. tenuifolia, E. sativa | [17] |
| n-Butyl | - | D. tenuifolia, E. sativa | [17] |
| 1-Methylethyl | Glucoputranjivin | D. tenuifolia | [17] |
| 4-Hydroxybenzyl | Sinalbin | D. tenuifolia, E. sativa | [17] |
| Benzyl | Glucotropaeolin | D. tenuifolia | [17] |
| 4-Phenylbutyl | Gluconasturtiin | D. tenuifolia, E. sativa | [23] |
| 2-Phenylethyl | Gluconasturtiin | D. tenuifolia, E. sativa | [23] |
| 4-(Methylsulphinyl)butyl | Glucoraphanin | D. tenuifolia, E. sativa | [17,23] |
| 5-(Methylsulphinyl)-pentyl | Glucoalyssin | D. tenuifolia, E. sativa | [17,23] |
| 4-(Methylthio)butyl | Glucoerucin | D. tenuifolia, E. sativa | [17,18,24] |
| 2(S)-2-Hydroxy-3-butenyl | Progoitrin/epiprogoitrin | D. tenuifolia, E. sativa | [25] |
| 2-Hydroxyethyl | - | D. tenuifolia, E. sativa | [17] |
| 4-Hydroxyindol-3-ylmethyl | 4-Hydroxyglucobrassicin | D. tenuifolia, E. sativa | [18] |
| Indol-3-ylmethyl | Glucobrassicin | D. tenuifolia, E. sativa | [18,23,25] |
| 4-Methoxyindol-3-ylmethyl | 4-Methoxyglucobrassicin | D. tenuifolia, E. sativa | [23] |
| 1-Methoxyindol-3-ylmethyl | Neoglucobrassicin | D. tenuifolia, E. sativa | [20] |
| 4-Mercaptobutyl (dimer)b | DMB-GLS | D. tenuifolia, E. sativa | [24] |
| 4-(β-d-Glucopyranosyldisulfanyl) butyl) | 4-GDB-GLS | D. tenuifolia, E. sativa | [22] |
| 4-Hydroxybenzyl | Glucosinalbin | D. tenuifolia | [19] |
| 4-Mercaptobutyl | Glucosativin | D. tenuifolia, E. sativa | [24] |
| 4-Pentenil | Glucobrassicanupin | D. tenuifolia, E. sativa | [23] |
| 3-(Methylsulfanyl)propyl | Glucoiberverin | D. tenuifolia, E. sativa | [26] |
3. Nutrients and Antioxidant Compounds
4. Rocket Compounds with Protective Roles on Cancer and Human Diseases
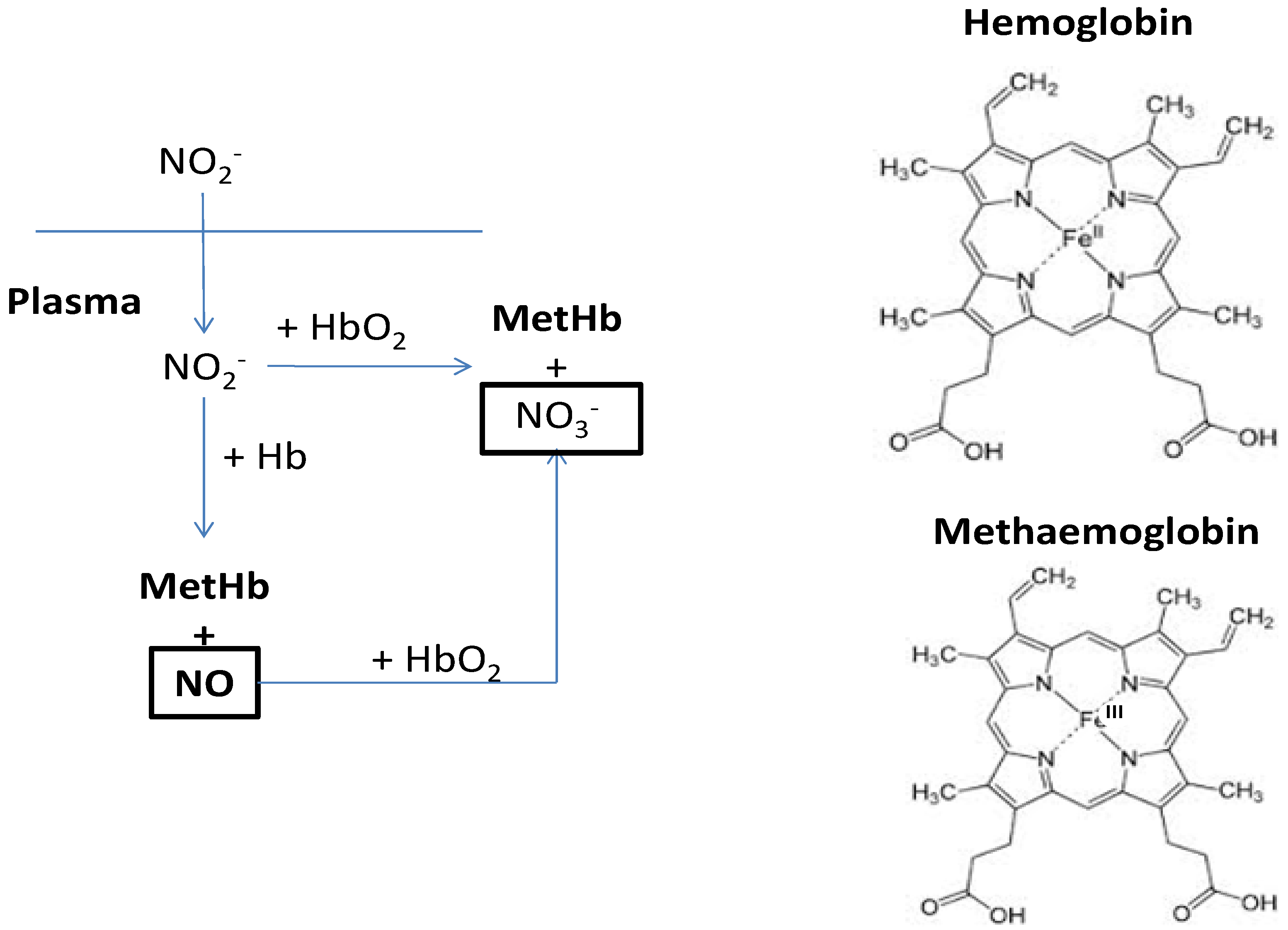
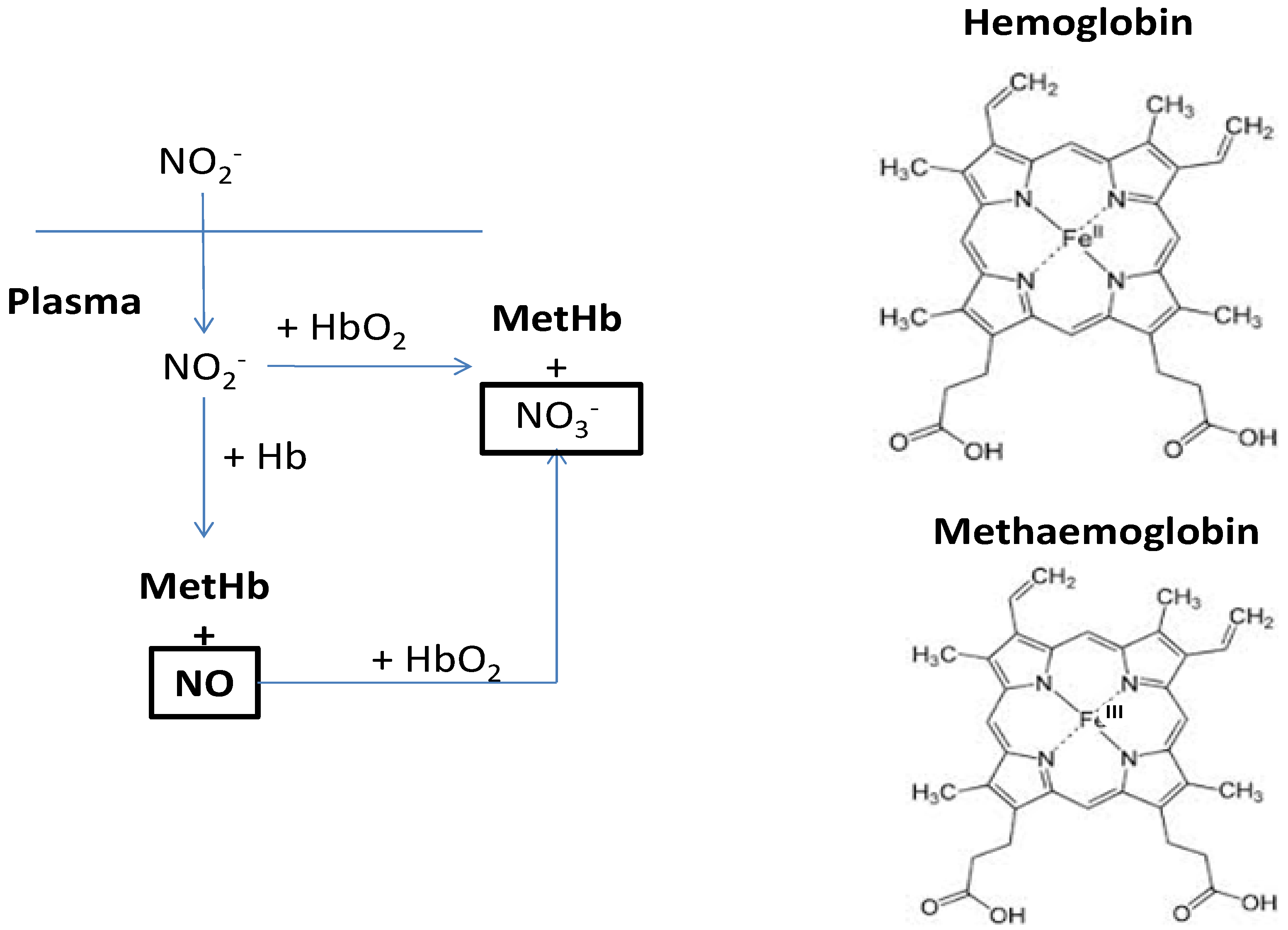
5. Nitrates in Rocket and the Effects on Human Health
| Leafy vegetable species | Harvesting period | Limits of NO3− (mg kg−1 FW) |
|---|---|---|
| Spinach (Spinacia oleracea L.) | 3500 | |
| 2500 | ||
| Frozen/canned spinach | 2000 | |
| Lettuce (Lactuca sativa L.) grown in protected environment or open field. | Harvesting from 1st October to 31st March:
| |
| 5000 | ||
| 4000 | ||
Harvesting from the 1st April to 30th September:
| ||
| 4000 | ||
| 3000 | ||
| Lettuce type “Iceberg” |
| 2500 |
| 2000 | ||
| Salad Rocket, wild Rocket (Eruca sativa, Diplotaxis tenuifolia) | Harvesting from 1st October to 31st March | 7000 |
| Salad Rocket, wild Rocket (Eruca sativa, Diplotaxis tenuifolia) | Harvesting from the 1st April to 30 September | 6000 |
| Species | Nitrate content (mg kg−1 FW) | Environment/technique | Reference |
|---|---|---|---|
| E. sativa | 1575–4139 | Organic | [105] |
| E. sativa | 2720–6036 | Soil | [105] |
| E. sativa | 4716–7083 | Hydroponics | [105] |
| D. tenuifolia | 5300–8100 | Peat, vermiculite, and perlite (3:2:1 v/v/v) | [71] |
| D. tenuifolia | 2500–3000 | Floating system | [73] |
| D. tenuifolia | 2524 | Soil (grown wild) | [106] |
| E. sativa | 1600–3300 | Floating system | [107,108] |
| E. sativa | 4400 | Hydroponically | [28] |
| E. sativa | 4000–4500 | Floating system | [72] |
| E. sativa | 7000 | Soil | [72] |
| E. sativa | 1282–2531 | Floating system | [103] |
| E. sativa | 217–341 | Soil | [108] |
| E. sativa | 432–467 | Floating system | [108] |
| E. sativa | 500–7200 | Floating system | [109] |
| E. sativa | 982 | Hydroponic Nutrient Film Technique | [110] |
| D. tenuifolia | 1859 | Hydroponic Nutrient Film Technique | [110] |
| E. sativa | 2574 | Soil (grown wild) | [106] |
| E. sativa | 4700 | Peat, vermiculite, and perlite (3:2:1 v/v/v) | [71] |
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Martinez-Sanchez, A.; Marin, A.; Llorach, R.; Ferreres, F.; Gil, M.I. Controlled atmosphere preserves quality and phytonutrients in wild rocket (Diplotaxis tenuifolia). Postharvest Biol. Technol. 2006, 40, 26–33. [Google Scholar] [CrossRef]
- Bianco, V.V. Rocket, An Ancient Underutilized Vegetable Crop and Its Potential. Rocket Genetic Resources Network. Report of the First Meeting, Lisbon, Portugal, 13–15 November 1994; Padulosi, S., Ed.; International Plant Genetic Resources Institute: Rome, Italy, 1995; pp. 35–59. [Google Scholar]
- Miyazawa, M.; Maehara, T.; Kurose, K. Composition of the essential oil from the leaves of Eruca sativa. Flavour Fragr. J. 2002, 17, 187–190. [Google Scholar]
- De Vos, A.C.; Broekman, R.; de Almeida Guerra, C.C.; van Rijsselberghe, M.; Rozema, J. Developing and testing new halophyte crops: A case study of salt tolerance of two species of the Brassicaceae, Diplotaxis tenuifolia and Cochlearia officinalis. Environ. Exp. Bot. 2013, 92, 154–164. [Google Scholar] [CrossRef]
- Hall, M.K.D.; Jobling, J.J.; Rogers, G.S. Some perspectives on rocket as a vegetable crop: A review. Veg. Crop. Res. Bull. 2012, 76, 21–41. [Google Scholar]
- De Feo, V.; Senatore, F. Medicinal plants and phytotherapy in the Amalfitan coast, Salerno Province, Campania, Southern Italy. J. Ethnopharmacol. 1993, 39, 39–51. [Google Scholar]
- Chopra, R.N.; Nayar, S.L.; Chopra, I.C. Glossary of Indian Medicinal Plants; Council of Scientific & Industrial Research: New Delhi, India, 1956; pp. 92–110. [Google Scholar]
- Alqasoumi, S. Carbon tetrachloride-induced hepatotoxicity: Protective effect of ‘Rocket’ Eruca sativa L. in rats. Am. J. Chin. Med. 2010, 38, 75–88. [Google Scholar] [CrossRef]
- Barillari, J.; Canistro, D.; Paolini, M.; Ferroni, F.; Pedulli, G.F.; Iori, R. Direct antioxidant activity of purified glucoerucin, the dietary secondary metabolite contained in rocket (Eruca sativa Mill.) seeds and sprouts. J. Agric. Food Chem. 2005, 53, 2475–2482. [Google Scholar] [CrossRef]
- Tommaso, R.I.; Cataldi, A.R.; Lelario, F.; Bufo, S.A. Naturally occurring glucosinolates in plant extracts of rocket salad (Eruca sativa L.) identified by liquid chromatography coupled with negative ion electrospray ionization and quadrupole ion-trap mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 2374–2388. [Google Scholar] [CrossRef]
- Sønderby, I.E.; Geu-Flores, F.; Halkier, B.A. Biosynthesis of glucosinolates—Gene discovery and beyond. Trends Plant Sci. 2010, 15, 283–290. [Google Scholar] [CrossRef]
- Wang, H.; Wu, J.; Sun, S.; Liu, B.; Cheng, F.; Sun, R.; Wang, X. Glucosinolate biosynthetic genes in Brassica rapa. Gene 2011, 487, 135–142. [Google Scholar] [CrossRef]
- Wallsgrove, R.M.; Doughty, K.J.; Bennett, R.N. Glucosinolates. In Plant Amino Acids: Biochemistry and Biotechnology; Marcel Dekker: BK Singh, NY, USA, 1999; pp. 523–562. [Google Scholar]
- Helmlinger, J.; Rausch, T.; Hilgenberg, W. Localization of newly synthesized indole-3-methylglucosinolate (=glucobrassicin) in vacuoles from horseradish (Armoracia rusticana). Physiol. Plant 1983, 58, 302–310. [Google Scholar] [CrossRef]
- Holst, B.; Williamson, G. A critical review of the bioavailability of glucosinolates and related compounds. Nat. Prod. Rep. 2004, 21, 425–447. [Google Scholar]
- Chin, H.W.; Lindsay, R.C. Modulation of volatile sulfur compounds in cruciferous vegetables. In Sulfur Compounds in Foods; Mussinan, C.J., Keelan, M.E., Eds.; American Chemical Society: Washington, DC, USA, 1994; pp. 90–104. [Google Scholar]
- D’antuono, L.F.; Elementi, S.; Neri, R. Glucosinolates in Diplotaxis and Eruca leaves: Diversity, taxonomic relations and applied aspects. Phytochemistry 2008, 69, 187–199. [Google Scholar] [CrossRef]
- Cataldi, T.R.; Rubino, A. Naturally occurring glucosinolates in plant extracts of rocket salad (Eruca sativa L.) identified by liquid chromatography coupled with negative ion electrospray ionization and quadrupole ion-trap mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 2374–2388. [Google Scholar] [CrossRef]
- Bennett, R.N.; Rosa, E.A.S.; Mellon, F.A.; Kroon, P.A. Ontogenic profiling of glucosinolates, flavonoids, and other secondary metabolites in Eruca sativa (salad rocket), Diplotaxis erucoides (wall rocket), Diplotaxis tenuifolia (wild rocket), and Bunias orientalis (turkish rocket). J. Agric. Food Chem. 2006, 54, 4005–4015. [Google Scholar] [CrossRef]
- Pasini, F.; Verardo, V.; Cerretani, L.; Caboni, M.F.; D’Antuono, L.F. Rocket salad (Diplotaxis and Eruca spp.) sensory analysis and relation with glucosinolate and phenolic content. J. Sci. Food Agric. 2011, 91, 2858–2864. [Google Scholar]
- Bennett, R.N.; Carvalho, R.; Mellon, F.A.; Eagles, J.; Rosa, E.A. Identification and quantification of glucosinolates in sprouts derived from seeds of wild Eruca sativa L.(salad rocket) and Diplotaxis tenuifolia L. (wild rocket) from diverse geographical locations. J. Agric. Food Chem. 2007, 55, 67–74. [Google Scholar]
- Adhikari, J.; Adhikari, S.; Achaya, K.T. Glucosinates in the seeds of Indian brassicas and Eruca sativa. Oil Technol. Assoc. India 1989, 21, 13–14. [Google Scholar]
- Chun, J.H.; Arasu, M.V.; Lim, Y.P.; Kim, S.J. Variation of major glucosinolates in different varieties and lines of rocket salad. Hortic. Environ. Biotechnol. 2013, 54, 206–213. [Google Scholar]
- Bennett, R.N.; Mellon, F.A.; Botting, N.P.; Eagles, J.; Rosa, E.A.S.; Williamson, G. Identification of the major glucosinolate (4-mercaptobutylglucosinolate) in leaves of Eruca sativa L. Phytochemistry 2002, 61, 25–30. [Google Scholar]
- Kiddle, G.; Bennett, R.N.; Botting, N.P.; Davidson, N.E.; Robertson, A.A.B.; Wallsgrove, R.M. High-performance liquid chromatographic separation of natural and synthetic desulphoglucosinolates and their chemical validation by UV, NMR and chemical ionisation-MS methods. Phytochem. Anal. 2001, 12, 226–242. [Google Scholar] [CrossRef]
- Jin, J.; Koroleva, O.A.; Gibson, T.; Swanston, J.; Magan, J.; Zhang, Y.; Rowland, I.R.; Wagstaff, C. Analysis of phytochemical composition and chemoprotective capacity of rocket (Eruca sativa and Diplotaxis tenuifolia) leafy salad following cultivation in different environments. J. Agric. Food Chem. 2009, 57, 5227–5234. [Google Scholar] [CrossRef]
- Verkerk, R.; Schreiner, M.C.; Krumbein, A.; Ciska, E.; Holst, B.; Rowland, I.; de Schrijver, R.; Hansen, M.; Gerhäuser, C.; Mithen, R.; et al. Glucosinolates from Brassica vegetables; The influence of the food supply chain on intake, bioavailability and human health. Mol. Nutr. Food Res. 2009, 53, 219. [Google Scholar] [CrossRef]
- Kim, S.; Ishii, G. Effect of storage temperature and duration on glucosinolate, total vitamin C and nitrate contents in rocket salad (Eruca sativa Mill.). J. Sci. Food Agric. 2007, 87, 966–973. [Google Scholar] [CrossRef]
- Force, L.E.; O’Hare, T.J.; Wong, L.S.; Irving, D.E. Impact of cold storage on glucosinolate levels in seed-sprouts of broccoli, rocket, white radish and kohl-rabi. Postharvest Biol. Technol. 2007, 44, 175–178. [Google Scholar] [CrossRef]
- Selma, M.V.; Martinez-Sanchez, A.; Allende, A.; Ros, M.; Hernandez, M.T.; Gil, M.I. Impact of organic soil amendments on phytochemicals and microbial quality of rocket leaves (Eruca sativa). J. Agric. Food Chem. 2010, 58, 8331–8337. [Google Scholar] [CrossRef]
- Kim, S.J.; Ishii, J. Glucosinolate profiles in the seeds, leaves and roots of rocket salad (Eruca sativa Mill.) and anti-oxidative activities of intact plant powder and purified 4-methoxyglucobrassicin. Soil Sci. Plant Nutr. 2006, 52, 394–400. [Google Scholar] [CrossRef]
- Omirou, M.; Papastefanou, C.; Katsarou, D.; Papastylianou, I.; Passam, H.C.; Ehaliotis, C.; Papadopoulou, K.K. Relationships between nitrogen, dry matter accumulation and glucosinolates in Eruca sativa Mills. The applicability of the critical NO3-N levels approach. Plant Soil 2012, 354, 347–358. [Google Scholar] [CrossRef]
- Labague, L.; Gardrat, G.; Coustille, J.L.; Viaud, M.C.; Rollin, P. Identification of enzymatic degradation products from synthesised glucobrassicin by gas chromatography-mass spectrometry. J. Chromatogr. 1991, 586, 166–170. [Google Scholar] [CrossRef]
- Jirovetz, L.; Smith, D.; Buchbauer, G. Aroma compound analysis of Eruca sativa (Brassicaceae) SPME headspace leaf samples using GC, GC-MS, and Olfactometry. J. Agric. Food Chem. 2002, 50, 4643–4646. [Google Scholar] [CrossRef]
- Villatoro-Pulido, M.; Priego-Capote, F.; Álvarez-Sánchez, B.; Saha, S.; Philo, M.; Obregón-Cano, S.; de Haro-Bailón, A.; Font, R.; del Río-Celestino, M. An approach to the phytochemical profiling of rocket [Eruca sativa (Mill.) Thell]. J. Sci. Food Agric. 2013, 93, 3809–3819. [Google Scholar] [CrossRef]
- Hanley, B.A.; Parsley, K.R.; Lewis, J.A.; Fenwick, R.G. Chemistry of indole glucosinolates: Intermediacy of indole-3-ylmethyl isothiocyanates in the enzyme hydrolysis of indole glucosinolates. J. Chem. Soc. 1990, 1, 2273–2276. [Google Scholar]
- Hasapis, X.; MaCleod, A.J. Benzylglucosinolate degradation in heat-treated Lepidium sativum seeds and detection of a thiocyanate-forming factor. Phytochemistry 1982, 21, 1009–1013. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Cocetta, G.; Ferrante, A. The antioxidants changes in ornamental flowers during development and senescence. Antioxidants 2013, 2, 132–155. [Google Scholar] [CrossRef]
- Bozokalfa, M.K.; Esiyok, D.; Yagmur, B. Use of multivariate analysis in mineral accumulation of rocket (Eruca sativa) accessions. Genetika 2011, 43, 437–448. [Google Scholar] [CrossRef]
- Bozokalfa, M.K.; Yagmur, B.; Ilbi, H.; Esiyok, D.; Kavak, S. Genetic variability for mineral concentration of Eruca sativa L. and Diplotaxis tenuifolia L. accessions. Crop Breed. Appl. Biotechnol. 2009, 9, 372–381. [Google Scholar] [CrossRef]
- Kawashima, L.M.; Valente-Soares, L.M. Mineral profile of raw and cooked leafy vegetables consumed in southern Brazil. J. Food Compos. Anal. 2003, 16, 605–611. [Google Scholar] [CrossRef]
- Bukhashi, E.; Maliki, S.; Ahmed, S.S. Estimation of nutritional value and trace elements content of Carthamus oxycantha, Eruca sativa and Plantago ovanta. Pak. J. Bot. 2007, 30, 1181–1187. [Google Scholar]
- Branca, F. Work Conducted by the Institute of Horticulture and Floriculture of the University of Catania, Sicily. Rocket Genetic Resources Network. In Report of the First Meeting, Lisbon, Portugal, 13–15 November 1994; Padulosi, S., Ed.; International Plant Genetic Resources Institute: Rome, Italy, 1995; p. 7. [Google Scholar]
- Esiyok, D.; Ongun, A.R.; Bozolkalfa, K.; Tepecik, M.; Okur, B.; Kaygisiz, T. Organic Rocket Growing. In Proceedings of the VI Vegetable Agriculture Symposium, Kahramanmaras, Turkey, 19–22 September 2006; pp. 85–89.
- Acikgoz, F.E. The effects of different sowing time practices on vitamin C and mineral material content for rocket (Eruca vesicaria subsp. sativa (Mill)). Sci. Res. Essays 2011, 6, 3127–3131. [Google Scholar]
- Francke, A. Effect of cultivation time and soil kind on yielding of garden rocket (Eruca sativa LDC.). Folia Univ. Agric. Stetin. 2004, 95, 81–85. [Google Scholar]
- Fraszczak, B.; Ziombra, M.; Knaflewski, M. The content of vitamin C and essential oils in herbage of some spice plants depending on light conditions and temperature. Roczniki Akademii Rolniczej w Poznaniu 2006, 379, 15–21. [Google Scholar]
- Martinez-Sanchez, A.; Llorach, R.; Gil, M.I.; Ferreres, F. Identification of new flavonoid glycosides and flavonoid profiles to characterize rocket leafy salads (Eruca vesicaria and Diplotaxis tenuifolia). J. Agric. Food Chem. 2007, 55, 1356–1363. [Google Scholar]
- Weckerle, B.; Michel, K.; Balázs, B.; Schreier, P.; Tóth, G. Quercetin 3,3′,4′-tri-O-β-d-glucopyranosides from leaves of Eruca sativa (Mill.). Phytochemistry 2001, 57, 547–551. [Google Scholar]
- Znidarcic, D.; Ban, D.; Helena Šircelj, H. Carotenoid and chlorophyll composition of commonly consumed leafy vegetables in Mediterranean countries. Food Chem. 2011, 129, 1164–1168. [Google Scholar] [CrossRef]
- Bryan, R.; Moser, J.K.; Moser, W.; Shah, S.N.; Vaughn, S.F. Composition and physical properties of arugula, shepherd’s purse, and upland cress oils. Eur. J. Lipid Sci. Technol. 2010, 112, 734–740. [Google Scholar] [CrossRef]
- Brock, A.; Herzfeld, T.; Paschke, R.; Koch, M.; Dräger, B. Brassicaceae contain nortropane alkaloids. Phytochemistry 2006, 67, 2050–2057. [Google Scholar]
- Gulfraz, M.; Sadiq, A.; Tariq, H.; Imran, M.; Qureshi, R.; Zeenat, A. Phytochemical analysis and antibacterial activity of Eruca sativa seed. Pak. J. Bot. 2011, 43, 1351–1359. [Google Scholar]
- Flanders, A.; Abdulkarim, S.M. The composition of seed and seed oils of Taramira (Eruca sativa). J. Am. Oil Chem. Soc. 1985, 62, 1134–1135. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Ali, J.; Ali, M.; Zia, M.; Bokhari, S.A.; Khan, M.A. Free radical scavenging activity in in vitro-derived tissues of Eruca sativa. Toxicol. Ind. Health 2013. in publish. [Google Scholar]
- Johnson, I.T. Glucosinolates in the human diet: Bioavailability and implications for health. Phytochem. Rev. 2002, 1, 183–188. [Google Scholar] [CrossRef]
- Traka, M.; Mithen, R. Glucosinolates, isothiocyanates and human health. Phytochem. Rev. 2009, 8, 269–282. [Google Scholar] [CrossRef]
- Fahey, J.W.; Talalay, P. Antioxidant functions of sulforaphane: A potent inducer of Phase II detoxication enzymes. Food Chem. Toxicol. 1999, 37, 973–979. [Google Scholar] [CrossRef]
- Rodriguez, A.S.; Gurovic, M.S.V.; Mulet, M.C.; Murray, A.P. Diplotaxis tenuifolia (L.) DC., a source of a potentially antifungal essential oil containing nitrile. Biochem. Syst. Ecol. 2006, 34, 353–355. [Google Scholar]
- Shaaya, E.; Kostyukovsky, M.; Demchenko, N. Alternative fumigants for the control of stored-product insects. In Advances in Stored-Product Protection; Proceedings of the Eighth International Working Conference on Stored-Product Protection, York, UK, 22–26 July 2002; Credland, P.F., Armitage, D.M., Bell, C.H., Cogan, P.M., Highley, E., Eds.; CAB International: Wallingford, UK, 2002; pp. 556–560. [Google Scholar]
- Alqasoumi, S.; Al-Sohaibani, M.; Al-Howiriny, T.; Al-Yahya, M.; Rafatullah, S. Rocket “Eruca sativa”: A salad herb with potential gastric anti-ulcer activity. World J. Gastroenterol. 2009, 15, 1958–1965. [Google Scholar]
- Lamy, E.; Schröder, J.; Paulus, S.; Brenk, P.; Stahl, T.; Mersch-Sundermann, V. Antigenotoxic properties of Eruca sativa (rocket plant), erucin and erysolin in human hepatoma (HepG2) cells towards benzo(a)pyrene and their mode of action. Food Chem. Toxicol. 2008, 46, 2415–2421. [Google Scholar] [CrossRef]
- El-Missiry, M.A.; El Gindy, A.M. Amelioration of alloxan induced diabetes mellitus and oxidative stress in rats by oil of Eruca sativa seeds. Ann. Nutr. Metab. 2000, 44, 97–100. [Google Scholar] [CrossRef]
- Alama, M.S.; Kaurb, G.; Jabbara, Z.; Javeda, K.; Athar, M. Eruca sativa seeds possess antioxidant activity and exert a protective effect on mercuric chloride induced renal toxicity. Food Chem. Toxicol. 2007, 45, 910–920. [Google Scholar] [CrossRef]
- Khoobchandania, M.; Ganeshb, N.; Gabbaninic, S.; Valgimiglid, L.; Srivastavaa, M.M. Phytochemical potential of Eruca sativa for inhibition of melanoma tumor growth. Fitoterapia 2011, 82, 647–653. [Google Scholar] [CrossRef]
- Emtenan, M.; Hanafi, E.M.; Hegazy, R.M.; Riad, H.A. Bio-protective effect of Eruca sativa seed oil against the hazardous effect of aflatoxin b1 in male rabbits. Int. J. Acad. Res. 2010, 2, 67–74. [Google Scholar]
- Mona, A.R.S.; Nehal, A.M. Histological and quantitative study of the effect of Eruca sativa seed oil on the testis of albino rat. Egypt. J. Hosp. Med. 2001, 2, 148–162. [Google Scholar]
- Melchini, A.; Costa, C.; Traka, M.; Miceli, N.; Mithen, R.; de Pasquale, R.; Trovato, A. Erucin, a new promising cancer chemopreventive agent from rocket salads, shows anti-proliferative activity on human lung carcinoma A549 cells. Food Chem. Toxicol. 2009, 47, 1430–1436. [Google Scholar] [CrossRef]
- Yehuda, H.; Khatib, S.; Sussan, I.; Musa, R.; Vaya, J.; Tamir, S. Potential skin antiinflammatory effects of 4-methylthiobutylisothiocyanate (MTBI) isolated from rocket (Eruca sativa) seeds. Biofactors 2009, 35, 295–305. [Google Scholar] [CrossRef]
- Yehuda, H.; Soroka, Y.; Zlotkin-Frušić, M.; Gilhar, A.; Milner, Y.; Tamir, S. Isothiocyanates inhibit psoriasis-related proinflammatory factors in human skin. Inflamm. Res. 2012, 61, 735–742. [Google Scholar] [CrossRef]
- Jakubikova, J.; Sedlak, J.; Mithen, R.; Bao, Y. Role of PI3K/Akt and MEK/ERK signaling pathways in sulforaphane-and erucin-induced phase II enzymes and MRP2 transcription, G2/M arrest and cell death in Caco-2 cells. Biochem. Pharmacol. 2005, 69, 1543–1552. [Google Scholar] [CrossRef]
- Durazzo, A.; Azzini, E.; Lazzè, M.C.; Raguzzini, A.; Pizzala, R.; Maiani, G. Italian Wild Rocket [Diplotaxis Tenuifolia (L.) DC.]. Influence of agricultural practices on antioxidant molecules and on cytotoxicity and antiproliferative effects. Agriculture 2013, 3, 285–298. [Google Scholar] [CrossRef]
- Santamaria, P.; Elia, A.; Serio, F. Effect of solution nitrogen concentration on yield, leaf element content, and water and nitrogen use efficiency of three hydroponically-grown rocket salad genotypes. J. Plant Nutr. 2002, 25, 245–258. [Google Scholar] [CrossRef]
- Ferrante, A.; Incrocci, L.; Maggini, R.; Serra, G.; Tognoni, F. Preharvest and postharvest strategies for reducing nitrate content in rocket (Eruca sativa L.). Acta Hortic. 2003, 628, 153–159. [Google Scholar]
- Alberici, A.; Quattrini, E.; Penati, M.; Martinetti, L.; Marino Gallina, P.; Ferrante, A. Effect of the reduction of nutrient solution concentration on leafy vegetables quality grown in floating system. Acta Hortic. 2008, 801, 1167–1176. [Google Scholar]
- Bruning-Fann, C.S.; Kaneene, J.B. The effects of nitrate, nitrite and N-nitroso compounds on human health: A review. Vet. Hum. Toxicol. 1993, 35, 521–538. [Google Scholar]
- Magee, P.N.; Barnes, J.M. The production of malignant primary hepatic tumours in the rat by feeding dimethylnitrosamine. Br. J. Cancer 1956, 10, 114–122. [Google Scholar] [CrossRef]
- Gangolli, S.D.; van den Brandt, P.A.; Feron, V.J.; Janzowsky, C.; Koeman, J.H.; Speijers, G.J.A.; Spiegelhalder, B.; Walker, R.; Winshnok, J.S. Nitrate, nitrite and N-nitroso compounds. Eur. J. Pharmacol. Environ. Toxicol. 1994, 4, 1–38. [Google Scholar]
- Camp, N.E. Methemoglobinemia. J. Emerg. Nurs. 2007, 33, 172–174. [Google Scholar] [CrossRef]
- Milkowski, A.; Garg, H.K.; Coughlin, J.R.; Bryan, N.S. Nutritional epidemiology in the context of nitric oxide biology: A risk-benefit evaluation for dietary nitrite and nitrate. Nitric Oxide 2010, 22, 110–119. [Google Scholar] [CrossRef]
- Weitzberg, E.; Lundberg, J.O. Novel aspects of dietary nitrate and human health. Ann. Rev. Nutr. 2013, 33, 129–159. [Google Scholar] [CrossRef]
- McKnight, G.M.; Smith, L.M.; Drummond, R.S.; Duncan, C.W.; Golden, M.; Benjamin, N. Chemical synthesis of nitric oxide in the stomach from dietary nitrate in humans. Gut 1997, 40, 211–214. [Google Scholar]
- McKnight, G.M.; Duncana, C.W.; Leifert, C.; Golden, M.H. Dietary nitrate in man: Friend or foe? Br. J. Nutr. 1999, 81, 349–358. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Feelisch, M.; Björne, H.; Jansson, E.A.; Weitzberg, E. Cardioprotective effects of vegetables: Is nitrate the answer? Nitric Oxide 2006, 15, 359–362. [Google Scholar] [CrossRef]
- Gilchrist, M.; Shore, A.C.; Benjamin, N. Inorganic nitrate and nitrite and control of blood pressure. Cardiovasc. Res. 2011, 89, 492–498. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on Contaminants in the Food chain on a request for the European Commission to perform a scientific risk assessment on nitrate in vegetables. EFSA J. 2008, 689, 1–79. [Google Scholar]
- Speijers, G.; van den Brandt, P.A. Nitrate. In Food Additives Series; Joint FAO/WHO Expert Committee on Food Additives: Geneva, Switzerland, 2003. [Google Scholar]
- European Commission. Commission Regulation (EC) No 1882/2006 of 19 December 2006 laying down methods of sampling and analysis for the official control of the levels of nitrates in certain foodstuffs. Off. J. Eur. Union 2006, L364, 25–31. [Google Scholar]
- European Commission. Commission Regulation (EU) No 1258/2011 of 2 December 2011 amending Regulation (EC) No. 1881/2006 as regards maximum levels for nitrates in foodstuffs. Off. J. Eur. Union 2011, L320, 15–17. [Google Scholar]
- Weightman, R.M.; Huckle, A.J.; Roques, S.E.; Ginsburg, D.; Dyer, C.J. Factors influencing tissue nitrate concentration in field-grown wild rocket (Diplotaxis tenuifolia) in southern England. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2012, 29, 1425–1435. [Google Scholar] [CrossRef]
- Santamaria, P.; Elia, A.; Parente, A.; Serio, F. Fertilization strategies for lowering nitrate content in leafy vegetables: Chycory and rocket salad cases. J. Plant Nutr. 1998, 21, 1791–1803. [Google Scholar] [CrossRef]
- Elia, A.; Santamaria, P.; Serio, F. Ammonium and nitrate influence on artichoke growth rate and uptake of inorganic ions. J. Plant Nutr. 1996, 19, 1029–1044. [Google Scholar] [CrossRef]
- Serio, F.; Elia, A.; Signore, A.; Santamaria, P. Influence of nitrogen form on yield and nitrate content of subirrigated early potato. J. Sci. Food Agric. 2004, 84, 1428–1432. [Google Scholar]
- Bonasia, A.; Conversa, G.; Gonnella, M.; Serio, F.; Santamaria, P. Effects of ammonium and nitrate nutrition on yield and quality in endive. J. Hortic. Sci. Biotechnol. 2008, 83, 64–70. [Google Scholar]
- Podetta, N.; Antonacci, S.; Ferrante, A. Diurnal pattern of nitrate reductase in leaves of rocket (Diplotaxis tenuifolia L.) plants. Minerva Biotecnol. 2011, 23, 40–41. [Google Scholar]
- Antonacci, S.; Maggiore, T.; Ferrante, A. Nitrate metabolism in plants under hypoxic and anoxic conditions. Plant Stress 2007, 1, 136–141. [Google Scholar]
- Gaudreau, L.; Charbonneau, J.; Vezina, L.P.; Gosselin, A. Effects of photoperiod and photosynthetic photon flux on nitrate content and nitrate reductase activity in greenhouse-grown lettuce. J. Plant Nutr. 1995, 18, 437–453. [Google Scholar] [CrossRef]
- Hassan, C. Effect of supplementary lighting on growth and primary nitrogen metabolism of greenhouse lamb’s lettuce and spinach. Can. J. Plant Sci. 1999, 79, 421–426. [Google Scholar] [CrossRef]
- De Martin, S.; Restani, P. Determination of nitrates by a novel ion chromatographic method: Occurrence in leafy vegetables (organic and conventional) and exposure assessment for Italian consumers. Food Addit. Contam. 2003, 20, 787–792. [Google Scholar] [CrossRef]
- Council Regulation (EEC) No 2092/91 of 24 June 1991 on organic production of agricultural products and indications referring thereto on agricultural products and foodstuffs. Off J Eur Union. 1991, L198, 1–89.
- Ferrante, A.; Trivellini, A.; Malorgio, F.; Carmassi, G.; Vernieri, P.; Serra, G. Effect of seawater aerosol on leaves of six plant species potentially useful for ornamental purposes in coastal areas. Sci. Hortic. 2011, 128, 332–341. [Google Scholar] [CrossRef]
- Neocleous, D.; Koukounaras, A.; Siomos, A.S.; Vasilakakis , M. Assessing the salinity effects on mineral composition and nutritional quality of green and red “baby” lettuce. J. Food Qual. 2014, 37, 11–18. [Google Scholar]
- Bianco, V.V.; Santamaria, P.; Elia, A. Nutritional value and nitrate content in edible wild species used in southern Italy. Acta Hortic. 1998, 467, 71–87. [Google Scholar]
- Barbieri, G.; Bottino, A.; di Stasio, E.; Vallone, S.; Maggio, A. Proline and light as quality enhancers of rocket (Eruca sativa Miller) grown under saline conditions. Sci. Hortic. 2011, 128, 393–400. [Google Scholar]
- Guadagnin, S.G.; Rath, S.; Reyes, F.G.R. Evaluation of the nitrate content in leaf vegetables produced through different agricultural systems. Food Addit. Contam. 2005, 22, 1203–1208. [Google Scholar] [CrossRef]
- Vernieri, P.; Borghesi, E.; Tognoni, F.; Serra, G.; Ferrante, A.; Piaggesi, A. Use of biostimulants for reducing nutrient solution concentration in floating system. Acta Hortic. 2006, 718, 477–484. [Google Scholar]
- Vernieri, P.; Borghesi, E.; Ferrante, A.; Magnani, G. Application of biostimulants in floating system for improving rocket quality. J. Food Agric. Environ. 2005, 33, 86–88. [Google Scholar]
- Fontana, E.; Nicola, S. Traditional and soilless culture systems to produce corn salad (Valerianella olitoria L.) and rocket (Eruca sativa Mill.) with low nitrate content. J. Food Agric. Environ. 2009, 7, 405–410. [Google Scholar]
- Cavarianni, R.L.; Cecilio, A.B.; Cazetta, J.O.; May, A.; Corradi, M.M. Nutrient contents and production of rocket as affected by nitrogen concentrations in the nutritive solution. Sci. Agric. 2008, 65, 652–658. [Google Scholar]
- Kim, S.J.; Kawaharada, C.; Ishii, G. Effect of ammonium: Nitrate nutrient ratio on nitrate and glucosinolate contents of hydroponically-grown rocket salad (Eruca sativa Mill.). Soil Sci. Plant 2006, 52, 387–393. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cavaiuolo, M.; Ferrante, A. Nitrates and Glucosinolates as Strong Determinants of the Nutritional Quality in Rocket Leafy Salads. Nutrients 2014, 6, 1519-1538. https://doi.org/10.3390/nu6041519
Cavaiuolo M, Ferrante A. Nitrates and Glucosinolates as Strong Determinants of the Nutritional Quality in Rocket Leafy Salads. Nutrients. 2014; 6(4):1519-1538. https://doi.org/10.3390/nu6041519
Chicago/Turabian StyleCavaiuolo, Marina, and Antonio Ferrante. 2014. "Nitrates and Glucosinolates as Strong Determinants of the Nutritional Quality in Rocket Leafy Salads" Nutrients 6, no. 4: 1519-1538. https://doi.org/10.3390/nu6041519
APA StyleCavaiuolo, M., & Ferrante, A. (2014). Nitrates and Glucosinolates as Strong Determinants of the Nutritional Quality in Rocket Leafy Salads. Nutrients, 6(4), 1519-1538. https://doi.org/10.3390/nu6041519